Original Article Myeloid-derived suppressor cells promote the stem cell-like acquisition of lung adenocarcinoma through the Wnt/β-catenin pathway
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Int J Clin Exp Med 2020;13(8):5823-5830
www.ijcem.com /ISSN:1940-5901/IJCEM0111447
Original Article
Myeloid-derived suppressor cells promote the stem
cell-like acquisition of lung adenocarcinoma
through the Wnt/β-catenin pathway
Yongshu Xu1, Qiong Feng4, Jinyan Chen2, Wenjun Xu2, Jian Sun2, Huan Deng3,5
Departments of 1General Practice, 2Respiration, 3Pathology, The Fourth Affiliated Hospital of Nanchang University,
Nanchang, China; 4Department of Pathology, The Second Affiliated Hospital of Nanchang University, Nanchang,
China; 5Tumor Immunology Institute, Nanchang University, Nanchang, China
Received March 26, 2020; Accepted June 2, 2020; Epub August 15, 2020; Published August 30, 2020
Abstract: Heterogeneity remains the biggest obstacle in deepening the understanding of lung adenocarcinoma
(LAC) biology. The phenotype and functions of cancer cells may change to counteract the threat from the host.
However, the exact mechanisms by which myeloid-derived suppressor cells (MDSCs) contribute to the acquisition
of stem cell-like properties in lung adenocarcinoma are largely unknown. Pathological archives of 256 patients with
lung adenocarcinoma at early stages were collected and evaluated by immunohistochemistry and FACS to explore
the infiltration of MDSCs and the expression of stem cell marker p63. To further provide insights into the molecular
mechanisms and interactions between MDSCs and p63-expressing cancer cells, we established early LAC models
using histidine decarboxylase (Hdc)-CreERT2; eGFP; LSL-KrasG12D transgenic mice. MDSC-derived Wnts were blocked
by porcupineflox/flox and diphtheria toxin receptor (DTR). Patients with a distinct population of LAC cells at early stages,
indicated by p63 immunopositivity, exhibited a significantly poorer prognosis than their p63- negative counterparts.
The percentage of Hdc+ polymorphonuclear (PMN) MDSCs increased in direct proportion to the p63+ rate. Hdc+
PMN-MDSCs secreted high levels of Wnt1, 2, 3, and 8a to aberrantly activate Wnt/β-catenin in the majority of
p63+ cancer cells but not p63- cells. Blockade of Hdc+ PMN-MDSC-derived Wnts, or ablation of Hdc+ PMN-MDSCs
in transgenic mouse models decreased the LAC tumor burden and the p63+ rate. In the microenvironment of LAC,
PMN-MDSCs infiltrate and expand to promote p63 expression in cancer cells through upregulating Wnts. Thus, the
tailored immunotherapies targeting Hdc+ PMN-MDSCs might be of therapeutic benefit for LAC patients.
Keywords: Lung adenocarcinoma, p63, cancer stem cell, myeloid-derived suppressor cell, histamine decarboxyl-
ase
Introduction ease progression is still debated. Upregulated
p63 contributes to cell cycle arrest and apop-
Lung cancer (LC) is the leading cause of cancer- tosis [4, 5]. Genomic sequencing of p63 is
related mortality worldwide [1, 2]. Up to 46% of amplified in the majority of lung squamous cell
adenocarcinoma patients develop recurrence carcinomas (SCCs) and endows patients with
or distant metastasis within five years [3]. Ac- prolonged survival [6]. Substantial evidence
cumulating evidence provides more insights also supports that p63 can function as an
into the heterogeneity of cancer and suggests oncogene. The dysregulation of p63 is invol-
that different gene expression signatures of ved in the progression of preneoplastic lesions
prognosis can be observed in separate regions and lung adenocarcinoma (LAC) [7, 8]. Thus, it
of the same tumor. However, the exact identity seems that the crosstalk between p63 and
of cancer stem cells (CSCs) is still controversial, other risk factors can influence neoplastic
mainly because of the lack of unique markers. progression.
P63, a member of the p53 protein family, is Several pathways have been linked to stem
consistently expressed by basal/somatic stem cell phenotypes in which p63 is involved. Ac-
cells. The role of p63 in tumorigenesis and dis- tivation of canonical Wnt signaling promoted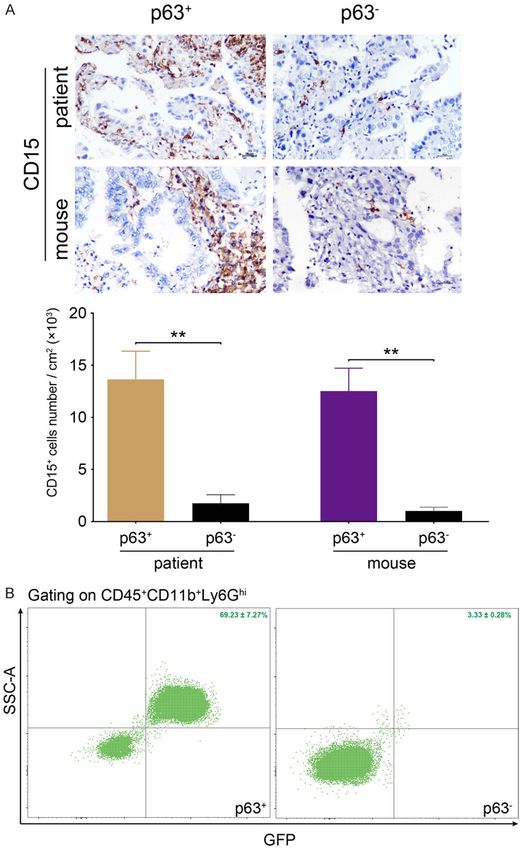
MDSCs and stem cell-like acquisition of lung adenocarcinoma
the expansion of bronchioalveolar stem cells eGFP mice were purchased from EMMA. LSL-
[9]. The downregulation of endogenous p63 KrasG12D, porcupineflox/flox, and iDTR mice were
enhanced the expression of Wnt-related genes obtained from the Jackson Laboratory. LSL-
[10]. However, cellular sources of Wnt ligands KrasG12D mice were crossed to Hdc-CreERT2;
are still unclear. An increased number of poly- eGFP followed by the administration of high
morphonuclear MDSCs (PMN-MDSCs) were ob- doses (5 × 108 PFU) of AdenoCre (Cell Biolabs,
served in patients with NSCLC compared with USA) by intranasal instillation at 6 weeks of
the general population [11]. The expansion of age as previously described, establishing early
PMN-MDSCs has been linked to a relatively LAC models [14]. To explore the role of Hdc+
poor outcome in NSCLC patients at late sta-
PMN-MDSC-derived Wnts, Hdc-CreERT2; eGFP;
ges, suggesting their pro-cancerous effects
LSL-KrasG12D mice were crossed with porcu-
[12]. They may serve as the cellular source of
pineflox/flox and iDTR, respectively. All mice were
tumor-associated immature myeloid cells in
the context of aging or tumors. Previous study backcrossed and maintained on the C57BL/6
suggests that Myeloid-derived Wnts play pivot- background and housed in the special patho-
al roles in the regeneration and differentiation gen-free animal facilities.
of intestinal stem cells [13].
Histopathological and immunohistochemical
In the current study, we explored the prognos- staining
tic role and regulatory mechanisms of p63.
Clinical data showed that a minority of pati- Formalin-fixed and paraffin embedded blocks
ents with early LAC displayed immunopositivity were embedded by the tissue arrayer TMAjr
for p63 and exhibited a tendency towards (Pathology Devices). Serial sections were cut
recurrence or metastasis. Hdc+ PMN-MDSCs into 3- to 4-μm slides followed by H&E and
were recruited and infiltrated around p63+ can- immunohistochemical staining. The histopath-
cer cells. The upregulation of canonical Wnts ological evaluation of all sections was per-
in Hdc+ PMN-MDSCs aberrantly activated Wnt/ formed by two independent pathologists (L
β-catenin signaling in cancer cells and pro- Zhou and J Hu), who were blinded regarding
moted p63 expression. Our data further unveil details. Primary antibodies for p63 (1:200;
interactions between Hdc+ PMN-MDSCs and ab53039, Abcam), CD15 (1:150; clone Carb-1,
LAC cells, raising the possibility that the inhibi- Novocastra, Leica), anti-GFP (1:200; ab13970,
tion or depletion of these cells with targeted
Abcam), and β-catenin (1:200; ab32572, Ab-
strategies might facilitate anti-lung cancer
cam) were used (Autostainer 360, Lab Vision,
treatment.
Thermo Fisher). A microscope (80i, Nikon) with
Materials and methods a CCD camera (DS-Ri2, Nikon) acquired the
images. Counting of positive cells was con-
Study cohort ducted in nonoverlapping fields using the 40 ×
objective. The average number of positive cells
This study included 256 patients with well-dif- in each square centimeter was calculated for
ferentiated LAC. Lymph node or secondary site each specimen.
metastasis was excluded by computed tomog-
raphy (CT) or ultrasonographic scanning. Achi- In vivo treatment
eves of tumor tissues were collected and eva-
luated. All patients gave written informed con- At 6 weeks of age, tamoxifen (Harlan Labo-
sent to participate in the study in accordance ratories) chow was fed to the mice in order to
with the Helsinki Declaration, and this study delete porpupine in Hdc+ myeloid cells, result-
was approved by the Medical Ethics Commit- ing in Wnts secretion blockade. To further
tee of The Second and Fourth Affiliated Hospi- exclude other influencing factors, intraperito-
tal of Nanchang University.
neal diphtheria toxin (DT, Sigma) injections
Mouse models combined with a tamoxifen chow were used to
abolish Hdc+ myeloid cell-derived Wnts in Hdc-
All animal experiment protocols were appro- CreERT2; eGFP; LSL-KrasG12D; iDTR and Hdc-
ved by the Nanchang University Institutional CreERT2; eGFP; LSL-KrasG12D; porcupineflox/flox;
Animal Care and Use Committee. Hdc-CreERT2; iDTR mice.
5824 Int J Clin Exp Med 2020;13(8):5823-5830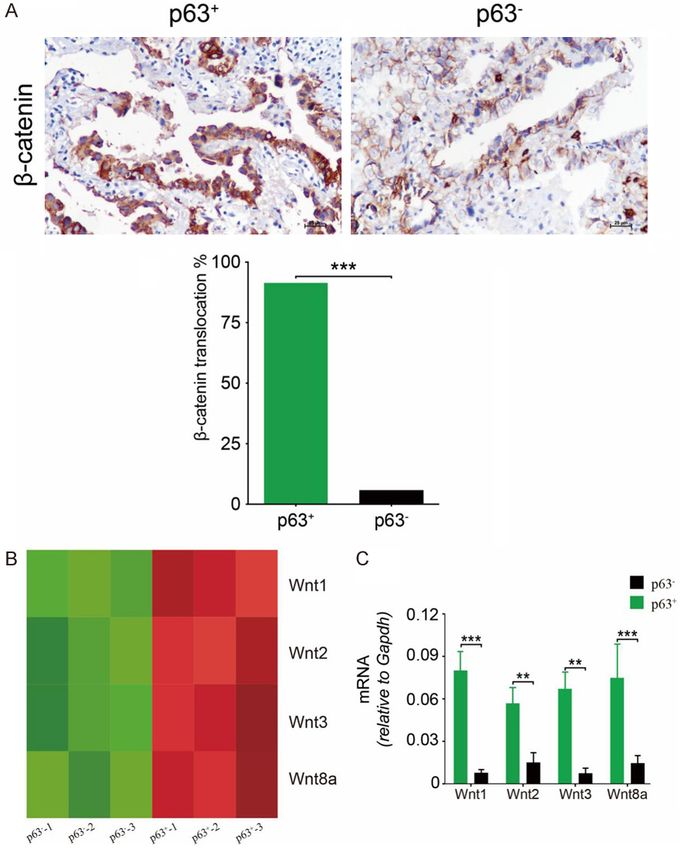
MDSCs and stem cell-like acquisition of lung adenocarcinoma
Table 1. Primers for quantitative RT-PCR
Gene
Organism Gene Name Forward Primer (5’-3’) Reverse Primer (5’-3’)
Symbol
Wnt1 Mus musculus wingless-type MMTV integration site family, member 1 TACCTACTAGCGTACTGATAC ACTGTCTAGTCTGGCGACTAG
Wnt2 Mus musculus wingless-type MMTV integration site family, member 2 CACCTCTCTTCCTGCTGATTT TCATTCTGCTTTCCATCCCTAC
Wnt3 Mus musculus wingless-type MMTV integration site family, member 3 GATGGTACTGGCTGGGTTTAG GCTCATGGTCACATGGTAGAG
Wnt8a Mus musculus wingless-type MMTV integration site family, member 8a TGCAGGACAGTGGAGATAGA CCACAAGCCAGGAGAACAA
Gapdh Mus musculus glyceraldehyde-3-phosphate dehydrogenase CTTTGTCAAGCTCATTTCCTGG TCTTGCTCAGTGTCCTTGC
incubated with the antibody
panel composed of CD45
(30-F11, eBioscience), CD11b
(M1/70, eBioscience), and Ly-
6G (1A8, eBioscience). PMN-
MDSCs were identified based
on their phenotype: CD45+
CD11b+Ly6Ghi. Hdc+ cells were
characterized by their high le-
vel of GFP expression (GFPhi).
CD45+CD11b+Ly6GhiGFPlo cells
harvested from eGFP wild-
type littermates were used to
set the gate. Stained cells we-
re fixed with Cytofix (BD Bio-
science) for 30 min on ice and
analyzed by an LSR II flow cy-
tometer (BD Bioscience).
RNA-seq analysis
Hdc+ PMN-MDSCs were sorted
and lysed in an ARCTURUS
PicoPure RNA isolation kit
according to the manufactur-
er’s instructions (Life Techno-
logies). Total RNA was isolated
followed by cDNA amplifica-
tion. Libraries were establish-
ed using the SMARTer Ultra
Low Input RNA kit (Clontech
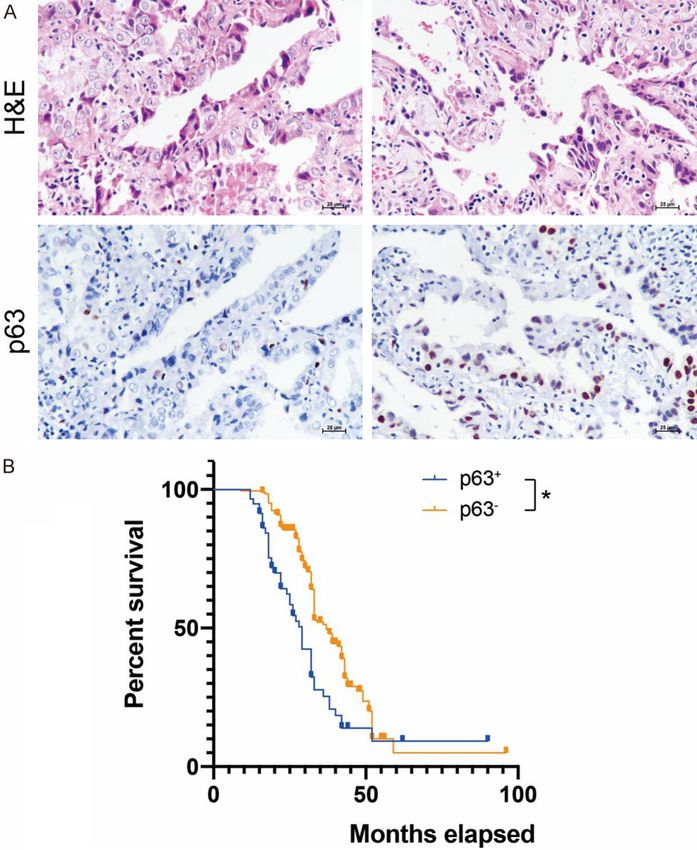
Figure 1. P63 expression in lung adenocarcinoma (LAC). (A) A minority of Laboratories) and the Nextera
LAC at early stages were positive for p63 antibody. (B) P63 positivity identi- XT DNA Library Preparation Kit
fied a distinct subpopulation of LAC characterized by a poorer prognosis
(Illumina). Sequencing was per-
compared with p63- counterparts. Original magnification × 400 (A). *P <
0.05 (B). formed on a HiSeq 2500
(Illumina).
Flow cytometry analysis Gene microarray analysis
Fresh tissues obtained from lung primary tu- Both Hdc+ and Hdc- PMN-MDSCs were harve-
mors were manually minced and incubated in sted from LAC tissues. Total mRNA was extra-
DMEM with collagenase A (Roche) and DNase cted using a RNeasy Micro Kit (Qiagen) and la-
I (Roche) for 45 min at 37°C. Suspensions beled with a 3’IVT Expression Kit (Affymetrix)
were filtered three times using a 70 μm nylon before hybridization to the Affymetrix Gene-
mesh to remove dead cell debris and enrich Chip mouse genome 430 2.0 array (Affymetrix).
leucocytes. No more than 1 × 106 cells were Arrays were performed using an Affymetrix
5825 Int J Clin Exp Med 2020;13(8):5823-5830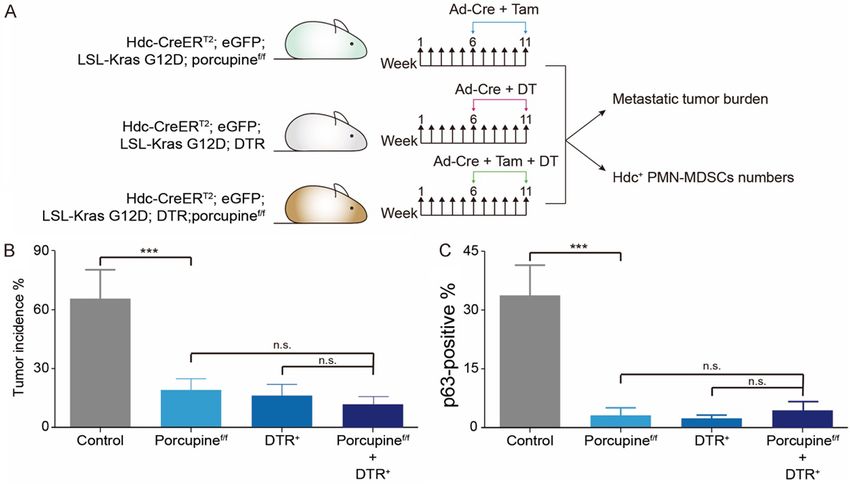
MDSCs and stem cell-like acquisition of lung adenocarcinoma
reverse transcription using a
SuperScript III First-Strand
Synthesis System (Life Tech-
nologies). The PrimerQuest
Tool (Integrated DNA Tech-
nologies) was used to design
sequences of SYBR Green for
Wnt1, Wnt2, Wnt3, and Wnt8a
(Table 1). Quantitative PCR
was performed with the Step-
One Plus instrument (Appli-
ed Biosystems). Relative gene
expression was normalized to
Gapdh.
Statistical analysis
Experimental results were rep-
licated at least once, unless
otherwise indicated. Sample
sizes for each study were esti-
mated on the basis of the
expected differences and pre-
vious experience with the par-
ticular assay. All data are
shown as the mean ± SEM.
Kaplan-Meier survival was sta-
tistically analyzed by the log-
rank test. The percentage of
cytoplasmic or nuclear β-cate-
nin positivity was compared by
Fisher’s exact test. Other sta-
tistical comparisons were eval-
uated with Student’s t test or
one-way ANOVA. Significance
levels were set at *P < 0.05;
**P < 0.01; ***P < 0.001;
n.s., not significant. N indica-
Figure 2. PMN-MDSCs infiltration in p63+ LAC. (A) In both patients and tes biological replicates. Data
mouse models, CD15+ PMN-MDSCs accumulated and expanded in p63+ analyses were carried out
rather than p63- LAC. The quantification further confirmed an increased using Prism 8 (GraphPad).
number of PMN-MDSCs. (B) Representative FACS analysis of mouse mod-
els indicated the expansion of Hdc+ PMN-MDSCs in p63+ tumors. Original Results
magnification × 400 (A). **P < 0.01 (A).
P63 promoted LAC progres-
Scanner 300-7G scanner with GCOS software. sion
A significance cut-off of a Benjamini-Hochberg
false discovery rate ≤ 0.05 was applied. Of the 256 patients with local LAC, 46 patients
(18.0%) exhibited nuclear immunoreactivity for
Quantitative RT-PCR p63 (Figure 1A). The retrospective analysis
suggested that p63+ LAC patients had a worse
Total mRNA of sorted cells was isolated using prognosis than p63 - patients (P < 0.05) (Figure
a RNeasy Micro Kit (Qiagen) and underwent 1B).
5826 Int J Clin Exp Med 2020;13(8):5823-5830MDSCs and stem cell-like acquisition of lung adenocarcinoma
Hdc+ PMN-MDSCs in p63+ LAC
tumors (69.23 ± 7.27%) com-
pared to that of p63- tumors
(3.33 ± 0.28%) (P < 0.001)
(Figure 2B).
Hdc+ myeloid cell-derived Wnts
promoted the expression of
p63
The majority of p63+ LAC tu-
mors (91.2%) showed the tr-
anslocation of β-catenin from
the membrane to the cyto-
plasm and nucleus (Figure
3A). Most p63 - cases (94.8%)
displayed membranous posi-
tivity for β-catenin (Figure 3A).
The cellular source of Wnt
ligands varies according to
different stages. RNA-seq
analysis of Hdc+ PMN-MDSCs
obtained from LAC masses
showed upregulated levels of
Wnts, including Wnt1, 2, 3, and
8a, indicating the involvement
of canonical Wnt signaling
(Figure 3B). RT-PCR analysis
supported that Hdc+ PMN-
MDSC-derived Wnts levels in
Figure 3. Aberrant activation of Wnt/β-catenin signaling in p63+ cells. (A) p63+ LAC tumors were signifi-
The majority of p63+ (91.2%), rather than p63- tumor cells (5.5%), showed
the translocation of β-catenin from the membrane to the cytoplasm and cantly higher than those in
nucleus (P < 0.001). (B) Wnt1, Wnt2, Wnt3, and Wnt5a were upregulated in p63 - tissues (Figure 3C).
Hdc+ PMN-MDSCs cells harvested from p63+ LAC tissues confirmed by RNA-
seq. (C) qRT-PCR revealed high levels of Hdc+ PMN-MDSCs-derived Wnt1, Blocking of Hdc+ PMN-MDSC-
2, 3, and 8a. Original magnification × 400 (A). **P < 0.01; ***P < 0.001 derived Wnts inhibited LAC
(A and C).
progression
MDSCs were recruited into LAC tissues Hdc-CreERT2; eGFP; LSL-KrasG12D mice were
crossed with porcupineflox/flox (Porcn) to elimi-
To further explore the identity of PMN-MDSCs,
nate the secretion of Wnts in Hdc-expressing
we established LAC animal models using Hdc-
cells (Figure 4A). FACS results indicated that
CreERT2; eGFP; LSL-KrasG12D mice, which were
the downregulation of porcupine decreased
treated with AdenoCre. In primary LAC tissues
obtained from patients and mouse models, the incidence of LAC (P < 0.001) rather than
PMN-MDSCs, defined by the combination of the percentage of Hdc+ PMN-MDSCs (P > 0.05)
morphology and CD15+, were recruited and sur- (Figure 4B). Hdc-CreERT2; eGFP; LSL-KrasG12D;
rounded cancer cells (Figure 2A). The quantifi- DTR mice were treated with DT to ablate Hdc+
cation results further revealed the propensity cells, resulting in a decreased percentage of
of CD15+ PMN-MDSCs to surround p63+ cancer both LAC incidence and p63 positivity (Figure
tissues (Figure 2A). Consistent with the immu- 4B and 4C). However, there were no significant
nohistochemical staining results, the FACS differences between Porcn, DTR, and Porcn/
results confirmed an increased percentage of DTR groups (P > 0.05) (Figure 4B and 4C).
5827 Int J Clin Exp Med 2020;13(8):5823-5830MDSCs and stem cell-like acquisition of lung adenocarcinoma
Figure 4. Block of Hdc+ PMN-MDSCs-derived Wnts attenuated p63 expression. (A) At 6 weeks, tamoxifen chow
induced the ablation of Hdc+ PMN-MDSCs-derived Wnts in porcupineflox/flox transgenic mice. Hdc+ cells were deleted
by DT in the combination with tamoxifen in Hdc-CreERT2; eGFP; LSL-KrasG12D; porcupineflox/flox; DTR mice. (B) A block
of Hdc+ cells or Hdc+ cells-derived Wnts decreased the number of LAC. Porcupineflox/flox only ruined the secretion of
Hdc+ cells-derived Wnts, the distinction of anti-LAC abilities between three transgenic animal groups were not sig-
nificant, suggesting the central role of Wnts/β-catenin pathway. (C) In line with Wnts levels, p63+ rates decreased in
transgenic animals. ***P < 0.001; n.s., not significant (B and C).
Discussion tionally considered as a tumor suppressor
gene, endowing cells with proapoptosis and
The heterogeneous nature of cancer remains cell cycle arrest-promoting properties [4, 5].
the rate-limiting step in the anti-LAC battle. Lung squamous cell carcinoma (LSCC) patients
Here we have identified a subgroup of p63- expressing high levels of p63 showed a better
expressing LAC cells at early stages that were prognosis than their p63 - counterparts [6].
associated with a poor prognosis. Hdc+ PMN- Another hypothesis argues that p63 can func-
MDSCs were recruited to and expanded in the tion as an oncogene. Both LAC and its precan-
p63+ tissues. They expressed high levels of cerous lesions exhibit strong immunopositivity
Wnts 1, 2, 3, and 8a to promote tumor progres- for p63 [8]. Upregulated expression of p63
sion and p63 acquisition. The close spatial causes hyperproliferation and terminal differ-
relationship indicated the possibility that PMN- entiation defects [16]. Our study provided pre-
MDSCs regulate the biological behaviors of liminary data that nuclear p63-positivity in the
p63+ cancer cells through a paracrine-like pat- minority of LAC is linked to a poor prognosis,
tern. The ablation of Hdc+ PMN-MDSC-derived further supporting the possibility that p63 pro-
Wnts attenuated tumor progression. motes LAC progression.
LAC is a heterogeneous disease composed of Myeloid-derived immune cells, including MD-
cancer cells with different behaviors. CSCs con- SCs, tumor-associated macrophages (TAMs),
tribute to recurrence or metastasis. However, a and tumor-associated neutrophils (TANs), are
lack of consensus regarding a universal marker essential for the tumorigenesis and progres-
of CSC hampers the clinical utility of CSC- sion of many solid tumors [17, 18]. MDSCs,
targeted therapies. P63 plays an important role which infiltrate and expand in NSCLC tissues
in normal lung development. The downregulat- but not in healthy donors, secret high levels of
ed expression of p63 in transgenic mice led to PD-L1 [17]. The levels of circulating S100A9+
dysplasia of the epithelium [15]. P63 is tradi- MDSCs in LAC patients are correlated with a
5828 Int J Clin Exp Med 2020;13(8):5823-5830MDSCs and stem cell-like acquisition of lung adenocarcinoma
poor prognosis and treatment response [19]. References
Our previous studies indicated that Hdc+ PMN-
MDSCs facilitated the tumorigenesis and pro- [1] Chen W, Zheng R, Baade PD, Zhang S, Zeng H,
gression of colon cancer [20, 21]. The tenden- Bray F, Jemal A, Yu XQ and He J. Cancer statis-
cy of Hdc+ PMN-MDSCs towards p63+ LAC tis- tics in China, 2015. CA Cancer J Clin 2016; 66:
sues suggests a close relationship between 115-132.
microenvironment immune cells and cancer [2] Siegel RL, Miller KD and Jemal A. Cancer sta-
tistics, 2020. CA Cancer J Clin 2020; 70: 7-30.
cells. Consistent with previous studies, Hdc+
[3] Kelsey CR, Marks LB, Hollis D, Hubbs JL, Ready
PMN-MDSCs expresses high levels of Wnts,
NE, D’Amico TA and Boyd JA. Local recurrence
including Wnt1, Wnt2, Wnt3, and Wnt8a, to after surgery for early stage lung cancer: an
aberrantly activate Wnt/β-catenin signaling in 11-year experience with 975 patients. Cancer
p63+ LAC cells. Ablation of Hdc+ PMN-MDSCs 2009; 115: 5218-5227.
in DTR transgenic mice significantly decreased [4] Yang A, Kaghad M, Wang Y, Gillett E, Fleming
the incidence of LAC and the percentage of MD, Dotsch V, Andrews NC, Caput D and McKe-
p63+ cases. Similar results were obtained in on F. p63, a p53 homolog at 3q27-29, encodes
Hdc-CreERT2; porcupineflox/flox mice, in which multiple products with transactivating, death-
Hdc+ myeloid cells were unable to synthesize inducing, and dominant-negative activities.
Wnts. Mol Cell 1998; 2: 305-316.
[5] Gressner O, Schilling T, Lorenz K, Schulze
In conclusion, we have identified a subpopu- Schleithoff E, Koch A, Schulze-Bergkamen H,
lation of LAC with a poor prognosis character- Lena AM, Candi E, Terrinoni A, Catani MV, Oren
ized by positive nuclear expression of p63. We M, Melino G, Krammer PH, Stremmel W and
demonstrated that Hdc+ PMN-MDSCs are re- Muller M. TAp63alpha induces apoptosis by
cruited to and expand in tumor tissues with a activating signaling via death receptors and
significant tendency towards p63+ cells. Sever- mitochondria. EMBO J 2005; 24: 2458-2471.
al canonical Wnts ligands, including Wnt1, [6] Massion PP, Taflan PM, Jamshedur Rahman
Wnt2, Wnt3, and Wnt8a, are upregulated in SM, Yildiz P, Shyr Y, Edgerton ME, Westfall MD,
Roberts JR, Pietenpol JA, Carbone DP and Gon-
Hdc+ PMN-MDSCs. The close spatial relation-
zalez AL. Significance of p63 amplification and
ship between Hdc+ PMN-MDSCs and p63+ LAC
overexpression in lung cancer development
cells endows the immune cells with the ability and prognosis. Cancer Res 2003; 63: 7113-
to regulate the behaviors of malignant cells in 7121.
a paracrine-like pattern. Abolishing of Hdc+ [7] Koster MI, Lu SL, White LD, Wang XJ and Roop
PMN-MDSCs or blocking of Hdc+ PMN-MDSC- DR. Reactivation of developmentally expressed
derived Wnts in transgenic mice attenuated the p63 isoforms predisposes to tumor develop-
tumor burden and p63+ rate in LAC models, ment and progression. Cancer Res 2006; 66:
suggesting a promising prospect for anti-LAC 3981-3986.
clinical treatment on the basis of targeting [8] Sheikh HA, Fuhrer K, Cieply K and Yousem S.
Hdc+ PMN-MDSCs. p63 expression in assessment of bronchioloal-
veolar proliferations of the lung. Mod Pathol
Acknowledgements 2004; 17: 1134-1140.
[9] Zhang Y, Goss AM, Cohen ED, Kadzik R, Lepore
This work was supported by the National Sci- JJ, Muthukumaraswamy K, Yang J, DeMayo FJ,
ence Foundation of China (81770624, 8186- Whitsett JA, Parmacek MS and Morrisey EE. A
0490), and the Science Foundation of Jiangxi Gata6-Wnt pathway required for epithelial
Province for Distinguished Young Scholars, stem cell development and airway regenera-
China (20192BCB23025). tion. Nat Genet 2008; 40: 862-870.
[10] Nekulova M, Holcakova J, Coates P and Vojte-
Disclosure of conflict of interest sek B. The role of p63 in cancer, stem cells and
cancer stem cells. Cell Mol Biol Lett 2011; 16:
None. 296-327.
[11] Heuvers ME, Muskens F, Bezemer K, Lambers
Address correspondence to: Huan Deng, Depart-
M, Dingemans AC, Groen HJM, Smit EF, Hoog-
ment of Pathology, The Fourth Affiliated Hospital steden HC, Hegmans JPJJ and Aerts JGJV. Argi-
of Nanchang University, 133 South Guangchang nase-1 mRNA expression correlates with my-
Road, Nanchang, Jiangxi, China. Tel: +86-791- eloid-derived suppressor cell levels in per-
87022537; Fax: +86-791-87022537; E-mail: bean- ipheral blood of NSCLC patients. Lung Cancer
deng@ncu.edu.cn 2013; 81: 468-474.
5829 Int J Clin Exp Med 2020;13(8):5823-5830MDSCs and stem cell-like acquisition of lung adenocarcinoma
[12] de Goeje PL, Bezemer K, Heuvers ME, Dinge- [18] Quail DF and Joyce JA. Microenvironmental
mans AC, Groen HJ, Smit EF, Hoogsteden HC, regulation of tumor progression and metasta-
Hendriks RW, Aerts JG and Hegmans JP. Immu- sis. Nat Med 2013; 19: 1423-1437.
noglobulin-like transcript 3 is expressed by [19] Feng PH, Yu CT, Chen KY, Luo CS, Wu SM, Liu
myeloid-derived suppressor cells and corre- CY, Kuo LW, Chan YF, Chen TT, Chang CC, Lee
lates with survival in patients with non-small CN, Chuang HC, Lin CF, Han CL, Lee WH and
cell lung cancer. Oncoimmunology 2015; 4: Lee KY. S100A9(+) MDSC and TAM-mediated
e1014242. EGFR-TKI resistance in lung adenocarcinoma:
[13] Saha S, Aranda E, Hayakawa Y, Bhanja P, Atay the role of RELB. Oncotarget 2018; 9: 7631-
S, Brodin NP, Li J, Asfaha S, Liu L, Tailor Y, 7643.
Zhang J, Godwin AK, Tome WA, Wang TC, Guha [20] Yang XD, Ai W, Asfaha S, Bhagat G, Friedman
C and Pollard JW. Macrophage-derived extra- RA, Jin G, Park H, Shykind B, Diacovo TG, Falus
cellular vesicle-packaged WNTs rescue intesti- A and Wang TC. Histamine deficiency promotes
nal stem cells and enhance survival after radi- inflammation-associated carcinogenesis thro-
ation injury. Nat Commun 2016; 7: 13096. ugh reduced myeloid maturation and accumu-
[14] Jackson EL, Willis N, Mercer K, Bronson RT, lation of CD11b+Ly6G+ immature myeloid
Crowley D, Montoya R, Jacks T and Tuveson cells. Nat Med 2011; 17: 87-95.
DA. Analysis of lung tumor initiation and pro- [21] Chen X, Takemoto Y, Deng H, Middelhoff M,
gression using conditional expression of onco- Friedman RA, Chu TH, Churchill MJ, Ma Y,
genic K-ras. Genes Dev 2001; 15: 3243-3248. Nagar KK, Tailor YH, Mukherjee S and Wang
[15] Yang A, Schweitzer R, Sun D, Kaghad M, Walk- TC. Histidine decarboxylase (HDC)-expressing
er N, Bronson RT, Tabin C, Sharpe A, Caput D, granulocytic myeloid cells induce and recruit
Crum C and McKeon F. p63 is essential for re- Foxp3(+) regulatory T cells in murine colon
generative proliferation in limb, craniofacial cancer. Oncoimmunology 2017; 6: e1290034.
and epithelial development. Nature 1999;
398: 714-718.
[16] Koster MI, Kim S, Mills AA, DeMayo FJ and
Roop DR. p63 is the molecular switch for initia-
tion of an epithelial stratification program.
Genes Dev 2004; 18: 126-131.
[17] Yamauchi Y, Safi S, Blattner C, Rathinasamy A,
Umansky L, Juenger S, Warth A, Eichhorn M,
Muley T, Herth FJF, Dienemann H, Platten M,
Beckhove P, Utikal J, Hoffmann H and Uman-
sky V. Circulating and tumor myeloid-derived
suppressor cells in resectable non-small cell
lung cancer. Am J Respir Crit Care Med 2018;
198: 777-787.
5830 Int J Clin Exp Med 2020;13(8):5823-5830You can also read

















































