Descending from the trees onto the mud to feed: observation of ophiophagy by Trimeresurus purpureomaculatus (Gray, 1832) on - Biotaxa
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Herpetology Notes, volume 14: 1281-1285 (2021) (published online on 8 October 2021)
Descending from the trees onto the mud to feed: observation of
ophiophagy by Trimeresurus purpureomaculatus (Gray, 1832) on
Fordonia leucobalia (Schlegel, 1837)
Alex Figueroa1,* and Ryan J.R. McCleary2
Ophiophagy, the feeding mode whereby animals prey bend the vertebral column of the prey snake into
on snakes, is considered exceptional in snakes (Greene, waves to accommodate it inside the gastrointestinal
1997) aside from a few taxa famed for this dietary tract (Jackson et al., 2004).
specialisation, most notably members of the genera Vipers, in particular, are not known to engage in
Clelia, Lampropeltis, and Micrurus, as well as the King ophiophagy as readily as other snakes, as they have
Cobra, Ophiophagus hannah (Cantor, 1836). Colston been documented to feed mainly on amphibians, lizards,
et al. (2010) estimated that approximately 20.4% birds, and mammals (Cundall & Greene, 2000). The
(~700 species) of all alethinophidian snakes (Uetz et diet of vipers is mostly attributed to them being more
al., 2021) exhibit ophiophagous behaviour. Numerous robust and having a large gape capacity relative to other
observations of ophiophagy have been recorded in snakes and utilising a sit-and-wait foraging mode. Thus,
other species, but these are generally limited to single vipers are regarded as feeding less frequently than other
events (e.g., McKelvy et al., 2013; Coelho-Lima et snakes. Trimeresurus purpureomaculatus (Gray, 1832) is
al., 2020), undoubtedly due to the secretive nature of an arboreal pitviper that inhabits mangroves and coastal
snakes that makes witnessing snakes feeding in the wild environments of southern Myanmar, Thailand, western
challenging. Indeed, much of what is known regarding Peninsular Malaysia, Singapore, and Sumatra (David
snake diets comes from the examination of museum and Vogel, 1996). Aside from being arboreal and feeding
specimens (Glaudas et al., 2017). on frogs, lizards, birds, and small mammals (David
Theoretically, as gape-limited predators, feeding and Vogel 1996), Pauwels et al. (2000) reported that a
on elongated prey should be functionally more captured individual in Thailand contained the digestive
manageable for snakes than feeding on bulkier prey remains of a congener. Otherwise, little is published
with limbs. One hypothesis why ophiophagy is not regarding the ecology of T. purpureomaculatus. Herein,
more prevalent in snakes is related to handling time, we present the first field observation of ophiophagy in
where ingesting elongated prey takes substantially T. purpureomaculatus (Fig. 1), in which we observed a
longer than non-elongated prey (Jackson et al., 2004; juvenile trail, capture, and feed on a Fordonia leucobalia
Banci et al., 2017). Handling time is a function of (Schlegel, 1837), a mangrove homalopsid (mudsnake)
prey size, and snakes as prey tend to approximate that spends a considerable amount of time in mud lobster
or exceed the size of the predatory snake, especially mounds (Karns et al., 2002). We also discuss a potentially
the consumer’s gut size (Jackson et al., 2004). One novel form of ophiophagous consumption whereby the
solution to overcoming this challenge is to forcibly prey is ingested not head or tail first (Jackson et al., 2004;
Braz and Marques, 2016), but from the midbody.
On the night of 25 November 2014, while surveying
Pasir Ris Park Mangroves (PRPM) in Singapore
1
Department of Biological Sciences, University of New Orleans,
New Orleans, Louisiana 70122, USA.
(1.37789°N, 103.9525°E), we spotted a snake writhing
2
Protein Science Laboratory, Department of Biological Sciences, in the mud at around 23:35 h. The snake was a ~30 cm
Faculty of Science, National University of Singapore, 117543, snout–vent length (SVL) F. leucobalia that appeared to
Singapore; and Department of Integrative Biology, University be in the stages of dying (Fig. 1A). After watching the
of South Florida, Sarasota, Florida 34243, USA. snake for a few seconds, we noticed another snake trailing
*
Corresponding author. E-mail: afigueroa21@gmail.com behind it. The second snake we quickly identified as T.
© 2021 by Herpetology Notes. Open Access by CC BY-NC-ND 4.0. purpureomaculatus (Fig. 1A). It is likely that just prior to
1282 Alex Figueroa & Ryan J.R. McCleary our arrival, the ~45 cm SVL viper had envenomated the incrementally (Fig. 2A). For further support, the viper mudsnake and was pursuing it. When we first approached, used its tail to anchor itself against the tree root, switching the viper seemed to become agitated, retreated a few from having its tail on its left side to positioning it on its meters, but quickly resumed its pursuit. By 23:40 h, the right side for better leverage (Fig. 2B). By 00:25 h, the viper had reached the mudsnake and began probing along viper had reached a point in the bend of the mudsnake’s its body, seemingly trying to find the head (Fig. 1B). This body where the head and tail began to meet, and the occurred at the base of a mound, and as the viper seized head entered the mouth before the tail (Fig. 2C). At the mudsnake’s head, the rest of the viper’s body elevated 00:27 h, the viper had reached the mudsnake’s tail and to the top of the mound. At this point, the viper released finished consuming its meal by 00:29 h (Fig. 2D). Shortly the mudsnake’s head, grasped it near the middle of its thereafter, the viper gave a characteristic snake yawn to body, and hoisted the mudsnake up onto the mound (Fig. realign its jaws and slowly slithered off. A video showing 1C). In what we believe to be an attempt by the viper to a portion of the predation event is available at https:// secure a better hold of its prey, the viper slid down a tree youtu.be/S2OT2fJf6OI. root that was sticking out of the mud next to it (Fig. 1C). Our encounter is the first live field observation of By the end of this action, 15 min had elapsed (time 00:05 ophiophagy in T. purpureomaculatus and the first h) and the viper commenced ingesting the mudsnake from observation of a F. leucobalia being eaten by a viperid the middle of its body (Fig. 1D). (Voris and Murphy, 2002). Previously, Pauwels et al. The entire ingestion phase lasted 24 min. The viper (2000) reported that a captured T. purpureomaculatus began eating the mudsnake in typical viper fashion by in Thailand contained the digestive remains of using its fangs to walk along the mudsnake’s body and another viper, which they identified as a congener. As using its throat muscles to forcefully ingest the snake part of a larger study characterizing the venom of T. Figure 1. A predatory encounter between a mudsnake (Fordonia leucobalia) and mangrove viper (Trimeresurus purpureomaculatus) in Pasir Ris Park Mangroves, Singapore. (A) The writhing mudsnake (red circle) is being pursued by the viper (blue circle). (B) The viper seized the mudsnake by its head, but then (C) switched its grasp to midbody in order to hoist its prey up onto a mud mound to (D) begin eating it. Photographs by Ryan McCleary.
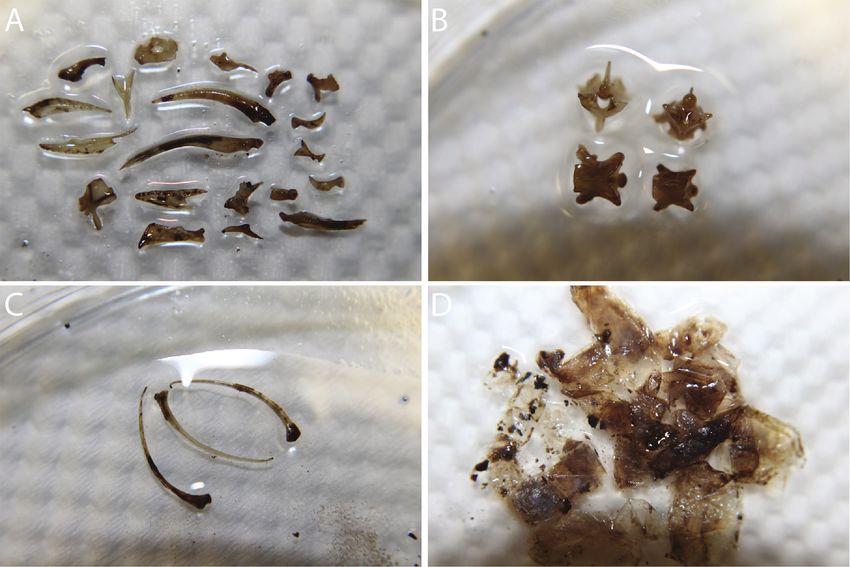
Ophiophagy by Trimeresurus purpureomaculatus on Fordonia leucobalia 1283 Figure 2. A predatory encounter between a mudsnake (Fordonia leucobalia) and mangrove viper (Trimeresurus purpureomaculatus) in Pasir Ris Park Mangroves, Singapore. (A) The viper is using its fangs to ingest the folded body of the mudsnake, while (B) positioning its tail on a mangrove root for support. The viper proceeded to consume the mudsnake at midbody with (C) the head entering the mouth first, before (D) the tail. Photographs by Alex Figueroa. purpureomaculatus, RJRM gathered an amalgamation (Karns et al., 2002). Cerberus schneiderii is often the of snake bones (Fig. 3A), that included vertebrae, ribs, most abundant snake at any mangrove in Southeast and scales (Fig. 3B–D) found in faecal samples excreted Asia (Murphy, 2007), including Sungei Buloh Wetland from three different T. purpureomaculatus collected at Reserve in Singapore (Chim and Diong, 2013), Peninsular PRPM. Mixed in one of the faecal samples was also the Malaysia (Jayne et al., 1987), and Bangladesh (Rahman partially digested claw of a crab, indicating the viper had et al., 2014), constituting a bountiful food source. Thus, if eaten one of two crustacean-eating homalopsid species, F. T. purpureomaculatus does indeed prefer to eat snakes, it leucobalia or Gerarda prevostiana (Eydoux & Gervais, has at least one dominant species to rely upon. 1837). Our observations, along with that of Pauwels et The method in which the T. purpureomaculatus ingested al. (2000), support that T. purpureomaculatus, at least the F. leucobalia is unusual. Snakes typically consume occasionally, forages on the ground (David and Vogel, their prey head-first, but sometimes may ingest prey from 1996), offering the possibility that T. purpureomaculatus the caudal region (Cundall and Greene, 2000) or tail first may prey on snakes regularly. Thus, the overarching in the case of ophiophagy in Erythrolamprus Boie, 1826 question that should be investigated in future studies (Braz and Marques, 2016). Ingesting the mudsnake from is whether T. purpureomaculatus prefers feeding on midbody was likely a failure or carelessness on the part snakes or whether they are only opportunistically of the viper to locate the mudsnake’s head, or urgency consuming snakes, possibly due to restricted availability to consume its prey. The viper did appear to search for of preferred prey. the mudsnake’s head but then abandoned the pursuit at In Southeast Asia, snakes are a prominent inhabitant midbody and began ingesting the snake. However, it is of mangroves. Pasir Ris Park Mangroves is home to at interesting to propose that perhaps feeding on elongate least eight other snake species, with Cerberus schneiderii prey at midbody, with the body of the prey doubled over, (Schlegel, 1837) being indisputably the most common may serve as a means of overcoming the biomechanical
1284 Alex Figueroa & Ryan J.R. McCleary
Figure 3. Photographs of snake remains found in the faeces of Trimeresurus purpureomaculatus from Pasir Ris Park Mangroves,
Singapore. The remains included (A) a mixture of cranial bones, (B) vertebrae, (C) ribs, and (D) scales. Photographs by Ryan
McCleary.
challenge of accommodating such a long prey item in the References
gastrointestinal tract (Jackson et al., 2004). Alternatively, Banci, K.R.S., Viera, N.F.T., Freitas, A.C., Marques, O.A.V. (2017):
ingesting elongated prey from midbody may also serve Feeding on elongate prey: additional data for the coral snake
to reduce handling time as it took the viper 24 min to Micrurus corallinus (Merrem, 1820) (Elapidae) and comments
fully ingest the mudsnake. This time is at the lower on aposematism. Herpetology Notes 10: 335–338.
limit of ingestion time of elongate prey in snakes but Braz, H.B., Marques, O.A. (2016): Tail-first ingestion of prey by
is still much greater than ingesting non-elongate prey the false coral snake, Erythrolamprus aesculapii: Does it know
where the tail is? Salamandra 52(2): 211–214.
(Banci et al., 2017). Our observation documents the
Chim, C.K., Diong, C.H. (2013): A mark-recapture study of a
first instance of a viper feeding on a homalopsid, and dog-faced water snake Cerberus schneiderii (Colubridae:
this substantiates that T. purpureomaculatus descends Homalopsidae) population in Sungei Buloh Wetland Reserve,
from the trees to forage on the ground. The frequency Singapore. Raffles Bulletin of Zoology 61: 811–825.
of these behaviours remains unknown and awaits further Coelho Lima, A.D., Oliveira Ramos, G., Martins, R.B.X., Castro
study. Perhaps, T. purpureomaculatus only uses arboreal Meira, L.P.D. (2020): Fist record of ophiophagy in the false
settings for escaping the daily mangrove tidal events, coral snake Oxyrhopus trigeminus Duméril, Bibron & Duméril,
1854. Cuadernos de Herpetología, 34(1) 89–91.
evading predators, and basking.
Cundall, D., Greene, H.W. (2000): Feeding in snakes. In: Feeding:
Form, Function, and Evolution in Tetrapod Vertebrates, p. 293–
Acknowledgements. This observation was made during a survey 333. Schwenk, K., Ed., New York, USA, Academic Press.
covered by research permit NP/RP10-095-3b issued to RJRM by David, P., Vogel, G. (1996): The Snakes of Sumatra, an Annotated
the National Parks Board. We would like to thank John Ascher Checklist and Key with Natural History Notes. Frankfurt am
at the National University of Singapore for allowing use of Main, Germany, Edition Chimaira.
his microscope. We also want to thank the reviewers for their Glaudas, X., Kearney, T.C., Alexander, G.J. (2017): Museum
comments and suggestions, and Jason Fantuzzi for his insight specimens bias measures of snake diet: a case study using the
into snake feeding. ambush-foraging puff adder (Bitis arietans). Herpetologica
73(2): 121–128.Ophiophagy by Trimeresurus purpureomaculatus on Fordonia leucobalia 1285
Greene, H.W. (1997): Snakes: the Evolution of Mystery in Nature. Murphy, J.C. (2007): Homalopsid Snakes. Evolution in the Mud.
Berkeley, California, USA, University of California Press. Malabar, Florida, USA, Krieger Publishing Company.
Jackson, K., Kley, N.J., Brainerd, E.L. (2004): How snakes Pauwels, O.S.G., Laohawat, O.-A., David, P., Bour, R., Dangsee,
eat snakes: the biomechanical challenges of ophiophagy for P., Puangjit, C., Chimsunchart, C. (2000): Herpetological
the California kingsnake, Lampropeltis getula californiae investigations in Phang-Nga Province, southern peninsular
(Serpentes: Colubridae). Zoology 107(3): 191–200. Thailand, with a list of reptile species and notes on their biology.
Jayne, B.C., Voris, H.K., Kiew, B.H. (1987): Diet, feeding Dumerilia 4: 123–154.
behavior, growth, and numbers of a population of Cerberus Rahman, S.C., Reza, A.A., Datta, R., Jenkins, C.L., Luiselli,
rynchops (Serpentes: Homalopsinae) in Malaysia. Fieldiana L. (2014): Niche partitioning and population structure of
Zoology 50: 1–15. sympatric mud snakes (Homalopsidae) from Bangladesh. The
Karns, D.R., Voris, H.K., Goodwin, T.G. (2002): Ecology of Herpetological Journal 24(2): 123–128.
oriental-Australian rear-fanged water snakes (Colubridae: Uetz, P., Freed, P., Hošek, J. (2019): The Reptile Database.
Homalopsinae) in the Pasir Ris Park mangrove forest, Singapore. Available at: http://www.reptile-database.org. Accessed on 21
Raffles Bulletin of Zoology 50: 487–498. March 2021.
McKelvy, A.D., Figueroa, A., Lewis, T.R. (2013): First record Voris, H.K., Murphy, J.C. (2002): The prey and predators of
of ophiophagy in the widely distributed snake Leptodeira homalopsine snakes. Journal of Natural History 36: 1621–1632.
septentrionalis (Kennicott, 1859) (Ophidia, Colubridae).
Herpetological Notes 6: 177–178.
Accepted by Justin BernsteinYou can also read

















































