HD-13 Induces Swine Pneumonia Progression via Activation of TLR9
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Hindawi
Computational and Mathematical Methods in Medicine
Volume 2022, Article ID 8660752, 11 pages
https://doi.org/10.1155/2022/8660752
Research Article
HD-13 Induces Swine Pneumonia Progression via
Activation of TLR9
Jing Liu ,1 Jiayi Xu ,2 Honglei Zhou ,1 and Lihua Wang 1
1
Jiangsu Agri-Animal Husbandry Vocational College, Taizhou, 225300 Jiangsu, China
2
Nanjing Agricultural University, Nanjing, 210095 Jiangsu, China
Correspondence should be addressed to Lihua Wang; ydliujing@sina.com
Received 14 November 2021; Revised 12 December 2021; Accepted 16 December 2021; Published 29 January 2022
Academic Editor: Jun Yang
Copyright © 2022 Jing Liu et al. This is an open access article distributed under the Creative Commons Attribution License, which
permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Swine pneumonia commonly known as swine pasteurellosis is an infectious disease of swine caused by Pasteurella multocida
infection. It has been reported that Toll-like receptors (TLRs) play a vital role in swine pneumonia progression. However, the
underlying mechanism has not been elucidated. This research was aimed at investigating the molecular mechanism by which TLR9
regulates swine pneumonia progression. Our findings illustrated that the HD-13 strain of Pasteurella multocida D (HD-13)
accelerated TLR9 expression in porcine alveolar macrophage 3D4/21 cells; HD-13 activated the inflammatory response via
accelerating TLR9 expression. Mechanistically, HD-13 activated mitogen-activated protein kinase (MAPK) and nuclear factor kB
(NF-κB) signals. In conclusion, HD-13 may activate MAPK and NF-κB pathways via accelerating TLR9 expression, thereby
accelerating the inflammatory response in the progression of swine pneumonia. TLR9 may serve as a novel therapeutic target for
swine pneumonia. Our research may provide a theoretical basis for the prevention and treatment of swine pneumonia.
1. Introduction ella multocida is divided into five types, A, B, D, E, and F,
depending on the serotype of the capsular antigen [7]. It
Swine pneumonia, commonly known as swine pasteurello- has been reported that the HD-13 strain of Pasteurella mul-
sis, is a contagious disease induced by Pasteurella multocida tocida D (HD-13) has strong infectivity [8]. In this paper, we
infection which is divided into chronic type, acute type, and aimed to study the specific mechanism of swine pneumonia
most acute type according to clinical manifestations and the induced by HD-13.
course of the disease [1]. What is more, swine pneumonia A Toll-like receptor (TLR) is a transmembrane protein
has a strong infectivity, and it also causes other infectious encoded by the Toll gene, which is one of cell transmem-
diseases, which seriously affects economic benefits [2]. At brane receptors and pathogen pattern recognition receptors
present, the prevention of swine pneumonia mainly depends (PRRs) in the innate immune system [9]. Prompt com-
on antibiotics, inactivated vaccines, attenuated vaccines, mencement of an inflammatory response is accomplished
subunit vaccines, etc. [3]. However, no obvious therapeutic with the help of unique and conserved PRRs present on
effect has been achieved, due to the emergence of drug resis- the surface of myeloid cells, lung epithelial cells, and lym-
tance and side effects [4]. Therefore, it is urgent to elucidate phoid tissue. PRRs comprise TLRs, RIG-I-like receptors
the specific pathogenesis of swine pneumonitis for seeking (RLRs), and NOD-like receptors (NLRs). TLRs activate viral
effective therapeutic drugs. responses by recognizing pathogenic microorganisms such
Pasteurella multocida, which is an important zoonotic as viruses, bacteria, and mycoplasmas to release inflamma-
pathogenic bacterium to infect a variety of livestock, such tory cytokines [10]. TLRs are important constituents of the
as poultry and wild animals, results in causing pasteurellosis innate immune system, which were applied to recognize
[5]. Pig is one of the susceptible animals of Pasteurella mul- and defend against foreign pathogens. Toll is a gene in fruit
tocida, which mainly causes swine lung disease [6]. Pasteur- fly and toll-like proteins which was discovered in animals
2 Computational and Mathematical Methods in Medicine
and also was named as TLRs. TLRs are conserved and ding should be changed once a day. Drinking water was pro-
approximately comprised 10 functional TLRs (TLR1 to vided continuously, and the pigs were provided fodder
TLR10) in humans and 12 (TLR1 to TLR9, TLR11 to (Zhong liang, COFCO, Beijing, China) every day. At the end
TLR13) in mice. TLR1, TLR2, TLR4, TLR5, TLR6, and of the experiment, the pigs were killed by neck detachment.
TLR10 are expressed on the human cell surface, while Spleen and lung tissues were cut and frozen at 80°C for func-
TLR3, TLR7, TLR8, and TLR9 are expressed on the mem- tional experiment. The animal experiments involved in this
brane of intracellular vesicles [9, 10]. TLR9 is a member of paper were carried out in accordance with the Guidelines for
the TLR family whose function is triggered by the unmethy- the Care and Use of Laboratory Animals issued by the
lated CpG motif of microbial DNA, which initiates the National Institutes of Health (NIH).
immune response by inducing cytokines and many mole-
cules [11]. It has been reported that various hosts recognize 2.2. Pasteurella multocida, Antibodies, and Reagents. HD-13
lipopolysaccharides in the cell membrane structure through was provided by China Agricultural University (Beijing,
TLR9 and activate downstream mitogen-activated protein China), which was provided ready to use. The HD-13
kinase (MAPK) and nuclear transcription factor-κB (NF-κB) (104 CFU/mL) strain was infected by nasal drip and respira-
pathways mediated by MyD88 to promote the MAPK protein tory tract infection, and the bacterial solution was dropped
phosphorylation, and NF-κB enters the nucleus, further into the nasal cavity of pigs. Reagents, including PD98059,
releasing cellular inflammatory factors [12]. Namely, TLR9 is which served as the p-ERK/ERK signaling pathway inhibi-
triggered by lipopolysaccharide in the same way that it is by tor, SB203580 which served as the p-p38/p38 signaling path-
CpG motifs, resulting in cellular inflammatory factor release. way inhibitor, SP600125 which served as the p-JNK/JNK
Therefore, it is speculated that TLR9, MAPK, and NF-κB signaling pathway inhibitor, and BAY11-7082 which served
pathways may play a vital role in the inflammatory response as the NF-κB signaling pathway inhibitor, were purchased from
of swine pneumonia. However, the underlying mechanism Solarbio (Beijing, China). Antibodies, including TLR9, MyD88,
remains unclear. TRIF, TRAF6, p-ERK, ERK, p-p38, p38, p-JNK, JNK, p-p65,
In this research, we aimed to explore TLR9 roles in swine p65, p-IKKα, IKKα, and β-actin, were provided by Abcam
pneumonia progression, and the underlying mechanism was (Cambridge, UK, 1 : 1000, monoclonal, species of mice).
illuminated by evaluating the levels of TLR9 downstream
effectors, including myeloid differentiation factor 88 2.3. Cell Culture. Primary 3D4/21 cells obtained from Kun-
(MyD88), Toll-like receptor-associated activator of interferon ming Medical University (Kunming, Yunnan) were cultured
(TRIF), and tumor necrosis factor receptor-associated factor 6 in Dulbecco’s modified Eagle’s medium (DMEM, Roche,
(TRAF6), and the inflammatory cytokines, including Basel, Switzerland) supplemented with 10% fetal bovine
interleukin- (IL-) 1β, IL-6, IL-12, and tumor necrosis factor serum (FBS, Roche, Basel, Switzerland) and 1% penicillin-
α (TNF-α). Our findings may provide a theoretical basis for streptomycin solution (Solarbio, Beijing, China) in a humid
the prevention and treatment of swine pneumonia. incubator containing 5% CO2 at 37°C.
2. Materials and Methods 2.4. Cell Transfection and Infection. Sangon Biotech
(Shanghai, China) synthesizes the random sequences of
2.1. Piglet and Tissue Specimen. 4-week-old Southern Yunnan TLR9 siRNA and the negative controls (Scra siRNA). When
small ear pigs (n = 60, 12 per group) used in this study were the cell confluency reached about 80%, TLR9 siRNA or Scra
given by China Agricultural University (Beijing, China). Pig- siRNA was transfected into 3D4/21 cells by using the Lipo-
lets were raised in houses with metal fences and concrete fectamine™ 3000 Transfection Reagent (Takara, Liaoning,
floors, with an area of 1.5 to 3.0 m2 per strip, and the fence China). Taken briefly, 3D4/21 cells were randomly divided
should be at least 0.6 m high, providing dry ground for each into 4 groups: Scra group—cells were transfected with Scra
animal to lie down and fully extend its body. The ground siRNA (2 μg) for 48 h, which serves as a negative control;
and playground are cement floors with obvious slopes to pre- TLR9 siRNA group—cells were transfected with TLR9
vent sewage from being trapped or in the pouring circle. The siRNA (2 μg) for TLR9 silencing for 48 h; HD-13+Scra
passageway and the ground have an effective drainage system group—cells were infected with HD-13 and transfected with
and are equipped with leaking fecal and urine floors and auto- Scra siRNA (2 μg) for 48 h; and HD-13+TLR9 siRNA
matic drinking fountains. Pig manure was treated with “dry group—cells were infected with HD-13 and transfected with
treatment”; that is, the feces were separated from the sewage, TLR9 siRNA for TLR9 silencing for 48 h. At 48 h posttreat-
the pig manure was accumulated and fermented, and the sew- ment, 1 × 106 3D4/21 cells were applied to subsequent
age was discharged to the treatment area far away from the pig experiments in each group. Furthermore, in vivo, plasmids
house through closure and sewage ditches. The suitable tem- (1 × 1012 PFU/mL) were injected into piglets via tail vein
perature for raising pigs is 18-25°C, and the relative tempera- injection.
ture is 40%-60%. The pigsty requires warm winter, no wind,
cool and ventilated summer, and shade. It is required to be 2.5. RT-qPCR. The TRIzol reagent (Takara, Liaoning, China)
ventilated at least 20 times per hour in order to effectively was used to extract the total RNA from 3D4/21 cells with
remove moisture and heat from the house and give it no less transfection of corresponding plasmids or piglet tissues.
than 10 h of light every day. Hay, sawn wood, chuanhua, or The M-MLV Reverse Transcriptase (RNase H) kit (Takara,
sawdust was selected as bedding materials for pigsty. The bed- Liaoning, China) was employed to synthesize cDNA. RT-
Computational and Mathematical Methods in Medicine 3
qPCR was performed as previously described [13]. Primers Table 1: Primer sequences.
used in RT-qPCR are shown in Table 1.
Primer name (5 ′ -3 ′ ) primer sequences
2.6. Western Blot. Total proteins were harvested from the F-TLR1 GCCTGGATGTGGTTCCTTTA
transfected 3D4/21 cells with corresponding plasmids or R-TLR1 ATGGATTGTTCCCTGCTTTG
infected with HD-13 by using cell lysis buffer (Beyotime,
F-TLR2 ACACCGCCATCCTCATTCTG
Nanjing, China). Western blots were conducted as previously
described [14]. In brief, proteins were resolved by sodium R-TLR2 CTTCCCGCTGCGTCTCATCC
dodecyl sulfate-polyacrylamide gel electrophoresis, transferred F-TLR3 ACTGATGCTCCGAAGGGTGG
onto polyvinylidene fluoride membranes (Millipore, Bedford, R-TLR3 CGTGTTTCCAGAGCCGTGCT
MA, USA), and incubated with primary antibodies overnight F-TLR4 CCGTCATTAGTGCGTCAGTT
at 4°C. A horseradish peroxidase-conjugated secondary anti- R-TLR4 TTCCTCACCCAGTCTTCGTC
body was incubated with the membrane at the second day. F-TLR5 GGTTCTCGCCCACCACATTA
The blots were visualized with the ECL reagent (Millipore).
R-TLR5 GGGTCCCAAAGAGTCGGAAG
All antibodies were obtained from Abcam (Cambridge, UK,
1 : 1000). The optical density of protein bands was quantified F-TLR6 TCTGCTCAAGGACTTCCGTGTA
by using ImageJ software (ImageJ Software Inc., USA). R-TLR6 CGGTCATGGTAAGAGCCAAT
F-TLR7 GTATGGGCAGACCTTGGACC
2.7. Immunofluorescence. Immunofluorescence was per- R-TLR7 TTGGCTGATGCTATTTCCTGAT
formed as described with a minor modification. Briefly, 1 F-TLR8 GTGGAAACCGCCTGGACCTT
× 105 3D4/21 cells were seeded in 12-well plates for trans-
R-TLR8 GGCTTCACTCGGGATGTGCT
fection or infection. Cells were washed with 1× PBS and
fixed for 15 min in 4% formaldehyde. Then, Triton-XTM F-TLR9 AGACTGGTTACCTGGCAAGACG
100 was added to increase membrane permeability. After R-TLR9 AGGAAACTGGCACGCAAGAG
washing and blocking, 3D4/21 cells were incubated with F-TLR10 TGGAAATCCTGGGTTTGAGT
the TLR9 or p-p65 antibody (Abcam, Cambridge, UK, R-TLR10 ATGGGCAGGCTACCTTCTTC
1 : 1000) overnight at 4°C. At the following day, cells were F-TLR9 5 ′ -ACAACAACATCCACAGCCAAGTGTC-3 ′
incubated with a secondary antibody (mouse anti-rabbit
IgG-HRP, Abcam, Cambridge, UK, 1 : 1000) for 1 h. Then, R-TLR9 5 ′ -AAGGCCAGGTAATTGTCACGGAG-3 ′
cells were counterstained with DAPI and sealed with a cover F-MyD88 5 ′ -CGACGCCTTCATCTGCTACTGC-3 ′
glass and finally observed with a confocal fluorescence
R-MyD88 5 ′ -CCACCACCATGCGACGACAC-3 ′
microscope (LSM880, Zeiss, Germany).
F-TRIF 5 ′ -TGAAGGAGGAAGCTGTAGCA-3 ′
2.8. ELISA. 1 mL of protein extraction reagent (Beyotime, 5 ′ -TGAAGGAGAAATGGCTTTCC-3 ′
R-TRIF
Nanjing, China) was applied to lyse lung tissues which were
separated from the piglet or HD-13-infected or plasmid- F-TRAF6 5 ′ -GTTGCTGAAATCGAAGCACA-3 ′
transfected 3D4/21 cells. Subsequently, the concentration R-TRAF-6 5 ′ -CGGGTTTGCCAGTGTAGAAT-3 ′
of inflammatory cytokines, including IL-6, IL-12, TNF-α,
F-IL-1β 5 ′ -CTCGTGCTGTCGGACCCAT-3 ′
and IL-1β, was determined by using an ELISA kit (Roche,
Basel, Switzerland) as depicted in the manual. R-IL-1β 5 ′ -CAGGCTTGTGCTCTGCTGTG-3 ′
F-IL-6 5 ′ -GGTACATCCTCGACGGCATCT-3 ′
2.9. Statistical Analysis. The representative data obtained
from this study was shown as the mean ± SD from three R-IL-6 5 ′ -GTGCCTCTTGCTGCTTTCAC-3 ′
independent experiments. GraphPad Prism version 5.0 soft- F-IL-12 5 ′ -GACATTCTGCGTTCAGGTCCAG-3 ′
ware (GraphPad Software, Inc.) is used for statistical analysis
of all data. A t-test was used for comparison between two R-IL-12 5 ′ -CATTTTTGCGGCAGATGACCGTG-3 ′
groups, and ANOVA followed by the Tukey post hoc F-TNF-α 5 ′ -ATCCGCGACGTGGAACTGG-3 ′
analysis was used for comparison between multiple groups.
R-TNF-α 5 ′ -CCATGCCGTTGGCCAGGAGG-3 ′
P value < 0.05 indicates that the difference is statistically
significant. F-GAPDH 5 ′ -GAGTCAACGGATTTGGTCGT-3 ′
R-GAPDH 5 ′ -TTGATTTTGGAGGGATCTCG-3 ′
3. Results TLR9 siRNA GCCCCACTTCTCGGCGGCAGC
3.1. HD-13 Upregulates TLR9 Level in 3D4/21 Cells. To Scra CCCAUUCGGGGGGGCAAACUCTT
explore the role of TLR receptor in swine pneumonia pro-
gression, RT-qPCR was conducted to assess the levels of
TLRs in 3D4/21 cells, following HD-13 infection at 0 h, 1 h, trol. For the TLR9 gene, HD-13 upregulated TLR9 level in
4 h, 8 h, 12 h, 24 h, and 48 h, respectively. Results indicated a time-dependent manner (Figure 1(a)). Following HD-13
that except for the TLR9 gene, there are no changes of infection of the piglet at 0 h, 12 h, 24 h, 36 h, and 48 h,
mRNA levels in other TLR genes, compared with the con- respectively, RT-qPCR was carried out to detect TLR level
4 Computational and Mathematical Methods in Medicine
8
8
Relative expression of mRNA
Relative expression of mRNA
6 6
4 4
2 2
0 0
0h 1h 4h 8h 12 h 24 h 48 h 0h 12 h 24 h 36 h 48 h
TLR1 TLR6 TLR1 TLR6
TLR2 TLR7 TLR2 TLR7
TLR3 TLR8 TLR3 TLR8
TLR4 TLR9 TLR4 TLR9
TLR5 TLR10 TLR5 TLR10
(a) (b)
15
Relative expression of TLR9
10 **
0h 1h 4h 12h 24h 48h **
TLR9 5 *
*
*
β-actin
0
0h 1h 4h 12h 24h 48h
(c)
0h 1h 4h 12h 24h 48h
DAPI
TLR9
Merge
(d)
Figure 1: HD-13 activates TLR9 expression. (a) RT-qPCR was performed to assess TLR expression in 3D4/21 cells, following HD-13
infection at 0 h, 1 h, 4 h, 8 h, 12 h, 24 h, and 48 h, respectively. (b) RT-qPCR was conducted to assess TLR expression in the spleen of pig,
following HD-13 infection at 0 h, 12 h, 24 h, 36 h, and 48 h, respectively. (c) Western blot was carried out to detect TLR9 expression in
3D4/21 cells, following HD-13 infection at 0 h, 1 h, 4 h, 12 h, 24 h, and 48 h, respectively. (d) Immunofluorescence was performed to
assess TLR9 expression in 3D4/21 cells. ∗ P < 0:05, ∗∗ P < 0:01 vs. control group.
in spleen tissues; the results were consistent with the above orescence (Figure 1(d)). Collectively, HD-13 may upregulate
(Figure 1(b)). To further verify the effect of HD-13 on TLR9 level in 3D4/21 cells.
TLR9, Western blot was performed to evaluate the level of
TLR9 in 3D4/21 cells, following HD-13 infection at 0 h, 3.2. HD-13 Activates the Inflammatory Response via
1 h, 4 h, 8 h, 12 h, 24 h, and 48 h, respectively. Findings Upregulating TLR9 Level. To investigate whether HD-13
showed that HD-13 promoted the level of TLR9 protein in mediates the inflammatory response in 3D4/21 cells by
a time-dependent manner (P < 0:05; Figure 1(c)). What is accelerating TLR9, following infection with HD-13 or
more, the same phenomenon was observed via immunoflu- TLR9 silencing in 3D4/21 cells, RT-qPCR was performed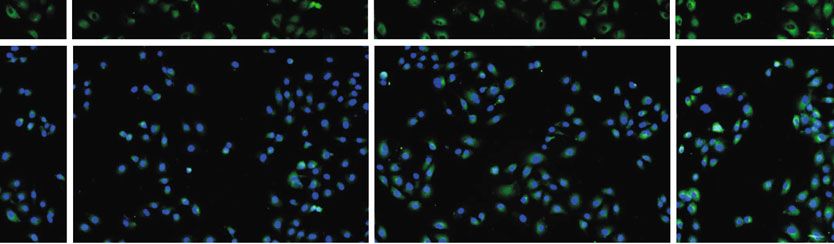
Computational and Mathematical Methods in Medicine 5
5
Relative expression of TLR9
6 ⁎⁎⁎ ⁎⁎⁎ ⁎⁎⁎
⁎⁎⁎
Relative expression
4
4
of mRNA
3
2
2 ⁎
1
⁎⁎ ⁎⁎ ⁎⁎ ⁎⁎
0 0
MyD88 TRIF TRAF6
Scra
TLR9 siRNA Scra
HD-13+Scra TLR9 siRNA
HD-13+TLR9 siRNA HD-13+Scra
HD-13+TLR9 siRNA
(a) (b)
HD-13+TLR9
TLR9 siRNA
HD-13+Scra
siRNA
2.0 6
Scra
⁎⁎⁎
Relative protein expression
⁎⁎⁎
⁎⁎ ⁎⁎
Relative expression
⁎⁎⁎
MyD88 1.5 ⁎⁎
4 ⁎⁎⁎
of mRNA
TRIF 1.0
⁎⁎ ⁎⁎ ⁎⁎
⁎⁎ 2 ⁎ ⁎ ⁎
TRAF6 0.5 ⁎⁎ ⁎⁎ ⁎⁎
⁎⁎ ⁎⁎ ⁎⁎ ⁎⁎
β-actin 0.0 0
MyD88 TRIF TRAF6 IL-1β IL-6 IL-12 TNF-α
Scra
Scra
TLR9 siRNA
TLR9 siRNA
HD-13+Scra
HD-13+Scra
HD-13+TLR9 siRNA
HD-13+TLR9 siRNA
(c) (d)
800 800 500 500
⁎⁎⁎ ⁎⁎⁎ ⁎⁎⁎ ⁎⁎⁎
400 400
TNF-α(pg/mL)
600 600
IL-6 (pg/mL)
IL-12 (pg/mL)
IL-1β(pg/mL)
300 300
400 400
⁎ 200 ⁎ 200 ⁎
⁎
200 200
100 100
⁎⁎ ⁎⁎ ⁎⁎ ⁎⁎
0 0 0 0
A
-1 A
ra
ra
A
LR cra
A
ra
A
R9 cra
a
A
A
ra
-1 A
ra
N
r
N
N
N
N
Sc
Sc
N
Sc
N
Sc
H siRN
Sc
Sc
S
iR
S
siR
siR
siR
siR
siR
siR
3+
3+
3+
3+
9s
-1
-1
R9
R9
R9
R9
R9
R9
D
D
D
D
TL
TL
TL
TL
TL
TL
H
TL
H
T
H
3+
3+
3+
3+
-1
-1
-1
-1
D
D
D
D
H
H
H
H
(e)
Figure 2: HD-13 activates the inflammatory response via accelerating TLR9 expression. (a, b) RT-qPCR was performed to assess the mRNA
levels of TLR9, MyD88, TRIF, and TRAF6. (c) Western blot was conducted to evaluate the levels of MyD88, TRIF, and TRAF6 proteins. (d)
RT-qPCR was carried out to detect the mRNA levels of IL-1β, IL-6, IL-12, and TNF-α. (e) ELISA was performed to detect the expression of
IL-1β, IL-6, IL-12, and TNF-α in the supernatant of 3D4/21 cells. ∗∗ P < 0:01 vs. control group, ###P < 0:001 vs. control group, and $$P < 0:01
vs. HD-13 group.
to assess TLR9 level. Results showed that HD-13 upregulated Figure 2(c)). IL-1β, IL-6, IL-12, and TNF-α serve as the vital
the level of TLR9 and rescued the downregulation of TLR9 inflammatory cytokines [15]; following infection with HD-
level caused by transfection of TLR9 siRNA (P < 0:05; 13 or TLR9 silencing in 3D4/21 cells, RT-qPCR and ELISA
Figure 2(a)). Further findings showed that TLR9 silencing were conducted to detect their levels. Results indicated that
inhibited the mRNA levels of the TLR9 downstream effector, TLR9 silencing inhibited the levels of IL-1β, IL-6, IL-12,
including MyD88, TRIF, and TRAF6, but HD-13 upregu- and TNF-α, whereas HD-13 upregulated their levels. Fur-
lated their levels. It is worth noting that TLR9 silencing thermore, TLR9 silencing suppresses the levels of IL-1β,
inhibited the mRNA levels of MyD88, TRIF, and TRAF6 IL-6, IL-12, and TNF-α promoted by HD-13 (P < 0:05;
accelerated by HD-13 (P < 0:05; Figure 2(b)). The above Figures 2(d) and 2(e)). Taken together, HD-13 may activate
results were further verified by Western blot (P < 0:05; the inflammatory response via accelerating TLR9 level.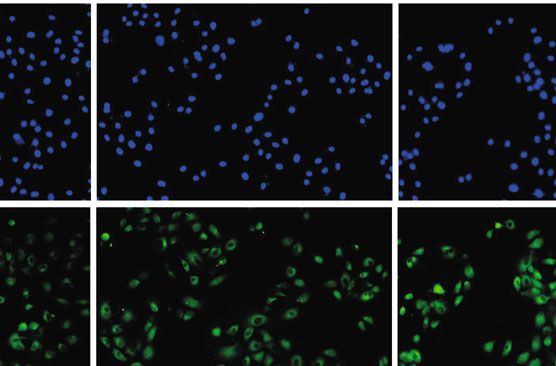
6 Computational and Mathematical Methods in Medicine
3 ( n)
)
)
in
in
i
m
m
m
60
90
90
3(
3(
H ol
r
-1
-1
-1
nt
D
D
D
Co
H
H
p-ERK
ERK
p-p38
Relative protein expression
15
p38 ⁎⁎⁎
⁎⁎⁎
10 ⁎⁎
p-JNK ⁎⁎
⁎⁎
JNK ⁎⁎ ⁎⁎ ⁎⁎
5 ⁎⁎
β-actin
0
p-ERK p-p38 p-JNK
Control
HD-13 (60min)
HD-13 (90min)
HD-13 (90min)
(a)
)
)
)
in
in
in
m
m
m
60
90
90
3(
3(
3(
l
ro
-1
-1
-1
nt
D
D
D
Co
H
H
H
p-p65
Relative protein expression
15
p65 ⁎⁎⁎
10
p-IKKα ⁎⁎
⁎⁎ ⁎⁎ ⁎⁎
IKKα 5
⁎
β-actin
0
p-p65 p-IKKα
Control
HD-13 (60min)
HD-13 (90min)
HD-13 (90min)
(b)
Control HD-13 (60min) HD-13 (90min) HD-13 (90min)
(c)
Figure 3: Continued.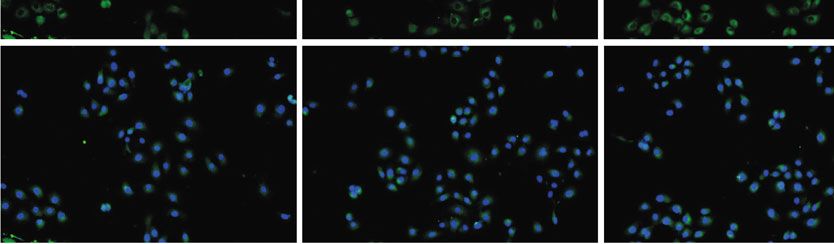
IL-1β (pg/mL) IL-1β (pg/mL) IL-1β (pg/mL) Relative expression of mRNA
0
2
4
6
100
150
200
100
150
200
100
150
200
50
50
50
0
0
0
Co Co
nt Co
ro nt nt
l ro ro
⁎⁎
l l
IL-1β
#
H H
D H
HD-13
D D
Control
-1
3 -1
⁎⁎
-1
⁎⁎
⁎⁎
H 3 H 3
D H D
D
IL-6
-1
⁎⁎⁎
3+ -1 -1
3+ 3+
##
PD PD
98 PD
05 98 98
9 05 05
9
##
##
##
9
HD-13+PD98059
⁎⁎
IL-12
##
IL-6(pg/mL) IL-6(pg/mL) IL-6(pg/mL)
⁎⁎
100
200
300
400
0
100
200
300
400
0
100
200
300
0
TNF-α
##
Co Co Co
nt
ro nt nt
ro
Relative expression Relative expression of mRNA
l ro
l l
of mRNA
0
1
2
3
4
5
H H H
0
2
4
6
D D D
-1 -1 -1
3 3 3
⁎⁎
⁎⁎
⁎⁎
H H
D H D
-1 D -1
3+ -1
Computational and Mathematical Methods in Medicine
P 3+ 3+
⁎⁎⁎
D
⁎⁎
PD
IL-1β
98 PD
IL-1β
98
#
##
05 98
#
#
9 05 05
9
##
9
HD-13
Control
(f)
(e)
(d)
⁎⁎⁎
IL-6
⁎⁎
IL-12(pg/mL) IL-12(pg/mL) IL-12(pg/mL)
IL-6
#
##
100
200
300
0
100
200
300
400
500
0
100
200
300
400
0
⁎⁎
IL-12
⁎⁎⁎
Co
Figure 3: Continued.
Co Co
nt n
##
ro nt
HD-13+BAY11-7082
##
tro ro
l l l
⁎⁎
H H
⁎⁎
D D H
#
D
TNF-α
-1 -1
H 3 -1
IL-12 TNF-α
3
⁎⁎
H H 3
⁎⁎⁎
D
⁎⁎
##
-1 D D
3+ -1 -1
PD 3+ 3+ Relative expression of mRNA
PD PD
98 98 98
05 05 05
9 9
##
9
##
##
0
1
2
3
4
5
TNF-α (pg/mL) TNF-α (pg/mL) TNF-α (pg/mL)
⁎⁎
IL-1β
##
0
200
400
600
800
1000
0
200
400
600
800
0
200
400
600
800
Co Co Co
⁎⁎
nt
IL-6
nt nt ro
ro ro l
l l
##
H H H
D D D
-1
⁎⁎
-1 -1 3
⁎⁎
⁎⁎⁎
3 3 H
⁎⁎
H H
IL-12
D D D
-1
##
-1 -1 3+
3+ 3+
PD PD PD
98 98 98
05
05 05 9
⁎⁎
##
##
##
9 9
TNF-α
##
7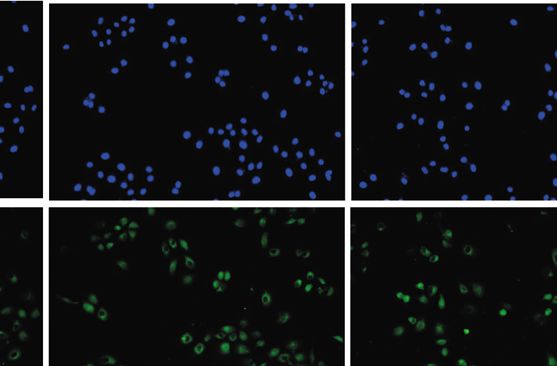
8 Computational and Mathematical Methods in Medicine
300 ⁎⁎ 400 500 ⁎⁎ 800 ⁎⁎
⁎⁎
TNF-α (pg/mL)
1L-12 (pg/mL)
IL-1β(pg/mL)
400
1L-6 (pg/mL)
300 # 600
200 # # ##
300
200 400
100 200
100 100 200
0 0 0 0
82
82
82
82
3
3
3
3
l
l
l
l
ro
ro
ro
ro
-1
-1
-1
-1
0
0
0
70
nt
nt
nt
nt
-7
-7
-7
D
D
D
D
1-
Co
Co
Co
Co
H
H
H
H
11
11
11
Y1
AY
AY
AY
BA
+B
+B
+B
3+
3
3
3
-1
-1
-1
-1
D
D
D
D
H
H
H
H
(g)
Figure 3: HD-13 activates the MAPK and NF-κB pathway. 3D4/21 cells were infected with HD-13 for 0 h, 0.5 h, 1 h, and 1.5 h, respectively.
(a) Western blot was performed to assess the levels of MAPK pathway-related proteins, including ERK, p-ERK, p38, p-p38, JNK, and p-JNK.
(b) The levels of NF-κB pathway-related proteins, including p65, p-p65, IKKα, and p-IKKα, were evaluated by Western blot, following
infection with HD-13 for 0 h, 1 h, 6 h, and 112 h, respectively. (c) Immunofluorescence was conducted to assess p-p65 expression. 3D4/
21 cells were randomly divided into 3 groups: control group—cells were untreated; HD-13 group—cells were infected with HD-13 for
1.5 h; and HD-13+PD98059/HD-13+SB203580/HD-13+SP600125/HD-13+BAY11-7082 group—cells were infected with HD-13 and
10 μM PD98059 or 10 μM SB203580 or 10 μM SP600125 or 10 μM BAY11-7082 for 1.5 h. PD98059 serves as the p-ERK/ERK pathway
inhibitor, SB203580 serves as the p-p38/p38 pathway inhibitor, SP600125 serves as the p-JNK/JNK pathway inhibitor, and BAY11-7082
serves as the NF-κB pathway inhibitor. (d, e) RT-qPCR was performed to assess the mRNA levels of IL-1β, IL-6, IL-12, and TNF-α. (f,
g) ELISA was carried out to detect the expression of IL-1β, IL-6, IL-12, and TNF-α in the supernatant of 3D4/21 cells. ∗ P < 0:05, ∗∗ P <
0:01, and ∗∗∗ P < 0:001 vs. control group; #P < 0:05, ##P < 0:01 vs. HD-13 group.
3.3. HD-13 Activates the MAPK and NF-κB Pathway. To ing TLR9 expression, thereby activating inflammatory
investigate the specific mechanism by which HD-13 acti- response in 3D4/21 cells, following infection with HD-13
vates the inflammatory response, following infection with or TLR9 silencing in 3D4/21 cells, Western blot was per-
HD-13 for 0 h, 0.5 h, 1 h, and 1.5 h, respectively, Western formed to measure the levels of MAPK pathway-related pro-
blot was conducted to assess the levels of MAPK pathway- teins, including ERK, p-ERK, p38, p-p38, JNK, and p-JNK.
related proteins, including ERK, p-ERK, p38, p-p38, JNK, Results showed that TLR9 silencing inhibited the levels of
and p-JNK in 3D4/21 cells. Findings showed that HD-13 p-ERK, p-p38, and p-JNK proteins, but HD-13 upregulated
accelerated the levels of p-ERK, p-p38, and p-JNK proteins, their levels. Attentively, TLR9 silencing inhibited the levels
and HD-13 phosphorylation was most effective at 1 h of of p-ERK, p-p38, and p-JNK proteins induced by HD-13
infection (P < 0:01; Figure 3(a)). Simultaneously, following (P < 0:05; Figure 4(a)), indicating that HD-13 may activate
infection with HD-13 for 0 h, 1 h, 6 h, and 12 h, respectively, MAPK pathways via upregulating TLR9 level. The NF-κB
in 3D4/21, Western blot was carried out to detect the levels pathway was explored by the similar way; results indicated
of NF-κB pathway-related proteins, including p65, p-p65, that TLR9 silencing inhibited the levels of p-p65 and
IKKα, and p-IKKα. Results indicated that the levels of p- p-IKKα, whereas HD-13 upregulates their level. Interestingly,
p65 and p-IKKα were promoted by HD-13, and the most the HD-13 effect was alleviated by TLR9 silencing (P < 0:05;
effective time point was 6 h, which was confirmed by immu- Figures 4(b) and 4(c)). These findings suggested that HD-13
nofluorescence (P < 0:05; Figures 3(b) and 3(c)). To further may activate the NF-κB pathway via upregulating TLR9 level.
verify whether HD-13 activates the inflammatory response Taken together, HD-13 may activate MAPK and NF-κB path-
via the MAPK and NF-κB pathway, while infecting 3D4/21 ways via upregulating TLR9 level.
cells with HD-13 for 1 h, the corresponding MAPK pathway
inhibitors were added, in which PD98059 acts as the inhibi- 4. Discussion
tor of the p-ERK/ERK pathway, SB203580 serves as the
inhibitor of the p-p38/p38 pathway, and SP600125 serves Swine pneumonia is a respiratory infectious disease caused
as the inhibitor of the p-JNK/JNK pathway. Western blot by Pasteurella multocida, which is highly contagious and
and ELISA results indicated that HD-13 promoted the levels often causes other infectious diseases, seriously damaging
of IL-1β, IL-6, IL-12, and TNF-α, which were alleviated by the benefits of pig industry [16]. It is critical to explore the
MAPK pathway inhibitors (P < 0:05; Figures 3(d) and pathogenesis of swine pneumonia and seek effective preven-
3(f)). The NF-κB pathway was investigated in the same man- tion and treatment methods. Our findings demonstrated
ner, and similar results were observed (P < 0:05; Figures 3(e) that HD-13 may activate MAPK and NF-κB pathways via
and 3(g)). Collectively, HD-13 may activate the MAPK and accelerating TLR9 expression, thereby activating the inflam-
NF-κB pathway. matory response in the progression of swine pneumonia.
Our research may provide a theoretical basis for the preven-
3.4. HD-13 Activates MAPK and NF-κB Pathways via tion and treatment of swine pneumonia.
Upregulating TLR9 Level. To further investigate whether Activated TLR initiates a series of events involving a
HD-13 activates MAPK and NF-κB pathways by accelerat- variety of protein kinases, eventually leading to some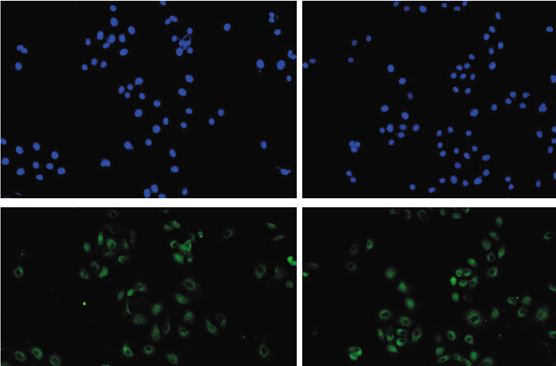
Computational and Mathematical Methods in Medicine 9
A
N
iR
s
R9
A
a
cr
TL
N
+S
iR
3+
s
3
-1
-1
R9
ra
D
D
TL
Sc
H
H
p-ERK
ERK
3 ⁎⁎⁎
p-p38 ⁎⁎⁎
Relative protein expression
p38 ⁎⁎
2 ⁎⁎
⁎⁎
p-JNK
1
JNK ⁎⁎ ⁎ ⁎
β-actin
0
p-ERK p-p38 p-JNK
Scra HD-13+Scra
TLR9 siRNA HD-13+TLR9 siRN
(a)
A
N
siR
R9
A
ra
TL
N
Sc
siR
3+
3+
-1
-1
4
R9
Relative expression of p65/IKKα
ra
D
D
TL
Sc
H
H
⁎⁎⁎
p-p65
3 ⁎⁎
p65
⁎⁎
p-IKKα 2 ⁎
IKKα
1
⁎ ⁎
β-actin
0
p-p65 p-IKKα
Scra HD-13+Scra
TLR9 siRNA HD-13+TLR9 siRNA
(b)
Scra TLR9 siRNA HD-13+Scra HD-13+TLR9 siRNA
(c)
Figure 4: HD-13 activates MAPK and NF-κB pathways via accelerating TLR9 expression. 3D4/21 cells were randomly divided into 4
groups: Scra group—cells were transfected with Scra siRNA for 48 h, which serves as a negative control; TLR9 siRNA group—cells were
transfected with TLR9 siRNA for TLR9 silencing for 48 h; HD-13+Scra group—cells were infected with HD-13 for 48 h; and
HD-13+TLR9 siRNA group—cells were infected with HD-13 and transfected with TLR9 siRNA for TLR9 silencing for 48 h. (a) Western
blot was conducted to evaluate the levels of MAPK pathway-related proteins, including ERK, p-ERK, p38, p-p38, JNK, and p-JNK. (b)
The levels of NF-κB pathway-related proteins, including p65, p-p65, IKKα, and p-IKKα, were measured by Western blot. (c)
Immunofluorescence was performed to assess p-p65 expression. ∗ P < 0:05, ∗∗ P < 0:01 vs. control group; ##P < 0:05, ###P < 0:001 vs. control
group; $P < 0:05, $$P < 0:01 vs. HD-13 group.10 Computational and Mathematical Methods in Medicine
transcription factors such as NF-κβ, activation of MAPK, was alleviated by HD-13. The above findings indicate that
IFN regulatory factor (IRF), and activator protein 1 (AP1). HD-13 may activate NF-κB pathways through accelerating
It is known that it plays an important role in the induction TLR9 expression, resulting in aggravating the development
of inflammatory mediators, so it induces antiviral innate of swine pneumonia. Besides, studies showed that human
immune response through TLR3- and TLR4-mediated signal melanocytes express functional TLR9, and the TLR9 ligand
transduction. This was confirmed by the deficiency of IFN- activates NF-κB signals and induces the expression of cyto-
related gene expression mediated by TLR3 and TLR4 in kines and chemokines in melanocytes. Cytosine guanine
TRIF-knockout mice [9, 11, 15, 16]. Many genetic associa- ODN2006 is an agonist of TLR9, which can be activated
tion studies have shown that TLR is also associated with by NF-κB activation increasing melanin synthesis. The com-
the exacerbation of allergic asthma. For example, TLR7 bination of odn2006 and UVB irradiation significantly
and TLR 8 are associated with the progression of asthma, increased the expression of tyrosinase and PMEL in melano-
and their ligands can prevent airway remodeling in experi- cytes. Therefore, TLR9 stimulation tends to enhance mela-
mentally induced asthma [12–16]. The ligands of TLR10 nin production and inflammatory cytokine induction in
have not been determined, but their SNPs share a correlation human melanocytes.
with asthma in two independent samples. TLR4 and TLR9 In summary, our findings illuminated that HD-13
are associated with wheezing, while TLR4 is associated with accelerated TLR9 expression in 3D4/21 cells, HD-13 acti-
the secretion of allergen-specific IgE. Therefore, the TLR9 vated the inflammatory response via accelerating TLR9
ligand is currently in clinical trials to prevent or treat asthma expression, HD-13 activated the MAPK and NF-κB path-
[2, 4, 8]. TLR9 is the main pattern recognition receptor in way, and HD-13 activated MAPK and NF-κB pathways via
the innate immune system, which selectively recognizes accelerating TLR9 expression. Our data collectively indi-
pathogen-related molecular patterns to initiate innate cated that HD-13 may activate MAPK and NF-κB pathways
immunity and induce the release of inflammatory factors via accelerating TLR9 expression, thereby activating the
[17]. Fang et al.’s study show that TLR2 and TLR4 are inflammatory response in the progression of swine pneumo-
closely associated with a variety of infectious diseases, partic- nia. Our research may provide a theoretical basis for the pre-
ularly lung diseases [18]. Our findings indicated that HD-13 vention and treatment of swine pneumonia.
promoted the levels of TLR9 and its downstream effector,
including MyD88, TRIF, and TRAF6. Besides, HD-13 acti- Abbreviations
vated the inflammatory response, because of the upregula-
tion of IL-1β, IL-6, Il-12, and TNF-α expression in 3D4/21 TLRs: Toll-like receptors
cells. The above results indicate that HD-13 may activate TNF-a: Tumor necrosis factor alpha
the inflammatory response via accelerating TLR9 expression NF-κB: Nuclear transcription factor-κB
in swine pneumonia. MAPK: Mitogen-activated protein kinase
The MAPK pathway is a vital component of innate immu- PRRS: Pattern recognition receptors
nity, which exists in animals, plants, and fungi [19]. The MyD88: Myeloid differentiation factor 88
MAPK pathway is more conservatively involved in mediating TRIF: Toll-like receptor-associated activator of interferon
cells to respond to various stimuli such as heat shock, mitosis, TRAF6: Tumor necrosis factor receptor-associated factor 6
and inflammation and transmits the signal amplification to HD-13: HD-13 strain of Pasteurella multocida D.
the nucleus, further promoting the transcription of cytokine
mRNAs [20]. Wang et al.’s study indicates that activation of Data Availability
MAPK and NF-κB signaling pathways can aggravate pneumo-
nia, myocarditis, and arthritis in piglets [21]. In this paper, our The raw data supporting the conclusions of this manuscript
findings showed that 1 h HD-13 infection significantly will be made available by the authors, without undue reser-
accelerated the levels of MAPK-related proteins, including vation, to any qualified researcher.
p-ERK, p-p38, and p-JNK. The above results indicate that
HD-13 activates the MAPK pathway. What is more, TLR9 Additional Points
silencing inhibited the levels of p-ERK, p-p38, and p-JNK pro-
teins, but HD-13 accelerated their expression, indicating that Highlights. (i) HD-13 activates TLR9 expression in 3D4/21
HD-13 may activate MAPK pathways via accelerating TLR9 cells. (ii) HD-13 activates inflammatory response via acceler-
expression to aggravate the development of swine pneumonia. ating TLR9 expression in 3D4/21 cells. (iii) HD-13 activates
NF-κB is a nuclear transcription factor with multidirec- the MAPK and NF-κB pathway. (iv) HD-13 activates MAPK
tional transcriptional regulation, which plays a vital role in and NF-κB pathways via accelerating TLR9 expression.
the regulation of natural immune response, acquired
immune response, and inflammatory response [22]. It has Conflicts of Interest
been reported that the NF-κB pathway is closely related to
the development of swine pneumonia [21]. Our results indi- There are no conflicts of interest to declare.
cated that HD-13 promoted the levels of NF-κB-related pro-
teins, including p-ERK, p-p38, and p-JNK, indicating that Authors’ Contributions
HD-13 activated the NF-κB pathway. Further results showed
that TLR9 silencing inhibited NF-κB pathway activity, which Jing Liu and Jiayi Xu are the co-first authors.Computational and Mathematical Methods in Medicine 11
Acknowledgments [12] K. K. Asanka Sanjeewa, T. U. Jayawardena, H. S. Kim et al.,
“Fucoidan isolated from Padina commersonii inhibit LPS-
This work was supported by grants from the Natural Sci- induced inflammation in macrophages blocking TLR/NF- κB
ence Basic Research Plan in Shanxi Province of China signal pathway,” Carbohydrate Polymers, vol. 224, article
(2019JM-133) and International Science and Technology 115195, 2019.
Cooperation Programme in Shanxi Province of China [13] J.-H. Cui, S. M. Dong, C. X. Chen et al., “Microplitis bicoloratus
(2018KW-062). bracovirus modulates innate immune suppression through the
eIF4E-eIF4A axis in the insect Spodoptera litura,” Develop-
mental & Comparative Immunology, vol. 95, pp. 101–107,
References 2019.
[14] S.-M. Dong, J. H. Cui, W. Zhang et al., “Inhibition of transla-
[1] J. Lin, S. Chen, K. Yeh, and C. Weng, “Mycoplasma hyorhinis tion initiation factor eIF4A is required for apoptosis mediated
in Taiwan: diagnosis and isolation of swine pneumonia patho- by Microplitis bicoloratus bracovirus,” Archives of Insect Bio-
gen,” Veterinary Microbiology, vol. 115, no. 1-3, pp. 111–116, chemistry and Physiology, vol. 96, no. 3, article e21423, 2017.
2006. [15] I. Maczynska, B. Millo, V. Ratajczakstefanska et al., “Proin-
[2] A. Ando, A. Shigenari, C. Kojima-Shibata et al., “Association flammatory cytokine (IL-1β, IL-6, IL-12, IL-18 and TNF-α)
of swine leukocyte antigen class II haplotypes and immune- levels in sera of patients with subacute cutaneous lupus erythe-
related traits in a swine line selected for resistance to myco- matosus (SCLE),” Immunology Letters, vol. 102, no. 1, pp. 79–
plasmal pneumonia,” Comparative Immunology, Microbiology 82, 2006.
and Infectious Diseases, vol. 48, pp. 33–40, 2016. [16] B. A. Cunha, “Swine influenza (H1N1) pneumonia: clinical
[3] Y. Shimoji, E. Oishi, Y. Muneta, H. Nosaka, and Y. Mori, considerations,” Infectious Disease Clinics of North America,
“Vaccine efficacy of the attenuated Erysipelothrix rhusiopathiae vol. 24, no. 1, pp. 203–228, 2010.
YS-19 expressing a recombinant protein of Mycoplasma hyop- [17] S. Zhang, Q. Zhang, F. Wang et al., “Hydroxychloroquine inhi-
neumoniae P97 adhesin against mycoplasmal pneumonia of biting neutrophil extracellular trap formation alleviates
swine,” Vaccine, vol. 21, no. 5-6, pp. 532–537, 2003. hepatic ischemia/reperfusion injury by blocking TLR9 in
[4] A. L. Vincent, K. M. Lager, B. H. Janke, M. R. Gramer, and J. A. mice,” Clinical Immunology, vol. 216, article 108461, 2020.
Richt, “Failure of protection and enhanced pneumonia with a [18] X. Fang, X. Liu, C. Meng et al., “Breed-linked polymorphisms
US H1N2 swine influenza virus in pigs vaccinated with an of porcine toll-like receptor 2 (TLR2) and TLR4 and the pri-
inactivated classical swine H1N1 vaccine,” Veterinary Microbi- mary investigation on their relationship with prevention
ology, vol. 126, no. 4, pp. 310–323, 2008. against Mycoplasma pneumoniae and bacterial LPS chal-
[5] C. Bourély, G. Cazeau, E. Jouy et al., “Antimicrobial resistance lenge,” Immunogenetics, vol. 65, no. 11, pp. 829–834, 2013.
of Pasteurella multocida isolated from diseased food- [19] M. Takano, T. Takeuchi, S. Kuriyama, and R. Yumoto, “Role of
producing animals and pets,” Veterinary Microbiology, peptide transporter 2 and MAPK signaling pathways in the
vol. 235, pp. 280–284, 2019. innate immune response induced by bacterial peptides in alve-
[6] M. Sahoo, S. Baloni, J. C. Thakor et al., “Localization of Pas- olar epithelial cells,” Life Sciences, vol. 229, pp. 173–179, 2019.
teurella multocida antigens in the brains of pigs naturally [20] G. Zhu, G. Zhao, J. Lin et al., “FCN-A mediates the inflamma-
infected with Pasteurellosis revealing a newer aspect of patho- tory response and the macrophage polarization in Aspergillus
genesis,” Microbial Pathogenesis, vol. 140, article 103968, 2020. fumigatus keratitis of mice by activating the MAPK signaling
[7] Y. Cheng, K. Wang, L. Lin, X. Zhao, Z. Pan, and Z. Zhou, “Dif- pathway,” International Immunopharmacology, vol. 83, article
ferences in pathogenicity and virulence-associated gene 106473, 2020.
expression among Pasteurella multocida strains with high [21] Q. Wang, H. Zhou, Q. Hao, M. Li, J. Liu, and H. Fan, “Coinfec-
and low virulence in a lung tissue model,” Microbial Pathogen- tion with porcine circovirus type 2 and Streptococcus suis sero-
esis, vol. 140, article 103911, 2020. type 2 enhances pathogenicity by dysregulation of the immune
[8] M. Petrocchi-Rilo, C. B. Gutiérrez-Martín, J. I. Méndez- responses in piglets,” Veterinary Microbiology, vol. 243, article
Hernández, E. F. Rodríguez-Ferri, and S. Martínez-Martínez, 108653, 2020.
“Antimicrobial resistance of Pasteurella multocida isolates [22] T. Lawrence, “The nuclear factor NF-κB pathway in inflamma-
recovered from swine pneumonia in Spain throughout 2017 tion,” Cold Spring Harbor Perspectives in Biology, vol. 1, no. 6,
and 2018,” Veterinary and Animal Science, vol. 7, article article a001651, 2009.
100044, 2019.
[9] S. Arora, S. Ahmad, R. Irshad et al., “TLRs in pulmonary dis-
eases,” Life Sciences, vol. 233, article 116671, 2019.
[10] V. Chaurasiya, S. Kumari, S. K. Onteru, and D. Singh, “Up-
regulation of miR-326 regulates pro-inflammatory cytokines
targeting TLR-4 in buffalo granulosa cells,” Molecular Immu-
nology, vol. 119, pp. 154–158, 2020.
[11] B. Zhou, J. Yan, L. Guo et al., “Hepatoma cell-intrinsic TLR9
activation induces immune escape through PD-L1 upregula-
tion in hepatocellular carcinoma,” Theranostics, vol. 10,
no. 14, pp. 6530–6543, 2020.You can also read

















































