Molecular Medicine YAP Controls Endothelial Activation and Vascular Inflammation Through TRAF6 - Chicago
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Molecular Medicine
YAP Controls Endothelial Activation and Vascular
Inflammation Through TRAF6
Yang Lv, Kyungho Kim, Yue Sheng, Jaehyung Cho, Zhijian Qian, You-Yang Zhao, Gang Hu,
Duojia Pan, Asrar B. Malik, Guochang Hu
Rationale: Microvascular inflammation and endothelial dysfunction secondary to unchecked activation of
endothelium play a critical role in the pathophysiology of sepsis and organ failure. The intrinsic signaling
mechanisms responsible for dampening excessive activation of endothelial cells are not completely understood.
Objective: To determine the central role of YAP (Yes-associated protein), the major transcriptional coactivator of the
Hippo pathway, in modulating the strength and magnitude of endothelial activation and vascular inflammation.
Methods and Results: Endothelial-specific YAP knockout mice showed increased basal expression of E-selectin
and ICAM (intercellular adhesion molecule)-1 in endothelial cells, a greater number of adherent neutrophils
in postcapillary venules and increased neutrophil counts in bronchoalveolar lavage fluid. Lipopolysaccharide
challenge of these mice augmented NF-κB (nuclear factor-κB) activation, expression of endothelial adhesion
proteins, neutrophil and monocyte adhesion to cremaster muscle venules, transendothelial neutrophil migration,
and lung inflammatory injury. Deletion of YAP in endothelial cells also markedly augmented the inflammatory
response and cardiovascular dysfunction in a polymicrobial sepsis model induced by cecal ligation and puncture.
YAP functioned by interacting with the E3 ubiquitin-protein ligase TLR (Toll-like receptor) signaling adaptor
TRAF6 (tumor necrosis factor receptor-associated factor 6) to ubiquitinate TRAF6, and thus promoted TRAF6
degradation and modification resulting in inhibition of NF-κB activation. TRAF6 depletion in endothelial cells
rescued the augmented inflammatory phenotype in mice with endothelial cell–specific deletion of YAP.
Conclusions: YAP modulates the activation of endothelial cells and suppresses vascular inflammation through
preventing TRAF6-mediated NF-κB activation and is hence essential for limiting the severity of sepsis-induced
inflammation and organ failure. (Circ Res. 2018;123:43-56. DOI: 10.1161/CIRCRESAHA.118.313143.)
Downloaded from http://ahajournals.org by on July 21, 2021
Key Words: acute lung injury ◼ E-selectin ◼ endothelial cells ◼ neutrophils ◼ sepsis ◼ ubiquitination
S epsis is the leading cause of intensive care unit admissions,
with high mortality and morbidity. In septic patients, wide-
spread vascular leakage and inflammation secondary to endo-
ultimately organ dysfunction.3 Therefore, maintenance or re-
constitution of vascular endothelial homeostasis after a major
systemic injury is a crucial determinant of outcome in sepsis.1,4
thelial activation and dysfunction is the primary mechanism of However, the underlying mechanisms limiting the severity of
multiorgan failure.1 Endothelial cells are the key sentinel cells endothelial activation during sepsis remain to be identified.
detecting microbial infection through the activation of pattern
Meet the First Author, see p 3
recognition receptors to mount an inflammatory response.
Endothelial activation during sepsis leads to the production of Activation of NF-κB (nuclear factor-κB) is critical for
multiple proinflammatory cytokines and upregulation of adhe- regulating the expression of inflammatory genes and adhesion
sion molecules E/P-selectins and ICAM (intercellular adhe- molecules.5 The NF-κB activation cascade is initiated by endo-
sion molecule)-1 essential for polymorphonuclear neutrophil toxin lipopolysaccharide binding to TLR (Toll-like receptor)-4,
(PMN) rolling, adhesion, and transendothelial PMN migra- which activates adaptor protein MyD88 (myeloid differen-
tion.2 Sustained activation of endothelium results in massive tiation factor)-dependent and MyD88-independent signaling
infiltration of PMNs, hyperinflammation, tissue damage, and pathways.6,7 MyD88 associates with IRAK (IL [interleukin]-1
Original received April 14, 2018; revision received May 16, 2018; accepted May 21, 2018. In April 2018, the average time from submission to first
decision for all original research papers submitted to Circulation Research was 13.36 days.
From the Department of Anesthesiology (Y.L., Guochang Hu) and Department of Pharmacology (K.K., J.C., A.B.M., Guochang Hu), University
of Illinois College of Medicine, Chicago; Department of Pharmacology, Nanjing Medical University, Jiangsu, China (Y.L., Gang Hu); Department of
Medicine and Cancer Research Center, The University of Illinois Hospital and Health Sciences System, Chicago (Y.S., Z.Q.); Program for Lung and
Vascular Biology, Stanley Manne Children’s Research Institute, Ann & Robert H. Lurie Children’s Hospital of Chicago, IL (Y.-Y.Z.); Departments of
Pediatrics and Pharmacology, Northwestern University Feinberg School of Medicine, Chicago, IL (Y.-Y.Z.); and Howard Hughes Medical Institute and
Department of Physiology, UT Southwestern Medical Center, Dallas, TX (D.P.).
The online-only Data Supplement is available with this article at http://circres.ahajournals.org/lookup/suppl/doi:10.1161/CIRCRESAHA.
118.313143/-/DC1.
Correspondence to Guochang Hu, MD, PhD, Department of Anesthesiology, University of Illinois College of Medicine, 3200W UICH, MC 515, 1740
W Taylor St, Chicago, IL 60612-7239. E-mail gchu@uic.edu
© 2018 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.118.313143
43
44 Circulation Research June 22, 2018
Novelty and Significance
What Is Known? Systemic endothelial activation and dysfunction induce vascu-
lar inflammation and disruption of microvascular barrier integ-
• Microvascular endothelial activation and dysfunction are critical deter-
minants in the progression from sepsis to organ failure.
rity leading to microvascular leak, tissue edema, shock, organ
• YAP (Yes-associated protein) transcriptionally promotes cell survival, failure, and ultimate death. However, the molecular mechanisms
chemotaxis, differentiation, and proliferation. responsible for controlling endothelial activation remains un-
known. Herein, we identified a crucial role of cytoplasmic YAP
What New Information Does This Article Contribute? as a checkpoint modulator of endothelial activation and vascular
inflammation. YAP suppressed inflammatory signaling by inter-
• Endothelial-specific deletion of YAP exaggerates sepsis-induced acting with TRAF6 and prevented the activation of NF-κB by en-
vascular inflammation, lung inflammatory injury, and cardiovascular hancing degradation of TRAF6 via K48 ubiquitination as well as
dysfunction. suppressing K63-linked ubiquitination. Our results suggest that
• YAP limits NF-κB (nuclear factor-κB) activation via modification of the
endothelial YAP may be a novel therapeutic target for prevention
signaling adaptor TRAF6 (tumor necrosis factor receptor-associated
and treatment of sepsis-induced tissue inflammatory injury and
factor 6).
organ failure.
TRAF6 leads to its degradation via the 26S proteasome, where-
Nonstandard Abbreviations and Acronyms
as K63-linked polyubiquitination is essential for activation of
CKO conditional knockout TAK1 kinase and subsequent NF-κB signaling.13,14
CLP cecal ligation and puncture The Hippo signaling pathway consists of a serine kinase
HA hemagglutinin cascade with associated regulatory and scaffolding proteins
HEK human embryonic kidney that transcriptionally regulate the expression of genes control-
HLMVECs human lung microvascular endothelial cells ling growth. The Ser/Thr kinase Hippo in Drosophila (Mst1
ICAM intercellular adhesion molecule [mammalian sterile 20-like kinase] and Mst2 in mammals)
IκB inhibitor protein κB
and Sav1 (salvador) form a complex that phosphorylates and
IKK IκB kinase
activates LATS1/2 (large tumor suppressor) kinases, which
in turn phosphorylate YAP (Yes-associated protein) and TAZ
IL interleukin
Downloaded from http://ahajournals.org by on July 21, 2021
(transcriptional coactivator with PDZ-binding motif), the 2
IRAK IL-1 receptor-associated kinase
downstream effectors of the Hippo pathway, causing them to
MyD88 myeloid differentiation factor
be retained in the cytoplasm.15–17 Dephosphorylation of YAP
NF-κB nuclear factor-κB
and TAZ induces their nuclear translocation and activates ex-
PMN polymorphonuclear neutrophil
pression of genes that promote cell proliferation and inhibit
sICAM soluble ICAM
apoptosis.18,19 YAP is a potent oncogene first identified by its
siRNA small interfering RNA ability to associate with Yes and Src tyrosine kinases.20 As a
sVCAM soluble vascular cell adhesion molecule transcriptional coactivator, YAP activates the expression of
TAK transforming growth factor-β–activated kinase target genes important for cell survival, chemotaxis, differen-
TAZ transcriptional coactivator with PDZ-binding motif tiation, and proliferation.21,22 Activation of YAP or loss of its
TLR Toll-like receptor upstream negative regulators leads to striking overgrowth and
TNF tumor necrosis factor tumor phenotypes.23 Although Hippo/YAP signaling is impor-
TRAF6 TNF receptor-associated factor 6 tant in oncogenesis, little is known about how YAP signals
WT wild-type inflammation. We previously showed that Gram-positive bac-
YAP Yes-associated protein teria activated Hippo signaling through Toll-dependent path-
way in Drosophila.24 It is unknown if Hippo/YAP signaling
also regulates inflammatory response induced by sepsis.
receptor-associated kinase), which recruits E3 ligase TRAF6 We identified here a previously unrecognized YAP-
(tumor necrosis factor [TNF] receptor-associated factor 6),6,8–10 dependent pathway regulating NF-κB activation in endothe-
which in turn interacts with the TAK1 (transforming growth lial cells via a direct interaction of YAP with TRAF6. YAP
factor-β–activated kinase 1) complex. TAK1 activates IKK induced the degradation of TRAF6 via K48 ubiquitination
(IκB [inhibitor protein κB] kinase), leading to the phosphoryla- while simultaneously suppressing K63-linked autoubiquitina-
tion and degradation of IκB, and consequent release of NF-κB tion to prevent downstream formation and activation of the
and its nuclear translocation.11,12 TRAF6 lies downstream of TAK1 complex and NF-κB signaling. We found that deletion
TNF receptor, IL-1 family and most TLR (TLR1-9) and func- of YAP expression in endothelial cells triggered inflammation
tions as a node controlling the activation of NF-κB. The ubiqui- even in the absence of infection and caused hyperinflamma-
tination of TRAF6 is a key regulatory event controlling the level tion in response to lipopolysaccharide or cecal ligation and
of TRAF6 expression and TAK1 activation.13,14 TRAF6 can be puncture (CLP) challenge through unfettered activation of
ubiquitinated through the E3 ubiquitin ligase in its RING (really NF-κB signaling. These results establish an important func-
interesting new gene) domain.14 K48-linked ubiquitination of tional linkage between endothelial YAP and inhibition of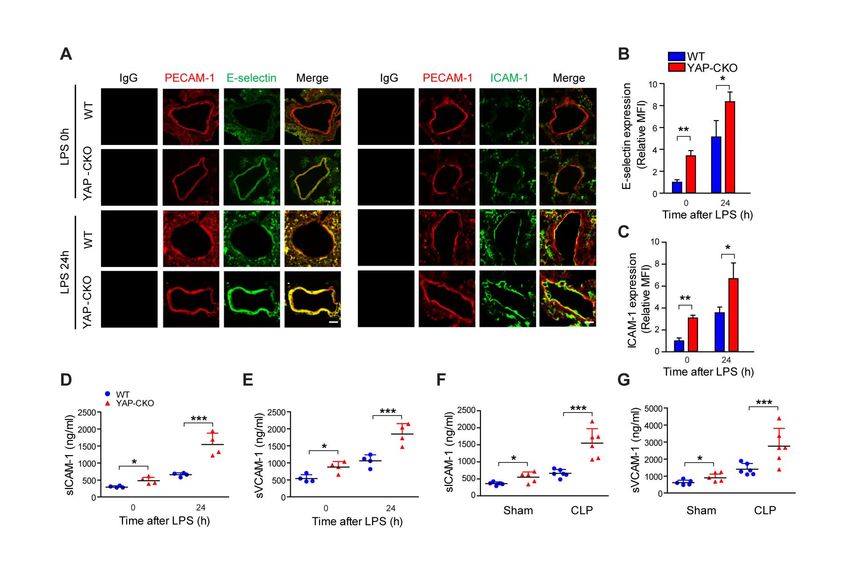
Lv et al YAP Limits Endothelial Activation 45
NF-κB signaling through direct interaction of cytoplasmic The proportion of born WT (YAP+/+), heterozygous (Tie2-Cre;
YAP with TRAF6 and point to a fundamental role of YAP in YAPf/+), and homozygous (Tie2-Cre; YAPf/f) mice complied
limiting activation of endothelial cells. with Mendel’s laws, and their appearance was undistinguish-
able from the littermates. Similarly, the male to female ratio
Methods was also consistent with the law. No difference was observed
The data that support the findings of this study are available from the with regard to body weight (Online Figure IIA) or organ size
corresponding author on reasonable request. (Online Figure IIB and IIC) in YAP-CKO mice. The ratios of
Details of materials and experimental procedures are available in
the Online Data Supplement.
organ to body weight were also not different in YAP-CKO mice
except for the slightly higher ratio in spleen/body weight as
Mice compared with WT mice (Online Figure IIC). YAP-CKO mice
Endothelial-specific YAP-deficient mice were generated by crossing also showed normal fertility and the oldest mice survived for >2
YAP-floxed mice (YAPf/f) with Tie2-Cre transgenic mice. All mice years. Importantly, YAP-CKO mice displayed mild lung inflam-
used in the experiments were the littermates of genotypes of YAPf/f
mation and endothelial activation as evidenced by increased
with Cre gene as endothelial YAP-conditional knockout (CKO) mice
(these mice are hereafter referred to as YAP-CKO). Mice only carry numbers of total cells, PMNs, macrophages, and PMN/mac-
YAPf/f as wild-type (WT) mice. rophage ratio in bronchoalveolar lavage fluid (Online Figures
IID, IIE, and IIIA through IIIC) and increased expression of
Mouse Models of Endotoxemia and Polymicrobial E-selectin and ICAM-1 in lung endothelial cells (Online Figure
Sepsis IVA through IVC) as compared with WT mice.
All experimental mice were sex- and age-matched. Endotoxemia
was induced with injection of lipopolysaccharide (5 mg/kg, IP). Enhanced Inflammatory Response to Endotoxin
Polymicrobial sepsis was created by CLP. and Polymicrobial Sepsis in YAP-CKO Mice
Intravital Microscopy We next examined whether baseline increases in endothelial
The cremaster muscle was exposed. PMNs and monocytes were vi- activation enhanced the susceptibility of YAP-CKO mice to
sualized by infusion of Alexa Fluor 647-conjugated anti-Ly-6G and endotoxemia. In YAP-CKO mice challenged with lipopolysac-
Alexa Fluor 647-conjugated anti-F4/80 antibody, respectively. The charide, the numbers of total cells (Online Figure IIIA), PMNs
rolling influx and number of adherent PMNs or monocytes were (Figure 1A), macrophages (Online Figure IIIB), and the ratio
determined.
of PMN/macrophages (Online Figure IIIC) in bronchoalveolar
Generation of Bone Marrow Chimeras lavage fluid as well as expressions of E-selectin and ICAM-
The irradiated mice (CD45.2) were transplanted with bone marrow 1 in pulmonary endothelial cells (Online Figure IVA through
IVC) dramatically increased compared with their WT coun-
Downloaded from http://ahajournals.org by on July 21, 2021
cells isolated from Ly5.1 (CD45.1). Six weeks after transplantation,
all the recipients were used for measurement of lung inflammation. terparts. The serum sICAM (soluble ICAM)-1 (Online Figure
IVD) and sVCAM (soluble vascular cell adhesion molecule)-1
Depletion of TRAF6 in Mouse Pulmonary
(Online Figure IVE) released by activated endothelium were
Endothelium
TRAF6 was depleted in mouse pulmonary vasculature by intrave- also elevated both basally and after lipopolysaccharide chal-
nous injection of cationic liposome-TRAF6 small interfering RNA lenge. Lung tissue from WT counterparts after lipopolysac-
(siRNA) complexes. charide challenge had severe histological changes, including
alveolar congestion, exudates, and infiltration of inflammatory
Statistics cells, whereas these alterations were markedly exacerbated in
One-way ANOVA and Student Newman-Keuls test for post hoc com-
parisons were used to test differences between the means of 3 or more lipopolysaccharide-challenged YAP-CKO mice, as evident by
groups. Mann-Whitney U test was performed to detect significant dif- the significantly higher lung injury score (Figure 1B). To as-
ferences between 2 experimental groups. Difference in the survival sess whether endothelial YAP knockout also augmented the
between 2 groups was determined by Mantel-Cox test. All data are increase in lipopolysaccharide-induced pulmonary vascular
expressed as mean±SD. Differences were considered significant permeability, we measured lung albumin-bound Evans blue
when P85% re- CLP-induced increase in serum sICAM-1 (Online Figure
duction in YAP mRNA (Online Figure IC) and protein levels IVF), sVCAM-1 (Online Figure IVG), IL-6 (Figure 1F),
in isolated mouse lung endothelial cells but not in nonendo- and TNF-α (Figure 1G). In addition, mean blood pressure
thelial cells (epithelial cells and fibroblasts; Online Figure ID). (Figure 1H), ejection fraction (Figure 1I and 1J), and frac-
Immunoblotting assessment of YAP protein expression also ex- tional shortening (Figure 1I and 1K) dramatically decreased
hibited 60% reduction in whole lung tissues (Online Figure IE). in YAP-CKO mice after CLP challenge compared with WT
46 Circulation Research June 22, 2018
Figure 1. YAP (Yes-associated protein) deletion in endothelial cells increases lung inflammatory injury, cardiovascular organ
failure, and mortality in mice challenged by lipopolysaccharide (LPS) or cecal ligation and puncture (CLP). Wild-type (WT) and
Downloaded from http://ahajournals.org by on July 21, 2021
YAP-conditional knockout (CKO) littermates were challenged with LPS (5 mg/kg, IP, A–D) or CLP (F–K). Bronchoalveolar lavage (BAL)
was performed at 0, 6, and 24 h after LPS injection. A, Polymorphonuclear neutrophil (PMN) count in BAL fluid by cytospin analysis.
n=6. **P
Lv et al YAP Limits Endothelial Activation 47
Downloaded from http://ahajournals.org by on July 21, 2021
Figure 2. Failure of wild-type (WT) bone marrow cells to rescue increased inflammatory response after lipopolysaccharide
(LPS) stimulation in YAP (Yes-associated protein)-conditional knockout (CKO) mice. A, An experimental protocol showing bone
marrow transplantation to lethally irradiated WT and YAP-CKO mice. B and C, FACS (fluorescence-activated cell sorting) analysis of
the percentage of the CD45.1 marker-expressing in the reconstituted WT and YAP-CKO mice. ***P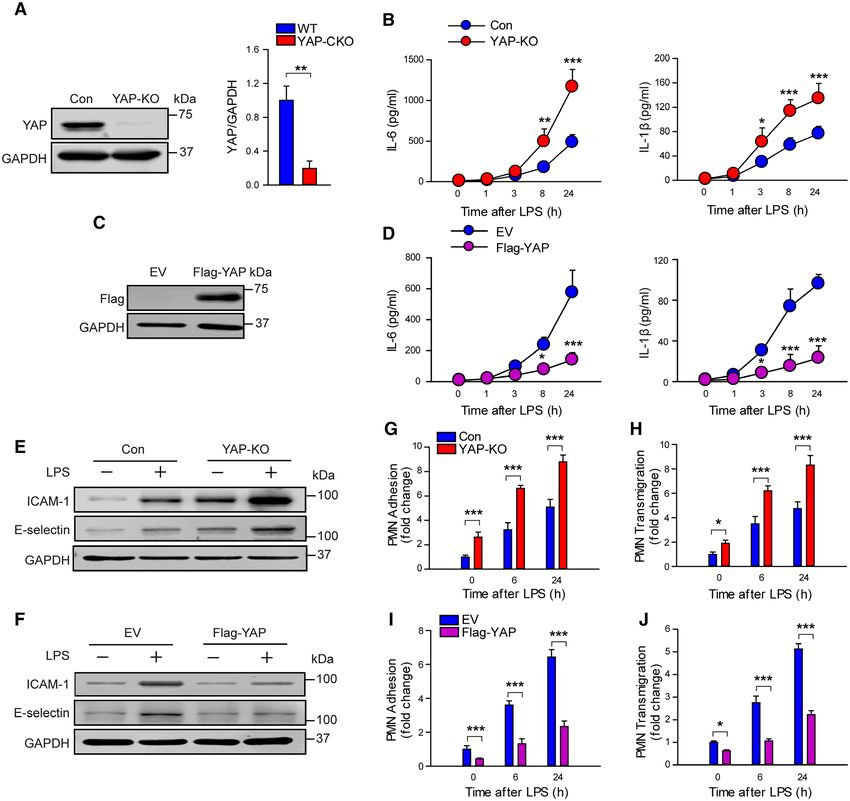
48 Circulation Research June 22, 2018
Downloaded from http://ahajournals.org by on July 21, 2021
Figure 3. Endothelial YAP (Yes-associated protein) deletion increases the number of adherent polymorphonuclear neutrophils
(PMNs) in vivo. A, Representative images of cremaster muscle venule demonstrating the role of YAP in regulating PMN rolling and
recruitment. PMN accumulation was visualized by infusion of Alexa Fluor 647-anti-Ly6G into wild-type (WT) or YAP-conditional knockout
(CKO) mice challenged with lipopolysaccharide (LPS) for 0 and 24 h. Scale bar: 10 μm. Arrows show direction of blood flow and yellow
dashed lines indicate the border of blood vessel. B and C, Numbers of adherent (B) and rolling (C) PMNs are shown. n=28 to 30 venules
in 3 to 4 mice per group. D, Representative images of E-selectin and ICAM (intercellular adhesion molecule)-1 staining. Cremaster
muscles from mice treated with LPS for 0 and 24 h were stained with PECAM-1 (platelet endothelial cell adhesion molecule-1) and
E-selectin or ICAM-1 antibodies. Scale bar: 10 μm. E and F, Quantitative data showing changes in mean fluorescent intensity (MFI) of
E-selectin (E) and ICAM-1 (F) expression. n=4 to 6. *P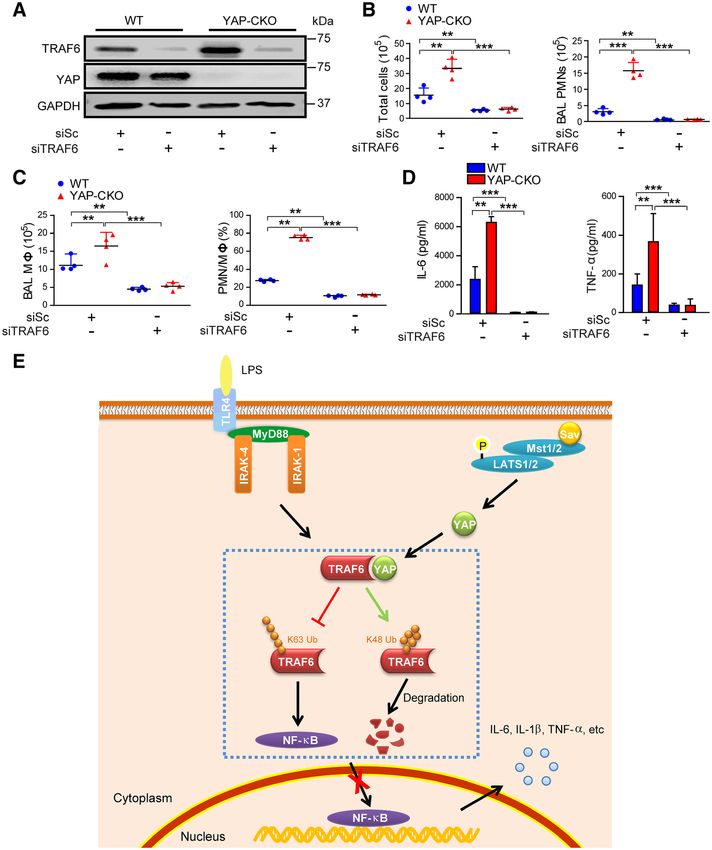
Lv et al YAP Limits Endothelial Activation 49
Downloaded from http://ahajournals.org by on July 21, 2021
Figure 4. Lipopolysaccharide (LPS) induces YAP (Yes-associated protein) expression in human endothelial cells and mouse lungs.
A, Immunoblotting showing expression of YAP in the HUVECs (human umbilical vein endothelial cells) after LPS stimulation (1 µg/mL) at
indicated times. Left, Representative Western blots for YAP and GAPDH (loading control); right, protein quantification by densitometry. n=3.
**P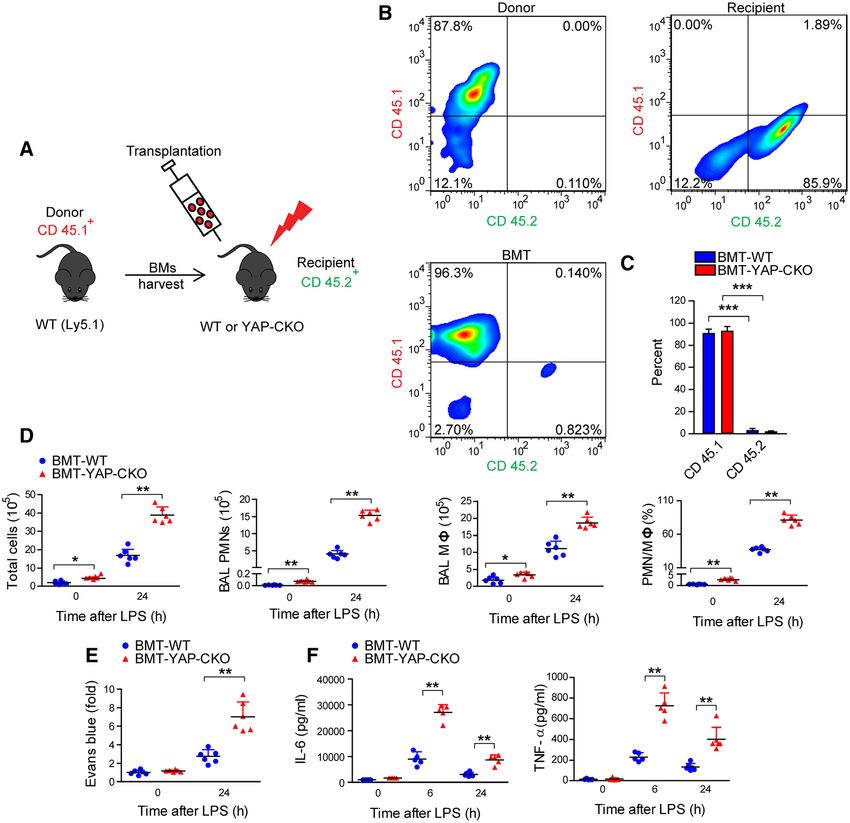
50 Circulation Research June 22, 2018
Downloaded from http://ahajournals.org by on July 21, 2021
Figure 5. YAP (Yes-associated protein) limits lipopolysaccharide (LPS)-induced endothelial inflammation. HUVECs (human
umbilical vein endothelial cells) were transfected with control CRISPR-Cas9 plasmid (Con), YAP CRISPR-Cas9 plasmid (YAP-KO), empty
vector (EV), or recombinant Flag-YAP cDNA. At 48 h posttransfection, HUVECs were stimulated with LPS (1 µg/mL) for the indicated
times. A and B, Effects of YAP deletion on the production of IL (interleukin)-6 and IL-1β. A, Representative Western blots for YAP (left)
and protein quantification by densitometry (right). B, The concentrations of IL-6 and IL-1β in HUVECs supernatants by ELISA. n=3. C and
D, Effects of YAP overexpression on the production of IL-6 and IL-1β. C, Representative Western blots for YAP; D, the concentrations of
IL-6 and IL-1β in HUVECs supernatants by ELISA. n=3 to 6. E and F, Representative immunoblots showing ICAM (intercellular adhesion
molecule)-1 and E-selectin expression after YAP knockout (E) and YAP overexpression (F) in HUVECs. G and H, Effects of YAP depletion on
polymorphonuclear neutrophil (PMN) adhesion to HUVECs (G) and PMN transendothelial migration (H). I and J, Effects of YAP overexpression
on PMN adhesion to HUVECs (I) and PMN transendothelial migration (J). n=3. *P
Lv et al YAP Limits Endothelial Activation 51
Downloaded from http://ahajournals.org by on July 21, 2021
Figure 6. YAP (Yes-associated protein) inhibits NF-κB (nuclear factor-κB) pathway secondary to direct interaction with TRAF6
(tumor necrosis factor receptor-associated factor 6). A, YAP associates with TRAF6 in HUVECs (human umbilical vein endothelial
cells) in the absence and presence of lipopolysaccharide (LPS; 1 µg/mL) stimulation. Immunoblot analysis was assessed followed by
coimmunoprecipitation with normal immunoprecipitation (IgG) or anti-TRAF6 antibody. B, YAP colocalizes with TRAF6 in HUVECs. Left,
Representative confocal images showing colocalization of YAP and TRAF6. Scale bars, 10 µm. Right, Quantification of YAP and TRAF6
colocalization. ***P
52 Circulation Research June 22, 2018
directly associated with Myc-his-TRAF6 (Figure 6D; Online Online Figure XIF) ubiquitination of TRAF6 in the presence
Figure XB). or absence of Flag-YAP plasmid, respectively. These findings
To determine whether YAP regulated NF-κB activation together indicate that YAP activates K48-linked ubiquitination
through its binding to TRAF6, we next expressed NF-κB lu- to induce TRAF6 degradation and inhibits K63-linked ubiqui-
ciferase reporter together with TRAF6 and increasing concen- tination of TRAF6, and thereby suppresses NF-κB signaling.
trations of YAP-WT or YAP-mutant cDNA in HEK293T cells.
YAP-WT strongly inhibited NF-κB activation in a concen- TRAF6 Depletion in Endothelial Cells Rescues
tration-dependent manner (Figure 6E), whereas YAP-mutant Augmented Inflammatory Phenotype of YAP-CKO
failed (Figure 6F). To further elucidate the effects of YAP on
Mice
To address whether TRAF6 expression was required and suffi-
NF-κB signaling in endothelial cells in vivo, we isolated lung
cient for lipopolysaccharide-induced increase in lung inflam-
endothelial cells from WT and YAP-CKO mice challenged
mation in YAP-CKO mice, we depleted TRAF6 in YAP-CKO
with lipopolysaccharide and observed that YAP deletion
lung vascular endothelium using liposomes incorporating
significantly increased NF-κB p65 phosphorylation (activa-
TRAF6 siRNA. Injection of liposomes containing scrambled
tion), IκBα degradation, and accumulation of p65 and p50 in
siRNA was used as control. Western blot analysis of pulmo-
nucleus both basally and after lipopolysaccharide challenge
nary endothelial cells isolated from YAP-CKO mice trans-
compared with control WT cells (Figure 6G; Online Figure
fected with siRNAs confirmed efficiency of TRAF6 depletion
XC through XE).
(Figure 8A; Online Figure XIIA). TRAF6 protein expression
YAP Modulates Ubiquitination and Degradation of in bone marrow cells was not altered by injection of liposome-
TRAF6 in Endothelial Cells TRAF6 siRNA mixture (Online Figure XIIB). Depletion of
We next determined the mechanism of YAP-mediated TRAF6 TRAF6 protein expression in YAP-CKO lung endothelial cells
suppression of NF-κB activation in endothelial cells. We ob- completely rescued lipopolysaccharide-induced increase in
served that CRISPR/Cas9 knockout of YAP in endothelial the numbers of total cells, PMNs (Figure 8B), macrophages,
cells basally upregulated TRAF6 protein expression and also PMN/macrophage ratio (Figure 8C) in bronchoalveolar la-
enhanced lipopolysaccharide-induced TRAF6 protein expres- vage fluid, and the release of IL-6 and TNF-α (Figure 8D),
sion as compared with control endothelial cells stimulated suggesting that YAP suppressed endothelial activation and
with lipopolysaccharide (Figure 7A; Online Figure XIA). We lung inflammation in vivo via TRAF6.
also observed in HEK293T cells transiently transfected with
Myc-his-TRAF6 and Flag-YAP that YAP inhibited TRAF6 Discussion
protein expression in a concentration-dependent manner YAP, a transcriptional coactivator, serves as a key regulator of
Downloaded from http://ahajournals.org by on July 21, 2021
(Figure 7B; Online Figure XIB). Based on these findings, tissue growth and organ size by promoting cell proliferation
we addressed the possibility that YAP functions by promot- and suppressing apoptosis.31 In cardiac and vascular smooth
ing TRAF6 degradation through ubiquitination. Proteins are muscle cells, YAP is an important regulator of cardiovascular
targeted for degradation by the proteasome with covalent at- development.32 Here, we identified a crucial role of cytoplas-
tachment of K48-linked polyubiquitin chains, whereas K63- mic YAP as a checkpoint modulator of endothelial activation
linked ubiquitin activates the downstream TAK1 activation and vascular inflammation. Specific deletion of YAP in endo-
important for NF-κB activation.13,30 We found that deletion thelial cells did not alter the size of organs; instead, it induced
of YAP in HUVECs significantly decreased total and K48- vascular inflammation in the absence of inflammatory stimuli
linked ubiquitination of TRAF6 but increased K63-linked and greatly augmented inflammation during endotoxemia and
ubiquitination of TRAF6 (Figure 7C; Online Figure XIC). To polymicrobial sepsis. YAP negatively regulated inflammatory
validate the role of YAP in the mechanism of ubiquitination signaling by directly interacting with the E3 ligase TRAF6
of TRAF6, we transfected HEK293T cells with Flag-YAP, which induced degradation of TRAF6 through K48-linked
Myc-his-TRAF6, and HA (hemagglutinin)-tagged ubiqui- TRAF6 ubiquitination and simultaneously inhibited K63-
tin (HA-Ub) and performed the in vivo ubiquitination assay. linked autoubiquitination of TRAF6.
Expression of Flag-YAP increased total Myc-his-TRAF6 Our study clearly demonstrated the requisite role of YAP
ubiquitination (Figure 7D; Online Figure XID). To identify in limiting endothelial activation, and thereby excessive vas-
the type of TRAF6 ubiquitination regulated by YAP, we trans- cular inflammation, and organ dysfunction. This notion was
fected HEK293T cells with Flag-YAP and Myc-his-TRAF6 strongly supported by studies in 2 murine experimental models
together with plasmids encoding K48-linked (HA-K48) of sepsis in which YAP was conditionally deleted in endothe-
(Figure 7E; Online Figure XIE) or K63-linked ubiquitin lial cells (YAP-CKO mice). Although mice with endothelial
(HA-K63; Figure 7F; Online Figure XIF). Ubiquitin mutants cell–specific deficiency of YAP did not display any gross phe-
containing single lysine-to-arginine mutation at positions notypic alteration up to adulthood, PMN infiltration into the
48 (HA-K48R) and 63 (HA-K63R) were individually trans- lung and enhanced expression of adhesion molecules suggest
fected into HEK293T cells. Expression of YAP resulted in a mild lung endothelial inflammatory activation in YAP-CKO
more K48-linked ubiquitination of TRAF6 (Figure 7E; Online mice. The inflammatory response to lipopolysaccharide and
Figure XIE) but less K63-linked ubiquitination of TRAF6 polymicrobial sepsis in YAP-CKO mice was markedly en-
(Figure 7F; Online Figure XIF). Coexpression of either HA- hanced as evident by higher tissue PMN infiltration, severe
K48R or HA-K63R prevented these changes in K48-linked lung histopathologic changes, and edema formation. Intravital
(Figure 7E; Online Figure XIE) and K63-linked (Figure 7F; microscopy studies in the cremaster muscle microcirculationLv et al YAP Limits Endothelial Activation 53
Downloaded from http://ahajournals.org by on July 21, 2021
Figure 7. YAP (Yes-associated protein) activates K48-linked ubiquitination to induce TRAF6 (tumor necrosis factor receptor-
associated factor 6) degradation and inhibits K63-linked TRAF6 ubiquitination. A, Effects of YAP depletion on TRAF6 protein
expression in HUVECs (human umbilical vein endothelial cells) stimulated with lipopolysaccharide (LPS). HUVECs were transfected with
control CRISPR-Cas9 plasmid (Con) or YAP CRISPR-Cas9 plasmid (YAP-KO). At 48 h posttransfection, cells were incubated with LPS (1
µg/mL). B, YAP decreased TRAF6 protein expression. Human embryonic kidney 293T (HEK293T) cells transfected Myc-tagged TRAF6
cDNA with increasing concentration of Flag-YAP-wild-type (WT) cDNA. C, Effects of YAP depletion on ubiquitination. Endogenous TRAF6
autoubiquitination was performed by immunoblot analysis of total ubiquitin (Ub), K48-linked ubiquitin (K48-Ub), or K63-linked ubiquitin
(K63-Ub) in HUVECs transfected with control CRISPR-Cas9 plasmid or YAP CRISPR-Cas9 plasmid, assessed after immunoprecipitation
with anti-TRAF6 antibody. D, Effects of exogenous YAP expression on total ubiquitination of exogenous TRAF6 expression. HEK293T
cells were transfected to express Myc-his-TRAF6 and HA-Ub (hemagglutinin-tagged total ubiquitin) in the presence (+) or absence (−)
Flag-YAP vector. TRAF6 ubiquitination was assessed by immunoprecipitation with anti-Myc antibody. E and F, Immunoblot analysis
of K48-linked ubiquitin (E) and K63-linked ubiquitin (F) of TRAF6 in HEK293T cells transfected to express Myc-his-TRAF6, K48-linked
ubiquitin (HA-K48-Ub), K48-linked ubiquitin mutant (HA-K48R-Ub), K63-linked ubiquitin (HA-K63-Ub), K63-linked ubiquitin mutant (HA-
K63R-Ub), with or without Flag-YAP expression. TRAF6 ubiquitination was assessed by immunoprecipitation with anti-Myc antibody.
also showed marked PMN adhesion to cremasteric vessel en- cells, as opposed to hematopoietic cells, was responsible for
dothelial cells of YAP-CKO mice both in the absence of in- endothelial cell activation and inflammation seen in the YAP-
flammatory stimulus and after lipopolysaccharide exposure. CKO mice.
Because Tie2 promoter/enhancer-driven Cre expression was TLR4/NF-κB signaling is essential for endothelial activa-
used to deplete the yap gene in endothelial cells in our study, a tion and the consequent inflammation during sepsis.5,28 Our
possible concern is that augmented endothelial activation and findings indicated that YAP prevented NF-κB activation by di-
vascular inflammation may also result from deletion of the rect interaction with TRAF6 in endothelial cells. We observed
yap gene in hematopoietic cells. To circumvent this concern, that coexpression of YAP-WT cDNA and TRAF6 cDNA
we transplanted either WT or YAP-CKO bone marrow cells inhibited NF-κB activation. Furthermore, amino acid sub-
into lethally irradiated YAP-CKO or WT mice and observed stitution mutations within the TRAF-binding site (Ser-Leu-
similar levels of endothelial activation and inflammatory re- Gln-Glu) of YAP abolished the interaction of YAP and TRAF6.
sponses in these mice. Thus, deletion of YAP in endothelial Interestingly, coexpression of YAP-mutant and TRAF6 cDNA54 Circulation Research June 22, 2018
Downloaded from http://ahajournals.org by on July 21, 2021
Figure 8. Deletion of TRAF6 (tumor necrosis factor receptor [TNF]-associated factor 6) in vascular endothelial cells prevents
lipopolysaccharide (LPS)-induced lung inflammation in YAP (Yes-associated protein)-conditional knockout (CKO) mice. Mice were
transfected with scrambled (Sc) or TRAF6 siRNA (si)-liposome complex through tail vein. At 48 h posttransfection, mice were challenged
with LPS for 24 h. A, Representative Western blot analysis showing TRAF6 knockdown in primary lung endothelial cells isolated from
wild-type (WT) and YAP-CKO mice. B and C, Total cells (B, left), polymorphonuclear neutrophils (PMNs; B, right), macrophages (Mϕ; C,
left), and PMN/Mϕ ratio (C, right) in bronchoalveolar lavage (BAL) fluid were enumerated at 24 h after LPS challenge. n=4. D, Levels of IL
(interleukin)-6 (left) and TNF-α (right) in serum measured by ELISA. n=4 to 6. **PLv et al YAP Limits Endothelial Activation 55
via suppression of K63-linked ubiquitination. Taken together, (Figure 8E). In this context, upregulation and stabilization of
YAP expression controls endothelial activation and thereby YAP expression in endothelial cells may be a novel pharmaco-
prevents hyperinflammatory response to endotoxin through logical approach for preventing sepsis-induced tissue inflam-
restraining TRAF6-mediated NF-κB activation. matory injury and organ failure.
In the present study, YAP expression in human and mouse
endothelial cells was upregulated by lipopolysaccharide, but Acknowledgments
then the expression recovered toward baseline after the in- We thank Yongkang Zhang (Department of Anesthesiology,
flammatory response exaggerated. In tumor-initiating stem- University of Illinois at Chicago) for technical assistance.
like cells39 and adipose tissue-derived mesenchymal stem
cells,40 TLR ligands also induced the expression of YAP1 Sources of Funding
and TAZ. These findings suggest a negative feedback loop This study was supported by National Institutes of Health Grant
between YAP and TLRs, whereby TLR signaling results in R01HL104092 (G. Hu), PPG1–P01 HL060678 (A.B. Malik), PPG2–
increased expression of YAP, which limits activation of NF- P01 HL077806 (A.B. Malik), and HL130028 (J. Cho).
κB to proceed and holds the inflammatory response in check.
Thus, YAP signaling may act as a molecular brake to restrict Disclosures
endothelial activation and prevent excessive inflammation None.
and organ failure.
YAP is known to play a key role in cell proliferation, References
1. Lee WL, Slutsky AS. Sepsis and endothelial permeability. N Engl J Med.
survival, differentiation, tissue regeneration, and organ size
2010;363:689–691. doi: 10.1056/NEJMcibr1007320.
determination.41 Global YAP knockout mice died at embry- 2. Andonegui G, Goyert SM, Kubes P. Lipopolysaccharide-induced leuko-
onic day 8.5.42 Kidney-specific CKO of YAP in mice resulted cyte-endothelial cell interactions: a role for CD14 versus Toll-like receptor
in defective kidneys with abnormal glomeruli, ducts with- 4 within microvessels. J Immunol. 2002;169:2111–2119.
3. Goldenberg NM, Steinberg BE, Slutsky AS, Lee WL. Broken barriers:
out lumen.43 Ectopic expression of YAP in the liver induced a new take on sepsis pathogenesis. Sci Transl Med. 2011;3:88ps25. doi:
liver enlargement and the liver reverted to a normal size on 10.1126/scitranslmed.3002011.
switching-off YAP overexpression.44,45 Liver enlargement was 4. Deutschman CS, Tracey KJ. Sepsis: current dogma and new perspectives.
also observed with knockout of Mst1/2 (molecules upstream Immunity. 2014;40:463–475. doi: 10.1016/j.immuni.2014.04.001.
5. Lawrence T. The nuclear factor NF-kappaB pathway in inflam-
of YAP). However, there was no discernable change in the mation. Cold Spring Harb Perspect Biol. 2009;1:a001651. doi:
size of the kidney, intestines, and lung in Mst1/2 knockout 10.1101/cshperspect.a001651.
mice.46 We observed that the size and weight of vital organs 6. Lomaga MA, Yeh WC, Sarosi I, et al. TRAF6 deficiency results in osteo-
Downloaded from http://ahajournals.org by on July 21, 2021
in endothelial YAP knockout mice were normal, indicating petrosis and defective interleukin-1, CD40, and LPS signaling. Genes Dev.
1999;13:1015–1024.
that YAP expression in endothelial cells is dispensable for 7. Zhang FX, Kirschning CJ, Mancinelli R, Xu XP, Jin Y, Faure E, Mantovani
organ growth and development. The primary role of YAP in A, Rothe M, Muzio M, Arditi M. Bacterial lipopolysaccharide activates
endothelial cells seems to modulate their activation second- nuclear factor-kappaB through interleukin-1 signaling mediators in cul-
tured human dermal endothelial cells and mononuclear phagocytes. J Biol
ary to YAP interaction with TRAF6, and thereby restrict the
Chem. 1999;274:7611–7614.
activation of NF-κB. 8. Muzio M, Ni J, Feng P, Dixit VM. IRAK (Pelle) family member
In contrast to our observation that YAP deletion in endo- IRAK-2 and MyD88 as proximal mediators of IL-1 signaling. Science.
thelial cells enhanced vascular inflammation, a study showed 1997;278:1612–1615.
9. Medzhitov R, Preston-Hurlburt P, Kopp E, Stadlen A, Chen C, Ghosh S,
that YAP/TAZ knockdown prevented oscillatory shear stress-
Janeway CA Jr. MyD88 is an adaptor protein in the hToll/IL-1 receptor
induced proinflammatory responses, as evidenced by reduced family signaling pathways. Mol Cell. 1998;2:253–258.
expression of endothelial adhesion molecules and THP1 10. Cao Z, Xiong J, Takeuchi M, Kurama T, Goeddel DV. TRAF6 is a
(Tamm-Horsfall protein 1) monocyte attachment.47 YAP/TAZ signal transducer for interleukin-1. Nature. 1996;383:443–446. doi:
10.1038/383443a0.
activation in HUVECs transfected with constitutively active 11. Covert MW, Leung TH, Gaston JE, Baltimore D. Achieving stabil-
YAP (S127A) and TAZ (S89A; CA-YAP/TAZ) also increased ity of lipopolysaccharide-induced NF-kappaB activation. Science.
monocyte-endothelial adhesion and the release of proinflamma- 2005;309:1854–1857. doi: 10.1126/science.1112304.
tory cytokines IL-6 and IL-8.48 The reasons for the discrepancy 12. Jiang D, Liang J, Fan J, et al. Regulation of lung injury and repair by
Toll-like receptors and hyaluronan. Nat Med. 2005;11:1173–1179. doi:
with our findings are not clear. It is possible that the differences 10.1038/nm1315.
are because of the cell types used and the inflammatory stimuli. 13. Schneider M, Zimmermann AG, Roberts RA, Zhang L, Swanson KV, Wen
Here, we determined the role of YAP in modulating lipopoly- H, Davis BK, Allen IC, Holl EK, Ye Z, Rahman AH, Conti BJ, Eitas TK,
saccharide-induced activation of NF-κB in which YAP func- Koller BH, Ting JP. The innate immune sensor NLRC3 attenuates Toll-
like receptor signaling via modification of the signaling adaptor TRAF6
tioned by interacting with TRAF6 to suppress NF-κB signaling. and transcription factor NF-κB. Nat Immunol. 2012;13:823–831. doi:
In summary, to our knowledge, this study was the first to 10.1038/ni.2378.
provide both in vitro and in vivo evidence that YAP is an es- 14. Chen ZJ. Ubiquitin signalling in the NF-kappaB pathway. Nat Cell Biol.
2005;7:758–765. doi: 10.1038/ncb0805-758.
sential regulator of endothelial activation and inflammation.
15. Gumbiner BM, Kim NG. The Hippo-YAP signaling pathway and
YAP functions by interacting with TRAF6 and prevents the contact inhibition of growth. J Cell Sci. 2014;127:709–717. doi:
activation of NF-κB by enhancing degradation of TRAF6 via 10.1242/jcs.140103.
K48 ubiquitination as well as suppressing K63-linked ubiqui- 16. Halder G, Dupont S, Piccolo S. Transduction of mechanical and cytoskel-
etal cues by YAP and TAZ. Nat Rev Mol Cell Biol. 2012;13:591–600. doi:
tination. The increase in endothelial YAP expression induced 10.1038/nrm3416.
by endotoxemia serves as an important checkpoint limit- 17. Sudol M, Harvey KF. Modularity in the Hippo signaling pathway. Trends
ing the severity of endothelial activation and inflammation Biochem Sci. 2010;35:627–633. doi: 10.1016/j.tibs.2010.05.010.56 Circulation Research June 22, 2018
18. Hao Y, Chun A, Cheung K, Rashidi B, Yang X. Tumor suppressor LATS1 35. Seijkens TTP, van Tiel CM, Kusters PJH, et al. Targeting CD40-induced
is a negative regulator of oncogene YAP. J Biol Chem. 2008;283:5496– TRAF6 signaling in macrophages reduces atherosclerosis. J Am Coll
5509. doi: 10.1074/jbc.M709037200. Cardiol. 2018;71:527–542. doi: 10.1016/j.jacc.2017.11.055.
19. Zhao B, Wei X, Li W, et al. Inactivation of YAP oncoprotein by the Hippo 36. Deng L, Wang C, Spencer E, Yang L, Braun A, You J, Slaughter C, Pickart
pathway is involved in cell contact inhibition and tissue growth control. C, Chen ZJ. Activation of the IkappaB kinase complex by TRAF6 requires
Genes Dev. 2007;21:2747–2761. doi: 10.1101/gad.1602907. a dimeric ubiquitin-conjugating enzyme complex and a unique polyubiq-
20. Sudol M, Bork P, Einbond A, Kastury K, Druck T, Negrini M, Huebner K, uitin chain. Cell. 2000;103:351–361.
Lehman D. Characterization of the mammalian YAP (Yes-associated pro- 37. Walsh MC, Lee J, Choi Y. Tumor necrosis factor receptor- associated
tein) gene and its role in defining a novel protein module, the WW domain. factor 6 (TRAF6) regulation of development, function, and homeostasis
J Biol Chem. 1995;270:14733–14741. of the immune system. Immunol Rev. 2015;266:72–92. doi: 10.1111/
21. Pan D. The hippo signaling pathway in development and cancer. Dev Cell. imr.12302.
2010;19:491–505. doi: 10.1016/j.devcel.2010.09.011. 38. Wang C, Deng L, Hong M, Akkaraju GR, Inoue J, Chen ZJ. TAK1 is a
22. Wang K, Degerny C, Xu M, Yang XJ. YAP, TAZ, and Yorkie: a conserved ubiquitin-dependent kinase of MKK and IKK. Nature. 2001;412:346–351.
family of signal-responsive transcriptional coregulators in animal de- doi: 10.1038/35085597.
velopment and human disease. Biochem Cell Biol. 2009;87:77–91. doi: 39. Chen CL, Tsukamoto H, Liu JC, Kashiwabara C, Feldman D, Sher L,
10.1139/O08-114. Dooley S, French SW, Mishra L, Petrovic L, Jeong JH, Machida K.
23. Mohseni M, Sun J, Lau A, Curtis S, Goldsmith J, Fox VL, Wei C, Frazier Reciprocal regulation by TLR4 and TGF-β in tumor-initiating stem-like
M, Samson O, Wong KK, Wong KK, Kim C, Camargo FD. A genetic cells. J Clin Invest. 2013;123:2832–2849. doi: 10.1172/JCI65859.
screen identifies an LKB1-MARK signalling axis controlling the Hippo- 40. Cho HH, Shin KK, Kim YJ, Song JS, Kim JM, Bae YC, Kim CD, Jung
YAP pathway. Nat Cell Biol. 2014;16:108–117. doi: 10.1038/ncb2884. JS. NF-kappaB activation stimulates osteogenic differentiation of mesen-
24. Liu B, Zheng Y, Yin F, Yu J, Silverman N, Pan D. Toll receptor-me- chymal stem cells derived from human adipose tissue by increasing TAZ
diated hippo signaling controls innate immunity in drosophila. Cell. expression. J Cell Physiol. 2010;223:168–177. doi: 10.1002/jcp.22024.
2016;164:406–419. doi: 10.1016/j.cell.2015.12.029. 41. Yimlamai D, Christodoulou C, Galli GG, Yanger K, Pepe-Mooney B,
25. Koni PA, Joshi SK, Temann UA, Olson D, Burkly L, Flavell RA. Gurung B, Shrestha K, Cahan P, Stanger BZ, Camargo FD. Hippo path-
Conditional vascular cell adhesion molecule 1 deletion in mice: impaired way activity influences liver cell fate. Cell. 2014;157:1324–1338. doi:
lymphocyte migration to bone marrow. J Exp Med. 2001;193:741–754. 10.1016/j.cell.2014.03.060.
26. Kim K, Li J, Tseng A, Andrews RK, Cho J. NOX2 is critical for hetero- 42. Morin-Kensicki EM, Boone BN, Howell M, Stonebraker JR, Teed J, Alb
typic neutrophil-platelet interactions during vascular inflammation. Blood. JG, Magnuson TR, O’Neal W, Milgram SL. Defects in yolk sac vascu-
2015;126:1952–1964. doi: 10.1182/blood-2014-10-605261. logenesis, chorioallantoic fusion, and embryonic axis elongation in mice
27. Varon D, Jackson DE, Shenkman B, Dardik R, Tamarin I, Savion N, with targeted disruption of Yap65. Mol Cell Biol. 2006;26:77–87. doi:
Newman PJ. Platelet/endothelial cell adhesion molecule-1 serves as a co- 10.1128/MCB.26.1.77-87.2006.
stimulatory agonist receptor that modulates integrin-dependent adhesion 43. Reginensi A, Scott RP, Gregorieff A, Bagherie-Lachidan M, Chung C,
and aggregation of human platelets. Blood. 1998;91:500–507. Lim DS, Pawson T, Wrana J, McNeill H. Yap- and Cdc42-dependent
28. Dauphinee SM, Karsan A. Lipopolysaccharide signaling in endothelial nephrogenesis and morphogenesis during mouse kidney development.
cells. Lab Invest. 2006;86:9–22. doi: 10.1038/labinvest.3700366. PLoS Genet. 2013;9:e1003380. doi: 10.1371/journal.pgen.1003380.
29. Ye H, Park YC, Kreishman M, Kieff E, Wu H. The structural basis for 44. Dong J, Feldmann G, Huang J, Wu S, Zhang N, Comerford SA, Gayyed
the recognition of diverse receptor sequences by TRAF2. Mol Cell. MF, Anders RA, Maitra A, Pan D. Elucidation of a universal size-control
Downloaded from http://ahajournals.org by on July 21, 2021
1999;4:321–330. mechanism in Drosophila and mammals. Cell. 2007;130:1120–1133. doi:
30. Jiao S, Zhang Z, Li C, et al. The kinase MST4 limits inflammatory re- 10.1016/j.cell.2007.07.019.
sponses through direct phosphorylation of the adaptor TRAF6. Nat 45. Camargo FD, Gokhale S, Johnnidis JB, Fu D, Bell GW, Jaenisch R,
Immunol. 2015;16:246–257. doi: 10.1038/ni.3097. Brummelkamp TR. YAP1 increases organ size and expands undif-
31. Zhao B, Tumaneng K, Guan KL. The Hippo pathway in organ size ferentiated progenitor cells. Curr Biol. 2007;17:2054–2060. doi:
control, tissue regeneration and stem cell self-renewal. Nat Cell Biol. 10.1016/j.cub.2007.10.039.
2011;13:877–883. doi: 10.1038/ncb2303. 46. Song H, Mak KK, Topol L, Yun K, Hu J, Garrett L, Chen Y, Park O,
32. Wang Y, Hu G, Liu F, Wang X, Wu M, Schwarz JJ, Zhou J. Deletion of yes- Chang J, Simpson RM, Wang CY, Gao B, Jiang J, Yang Y. Mammalian
associated protein (YAP) specifically in cardiac and vascular smooth muscle Mst1 and Mst2 kinases play essential roles in organ size control and tu-
cells reveals a crucial role for YAP in mouse cardiovascular development. mor suppression. Proc Natl Acad Sci USA. 2010;107:1431–1436. doi:
Circ Res. 2014;114:957–965. doi: 10.1161/CIRCRESAHA.114.303411. 10.1073/pnas.0911409107.
33. Chung JY, Park YC, Ye H, Wu H. All TRAFs are not created equal: com- 47. Wang KC, Yeh YT, Nguyen P, Limqueco E, Lopez J, Thorossian S,
mon and distinct molecular mechanisms of TRAF-mediated signal trans- Guan KL, Li YJ, Chien S. Flow-dependent YAP/TAZ activities regulate
duction. J Cell Sci. 2002;115:679–688. endothelial phenotypes and atherosclerosis. Proc Natl Acad Sci USA.
34. Lutgens E, Lievens D, Beckers L, et al. Deficient CD40-TRAF6 signaling 2016;113:11525–11530. doi: 10.1073/pnas.1613121113.
in leukocytes prevents atherosclerosis by skewing the immune response 48. Wang L, Luo JY, Li B, et al. Integrin-YAP/TAZ-JNK cascade me-
toward an antiinflammatory profile. J Exp Med. 2010;207:391–404. doi: diates atheroprotective effect of unidirectional shear flow. Nature.
10.1084/jem.20091293. 2016;540:579–582.1
Supplemental Material
Reagents
LPS (E.coli. 0111:B4), Hydrogen peroxide solution (H2O2, #216763) was purchased from Sigma Aldrich.
Ultrapure LPS (E. coli O111:B4) was obtained from InvivoGen. Anti-YAP (#4912), anti-TRAF6 (#8028),
anti-HA (#3724), anti-K63-Ub (#5621), anti-K48-Ub (#4289), anti-Flag (#8146), and anti-Myc (#2278),
anti-p50 (#13586), anti-p65 (#8242), and anti-IкBα (#4814) antibodies were from Cell Signaling. Anti-
TRAF6 (sc-7221), anti-YAP (sc-101199), anti-Ub (sc-8017), anti-ICAM-1 (sc-8439), anti-E-selectin (sc-
137054), anti-c-Myc (sc-40), anti-VE-cadherin (sc-28644), anti-p-p65 (sc-33020), and anti-Lamin A (sc-
20680) antibodies were from Santa Cruz. Recombinant Human TNF-α (300-01A) was from Peprotech.
Mice
Endothelial cell-specific YAP deficiency mice on a C57BL/6 background were generated by cross-
breeding Tie2-Cre transgenic mice with mice carrying the floxed YAP gene for at least 5 generations. All
the mice used in the experiments were the littermates of genotypes of YAPf/f with Cre gene as YAP-CKO
while the ones only carry YAPf/f as WT mice. All mice were bred and maintained in the Association for
Assessment and Accreditation of Laboratory Animal Care-accredited animal facilities at the University of
Illinois at Chicago according to NIH guidelines. The use of all mice (8-10 weeks old) was approved by
the University of Illinois Institutional Animal Care and Use Committee. Relatively equal proportions
(ratio 1:1) of male and female mice were used in each experiment. Experimental groups were assigned
using a simple randomization procedure by means of drawing lots.
Genotyping
All mice were genotyped prior to use. Genomic DNA was extracted from the tail tips (3-5 mm) of mice
and amplified by using Sigma REDExtract-N-AMP Tissue PCR kit (Sigma Aldrich, XNAT-100RXN)
according to manufacturers' instructions. Briefly, the tail samples were incubated in Tissue Preparation
Solution and Extraction Solution for 10 min at room temperature and heated at 95 °C for 3 min.
Neutralization Solution B was finally added to terminate reaction. An aliquot of the DNA extract was
then added directly to the optimized PCR mix supplied. PCR was performed using 0.5 µl genomic DNA
with 8 µl REDExtract-N-AMP PCR Ready mix, 1.6 µl H2O, 1.2 µl forward and 1.2 µl reverse primer (20
µmol/L, 1.25 µl Extract Solution and 1.25 µl Neutralization B Solution. The following PCR primer
sequences were used for analysis: mouse Yap flox 5’-CACCAAACCTGGCATAGACATGTGTTC-3’
and 5’-CAGTCTGTAACAACC-AGTCAGGGATAC-3’, mouse Tie-2 Cre 5’TTCCATGAGTGAACG-
AACCTGGTC-3’ and 5’-AGTGATGAGGTTCGCAAGAACCTG-3’. To detect YAP flox, the PCR
reaction was performed as 1 cycle on 94 °C for 3 min, followed by 35 cycles on 94 °C for 30 sec, 57 °C
for 45 sec, 72 °C for 1 min, and at last 72 °C for 10 min. That would generate a 557-base-pair (bp) band
from the wild-type allele and a 661-bp band from the YAP flox allele. To detect Tie2-Cre, the PCR
reaction was performed as 1 cycle on 94 °C for 3 min, followed by 35 cycles on 94 °C for 30 sec, 62 °C
for 1 min, 72 °C for 1 min, and at last 72 °C for 2 min, that generated a 360-bp band from the Tie-2 Cre
allele.
Cell culture
Human umbilical vein endothelial cells (HUVECs) and human lung microvascular endothelial cells
(HLMVECs) were cultured in EBM-2 (Lonza, CC-3156) supplemented with EGM-2 BulletKit (Lonza,
CC-4176) and EGM-2-MV BulletKit (Lonza, CC-4147), respectively. All experiments were conducted2 using HUVECs and HLMVECs between passages 3 and 6. HEK293T cells were maintained in high glucose DMEM (Cellgro, 10-013-CV) supplemented with 10% FBS (Sigma, F2442), penicillin and streptomycin. Isolation of primary mouse lung endothelial cell Primary lung endothelial cells were isolated by using immunoselection method.1 Lungs were removed and digested in HBSS supplemented with Collagenase/Dispase (Roche, 11097113001) for 40 min at 37 °C, followed by filtration with 70 µm and 40 µm strainers. Lung endothelial cells were then isolated using magnetic beads (Dynabeads Sheep anti-Rat IgG, Invitrogen, 11035) pre-coated with purified rat anti-mouse PECAM-1 specific antibody (BD Pharmingen, 553370). Primary purified endothelial cells and non-endothelial cells were lysed respectively with RIPA buffer (Boston BioProducts, BP-115) for Western blot. Isolation of bone marrow cells Bone marrow was harvested from mouse tibias and femurs using a 25 gauge needle, passed through a 70 μm nylon mesh cell strainer (Falcon). Red blood cells were lysed with ammonium-chloride-potassium (ACK) lysing buffer (Quality Biological, #118-156-101). Bone marrow cells were then counted, resuspended in PBS, and kept on ice prior to transplantation or lysed with RIPA lysis buffer for Western Blotting. Cell Fractionation Cells were fractionated using NE-PER™ nuclear and cytoplasmic extraction reagents (ThermoFisher) to generate cytosolic and nuclear pools as described by the manufacturer. CRISPR/Cas9 studies Human YAP CRISPR/Cas9 KO Plasmid (Santa Cruz, sc-400040) or control CRISPR/Cas9 Plasmid (Santa Cruz, sc-418922) were transfected into 80% confluent HUVECs or HLMVECs in antibiotic-free standard growth medium according to the manufacturer’s protocol. At 48 h posttransfection, Western blot analysis was performed to confirm the efficiency of YAP protein deletion. cDNA transfection HUVECs, HLMVECs or HEK293T cells were transfected with Flag-YAP-WT, Flag-YAP-Mutant, Myc- his-TRAF6, HA-Ub, HA-K48-Ub, HA-K48R-Ub, HA-K63-Ub, or HA-K63R-Ub with Lipofectamine 2000 (Invitrogen, 11668019) according to the manufacturer’s protocol. At 48 h posttransfection, Western blot analysis was performed to confirm the efficiency of transfection. Quantitative real time RT-PCR analysis Total RNA was isolated using Trizol (Invitrogen Life Technologies). Following reverse transcription with High-capacity cDNA reverse transcription kit (Applied Biosystems), quantitative real-time PCR analysis was performed using ABI 7500 system with Brilliant II SYBR Green QPCR Master Mix (Agilent). The primers used for analysis were listed as following: human YAP, 5’-GAACCCCAGATGACTTCCTG-3’ and 5’-CTCCTTCCAGTGTTCCAAGG-3’; human actin, 5’-GCACAGAGCCTCGCCTT-3’ and 5’- GTTGTCGACGACGAGCG-3’; mouse YAP, 5’-GTCCTCCTTTGAGATCCCTGA-3’ and 5’-TGTTG- TTGTCTGATCGTTGTGAT -3’; mouse GAPDH, 5’-CCCAATGTGTCCGTCGTGGAT-3’ and 5’- TGTAGCCCAAGATGCCCTTCAG-3’. Data were presented as the fold change relative to control.
3 Western blotting and immunoprecipitation Cells or lung tissues were lysed in RIPA buffer supplemented with 1 mmol/L Na3VO4, protease inhibitor cocktail (Sigma, P8340), and 1 mmol/L PMSF. Lysates were sonicated and incubated for 30 min on the ice followed by centrifuging at 13,200 rpm for 15 min at 4 °C. Protein concentration was determined with DC Protein Assay Reagent (Bio-Rad). Equal amounts of samples were loaded for SDS-PAGE and transferred onto PVDF (Millipore, 0.45 µm, IPVH00010) membranes. The membranes were blocked with 5% nonfat milk and incubated with primary antibody overnight at 4 °C. Then the membranes were probed with HRP-conjugated secondary antibody for 1 h at room temperature. The protein bands were detected with ECL SuperSignal reagent (Pierce, #34078). Relative band densities of the proteins were analyzed by using ImageJ Software (NIH). For immunoprecipitation,2, 3 cells were lysed in buffer containing 150 mmol/L NaCl, 50 mmol/L Tris- HCl, 1% NP-40, 1 mmol/L EDTA, 1 mmol/L NaF, 1 mmol/L Na3VO4, 1 mmol/L PMSF, and protease inhibitor cocktail. Proteins were precleared by using 1-2 µg normal IgG together with protein A/G PLUS- agarose beads (Santa Cruz, sc-2003) for 1 h, and then incubated with primary antibody at 4 °C overnight, followed by further incubation with 20 µl beads at 4 °C for 2 h. Beads were then washed 5-6 times with lysis buffer, and the pellets were eluted with SDS loading buffer and resolved by SDS-PAGE. Ubiquitination assay For in vivo ubiquitination assays,4 HEK293T cells were transfected with plasmids for 48 hours and then lysed in 1.5% SDS cell lysis buffer. Lysates were boiled for 10 min immediately followed by centrifuging for 10 min at 16,000 g. The supernatants were collected and diluted with 1% NP-40 Co-IP buffer, then immunoprecipitated and analyzed by SDS-PAGE. Immunofluorescence Immunofluorescence was performed as previously described.5 Cells plated on coverslips were fixed with 4% paraformaldehyde for 10 min, washed 3 times with 100 mmol/L glycine in HBSS for 10 min followed by washing once with HBSS for 10 min. Cells were permeabilized with 0.2% Triton X-100 in PBS for 10 min and then blocked with 5% donkey serum for 1 hour at RT followed by incubating with anti-YAP or anti-TRAF6 antibody overnight at 4°C. After washing for 4 times, the cells were incubated by donkey anti-mouse 488 (Invitrogen, A-21202) and donkey anti-rabbit 594 (Invitrogen, A-21207) secondary antibodies for 1 hour at RT. The cells were rinsed 4 times, and finally mounted on glass slides using Vectashield mounting medium (Vector, H-1500) containing DAPI. For the immunohistochemistry of lung and cremaster muscle, the tissues were embedded in paraffin, sectioned, and mounted to the slides. All the slides were rehydrated with xylene and decreasing concentrations of ethanol followed by antigen retrieval using sodium citrate buffer. The slides were then incubated with anti-PECAM-1 (Abcam, ab28364), anti- ICAM-1, and anti-E-selectin antibodies overnight at 4°C. After extensive washing with PBST, the slides were incubated with secondary antibody for 1 h and mounted with Vectashield containing DAPI. Confocal images were acquired with a laser-scanning confocal microscope (Zeiss LSM 510 META) using Hg lamp and UV-filter set to detect DAPI [band pass (BP) 385–470 nm emission], 488 nm excitation laser line to Alexa 488 (BP505-550 nm emission), and 568 nm excitation laser line to Alexa Fluor 568 (excitation/emission ~578/603 nm). Optical sections had a thickness of < 1 μm (pinhole set to achieve 1 Airy unit). Luciferase assay
You can also read

















































