Berichte Jahrgang 52 Heft 1 - Monash University
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Berichte Reports
Jahrgang 52 Volume 52
Heft 1 No. 1
2020
Wissenschaftliche Gesellschaft Swakopmund
Scientific Society Swakopmund
P.O. Box 361, Swakopmund, Namibia
Tel.: 264-(0)64-402695
Fax: 264-(0)64-400763
office@scientificsocietyswakopmund.com
www.scientificsocietyswakopmund.com
ISSN 2026-7649
Wissenschaftliche Gesellschaft Scientific Society
SwakopmundSwakopmund
NamibiaNamibia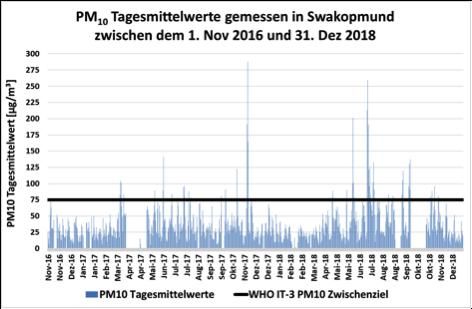
Herausgeber / Publisher Inhalt / Contents
Wissenschaftliche Gesellschaft Swakopmund
Scientific Society Swakopmund
P.O. Box 361, Swakopmund, Namibia Seite / Page
Tel: 264-(0)64-402695
Fax: 264-(0)64-400763 Crossing the line
office@scientificsocietyswakopmund.com The Ediacaran – Cambrian Transition in Southern
www.scientificsocietyswakopmund.com
Namibia – how the world began to change @ 538 million
Years ago
Kontaktadresse in Deutschland / German contact address P. Vickers-Rich et al. ......................................................................... 2
Deutsch-Namibische Gesellschaft e.V.
Sudetenlandstr. 18, 37085 Göttingen
Tel.: +49-551-7076781
Fax: +49-551-7076782 Feinstaub- und Radonkonzentration in Namibias
Erongo Region
Kontoverbindung / Account details Detlof von Oertzen & Hanlie Liebenberg-Enslin ........................... 26
Standard Bank Swakopmund, Namibia
Konto Nr. / Account No. 04 137 7230
Bankleitzahl / Branch code 082-172
Commerzbank, Düsseldorf-Königsallee, Deutschland Neueingänge in der Sam-Cohen-Bibliothek
IBAN DE91 3008 0000 0211 3508 02 New books in the Sam Cohen Library ........................................ 37
BIC DRESDEFF300
© Wissenschaftliche Gesellschaft Swakopmund, 2020
© Scientific Society Swakopmund, 2020
Layout: Gudrun Berens www.geeart.com
Druckerei / Printer: John Meinert Printing
Für den Inhalt der Beiträge sind die Verfasser verantwortlich.
The views of the authors are not necessarily those of the
Scientific Society Swakopmund.
ISSN 2026-7649CROSSING THE LINE Daten werden gesammelt, die sehr wichtige Hinweise geben, um die
Wechselwirkung von Leben und sich verändernden Umweltbedingungen
The Ediacaran – Cambrian Transition in Southern Namibia – zu verstehen. Dies ist auch höchst relevant, um die Zukunft unseres
How the World Began to Change @ 538 million years ago Planeten vorherzusagen.
P. Vickers-Rich, Monash University, Melbourne, Australia pat.rich@ ABSTRACT
monash.edu; K. Mhopjeni, Geological Survey of Namibia, Windhoek, Namibia hosts a near complete sequence of sediments and volcanics that
Namibia kombada.mhopjeni@mme.gov.na; G. Schneider Namibian record a period from the late Neoproterozoic into the early Phanerozoic
Uranium Institute, Swakopmund, Namibia director@namibianuranium.
(600-500 million years ago) when the biota on Planet Earth changed
org with H. Cui, S. Darroch, D. Elliott, M. Fedonkin, M. Hall, K.H.
from enigmatic life to life that we consider normal today, including the
Hoffmann, M. Hofmann, A. Ivantsov, A. Kaufman, L. Kriesfeld, M.
appearance of Animalia – forms with eyes, guts, the ability to move
Laflamme, U. Linnemann, H. Mocke, G. Narbonne, S. Pritchard, T.
and even hard skeletons. Older rocks of this sequence contain biota
Rich, A. Sharp, J. Smith, P. Swinkels, P. Trusler & M. Zakrevskaya
dominated by the so-called Ediacarans, which lacked burrowers,
depended greatly on massive microbial mats and chemistry of the
Zusammenfassung
surrounding ocean for nourishment, most of them lacked eyes, guts,
In Namibia kommt eine fast vollständige Abfolge von Sedimenten
and only few were able to move. All that changed when that Line
und vulkanischen Gesteinen vor, die die gesamte Zeit vom späten
Neoproterozoikum bis zum frühen Phanerozoikum (vor 600-500 was Crossed around 538-539 million years ago. Rocks in Southern
Millionen Jahre) umspannt. In dieser Zeit veränderte sich das Leben Namibia host many of the possible answers to just what happened at
auf der Erde von recht rätselhaften Kreaturen hin zu den Lebewesen, this time, and researchers have over the past few decades been able to
die wir heute kennen. Dazu gehörte auch das Auftreten von Mitgliedern more precisely date just when this transition occurred and what were
der Tierwelt – Kreaturen mit Augen, Verdauungsorganen, der Fähigkeit the environmental drivers to this change. Their research is showcasing
sich zu bewegen, und mit harten Knochen. Die älteren Gesteinsschichten early complex, megascopic lifeforms of the Ediacarans, some of which
enthalten allerdings vorwiegend die sogenannte Ediacara Fauna, die were predecessors of true animals. This research continues now with
für ihre Nahrungsaufnahme stark von massiven Algenmatten und der a global network of scientists continuing to add to the ever-increasing
Chemie des sie umgebenden Ozeans abhängig waren, die meisten database, which is so critical to understanding the interaction of life
hatten keine Augen und Verdauungsorgane, und nur ein paar von ihnen and changing environmental conditions. This is also very relevant for
konnten sich bewegen. Das alles änderte sich vor 538-539 Millionen predicting the future of Planet Earth.
Jahren, als die Grenze überschritten wurde. Gesteine im südlichen
Namibia geben Aufschluss über das, was genau in dieser Zeit passierte, Namibia has been a key region for understanding some unusual
und Wissenschaftler waren über die letzten Jahrzehnte in der Lage, den organisms called Ediacarans since the early days of the 20th century,
genauen Zeitpunkt dieser Übergangsperiode zu bestimmen, sowie die when geologists, such as Paul Range and German soldiers stationed at
Umweltveränderungen, die für den Übergang verantwortlich waren, isolated outposts in the Aus region of southern Namibia, first reported
zu untersuchen. Die Forschungsergebnisse zeigen die Ediacara Fauna strange fossils from there. These were the first large multicellular
als frühe, komplexe, mehrzellige Lebensformen, von denen manche organisms that prospered on Planet Earth before the development of
die Vorläufer der richtigen Tiere waren. Die Forschung wird nun von animals. Their fossils are preserved in the thick rock sequences of the
einem globalen Netzwerk von Wissenschaftlern fortgesetzt, und neue Nama Group in southern Namibia and showcase a time in the history
2 3of life when there were fundamental and pivotal changes from an Europe, that hosted true skeletonized animals – trilobites and shelled
enigmatic biota to the biota we know today (Fig. 1). forms related to clams and snails.
It has been more than 90 years since Gürich’s naming of Rangea,
and since that time a variety of other Ediacaran forms have also been
named and became bell-weather taxa for our changing interpretations
of the Ediacara biota. Rangea was originally regarded as a primitive
relative of the ctenophores or corals (Gürich, 1930, 1933; Dzik, 2002,
Richter, 1955; Jenkins, 1985, 1992). Much later, researchers such
as Seilacher and his colleagues (Seilacher, 1992; Grazhdankin and
Seilacher, 2005; Seilacher and Gishlick, 2014) removed Rangea from
the group Animalia and regarded it as a core member of what Seilacher
proposed as a new, extinct kingdom of organisms, which he named the
‘Vendobionta’. Hans Pflug (1970a, b, 1972) designated Rangea as the
type of a key Ediacaran division of life, the Rangeomorpha, which had
over time a global distribution. His view was subsequently endorsed
and refined by others (Narbonne, 2004; Gehling and Narbonne, 2007;
Laflamme and Narbonne, 2008; Xiao and Laflamme, 2009; and Erwin
Fig. 1. The Nama Group of Southern Namibia hosts a rich collection of Ediacaran et al., 2011, Laflamme et al., 2013). More discoveries (Vickers-Rich
organisms that prospered more than 530 million years ago, prior to the appearance et al., 2013) have added significant, complete, three-dimensional
of true animals on planet Earth
specimens recovered from ancient, small, near-shore marine channels
on a farm in southern Namibia. These discoveries have allowed
The first formal name for one of these complex Ediacaran life Rangea to be reconstructed in detail as a six-vaned, multifoliate frond
forms was given by Gürich in 1930. He proposed the name Rangea with an expanded basal bulb, which served as a weight-belt anchor,
schneiderhoehni for fossils of the enigmatic, cm-scale frond-like with the living organism situated on, or perhaps with the basal bulk
organism collected from the Dabis Formation in the south of the only slightly buried in the floor of an ancient, shallow ocean (Figs 2a,
country. Rangea predated Sprigg’s (1947) description of Dickinsonia 2b, 3a, 3b and 4).
from the Flinders Ranges in Australia by nearly two decades.
Dickinsonia fossils also formed part of the Ediacara biota and the
official name for this geological period is based on these fossils from
Ediacara. Likewise, the official naming of other Ediacaran fossils
in Ford’s (1958) description of Charnia from Charnwood Forest in
England happened nearly three decades after Gürich’s seminal paper.
Rangea schneiderhoehni was not a simple disc that could be described
and then forgotten – it was a frond covered with features so complex
that Gürich assumed Rangea must be of a younger Cambrian age, a
time period well known by then, especially from rocks in England and
4 5Fig. 3a
Fig. 2a. Peter Trusler’s reconstruction of Rangea
Figs 3a+b. 3-d scans of 2 of more than 200 specimens of Rangea discovered by
participants of IGCP 493, 587 and 673, provided by a cooperative study funded by
the Australian Synchrotron in Melbourne
Fig. 2b. Three-dimensional specimen of Rangea discovered by
participants of International Geological Cooperation
Programmes (IGCP) 493, 587 and 673
6 7lifestyle is still controversial, with some specimens reported to have
been preserved in situ at the top of the Swartpunt section (Narbonne
et al., 1997; Darroch et al., 2015). Traditional views (e.g. Jenkins,
1985) regarded them as fronds extending into the water column, but
this was disputed by Seilacher (1989; 1992) and Grazhdankin and
Seilacher (2005), who regarded Pteridinium as mainly or completely
living in soft sediments at the bottom of the water column. Subsequent
discoveries of material preserved in massive downslope avalanches,
again on farms in southern Namibia (Elliott et al., 2011; Meyer et al.,
2014a; 2014 b) have challenged this lifestyle interpretation, and so
just how these tubular organisms lived still remains a mystery.
Fig. 4. Interpretation of life position and preservation of Rangea in the shallow,
nearshore marine setting of the ancient Nama sea some 540 million years ago (art by
Peter Trusler)
Members of another group of organisms nearly unique to the Nama
Group sediments in southern Namibia are the Erniettomorphs –
Ernietta, Pteridinium, and Swartpuntia. Pteridinium simplex (Figs 5a
and 5b) was first described by Gürich in 1933, and its body consisted
Fig. 5a. A painting by palaeo-artist Peter Trusler of the preserved
of a double-layered palisade of tubes, an architecture quite distinct
past and the present, illustrating a lock containing Pteridinium with the
from the ‘fractal’ morphology of Rangea. Thus far, most specimens modern landscape where it was discovered in Neoproterozoic rocks
of Pteridinium have been found in transported sediments, and their
8 9fossils on the neighbouring rocks provide the graphic inspiration for
other meanings (Figs 6a, b+c).
Fig. 5b. Pteridinium fossil from Aarhäuser in southern Namibia in an
avalanche mass flow of an ancient ocean
Spectacular discoveries of the first ever truly complete specimens
of Ernietta (Pflug, 1966) were found by Andrey Ivantsov as part of
a UNESCO International Geosciences Project in the early 2000’s.
Ernietta seems to have been a bag-shaped organism, also constructed
of the double-layered palisade of parallel tubes diagnostic of the
Erniettomorpha. Before Andrey’s discovery, Ernietta was known
from hundreds of specimens, most of them transported, and not in
the place where they once lived (Elliott et al., 2016). All specimens Fig. 6a. Ernietta: one of the complete specimens found
up until this discovery were also incomplete. But the new specimens
were preserved at the base of small, near shore marine channels, and
appear to have lived in close-knit colonies. Some were absolutely
complete and consisted of a basal bag-shaped anchor, permanently
buried in the sediment, with the more distal part extending upwards
into the water column, consisting of two facing structures with fronds
at the distal opening (Ivantsov et al., 2015). Using a combination
of field observations and computational fluid dynamics modeling,
Gibson et al. (2019) recently illustrated that living sociably in
aggregated populations was almost certainly an ecological adaptation
that aided in feeding; multiple individuals living together would have
increased vertical mixing in the water column, helping to transport
food particles to their neighbors downstream, much in the way that
modern oysters and mussels do. San artists had long ago seen these
Fig. 6b. Ernietta: Reconstructions crafted by Peter Trusler,
fossils and left carved images in the rocks: could these have been the
based on these new discoveries
first to document the detail of the complete morphology or did these
10 11Other, rarer Ediacara-type impressions from Namibia include
Protechiurus (Glaessner, 1979) and Ausia (Hahn and Pflug, 1985). Most
of these taxa are based on one or a few specimens, and consequently of
uncertain affinities. Carbonaceous tubular compression fossils such as
Vendotaenia are locally common (Cohen et al., 2009). Until recently,
Protechiurus edmondsi was known from a single specimen, it has been
described as a cast of the body of a worm-like organism (Glaessner,
1979). The study of new findings made it possible to interpret this
fossil as remains of a conical hexahedral enclosure of an organism
close to the coelenterates (Ivantsov et al., 2019) (Figs 8a, b+c).
Fig. 6c. Ernietta: Engravings made long ago by San artists
Yet another mysterious fossil is Swartpuntia (Narbonne et al., 1997).
It was a multifoliate, frondose Erniettomorph that reached over a
decimeter in height. It is abundant at the top of Swartpunt where geo-
chronological and geochemical data imply it lived during the last one
million years of the Ediacaran Period, alongside Pteridinium (Fig. 7).
Fig. 8a. Joint burial of Ernietta, Pteridinium and Protechiurus
Fig. 7. Swartpuntia from near the top of the section on Swartpunt
12 13Beltanelliformis (Nemiana) is abundant in shallow-water Ediacaran
assemblages worldwide, and is most commonly interpreted as a
spherical ‘polyp’ of unknown affinities (Narbonne and Hofmann, 1987)
or as a Nostoc-like ball of bacteria (Ivantsov et al., 2015). Clearly,
there is still a lot to sort out with respect to the true relationships of
many of these early, multicellular, large organisms, and that is what
makes further work on the fossils from the late Precambrian and early
Phanerozoic in Namibia an exciting challenge.
Fig. 9. Beltanelliformis from the area near Aus in southern Namibia
Besides these megascopic mysterious organisms, another major
Fig. 8b. Isolated three-dimensional cast Fig. 8c. Reconstruction of contribution that the Namibian late Neoproterozoic fossil record
of theca of Protechiurus edmondsi Protechiurus edmondsi continues to make to understanding the progress of life on Earth is
the abundant occurrence of the world’s first skeletal fossils that are
Circular, cm-scale discs comprise another common taxon, and these
preserved in a number of carbonate rocks that occur throughout the
invariably occur in profusion, completely covering bedding planes
Nama Group. Some of these early forms with a hard skeleton played
with adjoining specimens, all of them exactly the same size and
a major role in building some of the world’s first skeletal-microbial
presumably the result of a single spatfall (the settling and attachment reefs (Germs, 1972; Grant, 1990; Grötzinger et al., 2000, 2005; Wood
of young organisms to the substrate). These fossils were originally et al., 2002; Wood et al., 2015; Penny et al., 2014; Zhuravlev et al.,
named Hagenetta aarensis (Hahn and Pflug, 1988) and were 2015). Cloudina (Germs, 1972), the first pre-Cambrian shelly fossil
interpreted as bivalve shells, but Ivantsov et al. (2015) suggested that named anywhere in the world, consists as mm-scale diameter calcified
the name Beltanelliformis has priority over the other common names tubes with a distinctive pattern of stacked, funnel-shaped transverse
(e.g. Nemiana) that have previously been used for this taxon (Fig. 9). partitions inside the tube. Most researchers consider it likely that
14 15Cloudina was a true animal. Namacalathus (Grötzinger et al., 2000) is The trigger of the earliest animal biomineralization, so well
another calcified reef-dweller, which, like Cloudina, occurs as skeletal documented in the Namibian record, has been proposed to be the
packstones between microbial elements in the reefs. Its structure is a ecological pressure from predation (Hua et al., 2003). However,
cm-scale cup that sits atop a short stem, with a hole in the top of the more recent chemostratigraphic studies of the late Ediacaran strata
cup and several holes along its sides. It also has been considered to in Namibia (Tostevin et al., 2019a), South China (Cui et al., 2016;
be an animal, a metazoan that Zhuravlev et al. (2015) considered as Cui et al., 2019a; Cui et al., 2019b), Siberia (Wood et al., 2017), and
a lophophorate, an animal with a fan of tentacles around the mouth, Brazil (Caetano-Filho et al., 2019; Macedo Paula-Santos et al., 2019)
regarded by others as an ancestral lophophotrochozoan (Darroch, reveal that the evolutionary novelty of animal biomineralization was
pers. comm., 2020) (Figs 10a+b). probably environmentally triggered by enhanced chemical weathering
and high seawater alkalinity at that time. Recent uranium isotope
analyses of the Ediacaran-Cambrian strata in Namibia (Tostevin et al.,
2019b) and South China (Wei et al., 2018; Zhang et al., 2018) suggest
an expansion of anoxia in terminal Ediacaran oceans and dynamic
marine redox fluctuations “across the line”, which may be the trigger
for the demise of the Ediacarans and the rise of the Cambrian animals
(Figs 11a+b).
Fig. 10a. Cloudina, highly magnified
Fig. 10b. Cloudina, with scale. Specimen from southern Namibia Fig. 11a
16 17and biological drivers of extinction, respectively. Only more data and
new fossil discoveries will allow us to test between these two models,
and determine what eventually led to the disappearance of the Ediacara
biota about 538-539 million years ago (Fig 12).
Fig. 11b. The changes across the Precambrian-Cambrian Boundary – the crossing
of a critical line for Life on Earth: (left a biota dominated by the Ediacarans,
which lacked burrowers, depended greatly on massive microbial mats and chemistry
Fig. 12. IGCP 673 field workshop in March 2019
of the surrounding ocean for nourishment, most of them lacked eyes, guts, and only
few were able to move (art by P Trusler); (right) life after that Line was Crossed,
when the Cambrian world looked more like the modern world (art by LM Tolpygin, Intriguingly, a biological driver of extinction would raise uncomfort-
courtesy of the Orlov Paleontological Museum, Moscow) able parallels with the present day, where humans are altering our envi-
ronment at a pace which threatens millions of species. In this fashion,
An intriguing question is what happened to the last of these first this supposed ‘first mass extinction’ and end of the Ediacaran may of-
complex and large organisms that developed on Earth? Recent fer clues and an invaluable lesson for interpreting the present biodiver-
analyses by Darroch et al. (2015; 2018) illustrate that the latest sity crisis. In the future, such knowledge will surely be of fundamental
Ediacaran communities preserved in the Nama Group were relatively value in using our understanding of what happened to the Ediacarans
species-poor, and perhaps already in the throes of an extinction event. and the dynamics of the dramatic biotic change at the beginning of the
This decrease in the diversity of Ediacara biota worldwide coincides Cambrian...and give much needed guidance on p lanning the future of
with both evidence for regional anoxia (e.g. Wood et al., 2015), and humanity! Just how we use that understanding will most certainly be
the rise of more complex Cambrian-type behaviors and ‘ecosystem critical to our survival as a species. We just hope humanity listens!
engineering’, providing potential support for competing environmental
18 19minal Ediacaran Earth-life system in South China and Arctic
Siberia. Estudios Geológicos, 75, e097
Cui, H, Xiao, S, Cai, Y, Peek, S, Plummer, RE & Kaufman, AJ
(2019b): Sedimentology and chemostratigraphy of the termi-
nal Ediacaran Dengying Formation at the Gaojiashan section,
South China. Geological Magazine, 156, 1924–1948 https://
doi.org/10.1017/S0016756819000293
Cui, H, Kaufman, AJ, Xiao, S, Peek, S, Cao, H, Min, X, Cai, Y, Siegel,
Z, Liu, XM, Peng, Y, Schiffbauer, JD, &Martin, AJ (2016)
Environmental context for the terminal Ediacaran biominer-
alization of animals. Geobiology, 14, 344–363, https://doi.
org/10.1111/gbi.12178
Darroch, SAF, Sperling, EA, Boag, T, Racicot, RA, Mason, SJ,
Morgan, AS, Tweedt, S, Myrow, P, Erwin, DH & Laflamme,
M (2015): Biotic replacement and mass extinction of the
Ediacara biota. Proceedings of the Royal Society, B, 282.
Fig. 13. Swartpunt in the distance – the “avenue” where the last vestiges
of the Ediacarans, are replaced by the Modern World of the Cambrian –
Darroch, SAF, Smith, E, Laflamme, M & Erwin, DH (2018): Ediacaran
where the Line is Crossed extinction and Cambrian Explosion. Trends in Ecology and
Evolution, 33, 653-663
References Dzik, J (2002): Possible Ctenophoran affinities of the Precambrian
Caetano-Filho, S, Macedo Paula-Santos, G, Guacaneme, C, Babinski, “seapen” Rangea. Journal of Morphology, 252, 315-334
M, Bedoya-Rueda, C, Peloso, M, Amorim, K, Afonso, Elliott, DA, Vickers-Rich, P, Trusler, P, & Hall, M (2011): New
J, Kuchenbecker, M, Reis, H & Trindade, RIF (2019): evidence on the taphonomic context of the Ediacaran
Sequence stratigraphy and chemostratigraphy of an Ediacaran- Pteridinium. Palaeontologica Polonica, 56 (3), 641-650
Cambrian foreland-related carbonate ramp (Bambuí Group, Elliott, DA, Trusler, P, Narbonne, G, Vickers-Rich, P, Hoffmann, K H
Brazil). Precambrian Research, https://doi.org/10.1016/j. & Schneider, G I C (2016): Ernietta from the Late Ediacaran
precamres.2019.105365 Nama Group, Namibia. Journal of Paleontology, 90, DOI
Cohen, PA, Bradley, A, Knoll, HA, Gr������������������������������
öt����������������������������
zinger, JP, Jensen, S, Abel- 10.1017/jpa.2016.94.
son, J, Hand, K, Love, G, Metz, J, McLoughlin, N, Meister, Erwin, DH, Laflamme, M, Tweedt, SM, Sperling, EA, Pisani, D
P, Shepard, R, Tice, M & Wilson, JP (2009): Tubular com- & Peterson, KJ (2011): The Cambrian conundrum: Early
pression fossils from the Ediacaran Nama Group, Namibia. divergence and later ecological success in the early history of
Journal of Paleontology, 83 (1), 110–122. animals. Science, 334, 1091-1097
Cui, H, Kaufman, AJ, Xiao, S, Grazhdankin, DV, Peek, S, Martin, Ford, T (1958): Precambrian fossils from Charnwood Forest.
AJ, Bykova, NV, Rogov, VI, Liu, X-M, Zhang, F, Romaniello, Proceedings of the Yorkshire Geological Society, 31, 211-217
SJ, Anbar, AD, Peng, Y, Cai, Y, Schiffbauer, JD, Meyer, M, Gehling, JG & Narbonne, G M (2007): Spindle-shaped fossils from
Gilleaudeau, GJ, Plummer, RE, Sievers, NE, Goderis, S & the Mistaken Point assemblage, Avalon Zone, Newfoundland.
Claeys, P (2019a): Recent advances in understanding the ter- Canadian Journal of Earth Sciences, 44, 367-387
20 21Germs, G.J B (1972): New shelly fossils from Nama Group South exceptional specimens in the Ediacaran of Namibia. Lethaia,
West Africa. American Journal of Science, 272, 752-761 DOI:10.1111/let.12164: 1-15
Gibson, BM, Rahman, IA, Maloney, KM, Racicot, RA, Mocke, H, Ivantsov, AY, Vickers-Rich, P, Zakrevskaya, MA & Hall, M
Laflamme, M & Darroch, SAF (2019): Gregarious suspension (2019) Conical thecae of Precambrian macroorganisms.
feeding in a modular Ediacaran organism. Science Advances, Paleontological Journal, 53 (11), 18-30
5(6), DOI: 10.1126/sciadv.aaw0260 Jenkins, R J F (1985): The enigmatic Ediacaran (Late Precambrian)
Glaessner, M F (1979): An echiruid worm from the Late Precambrian. genus Rangea and related forms. Paleobiology, 11 (3), 336-
Lethaia, 12, 121-124 355
Grant, SWF (1990): Shell, structure and distribution of Cloudina, a Jenkins, RJF (1992): Functional and ecological aspects of Ediacaran
potential index fossil for the terminal Proterozoic. American assemblages. In: Lipps, JH & Signor, PW: Origin and Early
Journal of Science, 290-A, 261-294 Evolution of the Metazoa, Plenum Press, New York, 131-176
Grazdankin, D & Seilacher, A (2005): A re-examination of the Nama Laflamme, M & Narbonne, G (2008) Ediacaran fronds.
Palaeogeography, Palaeoclimatology, Palaeoecology, 258,
type Vendian organism Rangea schneiderhoehni. Geological
162-179
Magazine, 142, 571-582
Laflamme, M, Darroch, SAF, Tweedt, S, Peterson, KJ & Erwin, DH
Grötzinger, J P, Adams, E W & Schröder, S (2005): Microbial-metazoan
(2013): The end of the Ediacara biota: extinction, biotic
reefs of the terminal Proterozoic Nama Group (c. 550–543
replacement, or Cheshire cat? Gondwana Research, 23,
Ma), Namibia. Geological Magazine, 142, 499-517
558-573
Grötzinger, JP, Watters, W A & Knoll, A H (2000): Calcified metazoans
Macedo Paula-Santos, G, Caetano-Filho, S, Enzweiler, J, Navarro,
in thrombolite-stromatolite reefs of the terminal Proterozoic MS, Babinski, M, Guacaneme, C, Kuchenbecker, M, Reis, H
Nama Group, Namibia. Paleobiology, 26 (3), 334-359 & Trindade, RIF (2019): Rare earth elements in the terminal
Gürich, G (1930): Die bislang ältesten Spuren von Organismen in Ediacaran Bambuí Group carbonate rocks (Brazil): evidence
Südafrika. Compte Rendu XV International Geological for high seawater alkalinity during rise of early animals. Pre-
Congress, 2, Palaeontologische Zeitschrift, VII, 670-681 cambrian Research, 336, 105506 https://doi.org/10.1016/j.
Gürich, G (1933): Die Kuibis-Fossilien der Nama Formation von precamres.2019.105506
Südwestafrika. Palaeontologische Zeitschrift, 15, 137-154 Meyer, M, Elliott, D, Schiffbauer, JD, Hall, M, Hoffmann, KH,
Hahn, G & Pflug, HD (1985): Polypenartige Organismen aus dem Schneider, G, Vickers-Rich, P & Xiao, S (2014 a): Taphonomy
Jung-Präkambrium (Nama-Gruppe) von Namibia. Geologica of the Ediacaran fossil Pteridinium simplex preserved three-
et Palaeontologica, 19, 1-13 dimensionally in mass flow deposits, Nama Group, Namibia.
Hahn, G & Pflug, HD (1988): Zweischalige Organismen aus dem Journal of Paleontology, 88 (2), 1-12
Jung- Präkambrium (Vendium) von Namibia (SW-Afrika). Meyer, M, Elliott, D, Wood, AD, Polys, NF, Colbert, M, Maisano,
Geologica et Palaeontologica, 22, 1-19 JA, Vickers-Rich, P, Hall, M, Hoffmann KH, Schneider, GIC
Hua, H, Pratt, BR & Zhang, L-Y (2003): Borings in Cloudina & Xioa, S (2014b): Three-dimensional microCT analysis of
shells: complex predator-prey dynamics in the terminal the Ediacara fossil Pteridinium simplex sheds new light on
Neoproterozoic. PALAIOS, 18(4-5), 454–459, https://doi. its ecology and phylogenetic affinity. Precambrian Research,
org/10.1669/0883-1351(2003)0182.0.co;2 249, 79-87, DOI 10.1016/j.precamres.2014.04.013
Ivantsov, AY, Narbonne, GM, Trusler, PW, Greentree, C & Vickers- Narbonne, G & Hoffman, HJ (1987): Ediacaran biota of the Wernecke
Rich, P (2015): Elucidating Ernietta: new insights from Mountains, Yukon, Canada. Palaeontology, 30, 647-676
22 23Narbonne, G M (2004): Modular construction of early Ediacaran oceans. Earth and Planetary Science Letters, 506, 104–112,
complex life forms. Science, 305, 1141-1144 https://doi.org/https://doi.org/10.1016/j.epsl.2018.10.045
Narbonne, GM, Saylor, BZ & Grötzinger, JP (1997): The youngest Vickers-Rich, P, Ivantsov, AY, Trusler, PW, Narbonne, G, Hall,
Ediacaran-fossils from southern Africa. Journal of M, Wilson, SA, Greentree, C, Fedonkin, MA, Elliott, DA,
Paleontology, 71, 953-967 Hoffmann, KH & Schneider, GIC (2013): Reconstructing
Penny, A M, Wood, R, Curtis, A, Bowyer, F, Tostevin, R & Hoffman, Rangea: New discoveries from the Ediacaran of Southern
KH (2014): Ediacaran metazoan reefs from the Nama Group, Namibia. Journal of Paleontology, 87(1), 1-15
Namibia. Science, 344(6191), 1504-1506 Wei, G-Y, Planavsky, NJ, Tarhan, LG, Chen, X, Wei, W, Li, D, Ling,
Pflug, HD (1966): Neue Fossilreste aus den Nama-Schichten in H-F (2018): Marine redox fluctuation as a potential trigger for
Südwest- Afrika. Paläontologische Zeitschrift, 40, 14-25 the Cambrian explosion. Geology, 46, 587–590 https://doi.
I. Pteridinia, Bau und systematische Zugehörigkeit. org/10.1130/G40150.1
Palaeontographica, Abteilung A, 134, 226-262 Wood, RA, Grötzinger, JP & Dickson, JAD (2002): Proterozoic
Pflug, HD (1970b): Zur Fauna der Nama-Schichten in Südwest-Afrika. modular biomineralised Metazoan from the Nama Group,
II. Rangeidae, Bau und systematische Zugehörigkeit. Namibia. Science, 296, 2383-2386
Palaeontographica, 139, 134-170 Wood, R, Ivantsov, AY, Zhuravlev, AY (2017): First macrobiota
Pflug, HD (1972): Zur Fauna der Nama-Schichten in Südwest-Afrika. biomineralization was environmentally triggered. Pro
III. Erniettomorpha, Bau und Systematik. ceedings of the Royal Society B: Biological Sciences,
Palaeontolgraphica Abteilung A, 135, 198-231 284(1851), 20170059 https://doi.org/10.1098/rspb.2017.0059
Richter, R (1955) Die ältesten Fossilien Süd-Afrikas. Lethaia, 36(3/4), Wood, RA, Poulton, SW, Prave, AR, Hoffmann, KH, Clarkson, MO,
243-289, Senckenberg Guilbaud, R, Lyne, JW, Tostevin, R, Bowyer, F, Penny, AM,
Seilacher, A (1989) Vendozoa. Organismic construction in the Curtis, A & Kasemann, SA (2015): Dynamic redox conditions
Proterozoic biosphere. Lethaia, 22, 229-239, Senckenberg control late Ediacaran metazoan ecosystems in the Nama
Seilacher, A (1992): Vendobionta and Psammocorallia: Lost Group, Namibia. Precambrian Research, 261, 252-271
constructions of Precambrian evolution. Journal of the Xiao, S & Laflamme, M (2009): On the eve of animal radiation:
Geological Society, 149, 607-613 Phylogeny, ecology and evolution of the Ediacara biota.
Seilacher, A & Gishlick, AD (2014): Vendobionts: Lost life forms of Trends in Ecology & Evolution, 24, 31-40
Ediacaran times. In Morphodynamics, 9, 133-148, CRC Press Zhang, F, Xiao, S, Kendall, B, Romaniello, SJ, Cui, H, Meyer,
Sprigg, RG (1947): Early Cambrian jellyfishes from the Flinders M, Gilleaudeau, GJ, Kaufman, AJ, Anbar, AD (2018):
Ranges, South Australia. Transactions of the Royal Society of Extensive marine anoxia during the terminal Ediacaran
South Australia, 71(2), 212-224 Period. Science Advances, 4(6), eaan8983 https://doi.
Tostevin, R, Bradbury, HJ, Shields, GA, Wood, RA, Bowyer, F, Pen- org/10.1126/sciadv.aan8983
ny, AM, Turchyn, AV (2019a): Calcium isotopes as a record Zhuravlev, AY, Wood, RA & Penny, AM (2015): Ediacaran skeletal
of the marine calcium cycle versus carbonate diagenesis metazoan interpreted as a lophophorate. Proceedings of the
during the late Ediacaran. Chemical Geology, 529, 119319 Royal Society B (Biological Sciences), 282, 20151860
https://doi.org/10.1016/j.chemgeo.2019.119319
Tostevin, R, Clarkson, MO, Gangl, S, Shields, GA, Wood, RA,
Bowyer, F, Penny, AM, Stirling, CH (2019b): Uranium isotope
evidence for an expansion of anoxia in terminal Ediacaran
24 25future assist to better compare the measured fine dust concentration,
Feinstaub- und Radonkonzentrationen in Namibias and to identify potential exceedences.
Erongo Region
The measurements show that PM10 concentrations are higher at
the coast, compared with further inland. They are mostly derived
Detlof von Oertzen1 & Hanlie Liebenberg-Enslin2 from natural sources, in particular from the Namib Desert and the
1
VO Consulting, P.O. Box 8168, Swakopmund, Namibia, detlof@ Atlantic Ocean. The most important man-made sources of fine dust
voconsulting.net are surfaced and unsurfaced roads, and the regional salt roads. These
contribute on average some 50 000 tonnes of PM10 dust per annum,
2
Airshed Planning Professionals (Pty) Ltd, PO Box 5260, followed by mining and quarrying activities with some 22 000 tonnes.
Halfway House, Südafrika, hanlie@airshed.co.za Point sources of fine dust, such as incinerators and fires, were found to
contribute only small amounts to the total regional fine dust emissions.
Abstract The study also looked at internal radiation doses, caused by inhalation
In 2016, the Namibian Ministry of Mines and Energy (MME), with of radioactive fine dust and radon decay products. Inhalation of
financial assistance from the German Federal Institute for Geosciences such fine dust causes an inhalation dosis of some 0.003 milliSievert
and Natural Resources, embarked upon an update of the air quality per annum (mSv/a) for adults respectively 0.002 for toddlers. The
study in Namibia’s Erongo Region which had been undertaken in 2010. contribution of this inhalation dose to the total dose of individuals in
The update also included the development of preliminary regional air the population of the Erongo Region is therefore very small, and does
quality guidelines. not pose a risk for the public health.
A regional monitoring network was established to measure air quality, The contribution of the dose through inhalation of radon decay
and to quantify atmospheric dust concentrations (dust with a size of products is some 0.1 mSv/a in Walvis Bay, 0.2 mSv/a in Swakopmund
less than 10 µm (abbreviated as PM10), and fine dust of less than 2.5 µm and 0.4 mSv/a at the measuring station between Arandis and Rössing.
(PM2.5), radon concentrations and some meteorological parameters These doses are well below the worldwide average radiation doses
in real time. Data was generated between November 2016 and end from radon concentrations in air for individual members of the
of December 2018, and was subsequently evaluated. Some of the population, which is given by UNSCEAR as 1.095 mSV/a.
PM10 samples were also analysed for their mineral and radionuclide
composition. In addition, the sources of the regional fine dust were
identified with the aid of a dispersion model.
Preliminary guidelines for regional concentrations of fine dust were
developed on the basis of measured and modelled results. It is suggested
to use the PM2.5 annual average of 15 microgramm per cubic meter
(µg/m3) as proposed by the World Health Organisation (WHO) as
regional guideline. Furthermore, the South African National Ambient
Air Quality Standard (NAAQS) annual upper limit of 40 µg/m3 for
PM10 is recommended as regional guideline. These guidelines will in
26 27ZUSAMMENFASSUNG bei, gefolgt von Bergbau- und Steinbruchbetrieben, die ca. 22 000
Ende 2016 veranlasste das namibische Ministerium für Bergbau Tonnen PM10 pro Jahr beisteuern. Feinstaubpunktquellen, z.B.
und Energie (Ministry of Mines and Energy, MME), mit finanzieller Verbrennungsanlagen und Feuer, tragen nur wenig zu den regionalen
Unterstützung der deutschen Bundesanstalt für Geowissenschaften Feinstaubgesamtemissionen bei.
und Rohstoffe, die Aktualisierung einer Luftqualitätsuntersuchung in
Namibias Erongo Region aus dem Jahr 2010, und die Entwicklung Die Studie schätzt auch die inneren Strahlendosen, die durch das
erster regionaler Luftqualitätsrichtwerte. Einatmen von radioaktivem Feinstaub und Radonfolgeprodukten
entstehen, ab. Das Einatmen von Feinstaub verursacht eine Inhala
Zur Überwachung der Luftqualität wurde ein regionales Messnetzwerk tionsdosis von ca. 0.003 Millisievert pro Jahr (mSv/a) bei Erwachsenen
eingerichtet, um atmosphärische Staubkonzentrationen (Feinstaub mit bzw. 0.002 bei Kleinkindern. Der Beitrag dieser Inhalationsdosis zur
einem Durchmesser von weniger als 10 µm (abgekürzt PM10), und Gesamtdosis von Einzelpersonen der Bevölkerung in der Erongo
Feinstaub der kleiner als 2.5 µm (PM2.5) ist), Radonkonzentrationen Region ist sehr gering und stellt deshalb kein Risiko für die öffentliche
und einige meteorologische Parameter in Echtzeit zu quantifizieren. Gesundheit dar.
Die zwischen November 2016 und Ende Dezember 2018 erhobenen
Daten wurden anschließend ausgewertet und einige der PM10-Proben Der Dosisbeitrag durch Radonzerfallsprodukte ist ca. 0.1 mSv/a in
auf ihren Mineral- und Radionuklidgehalt analysiert. Außerdem
Walvis Bay, 0.2 mSv/a in Swakopmund und 0.4 mSv/a an der Station
wurden mit Hilfe eines Dispersionsmodells die Hauptquellen des
zwischen Arandis und Rössing. Diese Dosen sind weit geringer als die
regionalen Feinstaubes identifiziert.
weltweiten durchschnittlichen Strahlendosen für Einzelpersonen der
Bevölkerung, die auf Radonkonzentrationen in der Luft zurückzuführen
Erste Richtwerte für regionale Feinstaubkonzentrationen beruhen
sind, welche die UNSCEAR auf 1.095 mSv/a beziffert.
auf gemessenen und modellierten Ergebnissen. So soll das von der
Weltgesundheitsorganisation (World Health Organisation, WHO)
vorgeschlagene IT-3 Zwischenziel für den PM2.5 Jahresmittelwert, 15 EINLEITUNG
Mikrogramm pro Kubikmeter (µg/m3), der regionale Richtwert sein. Kommen in der Umgebungsluft der Erongo Region radioaktiver Fein-
Für PM10 Feinstaub soll der Jahresgrenzwert des Südafrikanischen staub und Radongas in Konzentrationen vor, die gesundheitliche Aus-
National Ambient Air Quality Standards (NAAQS), 40 µg/m3, der wirkungen auf die Einwohner der Region zur Folge haben können?
hiesige Richtwert sein. Die Richtwerte sollen helfen, gemessene Diese und ähnliche Fragen wurden in einer kürzlich fertiggestellten
Feinstaubkonzentrationen in Zukunft besser vergleichen zu können Studie des namibischen Bergbau- und Energieministeriums (Minis-
und mögliche Überschreitungen eindeutig zu identifizieren. try of Mines and Energy, MME) untersucht (Ministry of Mines and
Energy, 2019). Der vorliegende Bericht ist von den Hauptautoren der
Die Messungen zeigen, dass die PM10-Konzentrationen an den Studie für eine deutsche Leserschaft verfasst.
Küstenstationen höher als an den Inlandstationen sind. Diese stammen
hauptsächlich aus natürlichen Quellen, insbesondere aus der Namib Im Jahr 2010 entwickelte das MME den ersten Strategischen Um-
und dem Atlantik. Die wichtigsten vom Menschen verursachten weltmaßnahmenplan für die Erongo Region (Strategic Environmental
Feinstaubquellen sind befestigte, unbefestigte und die regionalen Management Plan, SEMP). Diese Untersuchung schloss, unter ande-
Salzstraßen. Pro Jahr tragen diese ca. 50 000 Tonnen PM10 Feinstaubs rem, eine Luftqualitätsstudie ein, die die in der Region auftretende
Luftverschmutzung erstmals quantifizierte, einschließlich der dort
28 29auftretenden Feinstaub- und Radonkonzentrationen. Die Ergebnisse Das Messnetzwerk zur Ermittlung der aktuellen Feinstaub- und Ra-
zeigten, dass die Winderosion von natürlich exponierten Gebieten, der donkonzentration wurde in oder nahe den bevölkerungsreichsten regi-
Straßenverkehr und die in der Region aktiven Bergbauunternehmen onalen Zentren eingerichtet, wie in Bild 1 dargestellt wird. Die damit
die Hauptbeitragenden zur lokalen atmosphärischen Feinstaubkon- erhobenen Daten dienten als Grundlage für ein Dispersionsmodell,
zentration sind. Außerdem wurde festgestellt, dass die Radonkonzen- zur Ermittlung und Identifizierung der wichtigsten regionalen Staub-
trationen, die in der Nähe von Walvis Bay, Swakopmund und Arandis quellen und schufen eine erste empirische Grundlage, auf der erste
gemessen wurden, stark variieren und von den vorherrschenden Win- regionale Luftqualitätsrichtlinien aufgebaut werden konnten.
den geprägt werden, aber im internationalen Vergleich gering sind.
DIE LUFTQUALITÄT DER ERONGO REGION
Ende 2016 veranlasste das MME, unterstützt von der deutschen Bun-
Die durch das Messnetzwerk erhobenen Luftqualitätsdaten sind wich-
desanstalt für Geowissenschaften und Rohstoffe, die Luftqualitäts-
untersuchungen aus dem Jahr 2010 zu aktualisieren, und einen Maß- tige Indikatoren der Zusammensetzung der Luft an den jeweiligen
nahmenplan für die regionale Luftqualität (Air Quality Management Messstellen. Mittels einer Dispersionssimulation wurden diese auf
Plan, AQMP) zu entwickeln. Diese Studie, die hier SEMP-2019 ge- die Umgebung der Messstationen übertragen, und damit konnten die
nannt wird und das Thema der vorliegenden Abhandlung ist, hat die wichtigsten Feinstaubquellen der Region identifiziert werden. Derarti-
atmosphärischen Feinstaub- und Radonkonzentrationen in der Erongo ge Modelle sind nicht exakt, da sie nur einen Teil der tatsächlich rele-
Region quantifiziert und erste regionale Luftqualitätsrichtwerte als vanten Parameter, die die lokale Luftqualität bestimmen, berücksich-
Teil des AQMP für die Region vorgeschlagen (Ministry of Mines and tigen. Dennoch liefern sie wichtige Informationen, wenn realitätsnahe
Energy, 2019). Staubemissionsraten und meteorologische wie auch topographische
Daten verwendet werden.
EIN LUFTQUALITÄTSMESSNETZWERK IN DER ERONGO
REGION Man unterscheidet zwischen natürlichen und vom Menschen verur-
Um Feinstaub- und Radonkonzentrationen zu quantifizieren, wurde sachten Luftverschmutzungsquellen. Der vom Wind transportierte
Ende 2016 ein Messnetzwerk in der Erongo Region errichtet. Die Staub, der von unberührten Landschaften herstammt, trägt maßgeb-
erfassten Messgrößen sind in Tabelle 1 zusammengefasst. Es wurden lich zur Luftverschmutzung in der Erongo Region bei. Ebenso ist der
die Luftkonzentrationen von inhalierbaren Partikeln mit einem Atlantik eine natürliche Quelle, die eine Verschiedenheit von organi-
aerodynamischen Durchmesser von weniger als 10 Mikrometer (μm), schen Stoffen und Meersalz liefert, die ihrerseits die lokale Luftqua-
als PM10 abgekürzt, und weniger als 2.5 μm (PM2.5), gemessen, und
lität beeinflussen. Zudem gibt es eine Vielzahl von Staubquellen die
auch die Radonkonzentration in der Luft, sowie einige meteorologische
von Menschen verursacht werden: wichtig sind a) die von Fahrzeugen
Parameter.
und stationären Motoren erzeugten Feinstäube and Abgase von Ver-
Parameter PM10 PM2.5 Wind-
geschwin-
Wind-
richtung
Tem-
peratur
Feuch-
tigkeit
Ein-
strahlung
Atmos-
phärischer
Regen-
fall
Radon-
konzen-
brennungsmotoren aller Art, b) der vom Straßenverkehr verursachte
Messstation
digkeit Druck tration
Abrieb, und c) Bergbau- und Steinbruchaktivitäten einschließlich dem
Swakopmund x x x x x x x x x x Abbau von Sand. Außerdem tragen Emissionen im Hafen in Walvis
Walvis Bay
Arandis
x
x
x x
x
x
x
x
x
x
x x
x
x x
x
x
Bay bei, sowie Bauaktivitäten und Verbrennungsanlagen, wie auch die
Henties Bay x x x x Vielzahl von großen und kleinen Feuern in der Region. Die SEMP-
Jakalswater x x x x x x 2019 Studie hat die wichtigsten aus dieser großen Zahl von Staubquel-
Tabelle 1: Erfasste Messgrößen pro Messstation (Ministry of Mines and Energy, 2019) len identifiziert und quantifiziert.
30 31Die Verteilung von Feinstaub- und Radonkonzentrationen, die in der
26-monatigen Messperiode zwischen dem 1. November 2016 und 31.
Dezember 2018 gemessen wurde, variiert stark. Hierzu ist vor allem
der ständig wechselnde regionale Wind verantwortlich. An der Küste
kommt der Wind dominant aus südwestlicher und auch nordwestlicher
Richtung, in zentralen Gebieten der Region ist die Windrichtung über-
wiegend aus Nordwest und Ost-Nordost. Die Windgeschwindigkeiten
nahe Arandis und Jakalswater sind meist niedriger als die in Walvis
Bay und Swakopmund.
Die SEMP-2019 Studie zeigt, dass der von Fahrzeugen verursachte
Straßenabrieb auf befestigten, unbefestigten und regionalen Salzstra-
ßen die wichtigste vom Menschen verursachte Feinstaubquelle ist.
Pro Jahr trägt diese ca. 50 000 Tonnen PM10 Feinstaub bei, gefolgt
von Bergbau- und Steinbruchbetrieben die ca. 22 000 Tonnen PM10
pro Jahr beisteuern. Feinstaubpunktquellen, z.B. Verbrennungsanla-
gen und Feuer, tragen nur wenig zu den regionalen Gesamtemissionen
bei. Die wichtigsten natürlichen Feinstaubquellen der Region sind der
vom Wind transportierte Staub aus der Namib und aus dem Atlantik.
In Messperioden außerhalb der Ostwindepisoden wurden bei den
Messtationen in Swakopmund und Walvis Bay zum Teil hohe bzw.
erhöhte PM10 Konzentrationen gemessen, vor allem wenn der Wind
aus südwestlicher und westlicher Richtung kam. Derartige Feinstäube
stammen direkt aus dem Atlantik, und die Studie konnte mittels che-
mischer und elektronenmikroskopischer Analysen zeigen, dass sie aus
Meersalz und organischen Stoffen bestehen.
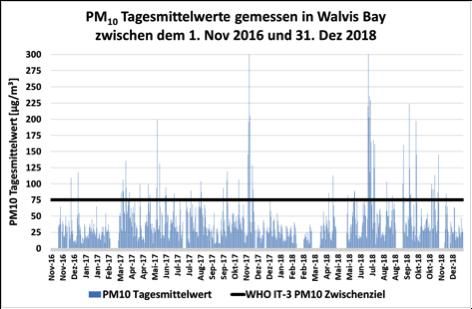
Abb. 2 (Abb. 3) zeigt die in Swakopmund (Walvis Bay) PM10 Tages-
mittelwerte und den von der Weltgesundheitsorganisation (WHO)
vorgeschlagenen IT-3 Luftqualitätsrichtlinienwert für PM10, um die
Luftkonzentrationen dieser Feinstaubsorte mit einem internationalen
Richtwert vergleichen zu können.
Mit einem Dispersionsmodell wurden die Hauptquellen des regionalen
Feinstaubes identifiziert. Das Modell beinhaltet ein Emissionsinventar
Abb. 1: Messstationen, Rezeptoren, Straßen, Bergwerke und Steinbrüche in der aus gemessenen und ermittelten Staubquellen, einschließlich a) dem
Erongo Region regionalen Straßennetzwerk, b) der Bergbaudeponien aller Art, den
32 33c) Bergbau- und Steinbruchbetrieben, und d) typischen Punktquellen
die in der Erongo Region Feinstäube emittieren.
Der Vergleich der gemessenen und simulierten Feinstaubkonzentra
tionen zeigt, dass die im Emissionsinventar enthaltenen Quellen nicht
komplett sind, weil die aus dem Hinterland und Atlantik stammenden
Emissionen unvollständig sind. Das erklärt, warum das Dispersions-
modell die meisten in der Region gemessenen atmosphärischen Fein-
staubkonzentrationen unterschätzt.
STRAHLENDOSEN FÜR EINZELPERSONEN DER
BEVÖLKERUNG
Die SEMP-2019 Studie präsentiert auch eine Abschätzung der durch
das Einatmen von radioaktivem Feinstaub und Radonfolgeprodukten
verursachten inneren Strahlendosis. Die Inhalationsexposition durch
Einatmen von Feinstaub in der Region ergibt bei Erwachsenen
Abb. 2: PM10 Tagesmittelwertkonzentrationen in Swakopmund, in μg/m3 (Kleinkindern) eine Strahlendosis von ca. 0.003 (0.002) Millisievert
pro Jahr (mSv/a), und ist damit etwas geringer als die weltweite
durchschnittliche Inhalationsdosis die vom United Nations Scientific
Committee on the Effects of Atomic Radiation (UNSCEAR) auf ca.
0.006 (0.005) mSv/a für Erwachsene (Säuglinge) geschätzt wird.
Die durchschnittlichen Dosisbeiträge, die vom Einatmen von
Radonzerfallsprodukten resultieren, belaufen sich auf 0.1 mSv/a
in Walvis Bay, 0.2 mSv/a in Swakopmund und 0.4 mSv/a an der
Messstation zwischen Arandis und Rössing. Diese Strahlendosen sind
weit geringer als die weltweiten durchschnittlichen Strahlendosen für
Einzelpersonen der Bevölkerung, die auf Radonkonzentrationen in
der Luft zurückzuführen sind, welche von der UNSCEAR auf 1.095
mSv/a beziffert wurden.
Die Ergebnisse der Dosisanalyse zeigen dass die Strahlendosis
durch das Einatmen von radioaktivem Staub in den wichtigsten
Bevölkerungszentren der Region Erongo kein Risiko für die öffentliche
Gesundheit darstellt und dass der Beitrag dieser Inhalationsdosis zur
Abb. 3: PM10 Tagesmittelwertkonzentrationen in Walvis Bay, in μg/m3
gesamten Strahlendosis für Einzelpersonen der Bevölkerung in der
Erongo Region sehr gering und daher unbedeutend ist.
34 35SCHLUSSFOLGERUNGEN regelmäßig und offiziell gemessen werden, um die Bevölkerung vor
Eingangs wurde gefragt ob der in der Luft der Erongo Region enthalte- möglichen Risiken zu schützen.
ne radioaktive Feinstaub und die Radonzerfallsprodukte maßgebliche
gesundheitliche Auswirkungen auf die Einwohner der Region haben DANKSAGUNG
können. Die SEMP-2019 Studie ist diesen Fragen nachgegangen, und Die Autoren danken dem MME für die freundliche Bereitstellung der
hat über einen Zeitraum von 26 Monaten die regionalen Feinstaub- Daten der SEMP-2019 Studie.
und Radonkonzentrationen gemessen und ausgewertet.
LITERATUR
Die Studie hat die Strahlendosen für Einzelpersonen der Bevölke- Ministry of Mines and Energy (2019): Advanced Air Quality
rung, die durch das Einatmen von radioaktivem Staub und Radon- Management for the Strategic Environmental Management Plan for
zerfallsprodukten entstehen, durch Echtzeitmessungen ermittelt. Da the Uranium and other Industries in the Erongo Region: Air Quality
die Messstationen nahe den wichtigsten Bevölkerungszentren in der Management Plan Report, Report 15MME01-4, Ministry of Mines
Erongo Region positioniert waren, sind die Ergebnisse der Studie and Energy, Windhoek, www.mme.gov.na
nicht auf die unmittelbare Umgebung von Uranminen zu beziehen.
Es konnte gezeigt werden, dass die bevölkerungsgewichtete Strahlen
dosis für Erwachsene (Kleinkinder) aufgrund des eingeatmeten Neueingänge in der Sam-Cohen-Bibliothek
radioaktiven Feinstaubs in der Außenluft der Region unter New books in the Sam Cohen Library
0.003 mSv/a (0.002 mSv/a) liegt. Derartige Strahlendosen sind als
sehr gering einzustufen. Auf empirischer Grundlage wurde ermittelt, Ahrens, Christian: Für Kaiser und Reich: Kriegstage in China und
dass die regionale bevölkerungsgewichtete Strahlendosis, die durch Südwestafrika. Windhoek, 2009
das Einatmen von Radonzerfallsprodukten in der Außenluft entsteht, Aiken, Bruce: The lions and elephants of the Chobe. Sandton, n.d.
kleiner als 0.2 mSv/a ist. Allofs, Theo & Sabine: Namibia. Hamm, 1996
Ammann, Kathrine & Karl: Geparden. Hannover, 1987
Die Studie schlägt auch erste Richtwerte für regionale Ammann, Karl & Kathrine: Jäger und Gejagte. Halfway House, 1992
Feinstaubkonzentrationen vor. So soll das von der Welt-Gesundsheits- Bannister, Anthony; Skinner, John: South African Animals in the
Organisation (WHO) vorgeschlagene IT-3 Zwischenziel für den Wild. Cape Town, 1987
PM2.5 Jahresmittelwert, also 15 µg/m3, der regionale Richtwert sein, Benjamin, Alfeus: Die nuwe skoolhoof. Tsumeb, 2018
und der Südafrikanische National Ambient Air Quality Standards Blüchel, Kurt; Medenbach, Olaf: Zauber der Mineralien: Kristalle,
(NAAQS) Jahresgrenzwert von 40 µg/m3 als hiesiger Richtwert für Gold und Edelsteine. München, 1981
PM10 Feinstaub dienen. Diese Richtwerte sollen helfen, gemessene Blumhagen, Hans Ernst: Die Doppelstaatigkeit der Deutschen
Feinstaubkonzentrationen besser vergleichen zu können, und mögliche im Mandatsgebiet Südwestafrika und ihre völkerrechtlichen
Überschreitungen eindeutig zu identifizieren. Auswirkungen. Berlin, 1938
Bollig, Michael; Klees, Frank: Überlebensstrategien in Afrika. Köln,
Die SEMP-2019 Studie schlägt auch vor, dass Feinstaubkonzentra- 1994
tionen in unmittelbarer Nähe zu den in der Region aktiven Uranmi- Bothma, J. du P. ed.: Game Ranch Management. Pretoria, 1989
nen, und Radonkonzentrationen in potenziell gefährdeten Gebäuden, Bothma, J. du P. ed.: Game Ranch Management. Pretoria, 1990
36 37Braam, Conny: Ek is Hendrik Witbooi. Pretoria, 2019 Fedonkin, Mikhail A. et al.: The rise of animals; evolution and
Bridgland, Fred: Jonas Savimbi: A key to Africa; South Africa’s ally diversification of the Kingdom Animalia. Baltimore, 2007
and Mao’s disciple. London, 1968 Frobenius, Leo; Pager, H.L.: Summary & Anhang: Madsimu
Butchart, Duncan: Wild about the Okavango. All-in-one guide to Dsangara; Südafrikanische Felsbilderchronik; Band 1 - Der
common animals and plants of the Okavango Delta, Chobe National archäologische Keilstil. Graz, 1962
Park and Caprivi Strip. Halfway House, 1999 Getta, Jelizaveta: Interpreting in Namibia: past and present. Overview
Conradie, Franz: Stargazing for the novice. Thabazimbi, 1996 from colonial times to the present day. Prague, 2019
Croan, Melvin: DDR - Neokolonialismus in Afrika. DAS Gisevius, Hans Bernd: Bis zum Bittern Ende: 1. Band: Vom
Schriftenreihe Heft 8. Bonn, 1981
Reichstagsbrand zur Fritsch-Krise. Zürich, 1946
Cubitt, Gerald; Robins, Eric: The Book of Kenya. London, 1980
Goss, Richard; Maberly, A.: Maberly’s Mammals of Southern Africa
Cubitt, Gerald; Owen-Smith, Garth: Namibia, Land der wilden
Schönheit. Cape Town, 1981 - a popular field guide. Craighall, 1986
Cubitt, Gerald; Joyce, Peter: Dies ist Namibia. Cape Town, 1992 Grobler, Hans et al.: Predators of Southern Africa. A guide to the
Lewis-Williams, J. David: The Rock Art of Southern Africa; the Carnivores. Johannesburg, 1984
imprint of Man. Cambridge, 1983 Gudemann, Wolf Eckhard, ed.: Flaggen und Wappen der Welt. Mit
Degré, Alain et al.: Meerkat Valley. Johannesburg, 1898 einem Schlüssel zur Flaggenbestimmung. Ausführliche Beschreibung
de Klerk, W. A.: Drie Swerwers oor die Einders. Kaapstad, 1962 der Geschichte und Symbolik von über 900 Flaggen und Wappen aller
Denker, Kai-Uwe; Denker, Helge: Jagen in Namibia. Windhoek, Staaten. Gütersloh, 1992
1994 Hagen, Walter: Die Geheime Front. Organisation, Personen und
Denker, Kai-Uwe: Entlang des Jägers Pfad. Gedanken und Erlebnisse Aktionen des Deutschen Geheimdienstes. Wien, 1950
eines Berufsjägers. 2000 Halenke, H.: Deutsche, Südafrikanische und Namibische
Denker, Kai-Uwe: Vom Wesen der afrikanischen Wildnis. Auf der Staatsangehörigkeit in Vergangenheit, Gegenwart und Zukunft; die
Fährte alter Elefantenbullen. Windhoek, 2014 rechtlichen Grundlagen in Stichworten. Windhoek, 1989
Dennis, Nigel; Macdonald, David: Meerkats. London, 1999 Hallier, Ulrich W.: Die Entwicklung der Felsbildkunst Nordafrikas.
Deutsche Schule Johannesburg: Festschrift der Deutschen Schule Stuttgart, 1990
zu Johannesburg zur 100 - Jahrfeier. Johannesburg, 1990 Hall-Martin, Anthony: Ein Tag im Leben eines Afrikanischen
Dowson, Thomas A.: Rock Engravings of Southern Africa.
Elefanten. Johannesburg, 1993
Johannesburg, 1992
Hansohm, Dirk et al., ed.: Monitoring Regional Integration in
Drechsler, Horst: Südwestafrika unter Deutscher Kolonialherrschaft.
Southern Africa - Yearbook, Volume 1. Windhoek, 2002
Der Kampf der Herero und Nama gegen den deutschen Imperialismus
(1884 - 1915). Berlin, 1984 Hes, Lex: The leopards of Londolozi Cape Town, 1991
Drechsler, Horst: Südwestafrika unter deutscher Kolonialherrschaft; Hilpisch, Hans: Ludwig von Estorff - Preußisch - eigenwillig -
die großen Land- und Minengesellschaften (1885 - 1914). Stuttgart, geachtet. Windhoek, 2019
1996 Hilpisch, Hans: Wo sind die Herero geblieben? Neue Erkenntnisse
du Plessis, Jéan: Namibia - a visual celebration. Cape Town, 2007 und Theorien zum Rückzug der Herero in die Omaheke 1904/1905.
Erkkilä, Antti; Siiskonen, Harri: Forestry in Namibia 1850 - 1990. Windhoek, 2019
Silva Carelica 20. Joensuu, Finland, 1992 Hoberman, Gerald & Marc: Namibia. Cape Town, 1999
38 39Hockey, Phil: The African penguin - a natural history. Cape Town, Maclean, Gordon Lindsay: Roberts’ Birds of Southern Africa; sixth
2001 edition. Cape Town, 1993
Holm, Erik; de Meillon, Elbie: Insects. Struik pocket guides for Malherbe, Candy; Bailey, Seton: These Small People. Hunter
Southern Africa. Cape Town, 1986 Gatherers. Pietermaritzburg, 1983
Holm, Erik sen.: Bushman Art. Pretoria, 1987 Marais, Christine: Swakopmund gestern und heute. Windhoek, 1992
Jackson, Helen: AIDS Africa: Continent in Crisis. Harare, 2002 Marais, Christine: Old Swakopmund today. Windhoek, 1992
Jouve, Lily & Marcel: Secret Namibia. Cape Town, 2009 Marc, Alexandre; ed.: African Art: The World Bank Collection.
Joyce, Peter: from Aardvark to Zebra; secrets of African Wildlife. Washington, 1998
Cape Town, 1999 Marker, L. et al.: Die oorlewing van jagluiperds op Namibiese plase.
Kandetu, Bob: Timeless Bonds: an appraisal of Namibia’s Windhoek, 1999
development through time. Windhoek, 2017 Masche, Bernd: Namibia - unser Land - unser Bier. Windhoek, 2019
Kauffman, Kyle D.; Lidnauer, David L. ed.: AIDS and South Africa. Mayer, Wolfgang et al.: Schwarz-Weiss-Rot in Afrika: die deutschen
The Social Expression of a Pandemic. New York, 2004 Kolonien 1883 - 1918. Puchheim, 1985
Knüsel, Beat et al.: Namibia - Jenseits des Ozeans. München, 1997 McCulloch, Elzanne, ed.: Large-scale black rhino conservation in
Koornhof, Anton: The Dive sites of South Africa. A concise illustrated north-western Namibia. Review of Progress 2012 - 2018. Windhoek,
guide to 170 coastal and inland dive destinations. Cape Town, 1991 2019
Kopij, Grzegorz: Namibian Wildlife Bibliography. Katima Mulilo, McGregor, Gordon: The History of Freemasonry in Namibia 1905 -
2014 2019. Windhoek, 2019
Künkel, Reinhard: Elephants. New York, 1989 McGregor, Gordon: South West Africa Wheel Tax Badges 1926 -
Lambrechts, Hugo A.: Namibia: Durstland und Wildnis. Cape Town, 1961. Windhoek, 2017
1985 McGregor, Gordon: The Dog Tax Badges of German South West
LEAD Project: A Place we want to call our own; a study on land Africa, South West Africa and Namibia 1898 – 2016. Swakopmund,
tenure policy and securing housing rights in Namibia. Windhoek, 2005 2016
Leipner, Nils: Marken und Token der ehemaligen deutschen Kolonien. McIntyre, Joan, ed.: Der Geist in den Wassern. Ein Buch zu Ehren
2016 des Bewußtseins der Wale und Delphine. Frankfurt, 1982
Lenoir, Philippe: Mauritius. Verona, 1985 McKenzie, Andrew A., ed.: The Capture and Care Manual. Capture,
le Roux, Annelise et al.: Namaqualand. South African Wildflower Care, Accommodation and Transportation of Wild African Animals.
Guide 1; revised edition. Kirstenbosch, 1988 Lynnwood Ridge, 1993
Lewis-Williams, David; Dowson, Thomas: Images of Power - Mead, Colin: Shadows of Sand. A photodocument of the Namib
Understanding Bushman Rock Art. Cape Town, 1989 Desert Dunes. Bryanston, 1988
Liebenberg, Louis: The Art of Tracking - the origin of science. Mead, Colin: Wilderness Heritage. Bryanston, nd
Claremont, 1990 Medenbach, Olaf; Wilk, Harry: Zauberwelt der Mineralien. Salzburg,
Loutit, Blythe: Der wundersame Elefant in der Namib. Windhoek, 1977
2001 Meinhof, Carl; Herausgeber: Afrikanische Märchen. Reinbek, 1993
Macintosh, E.K.: A guide to the rocks, minerals and gemstones of Miller, Jamie: An African Volk - the Apartheid regime and its search
Southern Africa. Cape Town, 1976 for survival. Oxford, 2016
40 41You can also read

















































