The anticancer Potency of Artemisinin and Its Derivatives
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Review Article
ISSN: 2635-0955
DOI: https://doi.org/10.52152/spr/2021.116
The anticancer Potency of Artemisinin and
Its Derivatives
Yimiao Lin1*, Zhixuan Song2, Zhian Xi3
1
School of International Education, Beijing University of Chemical Technology, Changping District,
Beijing 100029, P.R. China
2
Liaoning Province Shiyan High School, Shenyang, Liaoning 110031, China
3
Chemistry Department, the College of Science, Virginia Polytechnic Institute and State University,
Blacksburg, Virginia 24060, United States
*Corresponding author: Yimiao Lin, School of International Education, Beijing University of
Chemical Technology, Changping District, Beijing 100029, P.R. China.
Email:2019090121@mail.buct.edu.cn
Submission Date: 11/3/2021 Acceptance Date: 02/4/2021 Published Date: 09/04/2021
ABSTRACT
Artemisinin and its derivatives (Artemisinins) have long been used as antimalaria drugs with
considerable safety and efficacy. In recent preclinical researches, artemisinins also exert an anticancer
potency via induction of programmed cell death and inhibition of cell growth. Artemisinins can be
activated by heme to form Reactive Oxygen Species (ROS), which damage lipids or activate
mitochondria-mediated pathway to induce caspase cascades. Artemisinins also get involved in the
regulation of protein and gene expression to inhibit the VEGF signaling pathway that is responsible for
cell growth and survival. In this review, we focus on current knowledge of the modulation of some
detailed pathways that induce apoptosis and inhibit cell growth, including iron-dependent pathway,
mitochondrial-mediated pathway, and VEGF signaling pathway. We also collect up-to-date researches
to support their efficacy. With future researches and clinical investigation on artemisinins, more detailed
molecular mechanisms and anticancer effects will be identified and confirmed.
Keywords: Artemisinins, anticancer, heme, mitochondrial-mediated pathway, caspase, NF-κB pathway,
PI3K/Akt/mTOR pathway
Citation: Lin Y, Song Z, Xi Z, The anticancer Potency of Artemisinin and Its Derivatives, SPR, 2021, 43
Volume 1, issue, 2, Page No.: 43 - 51. DOI: https://doi.org/10.52152/spr/2021.116
INTRODUCTION
Artemisinin, with a molecular formula established ‘National Project 523’ to discover
C15H22O5, is a sesquiterpene lactone containing value drugs from traditional Chinese plants. Tu
a peroxide bridge. The first record of Youyou, who was influenced by Zhouhou
artemisinin (ARS) should date back to Beiji Fang in her childhood, showed interest in
thousands of years ago in China. The treatment the leaves of Artemesia annua L (sweet
of malaria was described in a handbook called wormwood), and her group finally extracted
Zhouhou Beiji Fang by Ge Hong in Jin ARS under cold temperature. (1)
Dynasty. In 1967, the Chinese government
Figure 1. The structure of Artesunate (ART) and Dihydroartemisinin (DHA) are designed for
better hydrophilic effect (2)
Starting from the test in mouse malaria in we will focus on some typical mechanisms of
1971, the anti-malaria effect of ARS was ARS-type drug-induced anticancer effects.
gradually recognized by the world. The
structure of the 1, 2, 4-trioxane ring in ARS 2. ART/DHA-induced apoptosis.
leads to the quick killing of parasites. Then,
2.1 Formation of ROS.
considering the short half-life and incomplete
course treatment of this drug, scientists began When DHA enters the human body, it is
to study the combination of ARS with other activated by the free heme derived from the
drugs for the protection and prevention of drug parasites digesting hemoglobin. A recent work
resistance. (1) Researchers also designed new done by Melissa with his coworkers shows that
drugs such as DHA (Dihydroartemisinin) and only trace amounts of heme within the parasite
ART (Artesunate), shown in figure 1 to are enough to stimulate and activate DHA. (3)
promote metabolism in the human body. As is shown in figure 2, there is an
The anticancer effect of ARS and its endo-peroxide bridge in DHA. (4) The reactive
derivatives is studied by scientists in recent endoperoxide bridge of DHA will be cleaved
years. They are considered anticancer drugs when it reacts with heme (Fe2+-FPIX). Then
due to the fact of the higher level of iron in DHA will be transformed into a short-lived
cancer cells. ARS-type drugs can react with Fe alkoxy radical. This alkoxy radical will go
(II) to create reactive oxygen species (ROS). through a rearrangement because of
These reactive radicals play a role in cell thermodynamic induction and will produce a
apoptosis, including cleavage of cell primary carbon-centered free radical. Such
membrane and activation of caspases carbon-centered free radicals can attack
regulating DNA fragmentations. Also, macromolecules and react with other structures
scientists have found that ARS-type drugs, in such as proteins and lipids, which leads to
the form of DHA, can regulate multiple widespread cellular damage, especially for
signaling pathways to inhibit the VEGF- tumor cells.
induced MAPK pathway which is responsible
for tumor cell proliferation. (2) In this review,
Citation: Lin Y, Song Z, Xi Z, The anticancer Potency of Artemisinin and Its Derivatives, SPR, 2021, 44
Volume 1, issue, 2, Page No.: 43 - 51. DOI: https://doi.org/10.52152/spr/2021.116
Figure 2. Mechanism of ROS formation.
Fe (II) is oxidized, and one of the electrons forms a carbon radical (4)
Also, in the fact that lysosomes are the is the damage of mitochondria and lysosomes
storage site for Fe2+, they are easily affected by that causes cell death, including the death of
drugs targeting iron, such as artemisinin cancer cells. Also, these ROS can lead to
derivatives. ARTs can release ROS via the protein and DNA damage, thus further causing
Fenton reaction in lysosomes. These ROS can the death of tumor cells.
influence the electron transfer chain in the
mitochondria and causes damage to lysosomes
and mitochondria, as Figure 3 shows. (5) And it
Figure 3. ROS cause programmed cell death (5)
2.2 Caspase-dependent anticancer effect. membrane, peroxidation of membrane lipid
and also permeability transition (6), finally
As is previously described, ROS can be
leading to the collapse of the mitochondria
very damaging to mitochondria. Reactive HO·
membrane. (7) At this point, cytochrome c,
and ROO· radicals, transformed from ROS via
originally stored in the mitochondria, would be
Fenton reaction, trigger a series of changes,
released into the cytoplasm.
including depolarization of the mitochondrial
Citation: Lin Y, Song Z, Xi Z, The anticancer Potency of Artemisinin and Its Derivatives, SPR, 2021, 45
Volume 1, issue, 2, Page No.: 43 - 51. DOI: https://doi.org/10.52152/spr/2021.116
Figure 4. The mechanism of the mitochondria-mediated pathway is induced by ROS.
ART supregulate the level of activated caspase-9, caspase-3 and nucleus AIF (11)
As shown in figure 4, released ARTs. (10) Caspases, a large family of
cytochrome c would first bind to APAF1 and proteases, cleave after an aspartate residue in
induce a conformational change. This change its protein substrate and thus selectively
allows it to combine with dATP to form a inactivate some important proteins, including
quaternary protein structure called the ICAD (the inhibitor of caspase-activated
apoptosome, which leads to the activation of DNase) and Poly (ADP-ribose) polymerase
caspase-9 and further the activation of caspase- (PARP) enzymes, which commits the cells to
3. (8), (6) Handrick and his colleagues observed death. (11) Caspases can promote DNA
the release of cytochrome c, activation of fragmentation via the cleavage of ICAD.
caspases, DNA fragmentation, and Normally, ICAD combines with CAD to
consequently apoptosis, in T-cell lymphoma complete its translation and maintains an
Jurkat cells after the dysfunction of association with CAD to block its dimerization
mitochondria induced by artemisinin and its and inhibit its DNase activity. (12) DNA
active metabolite DHA. (9) In short, the fragmentation occurs when activated caspase-3
initiation of the caspase-dependent pathway cleaves ICAD and thus activates CAD. (13)
has been shown to be involved with the This fragmentation appears to be significantly
generation of ROS induced by ARTs, as amplified with artemisinin treatment, which
chelators of Fe3+ significantly alleviate the causes an increased level of cleaved ICAD. (9)
extent of increased caspase and cytotoxicity of
Citation: Lin Y, Song Z, Xi Z, The anticancer Potency of Artemisinin and Its Derivatives, SPR, 2021, 46
Volume 1, issue, 2, Page No.: 43 - 51. DOI: https://doi.org/10.52152/spr/2021.116Figure 5.The level of caspases and cleaved-PARP in cells treated without(negative control group)
and with (1, 5, 10 µM) artemisinin: the level of caspase-3, caspase-8, caspase-9 and cleaved-PARP
shows a significant increase after artemisinin treatment (14)
Caspases also promote DNA caspase inhibitor (Z-VAD-FMK), Green-
fragmentation by blocking the repair shields and colleagues still observed the ART-
mechanism. For example, Poly(ADP-ribose) mediated cytotoxicity. (23) This finding
polymerase (PARP) enzymes, which are the indicates the existence of another caspase-
chief responders and contributors to DNA independent pathway, which is mediated by
repair, are influenced by caspases, especially Apoptosis-Inducing Factors (AIF).
caspase-3 and caspase-7. (15, (16) Artemisinin has AIF is a mammalian mitochondria
been observed to promote the cleavage of protein identified as a flavoprotein
PARP in a caspase-dependent manner in both oxidoreductase. After ROS damages the
HCT116 colon cancer cells and SKM1 cells, as mitochondria membrane, AIF will translocate
is shown in figure 5. (17), (18) The cleaved, or to the nucleus and bring about chromatin
inactivated PARP would fail to function condensation and DNA fragmentation without
normally and consequently accelerate the the help of caspase. (24) After ART treatment in
process of apoptosis. (19) ART and its cultured cells, significantly increased nAIF
metabolite DHA are responsible for the (nucleus AIF) detected in western blots in
apoptosis of human lung adenocarcinoma figure 6 verifies the involvement of AIF in
cells, colorectal cancer cells and breast cancer ART-induced apoptosis. (18) The translocation
cells via the caspase-dependent pathway, as and function of AIF can be independent of
evidenced by elevated levels of caspase-3, 8 caspases. Nevertheless, caspase-dependent and
and 9 (Figure 5) in cell lines treated with independent pathway are not mutually
ART. (20), (21), (14), (22) exclusive. Cleaved PARP, the substrate of
caspase-3 and caspase-7, also promotes the
2.3 Caspase-independent anticancer effect release of AIF. (25) Furthermore, the
combination of caspase-dependent and
Caspases really assume a pivotal role in
independent pathway can lead to synergistic
artemisinin-induced apoptosis. However, when
effects in apoptosis. (23)
pretreating the ovarian cancer cells with a pan-
Citation: Lin Y, Song Z, Xi Z, The anticancer Potency of Artemisinin and Its Derivatives, SPR, 2021, 47
Volume 1, issue, 2, Page No.: 43 - 51. DOI: https://doi.org/10.52152/spr/2021.116Figure 6.The level of nAIF (nucleus AIF) and tAIF (Total AIF) in cells treated with and
without artemisinin: nAIF significantly increased but tAIF remained unchanged after
artemisinin treatment (14)
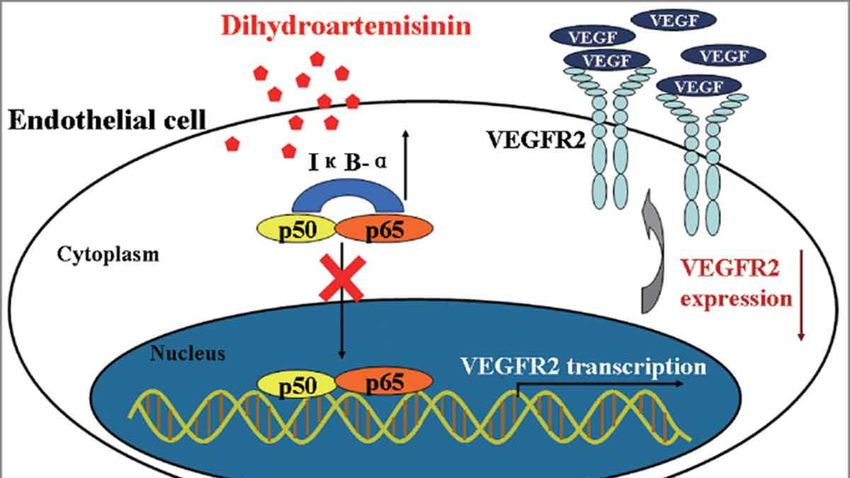
3. Inhibition of VEGF signaling by DHA. experiment that DHA can decrease the binding
of p65 protein to VEGFR-2 promotor by
The elevated level of VEGF expression
changing the motif of NF-κB instead of
is found in most types of cancer. The binding
changing the DNA sequences, which leads to
of VEGF-A to VEGFR-2 activates the
the suppression of the VEGFR-2 production in
mitogen-activated protein kinase (MAPK)
human umbilical vein endothelial cells. (27)
cascades and PI3K pathway that controls the
Inhibition of this transcription also causes the
cell proliferation and survival for tumor
decrease level of IL-8, a pro-angiogenic
angiogenesis. (26) Scientists have found that
cytokine acting as an autocrine growth factor
DHA can inhibit the process of VEGF binding
in colorectal cancer. (28), (29)
VEGFR-2. One of the examples is through the
inhibition of the NF-κB pathway. Fengyun
Dong and his colleagues have observed in the
Figure 7. VEGFR-2 production is reduced by changing the binding capacity of p65, which leads
to inhibition of transcription (27)
Citation: Lin Y, Song Z, Xi Z, The anticancer Potency of Artemisinin and Its Derivatives, SPR, 2021, 48
Volume 1, issue, 2, Page No.: 43 - 51. DOI: https://doi.org/10.52152/spr/2021.116DHA can also regulatePI3-kinase/Akt/mTOR phosphorylation processes in the PI3-
signaling pathway to decrease the formation of kinase/Akt/mTOR signaling pathway. Min
HIF-1α, which is a transcription factor Kong has compared the effect of drugs
responsible for VEGF expression. Compared between ART and AKT inhibitor VII on
with the role in the NF-κB pathway, DHA ccRCC cells in his experiment and observed
does not directly block the transcription the inactivation of AKT activity to mTOR. (31)
process. Instead, it reduces the formation of Studies have also shown that DHA directly
HIF-1α through down-regulation of the blocks the phosphorylation of 4E-BP1 by
translation factor. As shown in Figure 8, the mTOR. (30), (32) Since the formation of HIF-1α
4E-BP1/eIF-4E complex plays a significant requires degradation of the 4E-BP1/eIF-4E
role in inhibiting HIF-1α translation. (30) complex, translation of HIF-1α will be
Phosphorylation of 4E-BP1 can reduce its suppressed if 4E-BP1 cannot be
affinity to eIF-4E, which leads to upregulation phosphorylated.
of protein. (30) DHA can inhibit all the
Figure 8. PI3-kinase/Akt/mTOR signaling pathway, activated by VEGF, has positive
feedback on VEGF signaling. DHA can block the phosphorylation of 4E-BP1 to inhibit the
translation of HIF-1α, thus reducing VEGF expression (33)
Conclusion and Future Experiment
Artemisinin and its derivatives, known cancer cells. Studies have suggested that ARTs
as effective antimalarial drugs, may have induce cancer cell death and inhibit cell
transposable mechanisms between induction of proliferation via various mechanism, including
parasite death and cancer cell apoptosis and iron-dependent, caspase-dependent and -
exert a potential anticancer activity. Owing to independent pathway, and down-regulation of
the generation of ROS from the break of the VEGF signaling. Besides, the combination of
end peroxide bridge, ARTs theoretically artemisinin and some anticancer
exhibit high cytotoxicity specific to most pharmacophores contributes to additive or
Citation: Lin Y, Song Z, Xi Z, The anticancer Potency of Artemisinin and Its Derivatives, SPR, 2021, 49
Volume 1, issue, 2, Page No.: 43 - 51. DOI: https://doi.org/10.52152/spr/2021.116synergistic effects in mitochondria-mediated Drug Chem Toxicol. 2021; 44(1):47-57.
pathway and anti-angiogenesis. (22), (34) 7. Groninger E, Boer MD, De Graaf S,
However, there is still a long way to go Kamps W, De Bont E. Vincristine
before ARTs can be safely and effectively induced apoptosis in acute lympho-
applied to cancer therapy. In absence of blastic leukaemia cells: a mitochondrial
clinical trials and research, current insight into controlled pathway regulated by
ARTs remains insufficient to evidence their reactive oxygen species? Int. J. Oncol.
potency and practicability in cancer treatment. 2002; 21(6): 1339-1345.
Some molecular mechanisms of the ARTs- 8. Singh MH, Brooke SM, Zemlyak I,
induced cytotoxicity are still unclear or Sapolsky RM. Evidence for caspase
unknown to us. (35) Apart from that, several effects on release of cytochrome c and
clinical cases reported on the underlying AIF in a model of ischemia in cortical
embryotoxicity (36) and neurotoxicity (37) and neurons. Neurosci. Lett. 2010; 469(2):
raised doubt on the safety of ARTs, which led 179-183.
the World Health Organization to set a 9. Handrick R, Ontikatze T, Bauer KD,
restriction on the usage of ARTs. (38) In Freier F, Rübel A, Dürig J, Belka C,
general, more research should focus on the Jendrossek V. Dihydroartemisinin
molecular mechanisms and the clinical induces apoptosis by a Bak-dependent
treatment of artemisinin derivatives. intrinsic pathway. Mol Cancer Ther.
Artemisinin and its derivatives can be a 2010;9(9):2497-510.
promising candidate in fighting against the 10. Antoine T, Fisher N, Amewu R, O'Neill
global cancer pandemic. PM, Ward SA, Biagini G A. Rapid kill
of malaria parasites by artemisinin and
REFERENCE semi-synthetic endoperoxides involves
ROS-dependent depolarization of the
1. Miller LH, Su, X. Artemisinin:
membrane potential. J Antimicrob
discovery from the Chinese herbal
Chemother. 2014; 69(4): 1005-1016.
garden. Cell. 2011;146(6):855-858.
11. Riedl SJ, Shi Y. Molecular mechanisms
2. Mancuso, R, Foglio M, Saad SO.
of caspase regulation during apoptosis.
Artemisinin-type drugs for the
Nat. Rev. Mol. Cell Biol. 2004;
treatment of hematological malignan-
5(11):897-907.
cies. Cancer Chemother. Pharmacol.
12. Larsen BD, Sørensen CS. The caspase‐
2020; 1-22.
activated DN ase: apoptosis and
3. Rosenthal MR, Ng CL. Plasmodium
beyond. FEBS J. 2017; 284(8): 1160-
falciparum Artemisinin Resistance: The
1170.
Effect of Heme, Protein Damage, and
13. Nagata S. Apoptotic DNA fragment-
Parasite Cell Stress Response. ACS
ation. Exp. Cell Res. 2000; 256(1): 12-
Infect. Dis. 2020; 6(7): 1599–1614.
18.
4. Shen Y, Zhang B, Su Y, Badshah SA,
14. Lu YY, Chen TS, Qu JL, Pan WL, Sun
Wang X, Li X, Xue Y, Xie L, Wang Z,
L, Wei XB. Dihydroartemisinin (DHA)
Yang Z, Zhang G, Shang P. Iron
induces caspase-3-dependent apoptosis
promotes dihydroartemisinin cyto-
in human lung adenocarcinoma ASTC-
toxicity via ROS production and
a-1 cells. J. Biomed. Sci. 2009; 16(1):
blockade of autophagic flux via
1-15.
lysosomal damage in osteosarcoma.
15. Germain M, Affar EB, D’Amours D,
Front. Pharmacol. 2020; 11: 444.
Dixit VM, Salvesen GS, Poirier GG.
5. Morel I, Cillard J, Lescoat G, Sergent
Cleavage of automodified poly (ADP-
O, Pasdeloup N, Ocaktan AZ, Abdallah
ribose) polymerase during apoptosis
MA, Brissot P, Cillard P. Antioxidant
evidence for involvement of caspase-7.
and free radical scavenging activities of
J. Biol. Chem. 1999; 274(40): 28379-
the iron chelators pyoverdin and
28384.
hydroxypyrid-4-ones in iron-loaded
16. Pascal JM. The comings and goings of
hepatocyte cultures: comparison of their
PARP-1 in response to DNA damage.
mechanism of protection with that of
DNA repair. 2018; 71: 177-182.
desferrioxamine. Free Radic. Biol.
17. Jiang F, Zhou JY, Zhang D, Liu MH,
Med. 1992;13(5): 499-508.
Chen YG. Artesunate induces apoptosis
6. Olanlokun JO, Balogun FA,
and autophagy in HCT116 colon cancer
Olorunsogo OO. Chemotherapeutic and
cells, and autophagy inhibition
prophylactic antimalarial drugs induce
enhances the artesunate-induced
cell death through mitochondrial-
apoptosis. Int J Mol Med. 2018; 42(3):
mediated apoptosis in murine models.
Citation: Lin Y, Song Z, Xi Z, The anticancer Potency of Artemisinin and Its Derivatives, SPR, 2021, 50
Volume 1, issue, 2, Page No.: 43 - 51. DOI: https://doi.org/10.52152/spr/2021.1161295-1304. Biol Ther. 2014;15(11):1479-88.
18. Wang Y, Yang J, Chen L, Wang J, Wang 28. Brew R, Erikson JS, West DC, Kinsella
Y, Luo J, Pan L, Zhang X. Artesunate AR, Slavin J, Christmas SE.
induces apoptosis through caspase- Interleukin-8 as an autocrine growth
dependent and -independent factor for human colon carcinoma cells
mitochondrial pathways in human in vitro. Cytokine. 2000; 12(1): 78-85.
myelodysplastic syndrome SKM-1 29. Hwang SY, Kim JY, Kim KW, Park
cells. Chem Biol Interact. 2014;219:28- MK, Moon Y, Kim WU, Kim HY. IL-
36. 17 induces production of IL-6 and IL-8
19. Jackson SP, Bartek J. The DNA- in rheumatoid arthritis synovial
damage response in human biology and fibroblasts via NF-κB-and PI3-
disease. Nature, 2009; 461(7267): kinase/Akt-dependent pathways.
1071-1078. Arthritis Res Ther. 2004; 6(2): 1-9.
20. Lin L, Lu W, Dai T, Chen H, Wang T, 30. Masoud GN, Li W. HIF-1α pathway:
Yang, L., Yang X, Liu Y, Sun D. Novel role, regulation and intervention for
artemisinin derivatives with potent cancer therapy. Acta Pharm. Sin. B..
anticancer activities and the anti- 2015;5(5): 378-389.
colorectal cancer effect by the 31. Yu C, Sun P, Zhou Y, Shen B, Zhou M,
mitochondria-mediated pathway. Wu L, Kong M. Inhibition of AKT
Bioorg. Chem. 2020;106: 104496. enhances the anti-cancer effects of
21. Lu M, Sun L, Zhou J, Yang J. Artemisinin in clear cell renal cell
Dihydroartemisinin induces apoptosis carcinoma. Biomed. Pharmacother.
in colorectal cancer cells through the 2019; 118: 109383.
mitochondria-dependent pathway. 32. Odaka Y, Xu B, Luo Y, Shen T, Shang
Tumour Biol. 2014;35(6): 5307-5314. C, Wu Y, Zhou H, Huang S. Dihydro-
22. Mao H, Gu H, Qu X, Sun J, Song B, artemisinin inhibits the mammalian
Gao W, Liu J, Shao Q. Involvement of target of rapamycin-mediated signaling
the mitochondrial pathway and pathways in tumor cells. Carcinogene-
Bim/Bcl-2 balance in dihydroartemisin- sis. 2014;35(1):192-200.
in-induced apoptosis in human breast 33. Wei T, Liu J. Anti-angiogenic proper-
cancer in vitro. Int J Mol Med. ties of artemisinin derivatives. Int. J.
2013;31(1):213-8. Mol. Med. 2017; 40(4): 972-978.
23. Greenshields AL, Shepherd TG, 34. Wang SJ, Gao Y, Chen H, Kong R,
Hoskin DW. Contribution of reactive Jiang HC, Pan SH, Xue DB, Bai XW,
oxygen species to ovarian cancer cell Sun B. Dihydroartemisinin inactivates
growth arrest and killing by the anti‐ NF-kappaB and potentiates the anti-
malarial drug artesunate. Mol. tumor effect of gemcitabine on
Carcinog.2017; 56(1): 75-93. pancreatic cancer both in vitro and in
24. Zhang J, Ma G, Guo Z, Yu Q, Han L, vivo. Cancer Lett. 2010;293(1):99-108.
Han M, Zhu Y. Study on the apoptosis 35. Kiani BH, Kayani WK, Khayam AU,
mediated by apoptosis-inducing-factor Dilshad E, Ismail H, Mirza B.
and influencing factors of bovine Artemisinin and its derivatives: A
muscle during postmortem aging. Food promising cancer therapy. Mol. Biol.
Chem. 2018; 266: 359-367. Rep. 2020; 1-16.
25. Susin SA, Lorenzo HK, Zamzami N, 36. Gomes C, Boareto AC, Dalsenter PR.
Marzo I, Snow BE, Brothers GM, Clinical and non-clinical safety of
Mangion J, Jacotot E, Costantini P, artemisinin derivatives in pregnancy.
Loeffler M, Larochette N, Goodlett DR, Reprod. Toxicol. 2016; 65: 194-203.
Aebersold R, Siderovski DP, Penninger 37. Genovese RF, Newman DB. Under-
JM, Kroemer G. Molecular character- standing artemisinin-induced brainstem
ization of mitochondrial apoptosis- neurotoxicity. Arch. Toxicol. 2008;
inducing factor. Nature. 1999;397 82(6): 379-385.
(6718):441-6. 38. Organization, W. H. World malaria
26. Kerbel RS. Tumor angiogenesis. N report 2015: World Health
Engl J Med 2008;358(19): 2039-2049. Organization. 2016; https://www.who.
27. Dong F, Zhou X, Li C, Yan S, Deng X, int/malaria/publications/world-malaria-
Cao Z, Li L, Tang B, Allen TD, Liu J. report-2015/report/en
Dihydroartemisinin targets VEGFR2
via the NF-κB pathway in endothelial
cells to inhibit angiogenesis. Cancer
Citation: Lin Y, Song Z, Xi Z, The anticancer Potency of Artemisinin and Its Derivatives, SPR, 2021, 51
Volume 1, issue, 2, Page No.: 43 - 51. DOI: https://doi.org/10.52152/spr/2021.116You can also read

















































