SOCS: negative regulators of cytokine signaling for immune tolerance
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
International Immunology © The Japanese Society for Immunology. 2021. All rights reserved.
https://doi.org/10.1093/intimm/dxab055 For permissions, please e-mail: journals.permissions@oup.com
Advance Access publication 20 August 2021
SOCS: negative regulators of cytokine signaling for
immune tolerance
Akihiko Yoshimura1, Minako Ito2, Setsuko Mise-Omata1 and Makoto Ando1
Downloaded from https://academic.oup.com/intimm/advance-article/doi/10.1093/intimm/dxab055/6355553 by guest on 01 November 2021
1
Department of Microbiology and Immunology, Keio University School of Medicine, 35 Shinanomachi, Shinjyuku-ku, Tokyo
160-8582, Japan
2
Research Center for Systems Immunology, Medical Institute of Bioregulation Kyushu University, 3-1-1 Maidashi, Higashi-ku,
Fukuoka 812-8582, Japan
REVIEW
Correspondence to: A. Yoshimura; E-mail: yoshimura@keio.jp
Received 21 June 2021, editorial decision 17 August 2021; accepted 18 August 2021
Abstract
Cytokines are important intercellular communication tools for immunity. Many cytokines promote
gene transcription and proliferation through the JAK/STAT (Janus kinase/signal transducers and
activators of transcription) and the Ras/ERK (GDP/GTP-binding rat sarcoma protein/extracellular
signal-regulated kinase) pathways, and these signaling pathways are tightly regulated. The SOCS
(suppressor of cytokine signaling) family members are representative negative regulators of JAK/
STAT-mediated cytokine signaling and regulate the differentiation and function of T cells, thus being
involved in immune tolerance. Human genetic analysis has shown that SOCS family members are
strongly associated with autoimmune diseases, allergy and tumorigenesis. SOCS family proteins
also function as immune-checkpoint molecules that contribute to the unresponsiveness of T cells to
cytokines.
Keywords: anergy, autoimmunity, JAK kinase, regulatory T cells, systemic lupus erythematosus
Introduction
Cytokines of the immune system are soluble molecules that After cloning of the cytokine receptor genes was almost
mediate communication between immune cells or between completed in the 1980s, researchers started to clarify the
immune cells and non-immune cells, and some of them signaling mechanisms of the receptors. First, Src-type tyro-
are called ‘interleukins’. Cytokines with antiviral activity are sine kinases were thought to be involved in the signal trans-
called interferons. The importance of these cytokines in the duction of cytokine receptors. However, it was difficult to show
immune response is clear. Pro-inflammatory cytokines such the binding of the Src-like kinases (~60 kDa) to the receptors.
as tumor necrosis factor α (TNF-α), interleukin 6 (IL-6), IL-12 Instead, we reported that a 130-kDa tyrosine-phosphorylated
and IL-23 are produced mainly by macrophages and den- protein (pp130) binds to the erythropoietin (EPO) receptor in
dritic cells to promote inflammation and activate the ac- 1992 (1). Subsequently, it was demonstrated that the pp130
quired immune system. that bound to the EPO receptor is Janus kinase 2 (JAK2) (2).
T cells of the acquired immune system include CD4+ helper Now four JAK-type tyrosine kinases—JAK1, JAK2, JAK3 and
T (Th) cells and CD8+ cytotoxic T lymphocytes (CTLs). Th Tyk2—are known to non-covalently bind to cytokine recep-
cells are called the ‘command center of immunity’ and re- tors and are activated by receptor oligomerization initiated
lease a variety of T-cell cytokines such as interferon γ (IFN- by cytokine binding (3). For example, JAK1 and Tyk2 bind to
γ), IL-4 and IL-17, which activate B cells, CTLs and cells of IFN-α/β receptors, JAK1 and JAK2 bind to IFN-γ receptors,
the innate immune system. Immune-promoting Th cells are JAK2 binds to the IL-3 receptor β chain and the EPO receptor
referred to as effector T cells and are currently classified into and JAK3 binds to the IL-2 receptor γ chain (4).
four main subsets: Th1, Th2, Th17 and T follicular helper (Tfh) Upon binding of cytokines to their receptors, JAKs are acti-
cells. In addition, the fifth CD4+ T-cell subset comprises regu- vated by phosphorylation of the ‘kinase-activation loop’, which
latory T (Treg) cells, which suppress immune reactions. Treg induces a conformational change that allows the substrate to
cells play a central role in ‘immune tolerance’, a mechanism enter the catalytic pocket of the kinase. Then, activated JAKs
that controls excessive immune responses and immune re- phosphorylate tyrosine residues on the receptor, resulting in
sponses to ‘self’. Treg cells have multiple suppression mech- the recruitment of intracellular signaling molecules that con-
anism including producing anti-inflammatory cytokines such tain modules that recognize phosphotyrosine motifs such
as IL-10 and transforming growth factor β (TGF-β). as the SH2 domain. These molecules are phosphorylated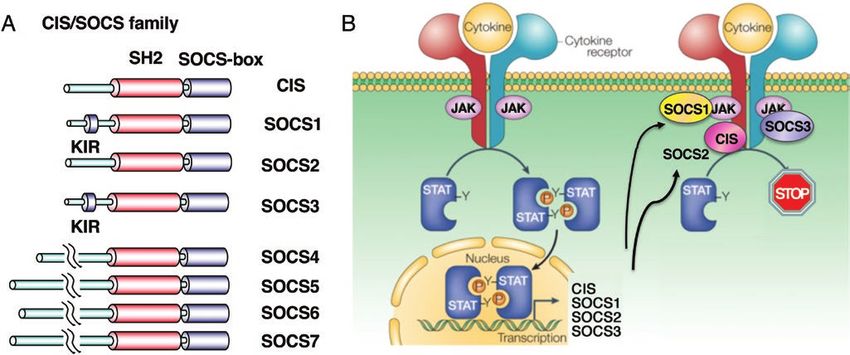
Page 2 of 6 Immune tolerance and tumor immunity by SOCS
by JAKs and are activated, then transmit signals to further family (10). In this family, the SH2 domain and the C-terminal
downstream. Signal transducers and activators of transcrip- SOCS-box are conserved (Fig. 1A).
tion (STAT) family proteins are transcription factors but con- Among the CIS/SOCS family, genes with high specificity for
tain the SH2 domain and C-terminal tyrosine residues that cytokinesis are CIS, SOCS1, SOCS2 and SOCS3. The role of
are phosphorylated by JAKs. STATs are usually present in the the SOCS-box will be described later. Because most SOCS
cytoplasm but move to the nucleus after phosphorylation and family proteins are rapidly induced by cytokines, they are a
dimerization through the SH2 domain. There are six members major negative feedback regulator of cytokine signaling (Fig.
in the STAT family and STATs transmit the information of cor- 1B).
responding cytokines to the nucleus.
Downloaded from https://academic.oup.com/intimm/advance-article/doi/10.1093/intimm/dxab055/6355553 by guest on 01 November 2021
CIS and SOCS2
The cytokine-inducible SH2 protein/suppressor of
The expression of CIS is induced by STAT5 and STAT6, which
cytokine signaling family
are activated by EPO, IL-2, IL-3 and IL-4, and it binds to
Signals from cytokines are needed to mediate effective im- tyrosine-phosphorylated receptors through the SH2 domain.
mune responses, and the negative regulation of cytokine Physical interaction between CIS and the receptor blocks
signaling in immunity is essential for maintaining immune- STAT5 recruitment to the phosphotyrosine residues of the re-
homeostasis and tolerance by suppressing excess immune ceptor and the SOCS-box mediates ubiquitination and deg-
responses. Several proteins have been shown to regulate radation of the receptor (11–13) (Figs 1B and 2A).
tyrosine kinase-associated signaling including SHP-1 tyro- Cis-deficient mice spontaneously develop asthma-like
sine phosphatase and c-Cbl E3 ubiquitin ligase (5). However, symptoms and their T cells have been shown to have increased
these are not specific to cytokine receptors; they are also in- sensitivity to IL-4 (14). Single-nucleotide polymorphisms
volved in negative regulation of receptor tyrosine kinases and (SNPs) in the human CISH gene correlate with susceptibility
Src-related kinase-associated receptors. to a number of infectious diseases, including tuberculosis
We discovered a new family of SH2 domain-containing pro- and malaria (15), and Cis–/– mice also show resistance to tu-
teins, the cytokine-inducible SH2 protein/suppressor of cyto- berculosis infection (16). Genome-wide association studies
kine signaling (CIS/SOCS) family, which are more specific to (GWASs) also revealed that the CISH gene is strongly as-
the JAK–STAT pathway of cytokine receptors. The CIS (gene sociated with the risk of death in sepsis patients (17). CIS
name CISH in humans) was the first gene discovered in this is also important in the regulation of the IL-15 sensitivity of
family that was cloned as a cytokine-inducible gene in 1995 natural killer (NK) cells (18), and CISH–/– NK cells derived
(6). The next discovered gene was SOCS1 [also called JAK- from induced pluripotent stem cells have strong anti-tumor
binding protein (JAB) and STAT-induced STAT inhibitor 1 (SSI- activity. Therefore, CISH could be a therapeutic target for NK
1)], which was reported simultaneously by three independent cell-mediated cancer therapy (19).
groups in 1997 (7–9). We isolated SOCS1, using a yeast two- SOCS2 associates with growth hormone (GH) receptor
hybrid system, as a factor that binds to JAKs and inhibits the and regulates its signaling including STAT5 (20). Therefore,
kinase activity (7). The other two groups discovered it as an SOCS2-deficient mice exhibit gigantism. SOCS2 in dendritic
IL-6-inhibitory molecule and a STAT-inducible SH2 protein, re- cells has shown to be induced by IFN-γ and negatively regu-
spectively. CIS and SOCS1 contain a central SH2 domain and late T-cell activation in human cancer patients (21). However,
similar C-terminal regions, so-called SOCS-box. A database precise molecular mechanism of SOCS2 for cytokine regula-
search revealed that there are currently eight members of this tion remains to be clarified.
Fig. 1. The CIS/SOCS family and their mode of action. (A) Basic structures of the CIS/SOCS family proteins. (B) CIS, SOCS1, SOCS2 and
SOCS3 are induced by STATs. CIS inhibits STAT5 activation by binding to the receptor, SOCS1 directly binds to JAKs and SOCS3 binds to both
the gp130-related receptors and JAKs. SOCS2 also binds to the receptor like CIS but is relatively specific to the GH receptor.Immune tolerance and tumor immunity by SOCS Page 3 of 6
Downloaded from https://academic.oup.com/intimm/advance-article/doi/10.1093/intimm/dxab055/6355553 by guest on 01 November 2021
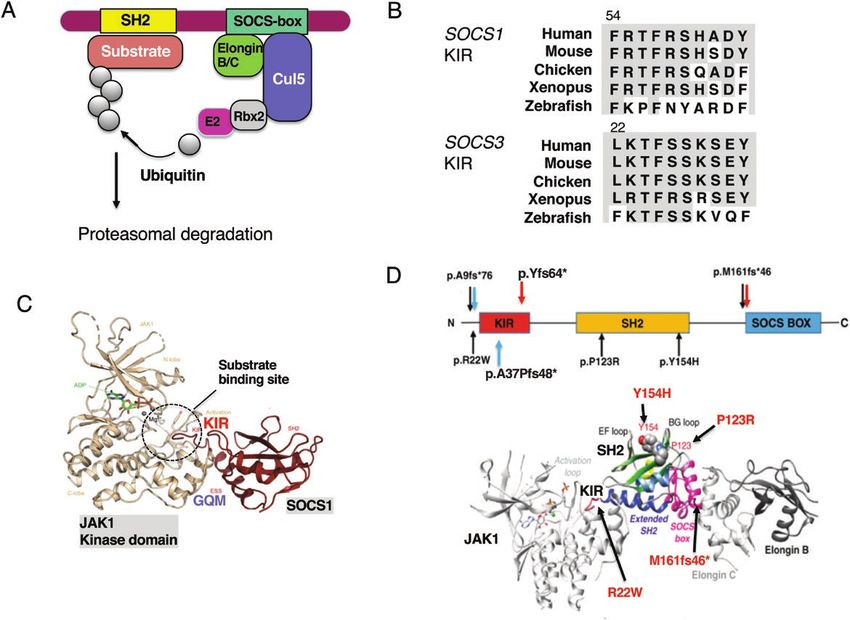
Fig. 2. Molecular mechanisms of suppression by the SOCS-box and KIR. (A) The SOCS-box consists of the BC-box and Cul5-box and recruits
Elongin B–Elongin C heterodimers and Cullin 5 (Cul5). Cul5 also binds Rbx2, which recruits ubiquitin transferase E2. Then ubiquitinated sub-
strates undergo proteasomal degradation. (B) Amino acid sequences of the KIR of SOCS1 and SOCS3 of various species (60). (C) X-ray crys-
tallography of the complex of JAK1 and SOCS1. There are no phosphorylated peptides in the SOCS1 SH2 domain in this structure. Modified
from refs (60–62). (D) SOCS1 structure and mutations found in five families from France (black arrows) (57), two mutations found in the UK (red
arrows) (56) and two mutations in the USA (blue arrows) (58). p.A9fs*76 and p.M161fs*46 were found in two cohorts. The SOCS1 M161Afs*46
mutant leads to disruption of the SOCS-box. The lower panel shows positions of mutations in the complex of JAK1–SOCS1–Elongin B/C. The
amino acids P123 and Y154 are located in the phosphotyrosine peptide-binding groove of the SH2 domain; therefore, mutations P123R and
Y154H probably impair the SH2 domain structure and/or function. Figures are modified from refs (56–58).
Characterization of SOCS1- and SOCS3-deficient mice suggesting a relationship between reduced SOCS1 expres-
and relationship to tumors sion and inflammation-mediated tumor development.
Deletion mutations and functionally defective missense
SOCS1 is highly expressed in thymic T cells as well as acti-
mutations of the SOCS1 gene have been reported in many
vated T cells (22). SOCS1-deficient mice die due to severe
lymphomas, including Burkitt lymphoma and Hodgkin’s
systemic inflammation, including fulminant hepatitis, by 3
lymphoma (28). SOCS1 deficiency is thought to result in the
weeks after birth (22). Since these phenotypes are ameli-
strong activation of JAKs, which contributes to excess cell
orated by an IFN-γ –/– background, and Socs1 expression is
proliferation.
strongly induced by IFN-γ, SOCS1 is a potent negative regu-
SOCS3-deficiecy in mice is prenatally lethal because of
lator of IFN-γ (23). SOCS1 is also involved in the regulation
placental abnormalities caused by abnormal signaling from
of toll-like receptor (TLR) signaling in macrophages. Tissue-
leukemia inhibitory factor (LIF) (29). In addition, analyses of
specific deletion of the Socs1 gene revealed that SOCS1 is
organ-specific knockout or transgenic mice have shown that
fundamentally important for anti-inflammation, homeostasis
SOCS3 plays essential roles in maintaining homeostasis in
and immune tolerance (24). Socs1-deficient mice in which
various tissues, such as heart, brain, liver, fat and joints (30–
SOCS1 expression was restored in T and B cells on a Socs1–
34). Most phenotypes are due to the dysregulated signaling
/–
background spontaneously developed intestinal tumors
of cytokines that activate gp130-related receptors, such
(25), and DNA methylation at the promoter site of the SOCS1
as cardiotrophin-1, leptin and IL-6. In the liver, SOCS3 also
gene is frequently found in human hepatocarcinoma (26, 27),Page 4 of 6 Immune tolerance and tumor immunity by SOCS
regulates insulin-mediated IRS-1 (insulin receptor substrate and hyperglobulinemia within a few months after birth (37).
1) signaling (32). In addition, Th-cell differentiation to Th17, CD4+ T cells in cKO mice are thought to be Th1 or Th2 dom-
which is promoted by IL-6, IL-23 and IL-27, is enhanced by inant depending on their genetic background and environ-
Socs3 deficiency (35–37). In tumors, SOCS3 down-regulation ment. Peripheral T cells in cKO mice mostly showed activated
may contribute to the development of various tumors be- memory types and produced a much higher amount of IFN-γ
cause of IL-6-mediated STAT3 hyperactivation (38, 39). A re- and other cytokines than wild-type T cells did. However, Th17
cent study showed that SOCS3 may be a prognostic factor differentiation was rather suppressed due to Th1 predomin-
and SOCS2 may be a potential therapeutic target in breast ance (37).
cancer (40). These reports indicate that SOCS1 and SOCS3 SOCS1 also plays an important role in suppressing activa-
Downloaded from https://academic.oup.com/intimm/advance-article/doi/10.1093/intimm/dxab055/6355553 by guest on 01 November 2021
are tumor-suppressor genes in humans. tion of CTLs; the deletion of SOCS1 in CD8+ T cells enhanced
T-cell responsiveness and resulted in strong anti-tumor im-
Mechanisms of CIS/SOCS-mediated inhibition of JAKs mune activity (48–50). Genome-wide CRISPR screening
confirmed that SOCS1, Cbl-b and other negative regulators
The SOCS-box is found not only in the CIS/SOCS family but are important for the restriction of human T-cell proliferation
also in many other proteins, including von Hippel–Lindau and activation in response to TCR stimulation (51). Of note,
tumor-suppressor protein, which are E3 ubiquitin ligases. The SOCS1 is considered to be a target of microRNA miR-155,
SOCS-box recruits ubiquitin transferases (Elongin B/C, Rbx2, and forced expression of miR-155 reduced the expression
E2 enzyme complex) and promotes the ubiquitin-dependent level of SOCS1 and enhanced the anti-tumor immunity (52).
proteasomal degradation of molecules, including receptors SOCS1 also plays important roles in the regulation of Treg
and JAKs that associate with the SH2 domain (41) (Fig. 2A). cells (53). SOCS1 is highly expressed in Treg cells, and Treg-
The SH2 domain of SOCS1 binds directly to the phosphor- specific Socs1-deficient mice exhibit symptoms of inflamma-
ylated tyrosine residues of the kinase-activation loop of JAKs, tory diseases, such as dermatitis, hepatitis and splenomegaly
and the SH2 domain of SOCS3 binds with high affinity to the (54, 55). Socs1-deficient Treg cells lose Foxp3 expression
Y759 tyrosine residue of gp130 (see Fig. 1B). In addition to the and convert to Th1-like or Th17-like cells, which produce
SOCS-box-mediated JAK and receptor degradation, SOCS1 IFN-γ and IL-17, respectively, because of STAT 1 and STAT3
and SOCS3 have a kinase inhibitory region (KIR) that, in add- hyperactivation (55).
ition to the SOCS-box, directly suppresses the kinase activity
of JAKs. KIR is highly conserved among various species (Fig.
2B). We proposed an inhibitory mechanism in which the KIR Human SOCS1 mutations and autoimmune diseases
acts as a pseudosubstrate and inhibits the binding of JAK GWASs have shown that SOCS1 SNPs are found in a variety
substrates (42, 43) (Fig. 2C). The non-canonical surface of of immune diseases, including primary biliary cirrhosis, mul-
the SH2 domain that opposes the phosphopeptide-binding tiple sclerosis, Crohn’s disease and celiac disease. However,
surface and the N-terminal extended SH2 domain (ESS) of inherited mutations of the SOCS1 gene in human auto-
SOCS1 and SOCS3 bind to the GQM motif of JAKs. Since immunity have just recently been reported in 2020. SOCS1
JAK3 does not have a GQM motif, SOCS1 and SOCS3 do haploinsufficiency leads to early onset autoimmune diseases
not inhibit the kinase activity of JAK3. This pseudosubstrate associated with the cytokine hypersensitivity of immune cells.
mechanism of inhibition is demonstrated by the analysis of In other words, reducing the expression of SOCS1 by half
the co-crystal structure of JAKs and SOCS1 or SOCS3 (41, induces autoimmune diseases, indicating that SOCS1 is an
42, 44) (Fig. 2C). important gene for the maintenance of immune tolerance
Since SOCS3 binds to the gp130-related cytokine recep- in humans.
tors, SOCS3 is relatively specific to STAT3 inhibition. In add- First, the whole-genome sequencing of large sporadic (or
ition, an important function of SOCS3 is to distinguish the non-familial) primary immunodeficiency disease patients in
pro-inflammatory and anti-inflammatory effects of IL-6 and the UK discovered two heterozygous mutations (56) (Fig.
IL-10 (45). In macrophages, SOCS3 is induced by IL-6 and 2D). Patients with p.Met161Alafs*46 and p.Tyr64* mutations
inflammatory signals including TLRs and then associates with showed decreased B cells, increased Th1 cells and de-
gp130; therefore, IL-6-mediated STAT3 activation is weak and creased Treg cell numbers. Patients with p.Met161Alafs*46
transient. In contrast, although SOCS3 is also induced by mutation also had lung and liver inflammation. The SOCS1
IL-10, it does not bind to the IL-10 receptor; therefore, STAT3 M161Afs*46 mutation disrupts the function of the SOCS-box.
activation is strong and sustained. Hyperactivation of STAT3 T cells derived from patients with these SOCS1 mutations
suppresses NF-kB and TLR signaling, thus limiting the induc- showed reduced levels of SOCS1 protein and increased IFN-
tion of inflammatory responses such as induction of TNF-α γ-induced phosphorylation of STAT1.
and IL-12. When a mutant gp130 that cannot associate with Next, heterozygous germline mutations in SOCS1 were re-
SOCS3 is expressed in macrophages, IL-6 behaves like ported in 10 patients with early onset autoimmune disease
IL-10, an anti-inflammatory cytokine. This model has been in five families from France (57) (Fig. 2D). The mutations
proven to be correct in a number of follow-up studies (46, 47). are amino acid substitutions (c.368 C > G, p.P123R, c.64
C > T, p.R22W, c.460 T > C, p.Y154H) and deletions, all
of which were considered loss-of-function. The two amino
SOCS1 and tolerance of effector T cells and Treg cells
acids (P123 and Y154) are located in the phosphotyrosine
T-cell-specific Socs1 conditional knockout (cKO) mice devel- peptide-binding groove of the SH2 domain; therefore, muta-
oped autoimmune diseases such as dermatitis, splenomegaly tions P123R and Y154H impair the SH2 domain structure and/Immune tolerance and tumor immunity by SOCS Page 5 of 6
or function. Symptoms are early onset autoimmune disease; 3 Schwartz, D. M., Bonelli, M., Gadina, M. et al. 2016. Type I/II
60% of cases occur under 10 years of age and include im- cytokines, JAKs, and new strategies for treating autoimmune dis-
eases. Nat. Rev. Rheumatol. 12:25.
mune thrombocytopenic purpura, psoriasis, celiac disease, 4 O’Shea, J. J., Gadina, M. and Schreiber, R. D. 2002. Cytokine
systemic lupus erythematosus, thyroiditis and hepatitis. signaling in 2002: new surprises in the Jak/Stat pathway. Cell
Patients have higher levels of cytokines in the blood, similar 109(Suppl.):S121.
to patients carrying gain-of-function mutations in STAT1 5 Yokouchi, M., Kondo, T., Sanjay, A. et al. 2001. Src-catalyzed
and STAT3. Some patients developed splenomegaly and phosphorylation of c-Cbl leads to the interdependent ubiquitin-
ation of both proteins. J. Biol. Chem. 276:35185.
Hodgkin’s lymphoma. Heterozygous lymphocytes with the 6 Yoshimura, A., Ohkubo, T., Kiguchi, T. et al. 1995. A novel
mutation are more sensitive to cytokines such as IFN-γ, IL-2 cytokine-inducible gene CIS encodes an SH2-containing protein
Downloaded from https://academic.oup.com/intimm/advance-article/doi/10.1093/intimm/dxab055/6355553 by guest on 01 November 2021
and IL-4, and Treg functions are impaired (57). Hyper STAT that binds to tyrosine-phosphorylated interleukin 3 and erythro-
activation in these cells with SOCS1 mutations was restored poietin receptors. EMBO J. 14:2816.
7 Endo, T. A., Masuhara, M., Yokouchi, M. et al. 1997. A new protein
by the JAK1/JAK2 inhibitor ruxolitinib. T-cell activation was containing an SH2 domain that inhibits JAK kinases. Nature
also increased in ~30% of asymptomatic carriers. 387:921.
In addition, one sporadic case and one familial deletion 8 Naka, T., Narazaki, M., Hirata, M. et al. 1997. Structure and func-
of SOCS1 have been reported in multisystem inflammatory tion of a new STAT-induced STAT inhibitor. Nature 387:924.
syndrome in children (MIS-C) (58) (Fig. 2D). One patient de- 9 Starr, R., Willson, T. A., Viney, E. M. et al. 1997. A family of
cytokine-inducible inhibitors of signalling. Nature 387:917.
veloped a severe acute respiratory syndrome (SARS) after 10 Masuhara, M., Sakamoto, H., Matsumoto, A. et al. 1997. Cloning
SARS coronavirus 2 (SARS-CoV-2) infection, suggesting that and characterization of novel CIS family genes. Biochem.
SOCS1 deficiency may be related to the severity or prog- Biophys. Res. Commun. 239:439.
nostic symptoms of coronavirus disease 2019 (COVID-19). 11 Matsumoto, A., Masuhara, M., Mitsui, K. et al. 1997. CIS, a cyto-
kine inducible SH2 protein, is a target of the JAK-STAT5 pathway
and modulates STAT5 activation. Blood 89:3148.
Conclusion 12 Matsumoto, A., Seki, Y., Kubo, M. et al. 1999. Suppression of
STAT5 functions in liver, mammary glands, and T cells in cytokine-
For the past 30 years, we have been working to elucidate the inducible SH2-containing protein 1 transgenic mice. Mol. Cell.
molecular mechanisms of the regulation of cytokine signaling Biol. 19:6396.
by the CIS/SOCS family and their physiological and patho- 13 Verdier, F., Chrétien, S., Muller, O. et al. 1998. Proteasomes regu-
logical significance. The molecular mechanism of suppres- late erythropoietin receptor and signal transducer and activator
of transcription 5 (STAT5) activation. Possible involvement of the
sion has been demonstrated at the atomic level by X-ray ubiquitinated Cis protein. J. Biol. Chem. 273:28185.
crystallography, and SOCS1 has been confirmed as an im- 14 Yang, X. O., Zhang, H., Kim, B. S. et al. 2013. The signaling sup-
mune tolerance-related gene by the discovery of human mu- pressor CIS controls proallergic T cell development and allergic
tations inherited by families. We have shown that SOCS is airway inflammation. Nat. Immunol. 14:732.
15 Khor, C. C., Vannberg, F. O., Chapman, S. J. et al. 2010. CISH and
necessary for immune regulation and that SOCS1 also acts in susceptibility to infectious diseases. N. Engl. J. Med. 362:2092.
immune-checkpoints in tumor immunity (48, 59). We will con- 16 Queval, C. J., Song, O. R., Carralot, J. P. et al. 2017.
tinue our research in pursuit of the application of SOCS to the Mycobacterium tuberculosis controls phagosomal acidification
treatment of autoimmune diseases and cancers in humans. by targeting CISH-mediated signaling. Cell Rep. 20:3188.
17 Rosier, F., Brisebarre, A., Dupuis, C. et al. 2021. Genetic predis-
position to the mortality in septic shock patients: from GWAS to
Funding the identification of a regulatory variant modulating the activity of
a CISH enhancer. Int. J. Mol. Sci. 22:5852.
18 Delconte, R. B., Kolesnik, T. B., Dagley, L. F. et al. 2016. CIS is
This work was supported by JSPS KAKENHI 17H06175, 21H05044, a potent checkpoint in NK cell-mediated tumor immunity. Nat.
19H04817, 21K19382, 21H02719, 21H00432, AMED-CREST Immunol. 17:816.
21gm1110009, AMED-PRIME 21gm6210012, the Princess 19 Zhu, H., Blum, R. H., Bernareggi, D. et al. 2020. Metabolic repro-
Takamatsu Cancer Research Fund, the Yasuda Medical Foundation, graming via deletion of CISH in human iPSC-derived NK cells
the Kishimoto Family Foundation, the Tomizawa Jun-ichi & Keiko promotes in vivo persistence and enhances anti-tumor activity.
Fund of Molecular Biology Society of Japan for Young Scientist, the Cell Stem Cell 27:224.
Mitsubishi Foundation, the Mochida Memorial Foundation for Medical 20 Letellier, E. and Haan, S. 2016. SOCS2: physiological and patho-
and Pharmaceutical Research, the Takeda Science Foundation, logical functions. Front. Biosci. (Elite Ed.) 8:189.
the Uehara Memorial Foundation, the Naito Foundation, the Kanae 21 Nirschl, C. J., Suárez-Fariñas, M., Izar, B. et al. 2017. IFNγ-
Foundation, the SENSHIN Medical Research Foundation, the Astellas dependent tissue-immune homeostasis is co-opted in the tumor
Foundation for Research on Metabolic Disorders, the Inoue Research microenvironment. Cell 170:127.
Award for Young Scientists, a Life Science Research Award and Keio 22 Marine, J. C., Topham, D. J., McKay, C. et al. 1999. SOCS1 de-
Gijuku Academic Developmental Funds. ficiency causes a lymphocyte-dependent perinatal lethality. Cell
Conflicts of interest statement: the authors declared no conflicts of 98:609.
interest. 23 Sakamoto, H., Kinjyo, I. and Yoshimura, A. 2000. The Janus kinase
inhibitor, Jab/SOCS-1, is an interferon-gamma inducible gene
and determines the sensitivity to interferons. Leuk. Lymphoma
38:49.
References 24 Yoshimura, A., Naka, T. and Kubo, M. 2007. SOCS proteins, cyto-
1 Yoshimura, A. and Lodish, H. F. 1992. In vitro phosphorylation kine signalling and immune regulation. Nat. Rev. Immunol. 7:454.
of the erythropoietin receptor and an associated protein, pp130. 25 Hanada, T., Kobayashi, T., Chinen, T. et al. 2006. IFNgamma-
Mol. Cell. Biol. 12:706. dependent, spontaneous development of colorectal carcinomas
2 Witthuhn, B. A., Quelle, F. W., Silvennoinen, O. et al. 1993. JAK2 in SOCS1-deficient mice. J. Exp. Med. 203:1391.
associates with the erythropoietin receptor and is tyrosine phos- 26 Yoshida, T., Ogata, H., Kamio, M. et al. 2004. SOCS1 is a sup-
phorylated and activated following stimulation with erythropoietin. pressor of liver fibrosis and hepatitis-induced carcinogenesis. J.
Cell 74:227. Exp. Med. 199:1701.Page 6 of 6 Immune tolerance and tumor immunity by SOCS
27 Yoshikawa, H., Matsubara, K., Qian, G. S. et al. 2001. SOCS- 44 Yoshimura, A., Mori, H., Ohishi, M. et al. 2003. Negative regu-
1, a negative regulator of the JAK/STAT pathway, is silenced lation of cytokine signaling influences inflammation. Curr. Opin.
by methylation in human hepatocellular carcinoma and shows Immunol. 15:704.
growth-suppression activity. Nat. Genet. 28:29. 45 Yasukawa, H., Ohishi, M., Mori, H. et al. 2003. IL-6 induces an
28 Lennerz, J. K., Hoffmann, K., Bubolz, A. M. et al. 2015. Suppressor anti-inflammatory response in the absence of SOCS3 in macro-
of cytokine signaling 1 gene mutation status as a prog- phages. Nat. Immunol. 4:551.
nostic biomarker in classical Hodgkin lymphoma. Oncotarget 46 El Kasmi, K. C., Holst, J., Coffre, M. et al. 2006. General nature
6:29097. of the STAT3-activated anti-inflammatory response. J. Immunol.
29 Takahashi, Y., Carpino, N., Cross, J. C. et al. 2003. SOCS3: an 177:7880.
essential regulator of LIF receptor signaling in trophoblast giant 47 Fowler, K. D., Kuchroo, V. K. and Chakraborty, A. K. 2012. A
cell differentiation. EMBO J. 22:372. model for how signal duration can determine distinct outcomes
Downloaded from https://academic.oup.com/intimm/advance-article/doi/10.1093/intimm/dxab055/6355553 by guest on 01 November 2021
30 Hamanaka, I., Saito, Y., Yasukawa, H. et al. 2001. Induction of gene transcription programs. PLoS One 7:e33018.
of JAB/SOCS-1/SSI-1 and CIS3/SOCS-3/SSI-3 is involved in 48 Chikuma, S., Kanamori, M., Mise-Omata, S. et al. 2017.
gp130 resistance in cardiovascular system in rat treated with Suppressors of cytokine signaling: potential immune checkpoint
cardiotrophin-1 in vivo. Circ. Res. 88:727. molecules for cancer immunotherapy. Cancer Sci. 108:574.
31 Shouda, T., Yoshida, T., Hanada, T. et al. 2001. Induction of the 49 Lawson, K. A., Sousa, C. M., Zhang, X. et al. 2020. Functional
cytokine signal regulator SOCS3/CIS3 as a therapeutic strategy genomic landscape of cancer-intrinsic evasion of killing by T
for treating inflammatory arthritis. J. Clin. Invest. 108:1781. cells. Nature 586:120.
32 Torisu, T., Sato, N., Yoshiga, D. et al. 2007. The dual function 50 Wei, J., Long, L., Zheng, W. et al. 2019. Targeting REGNASE-1
of hepatic SOCS3 in insulin resistance in vivo. Genes Cells programs long-lived effector T cells for cancer therapy. Nature
12:143. 576:471.
33 Yasukawa, H., Hoshijima, M., Gu, Y. et al. 2001. Suppressor of 51 Shifrut, E., Carnevale, J., Tobin, V. et al. 2018. Genome-wide
cytokine signaling-3 is a biomechanical stress-inducible gene CRISPR screens in primary human T cells reveal key regulators of
that suppresses gp130-mediated cardiac myocyte hypertrophy immune function. Cell 175:1958.
and survival pathways. J. Clin. Invest. 108:1459. 52 Ji, Y., Wrzesinski, C., Yu, Z. et al. 2015. miR-155 augments CD8+
34 Seki, Y., Kai, H., Shibata, R. et al. 2000. Role of the JAK/STAT T-cell antitumor activity in lymphoreplete hosts by enhancing re-
pathway in rat carotid artery remodeling after vascular injury. sponsiveness to homeostatic γc cytokines. Proc. Natl Acad. Sci.
Circ. Res. 87:12. USA 112:476.
35 Stumhofer, J. S., Laurence, A., Wilson, E. H. et al. 2006. Interleukin 53 Takahashi, R. and Yoshimura, A. 2014. SOCS1 and regulation of
27 negatively regulates the development of interleukin 17-produ- regulatory T cells plasticity. J. Immunol. Res. 2014:943149.
cing T helper cells during chronic inflammation of the central ner- 54 Lu, L. F., Thai, T. H., Calado, D. P. et al. 2009. Foxp3-dependent
vous system. Nat. Immunol. 7:937. microRNA155 confers competitive fitness to regulatory T cells by
36 Taleb, S., Romain, M., Ramkhelawon, B. et al. 2009. Loss of targeting SOCS1 protein. Immunity 30:80.
SOCS3 expression in T cells reveals a regulatory role for inter- 55 Takahashi, R., Nishimoto, S., Muto, G. et al. 2011. SOCS1 is es-
leukin-17 in atherosclerosis. J. Exp. Med. 206:2067. sential for regulatory T cell functions by preventing loss of Foxp3
37 Tanaka, K., Ichiyama, K., Hashimoto, M. et al. 2008. Loss of expression as well as IFN-gamma and IL-17A production. J. Exp.
suppressor of cytokine signaling 1 in helper T cells leads Med. 208:2055.
to defective Th17 differentiation by enhancing antagonistic 56 Thaventhiran, J. E. D., Lango Allen, H., Burren, O. S. et al.;
effects of IFN-gamma on STAT3 and Smads. J. Immunol. Primary Immunodeficiency Consortium for the NIHR Bioresource.
180:3746. 2020. Whole-genome sequencing of a sporadic primary immuno-
38 Ogata, H., Kobayashi, T., Chinen, T. et al. 2006. Deletion of the deficiency cohort. Nature 583:90.
SOCS3 gene in liver parenchymal cells promotes hepatitis- 57 Hadjadj, J., Castro, C. N., Tusseau, M. et al. 2020. Early-onset
induced hepatocarcinogenesis. Gastroenterology 131:179. autoimmunity associated with SOCS1 haploinsufficiency. Nat.
39 Lesina, M., Kurkowski, M. U., Ludes, K. et al. 2011. Stat3/Socs3 Commun. 11:5341.
activation by IL-6 transsignaling promotes progression of pan- 58 Lee, P. Y., Platt, C. D., Weeks, S. et al. 2020. Immune dysregulation
creatic intraepithelial neoplasia and development of pancreatic and multisystem inflammatory syndrome in children (MIS-C)
cancer. Cancer Cell 19:456. in individuals with haploinsufficiency of SOCS1. J. Allergy Clin.
40 Sun, M., Tang, C., Liu, J. et al. 2021. Comprehensive analysis Immunol. 146:1194.
of suppressor of cytokine signaling proteins in human breast 59 Hibino, S., Chikuma, S., Kondo, T. et al. 2018. Inhibition of Nr4a re-
cancer. BMC Cancer 21:696. ceptors enhances antitumor immunity by breaking Treg-mediated
41 Yoshimura, A., Nishinakamura, H., Matsumura, Y. et al. 2005. immune tolerance. Cancer Res. 78:3027.
Negative regulation of cytokine signaling and immune responses 60 Babon, J. J., Kershaw, N. J., Murphy, J. M. et al. 2012. Suppression
by SOCS proteins. Arthritis Res. Ther. 7:100. of cytokine signaling by SOCS3: characterization of the mode of
42 Yasukawa, H., Misawa, H., Sakamoto, H. et al. 1999. The JAK- inhibition and the basis of its specificity. Immunity 36:239.
binding protein JAB inhibits Janus tyrosine kinase activity through 61 Liau, N. P. D., Laktyushin, A., Lucet, I. S. et al. 2018. The mo-
binding in the activation loop. EMBO J. 18:1309. lecular basis of JAK/STAT inhibition by SOCS1. Nat. Commun.
43 Sasaki, A., Yasukawa, H., Suzuki, A. et al. 1999. Cytokine- 9:1558.
inducible SH2 protein-3 (CIS3/SOCS3) inhibits Janus tyrosine 62 Kershaw, N. J., Murphy, J. M., Liau, N. P. et al. 2013. SOCS3 binds
kinase by binding through the N-terminal kinase inhibitory region specific receptor-JAK complexes to control cytokine signaling by
as well as SH2 domain. Genes Cells 4:339. direct kinase inhibition. Nat. Struct. Mol. Biol. 20:469.You can also read

















































