Snowdrift game induces pattern formation in systems of self-propelled particles
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

PHYSICAL REVIEW E 104, 044408 (2021)
Editors’ Suggestion
Snowdrift game induces pattern formation in systems of self-propelled particles
Johanna Mayer,1,* Michael Obermüller ,1,* Jonas Denk ,1,2,3 and Erwin Frey 1,†
1
Arnold Sommerfeld Center for Theoretical Physics (ASC) and Center for NanoScience (CeNS), Department of Physics,
Ludwig-Maximilians-Universität München, Theresienstrasse 37, D-80333 München, Germany
2
Department of Physics, University of California, Berkeley, California 94720, USA
3
Department of Integrative Biology, University of California, Berkeley, California 94720, USA
(Received 5 May 2021; accepted 29 July 2021; published 14 October 2021)
Evolutionary games between species are known to lead to intriguing spatiotemporal patterns in systems of
diffusing agents. However, the role of interspecies interactions is hardly studied when agents are (self-)propelled,
as is the case in many biological systems. Here, we combine aspects from active matter and evolutionary game
theory and study a system of two species whose individuals are (self-)propelled and interact through a snowdrift
game. We derive hydrodynamic equations for the density and velocity fields of both species from which we
identify parameter regimes in which one or both species form macroscopic orientational order as well as regimes
of propagating wave patterns. Interestingly, we find simultaneous wave patterns in both species that result from
the interplay between alignment and snowdrift interactions—a feedback mechanism that we call game-induced
pattern formation. We test these results in agent-based simulations and confirm the different regimes of order
and spatiotemporal patterns as well as game-induced pattern formation.
DOI: 10.1103/PhysRevE.104.044408
I. INTRODUCTION e.g., Refs. [10–14]). To the best of our knowledge, however,
there are no studies that deal with the effect of self-propelled
The composition and temporal evolution of microbial as
movement—a characteristic feature of many living beings—
well as other populations depends on a variety of factors,
on the outcome of spatially extended evolutionary games.
including environmental conditions, the population structure,
Since collectives of self-propelled particles (SPP) are known
and the degree of mobility of each species [1–3]. Evolutionary
to generically exhibit intriguing self-organisation phenomena
game theory (EGT) [4,5] has been used as a mathematical
like flocking, clustering, and motility-induced phase separa-
framework to conceptually describe the evolutionary dynam-
tion (for reviews see, e.g., Refs. [15–19]), this will affect
ics of such populations employing methods from nonlinear
the spatial proximity between individuals and thereby their
dynamics [6]. Two-player games, including the prisoner’s
competitive interaction (described as a game). Changes in the
dilemma and the snowdrift game, have played an important
composition of the population could in turn affect the ordering
role in the development of the research field. In EGT, games
processes of the SPPs.
are regarded as a population dynamics problem, where in-
Here, we investigate this interplay of collective effects
dividuals who follow different strategies (“species”) interact
due to self-propelled motion and nonlinear dynamics based
over many generations according to rules set by the respective
on competitive (game theory) interactions. For specificity,
game. The ensuing (nonlinear) dynamic process then deter-
we will address this question for the snowdrift game, one
mines which species survive, i.e., “win the game.”
of the classical two-player games. Using both agent-based
The mobility of individuals in particular has been shown
simulations and a hydrodynamic approach, we discover a
to play a crucial role in the evolutionary success of a species.
phenomenon that we term game-induced pattern formation:
Previous theoretical studies have modeled mobility either as
When one of the two species starts to exhibit collectively
a diffusive process, in which individuals perform random
moving patterns due to self-propelled motion and alignment
walks (see, e.g., Refs. [7–9]) or alternatively as a migration
of its agents, a pattern is induced in the second species. We
process, in which individuals move by winning a competition
attribute this to a nonlinear feedback mechanism between the
with other species that are spatially adjacent to them (see,
local snowdrift game interaction and the alignment between
particles.
Upon exploring the model parameters that determine the
*
These authors contributed equally to this work. alignment and the interaction probability, we find that the
†
frey@lmu.de system exhibits five distinct phases, each with qualitatively
different collective behavior. If the game interaction is suffi-
Published by the American Physical Society under the terms of the ciently weak, then we find that reducing the noise level leads
Creative Commons Attribution 4.0 International license. Further to a phase transition from a disordered phase to an ordered
distribution of this work must maintain attribution to the author(s) phase in which one species induces a polar wave pattern in the
and the published article’s title, journal citation, and DOI. second species. These patterns vanish when the noise strength
2470-0045/2021/104(4)/044408(24) 044408-1 Published by the American Physical Society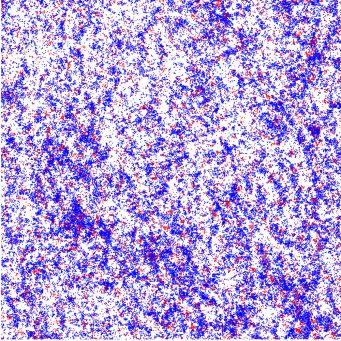
MAYER, OBERMÜLLER, DENK, AND FREY PHYSICAL REVIEW E 104, 044408 (2021)
is reduced, leading to a partially ordered phase in which one two strategies. These include the prisoner’s dilemma, the
species shows uniform polar order while the second species coordination, and the harmony game. Their dynamics in a
is disordered. A further reduction of the noise strength then well-mixed environment (where diffusion is much faster than
also leads to an ordering transition in the second species, the reaction kinetics) can be formulated in terms of concepts
which first induces a wave pattern in the previously uniformly from nonlinear dynamics [6]. The main difference between
ordered species and finally also shows uniform polar order. these four games is their evolutionary outcome, i.e., the com-
All of these phases are generalizations of the phases observed position of the population in the long run. While, for example,
in single-species systems of SPPs, and we show that their in the snowdrift game, cooperators and defectors coexist,
existence depends on the interplay between alignment and defectors always dominate in the prisoner dilemma and coop-
interspecies interactions. erators go extinct. This is related to the rules of the prisoner’s
This paper is organized as follows: In the remainder of the dilemma, which can be formulated as a public goods game
introduction we give a concise overview over previous work where an individual can either cooperate by providing some
on two-player games in a well-mixed environment and in kind of public good or defect by exploiting this public good.
spatially extended systems. This is mainly intended to inform Under these conditions, defectors will prevail as they can
readers in the active matter field not familiar with this topic benefit from the interaction with cooperators without paying
about those results relevant for the present study. In Sec. II any costs.
we first summarize the Boltzmann approach for SPPs and the
dynamics of the snowdrift game to then combine both models B. Evolutionary games in spatially extended systems
to find equations governing the dynamics of two SPP species
with interspecies snowdrift game interaction. Section III takes The spatial extension of a population and the ensuing spa-
these equations as a starting point to derive simpler hydro- tiotemporal arrangement of individuals can strongly influence
dynamic equations and analyses them using both analytical the evolutionary dynamics [22–25]. It is well known that this
and numerical methods. In this section we obtain the phase plays an important role for microbial life, as many microbes
diagram of the system in the space spanned by parameters on our planet are found in the form of dense microbial com-
of self-propelled motion and game interaction and discuss munities such as colonies and biofilms [23,26].
their properties. The results of the hydrodynamic approach A key factor for the dynamics is the spatial clustering of
and the overall phenomenology of the system are qualitatively individuals, which has already been argued by Hamilton to
validated in Sec. IV by agent-based simulations. We conclude support cooperation in populations [27], because cooperat-
with a summary and discussion of our results in Sec. V. ing individuals are more likely to interact with each other
and therefore are less likely to be exploited by defectors. To
what degree this determines the composition of a population
A. Two-player public good games has been studied extensively on different kinds of lattices as
Originally, the “snowdrift” game (also known as “hawk- well as for different kinds of interaction rules [8,10–12,28–
dove” game) was formulated as a strategic “cooperator- 34]. Specifically, for the snowdrift game, it was shown that
defector” game [20]. It owes its name to its prominent accounting for spatial extension can change population ratios
explanatory framework: Two drivers are caught in a winter compared to well-mixed systems [29,35].
storm and a snowdrift is blocking the road. Both drivers now While all the studies mentioned above consider diffusive
have the option of either remaining in the car or getting out dynamics, self-propulsion is an essential part of biological
and removing the snowdrift. Since both drivers want to con- reality. In fact, there are a large variety of microbes that show
tinue their journey, but getting out is not really desirable, the directed mobility, often along chemotactic gradients [36–39]
best strategy for each driver is to do exactly the opposite of or due to chemical warefare [40]. Here, motility is an impor-
what the other driver does. tant factor of their strategy, e.g., for directed motion away
This strategic game can be transferred to the field of EGT from hostile environments, chemotaxis or self-organisation
and microbial systems such as yeast cells competing for into flocks.
resources and using different survival strategies [21]: In a
metabolically costly process, part of the yeast cells (producer II. MODEL DESCRIPTION
cells) convert sucrose to glucose, which it can use as an energy
A. Active matter model
source. This process yields more glucose than can be metab-
olized, and the extra glucose diffuses into the surrounding To implement propulsion of particles of both species, A
medium. There it can be used by all yeast cells, regardless and B, we employ a kinetic Boltzmann approach [41,42].
of whether they produce glucose or not. As long as there is an Here, particles with a diameter d0 move ballistically on a two-
excess of glucose in the system, it is therefore advantageous to dimensional substrate with constant speed v0 and orientation
save the metabolic costs for its production and only consume θ . Ballistic motion is interrupted by stochastic self-diffusion
it (nonproducer cells). In contrast, when there is a lack of events: Particles change their orientation by an angle η at a
glucose, it becomes advantageous to produce glucose as this rate ω to
provides an energy source. This leads to the coexistence of θ = θ + η, (1)
producer (cooperator) and nonproducer (defector) cells in the
yeast population. where η is a stochastic variable (noise); see Fig. 1(a). Hence,
The snowdrift game is one out of four possible two-player the particles move in a run-and-tumblelike fashion with a
games, i.e., games where the individuals can adopt one of mean time ω−1 between tumbling events.
044408-2
SNOWDRIFT GAME INDUCES PATTERN FORMATION IN … PHYSICAL REVIEW E 104, 044408 (2021)
FIG. 1. Illustration of self-propelled dynamics: (a) Particles
change their orientation θ by a stochastic angle η to θ = θ + η FIG. 2. Illustration of the snowdrift game. (a) Payoff matrix:
during self-diffusion events. (b) Binary collisions between particles There is a nonzero payoff only if individuals belonging to different
of the same species lead to polar half-angle alignment of their ori- species meet. Then, A receives a payoff τ , while B receives a payoff
entations. We allow for additional stochastic noise terms η1 and η2 λτ . The relative fitness λ and game strength τ serve as control
during the collision event. parameters. (b) Flow diagram of the replicator equation, Eq. (7), for
species A with population density a: Unstable fixed points are marked
Further, we assume that intraspecies interactions are due with an open circle, while the stable fixed point at a0 is marked with
to binary collisions between particles, that lead to alignment a filled circle. The arrows indicate the flow toward the stable fixed
of their orientations, while interspecies interactions follow a point.
snowdrift game (see the following Sec. II B). We implement
intraspecies aligning collisions in terms of half-angle align-
ment rules; see Fig. 1(b). The particle orientations after a Here, a and b denote the densities of species A and B,
collision event are given by respectively. In the following, we will refer to λ as the relative
fitness parameter of particles of species B relative to A, and τ
θ1 + θ2 θ1 + θ2 as the game strength.
θ1 = + η1 , θ2 = + η2 , (2)
2 2 The time evolution now follows “survival of the fittest”:
where we allowed for additional noise terms η1 and η2 . We The species with higher fitness value increases in population
have chosen these rules to keep our analysis conceptually size, while the other species decreases in number. Specifically,
simple. In actual systems one would need to account for the the dynamics of the densities a and b are given by the replica-
problem-specific interaction between the agents of the active tor equations:
system. This has, for example, been achieved for the actin-
∂t a = ( fA − fB )b a, (5a)
myosin motility assay [43–45]. For simplicity, diffusion and
collision noise variables are drawn from a Gaussian distribu- ∂t b = ( fB − fA )a b. (5b)
tion with the same standard deviation σ .
For a dilute system of only one species of propelled These replicator equations govern the dynamics of the
particles, Bertin et al. [41,42] proposed a kinetic Boltz- game. The fitness difference ( fA − fB ) indicates the relative
mann equation. It describes the dynamics of the one-particle strength of the interaction partner, while the factor a b encodes
probability density function, f (r, θ , t ), which indicates the the probability that particles of species A and B meet and in-
probability that a particle is located at position r with the teract with each other. Note that the overall density ρ = a + b
orientation θ at time t. The kinetic Boltzmann equation for is constant. Inserting the expressions for the fitness values fA
self-propelled particles for one species reads [41,42] and fB , Eqs. (4), into Eqs. (5) one can rewrite the replicator
equations as
∂t f (r, θ , t ) + v0 eθ · ∇ f = Idiff [ f ] + Icoll [ f , f ], (3)
∂t a = τ (b − λa)a b, (6a)
where Idiff and Icoll denote contributions resulting from dif-
fusion and collision events, respectively, while eθ is the unit ∂t b = τ (λa − b)a b. (6b)
vector pointing in θ direction. Their explicit expressions can The meaning of the terms relative fitness and game strength
be found in Appendix A 1. In the present context, the most for λ and τ , respectively, now become evident: If b > λa, then
important feature of Idiff and Icoll is that they depend on the there is a fitness advantage for species A and its density grows
parameter σ , which characterizes the noise strength. relative to species B. In contrast, if b < λa, then the growth of
species B is favored relative to species A. Since the equations
B. Snowdrift game are antisymmetric under the exchange of species A and B, we
In addition to the alignment interactions, we assume that can, without loss of generality, impose that species A is always
particles during collisions also play a snowdrift game. The fitter than species B, i.e., constraining λ ∈ [0, 1]. By changing
outcome of this interaction is specified by the payoff matrix λ we can tune the relative fitness from the two species being
[Fig. 2(a)], which is nonzero only if two different species equally competitive (λ = 1) to species A being strongly domi-
meet. The fitness of a particular strategy, A or B, is defined nant over B (λ 1). The second game parameter, τ ∈ [0, ∞),
as a constant background fitness (set to 1) plus the average can be understood as the game strength.
payoff obtained from playing the game [6,46]: Since the total particle number is conserved (b =ρ − a),
Eqs. (6) can be reduced to the temporal evolution of a single
fA = 1 + τ b, (4a) species (say A):
fB = 1 + τ λ a. (4b) ∂t a = τ (b − λa)ab = τ [(ρ − a) − λa]a(ρ − a). (7)
044408-3
MAYER, OBERMÜLLER, DENK, AND FREY PHYSICAL REVIEW E 104, 044408 (2021)
Figure 2(b) shows the flow diagram of Eq. (7) with arrows dilute system of two species of self-propelled particles with
pointing to the right for increasing a (∂t a > 0) and to the left a snowdrift interaction. It includes convection, self-diffusion,
for ∂t a < 0. There are three fixed points (stationary points with intraspecies binary collision (with polar alignment), as well as
∂t a = 0): a snowdrift interaction between particles. As solving the ki-
ρ netic Boltzmann equations, Eqs. (9), analytically is generally
aB = 0, aA = ρ, and a0 = . (8) impractical, one has to resort to numerical implementations
1+λ
[45,47,48]. However, as exemplified by studies of the kinetic
The fixed points aB and aA are unstable in the snowdrift game, Boltzmann equation for a single species [41,42,48–51], one
while a0 is stable. For aB = 0 and aA = 0 species A and B go can also use Eqs. (9) to derive hydrodynamic equations for
extinct, respectively, and therefore the game ends (absorbing those collective variables that vary slowly in space and time.
fixed points). The fixed point a0 refers to a stable coexistence In the next section, we first derive these hydrodynamic
of both species with population ratio b/a = λ. equations for the system of two species, Eqs. (9), and then
study them analytically for spatially uniform systems. Then
C. Active matter snowdrift game we investigate spatially nonuniform solutions using linear sta-
To describe our system of two interacting self-propelled bility analysis of the uniform states and numerical solutions
particle species, we introduce the one-particle probability den- of the full hydrodynamic equations.
sity functions α(r, θ , t ) and β(r, θ , t ), governing the temporal
evolution of species A and B, respectively. We use the ki- A. Derivation of hydrodynamic equations
netic Boltzmann equation, Eq. (3), and extend it for each As the probability densities α(r, θ , t ) and β(r, θ , t ) are
species separately by snowdrift interaction terms, Igame A
[α, β] periodic functions in θ , it is convenientto work with their
and Igame [β, α], chosen in analogy to the replicator equations,
B π
Fourier modes, defined as αk (r, t ) := −π dθ eiθk α(r, θ , t )
Eqs. (6). This leads to the general form of the two-species π
and βk (r, t ) := −π dθ eiθk β(r, θ , t ). Note that the zeroth
Boltzmann equations Fourier modes are the densities of the respective species,
∂t α+v0 eθ ∇α = Idiff [α]+Icoll [α, α]+Igame
A
[α, β], (9a) α0 = a, β0 = b, defined by Eq. (10). Besides the densities of
species A and B, the second set of slow, hydrodynamic vari-
∂t β+v0 eθ ∇β = Idiff [β]+Icoll [β, β]+Igame
B
[β, α]. (9b) ables are the polar order fields PA (r, t ) and PB (r, t ), which are
related to the first Fourier modes
We assume snowdrift interactions to be local and inde-
pendent of particle orientation as in Eqs. (6). With the local Re(α1 ) Re(β1 )
a PA = , b PB = . (12)
densities a(r, t ) and b(r, t ) of species A and B, related to α Im(α1 ) Im(β1 )
and β by
π π Hence, the modes α1 and β1 are a measure for the polar order
a(r, t ) = dθ α(r, θ , t ), b(r, t ) = dθ β(r, θ , t ), multiplied by the density at a given point in space and time of
−π −π the system. If α1 = β1 = 0, then the system is disordered. If α1
(10) or β1 is nonzero, then there is a preferred direction of motion,
meaning the respective species moves collectively. We will
the snowdrift interaction terms read
refer to α1 and β1 in the following as velocity fields.
A
Igame [α, β] = τ (b − λa) b α, (11a) The expansion of the kinetic Boltzmann equations,
Eqs. (9), in Fourier modes is given by (see Appendix A for
B
Igame [β, α] = τ (λa − b) a β. (11b) details)
The dependency on α and β in Eqs. (11a) and (11b), 1
∂t αk + ∇αk−1 + ∇ ∗ αk+1 = − 1 − e−(kσ ) /2 αk
2
respectively, is chosen to match the form of the Boltzmann 2
equation with its dependency on the one-particle densities α
and β. The game strength τ tunes the relative weight of the + αk−p α p I p,k + τ (b − λa) b αk , (13a)
p
snowdrift interaction compared to the alignment interaction
(collision terms). If one sets τ = 0, then the snowdrift inter- 1
∂t βk + ∇βk−1 + ∇ ∗ βk+1 = − 1 − e−(kσ ) /2 βk
2
action terms vanish and Eqs. (9) reduce to two independent 2
equations for species A and B. However, in the absence of con-
vection (v0 = 0), diffusion (Idiff = 0), and collision (Icoll = 0), + βk−p β p I p,k − τ (b − λa) a βk , (13b)
p
one recovers the replicator Eqs. (6) by integrating Eqs. (9)
over the angle θ . where we introduced the abbreviations ∇ = ∂x + i∂y and
In the following, time, space, and densities are rescaled ∇ ∗ = ∂x − i∂y . The explicit expressions for the collision
such that one measures time in units of 1/ω, position in kernels I p,k (σ ) are given in Appendix A. Equation (13) con-
units of v0 /ω and spatial densities in units of ω/(v0 d0 ). This stitutes an infinite set of differential equations that couple
amounts to setting v0 = d0 = ω = 1. lower- with higher-order Fourier modes. For k = 0, Eqs. (13)
yield
III. HYDRODYNAMIC THEORY
∂t a = − 21 (∇α1∗ + ∇ ∗ α1 ) + τ (b − λa) b a, (14a)
In the previous section, we have formulated an extended
version of two coupled kinetic Boltzmann equations for a ∂t b = − 21 (∇β1∗ ∗
+ ∇ β1 ) + τ (λa − b) a b. (14b)
044408-4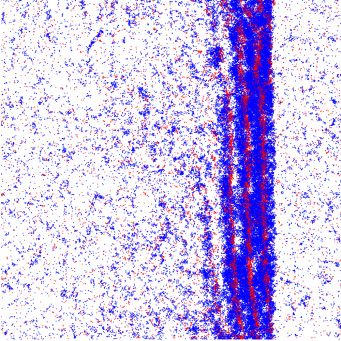
SNOWDRIFT GAME INDUCES PATTERN FORMATION IN … PHYSICAL REVIEW E 104, 044408 (2021)
The first terms in Eqs. (14) can be understood as the
temporal change of the densities due to spatial variations in
the velocity fields, while the last terms can be interpreted
as source or sink terms given by the replicator equations,
Eqs. (6). These source terms couple the densities of species A
and B and we will refer to them in the following as snowdrift
terms. Note, that the sum of both snowdrift terms is zero
at every point in space and time. Hence, the overall density
ρ = a + b follows a continuity equation
∂t ρ = − 21 [∇(α1∗ + β1∗ ) + ∇ ∗ (α1 + β1 )]. (15)
The spatial average of the density, ρ̄, is therefore a conserved
quantity and a control parameter of the system.
For a spatially uniform system, the spatial derivatives in FIG. 3. Solutions of the spatially uniform equations for the ve-
the equations for the particle densities, Eqs. (14), vanish and locity fields α1 and β1 [see Eqs. (21)]. (a) Amplitudes of the solutions
Eqs. (14) reduce to the replicator equations. The correspond- as functions of the noise strength σ with the relative fitness set to
ing steady states are given by the stable snowdrift fixed point λ = 1, λ = 0.6, and λ = 0.2, as indicated in the graph. Solid lines are
with a(0) = ρ̄/(1 + λ) and b(0) = ρ̄λ/(1 + λ). Hence, a state stable solutions, dashed lines unstable solutions. If the noise strength
with uniform densities a = a(0) and b = b(0) and all higher is smaller than a threshold value, σ σtA (σ σtB ), then the disor-
Fourier modes vanishing is a solution to Eqs. (13). This so- dered solution |α1(0) | = 0 (|β1(0) | = 0) becomes unstable and a new
lution describes a spatially uniform, disordered system with stable solution with |α1(0) | > 0 (|β1(0) | > 0) appears. (b) Bifurcation
(phase) diagram for the spatially uniform solution in control param-
densities set to their snowdrift fixed points. Small spatially
eter space (λ, σ ). At σtA (λ), there is a transitions from a disordered
uniform perturbations δαk , δβk to this solution evolve to linear
state (fixed point |α1(0) | = |β1(0) | = 0) to a broken symmetry state with
order as
macroscopic polar order in A only (|α1(0) | > 0, |β1(0) | = 0). A second
∂t δαk = μAk (a(0) , b(0) ) δαk , (16a) transition to a state with polar order in both species (|α1(0) | > 0 and
|β1(0) | > 0) occurs at σtB (λ) σtA (λ). The threshold values for the
∂t δβk = μBk (a(0) , b(0) ) δβk , (16b) noise strengths coincide if the relative game strength λ = 1.
with
σ2 Bertin et al. [41] and use the following truncation scheme: We
μAk (a, b) = e− 2 −1 + (I0,k +Ik,k )a + τ (b−λa)b, (17a) assume that close to the onset of polar order the velocity fields
σ2 are small, α1 , β1 1, and obey the scaling relations (a −
μBk (a, b) = e− 2 −1 + (I0,k +Ik,k )b + τ (λa−b)a. (17b) a(0) ) ∼ α1 , αk ∼ α1|k| (and analogously for βk ). Furthermore,
we assume that spatial and temporal variations ∇, ∂t ∼ α1 are
The respectively first two terms in μAk and μBk are due to small. With these assumptions, one can truncate and close
the self-propelled motion of both species and are familiar the Boltzmann equation in Fourier space, Eq. (13), at the
expressions from one-species SPP systems [41,42]. Here, we third order term for the velocity field, α13 . We refer the inter-
encounter an additional term due to the snowdrift game, which ested reader to Appendix A and Refs. [41,42,49] for a more
implies that the growth rate of every Fourier mode depends on detailed discussion on the truncation scheme. Following the
the densities of both species. When we insert the stable solu- described procedure, we obtain hydrodynamic equations for
tion for the densities, a(0) = ρ̄/(1 + λ) and b(0) = ρ̄λ/(1 + λ), the velocity fields of species A and B:
into the expressions for μAk and μBk , the snowdrift terms van-
ish and we are left with the first two terms, which are then ∂t α1 = μA1 (a, b) − ξ (a)|α1 |2 α1 + ν(a) ∇ ∗ ∇α1
functions of σ , ρ̄ and λ. Varying these parameters, one finds
that all Fourier modes μAk and μBk with k 2 are negative −γ (a) α1 ∇ ∗ α1 − κ (a) α1∗ ∇α1 − 21 ∇a, (18a)
and only the growth rates for the first Fourier mode, μA1 and ∂t β1 = μB1 (a, b) − ξ (b)|β1 |2 β1 + ν(b) ∇ ∗ ∇β1
μB1 , can become positive, meaning that small perturbations
in the velocity fields α1 or β1 , respectively, will grow ex- − γ (b) β1 ∇ ∗ β1 − κ (b) β1∗ ∇β1 − 21 ∇b. (18b)
ponentially. For fixed overall density, ρ̄, and relative game For explicit expressions of the coefficients please refer
strength, λ, the growth rates μA1 and μB1 are negative for high to Appendix A. The essential difference between Eqs. (18)
values of the noise strength σ and become positive when the and the hydrodynamic equation for the velocity field of one-
noise strength is low enough. This defines threshold values species SPP [41] lies in the coefficients μA1 and μB1 , which here
for the noise strength, σtA (λ, ρ̄) and σtB (λ, ρ̄), at μA1 |σtA ,ρ̄,λ = 0 couple the velocity fields to the density of the respective other
and μB1 |σtB ,ρ̄,λ = 0, respectively [see Fig. 3(b)]. Below these species [see Eqs. (17)]. Note that the snowdrift interaction
thresholds (σ < σtA or σ < σtB ), the spatially uniform solution term only appears in the term linear in the velocity fields in
is unstable and small perturbations in the velocity fields, α1 or Eqs. (18). This is a consequence of the truncation scheme and
β1 , have positive growth rates. is explained in more detail in Appendix A.
Close to the threshold values σtA and σtB , a weakly non- Taken together, Eqs. (14) and (18) constitute a minimal set
linear analysis yields further insights into the dynamics of of hydrodynamic equations that describe the dynamics of the
the system and the resulting steady states. Here we follow slow variables of our system, the densities and velocity fields
044408-5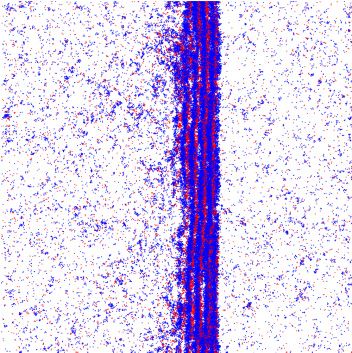
MAYER, OBERMÜLLER, DENK, AND FREY PHYSICAL REVIEW E 104, 044408 (2021)
of both species. Overall, we identify four control parameters: For values of the noise strength below the respective
the noise strength σ , the overall density ρ̄, the game strength threshold, the solution α1 = β1 = 0 is linearly unstable and
τ , and the relative fitness λ. For specificity, we use a fixed additional, stable solutions appear, given by
value for the average density, ρ̄ = 1 [in units of ω/(v0 d0 )]
and vary the remaining parameters. We observe the same μA1 iθa
α1(0) = e for σ < σtA , (22a)
phenomenology if we instead fix σ and use ρ̄ as control ξ
parameter [52] (this is due to the opposing roles of density and
noise changes in the self-organization of active matter systems μB1 iθb
[16,17,19]). β1(0) = e for σ < σtB . (22b)
ξ
In the next sections we will first determine solutions for the
spatially uniform case of Eqs. (14) and (18) and subsequently These solutions correspond to collective states which spon-
analyze their linear stability against spatially nonuniform per- taneously break the rotational symmetry of the system by
turbations. choosing random directions of macroscopic order θa and θb ,
respectively. Figure 3(a) shows the absolute values (ampli-
tudes) |α1(0) | and |β1(0) | as functions of the noise strength σ for
B. Spatially uniform solutions different values of λ. For equally competitive species, λ = 1,
The stationary uniform solutions of the hydrodynamic the densities are the same for both species: a(0) = b(0) = 1/2
equations, Eqs. (14) and (18), can be found by setting all tem- [see Eqs. (20)]. In this case the growth rates μA1 (a(0) , b(0) ) and
poral and spatial derivatives to zero. Then, the equations for μB1 (a(0) , b(0) ) are identical for all values of the noise strength
the densities and velocity fields decouple. For the densities, σ [see Eqs. (17)]. Thus, the amplitudes of the velocity fields,
Eqs. (14), one obtains |α1(0) | and |β1(0) |, are also identical as shown in the upper panel
of Fig. 3. When σ is decreased below σtA = σtB , the ampli-
∂t a = τ (b − λa) a b, (19a) tudes of the velocity fields become nonzero, which means that
both species exhibit spatially uniform macroscopic order. For
∂t b = τ (λa − b) a b. (19b) λ < 1, the amplitudes |α1(0) | and |β1(0) | differ. Since a relative
game strength λ < 1 indicates that species A is “stronger” than
These are precisely the replicator equations, Eqs. (6), with species B, its stationary density (fixed point) is larger than
the stable solution (note we have set ρ̄ = 1) that of species B: a(0) > b(0) . As a consequence it has the
larger growth rate, μA1 > μB1 , and thus polar order of A is less
1 λ prone to noise resulting in a larger threshold value, σtA > σtB ,
a(0) = , b(0) = . (20)
1+λ 1+λ see Figs. 3(b) and 3(a) for λ = 0.6 and λ = 0.2, respectively.
This underlines an important aspect of the game interactions.
Hence, the spatially uniform densities of each species are
These determine the relative density of the species A and B,
determined by the relative fitness λ, and are independent of
which directly affects the ability of these species to establish
the game strength τ .
macroscopic polar order, i.e., the threshold values of the noise.
The hydrodynamic equations for the velocity fields,
In summary, for our two-species system with game inter-
Eqs. (18), of a spatially uniform system are given by
actions, the spatially uniform solutions show a new aspect of
macroscopic order that is absent in one-species SPP systems
∂t α1 = μA1 − ξ |α1 |2 α1 , (21a) [41,42,49]. There is not only one transition between a polar-
∂t β1 = μB1 − ξ |β1 |2 β1 . (21b) ordered and a disordered state at some threshold value for the
noise strength but two successive transitions. The underlying
Hence, α1 = β1 = 0 (corresponding to a disordered state) is reason is that the competition of the two species, as defined
always a trivial solution. A nontrivial solution occurs when by the (snowdrift) game, implies that their stationary densities
the terms in the brackets balance each other. Since ξ > 0 for are different: the species with the lower fitness also has a lower
all values of the control parameters (see Appendix A), such a density. As the competition between alignment, favoring polar
solution for α1 (β1 ) exists for μA1 > 0 (μB1 > 0). We know from order, and noise, favoring disorder, crucially depends on the
the last section that the growth rate μA1 (μB1 ) is negative at high particle density, the noise thresholds for the ordering transition
values of the noise strength σ and becomes positive below the of the two species are different. The more competitive species
threshold value σtA (σtB ), which depends on the relative fitness has a higher density and, therefore, orders at a higher noise
λ, as shown in Fig. 3(b). Note that the threshold values for strength than the less competitive species. This is summarized
the noise strength are different for species A and B because by the phase diagram shown in Fig. 3(b). By varying the
their densities are different in steady state for games with noise strength σ and the relative fitness λ there are three
λ = 1. The “weaker” species (here B) has a lower density and phases: a disordered phase above σtA , an intermediate phase
hence is more prone to the disordering effect of noise, i.e., for σtA > σ > σtB where only the more competitive species,
σtB σtA . Moreover, all results are independent of the game A, shows polar order, and a phase where both species exhibit
strength τ , since the terms involving τ in μA1 and μB1 [see polar order below σtB .
Eqs. (17)] cancel as the densities reach their spatially uniform
steady state solutions a(0) and b(0) given by Eqs. (20). The C. Stability against spatially nonuniform perturbations
game strength τ will, however, play an important role for the In this section we will test the linear stability of the spa-
dynamics of spatially nonuniform systems. tially uniform solutions given by Eqs. (20) and Eqs. (22),
044408-6
SNOWDRIFT GAME INDUCES PATTERN FORMATION IN … PHYSICAL REVIEW E 104, 044408 (2021)
against perturbations with a finite wavelength. To this end,
we introduce spatially nonuniform perturbations around the
uniform solutions
a(r, t ) = a(0) + δa(r, t ), α1 (r, t ) = α1(0) + δα1 (r, t ),
(23)
b(r, t ) = b(0) + δb(r, t ), β1 (r, t ) = β1(0) + δβ1 (r, t ).
Assuming that the perturbations are small, we insert the per-
turbed fields in the full hydrodynamic equations, Eqs. (14)
and (18), and keep only terms up to linear order in the per-
turbation. Note that one has to account for both perturbations
δα1 , δβ1 and its complex conjugates
∞ δα1∗ , δβ1∗ . Introducing
Fourier modes δa(r, t ) = 2π −∞ dq e δa(q, t ) and analo-
1 iq·r
gously for all other fields, one obtains a set of linear equations
for δmq = [δa(q), δb(q), δα1 (q), δα1∗ (q), δβ1 (q), δβ1∗ (q)]T ,
∂t δmq = Jq δmq , (24)
with the Jacobian Jq . It is a 6×6 matrix that depends
on the wave vector q and the spatially uniform solution
(a(0) , b(0) , α1(0) , β1(0) ) and therefore on all control parameters
(λ, τ, σ ). An explicit expression for Jq can be found in
Appendix B. For simplification, we assume that the spa-
tially uniform solutions of the velocity fields are parallel,
α1(0) β1(0) . Numerical solutions of the hydrodynamic equa-
tions and agent-based simulations in the next sections will FIG. 4. Stability analysis of the hydrodynamic equations
show that this assumption is justified. Eqs. (14) and (18). (a) Dispersion relation of the eigenvalue with
The real parts of the eigenvalues of the Jacobian Jq the largest real part, sq , for different values of the noise strength
determine the linear growth rate of the perturbations: An σ , and fixed game strength τ = 0.2 and relative fitness λ = 0.4. sq
eigenvalue with a positive real part indicates that the cor- is plotted for wave vector q pointing in the direction parallel to
responding uniform solution is linearly unstable and the macroscopic order, q α1 , β1 (denoted by q ). The spatially uniform
perturbation given by the respective eigenvector of the Ja- state, Eqs. (22), is unstable when the maximum of the dispersion re-
cobian will grow (exponentially) in time. Figure 4(a) shows lation, smax , is larger zero. (b) Parameter regime (bands) of λ (relative
examples of the real part of the eigenvalue with the largest real fitness) and σ (noise strength) where the spatially uniform polar-
part, sq , as a function of q for different values of σ and fixed ordered states are linearly unstable against spatial perturbations (i.e.,
λ = 0.4 and τ = 0.2. For certain values of the noise strength smax > 0), for different values of the game strength τ (shades of blue)
as indicated in the graph. The two bands are bounded from above
σ there are long wavelength instabilities (i.e., for small values
by the threshold values σtA and σtB indicating the onset of spatially
of the wave vector q).
uniform polar order for species A and B, respectively (dashed lines).
Our linear stability analysis shows that the spatially uni-
The width of the bands decreases with increasing game strength
form ordered states are unstable with respect to spatial τ . White regions indicate parameter regimes where the spatially
perturbations right at the onsets of both transitions to macro- uniform ordered states are stable against spatial perturbations. (c, d)
scopic order, σtA and σtB [Fig. 4(b)]. In other words, the Linearly unstable regions in (τ, σ ) space and (λ, σ ) space, respec-
disordered phase (σ > σtA ) is stable, while the phases with tively. The color code compares the two density field components
spatially uniform polar order in species A (σtA > σ > σtB ) (δa and δb) of the eigenvector corresponding to the eigenvalue with
and uniform polar order in both species (σtB > σ ) are both the largest positive real part: δa := δa/(δa + δb) which ranges from
unstable toward spatial perturbations for σ close enough to 0 (the largest linear growth rate corresponds to an eigenvector with
the respective onset of order. By computing the real part δa component equal to zero and a nonzero δb component) to 1 (δb
of the largest eigenvalue, sq , for different wave vectors q, component equal to zero and nonzero δa component).
we find that sq is largest for wave vectors parallel to the
direction of macroscopic order (q α1(0) , β1(0) ); i.e., instabil-
ities are strongest when longitudinal. This suggests that the that the snowdrift interaction has a stabilizing effect on the
ensuing patterns will form along the direction parallel to spatially uniform solutions.
macroscopic order, α1(0) and β1(0) , while the system remains ho- The perturbation with the largest linear growth rate is
mogeneous along the direction perpendicular to macroscopic given by the eigenvector δmq corresponding to the eigen-
order. For τ = 0, i.e., in the absence of snowdrift interactions, value sq with the largest positive real part (and with wave
we recover the same unstable regions in parameter space as vector q fixed such that sq is maximal); see Eq. (24). We
previously obtained from one-species SPP systems [41,42] compute this eigenvector as a function of the three control
[see Fig. 4(b)]. For a finite game strength, τ > 0, the upper parameters σ, λ and τ to see how the interplay of snowdrift
boundaries of the unstable regions in parameter space are still game and self-propelled motion affects which perturbations
σtA and σtB (as in the case of no snowdrift interaction). The have the largest growth rate. To compare the growth rates of
width of the unstable regions, however, is smaller indicating the fields of the two species, we first look at the two density
044408-7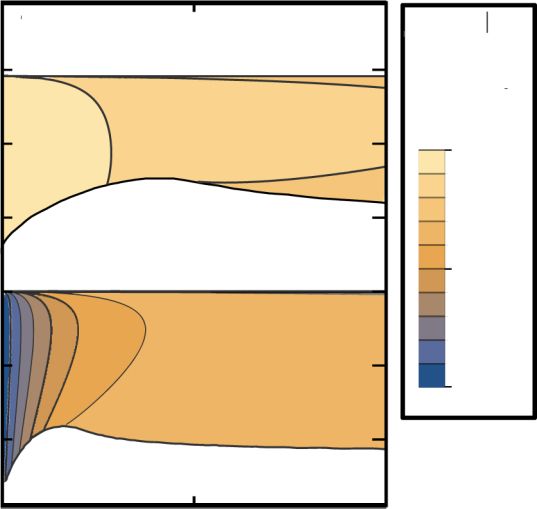
MAYER, OBERMÜLLER, DENK, AND FREY PHYSICAL REVIEW E 104, 044408 (2021)
field components of the eigenvector, δa and δb. We use the
expression δa := δa/(δa + δb) to measure the direction of the
eigenvector projected onto the plane spanned by the two den-
sity fields. This value can range from 0 (δa = 0 and nonzero
δb component), meaning that only perturbations in the density
of species B grow, to 1 (δb = 0 and nonzero δa component)
meaning that only the density of species A is unstable toward
perturbations. Figures 4(c) and 4(d) show the value of δa as a
function of the control parameters in (τ, σ ) and (λ, σ ) space,
respectively. Analyzing the results in (τ, σ ) space [Fig. 4(c)],
we can read off how the game strength, τ , affects the relative
growth rates of perturbations of the two densities, a and b: For
large enough game strength, τ , perturbations in the densities
of both species grow equally fast (δa → 0.5). Decreasing τ
changes the value of δa dependent on the region in parameter
space: In the unstable region with upper boundary σ = σtA ,
perturbations in the density of species A are growing faster FIG. 5. Phase diagram in control parameter space (λ, σ ) for
than perturbations in the density of species B such that δa → 1 game strength τ = 0.2. Colored regions indicate parameter regimes
for small enough game strength τ [see Fig. 4(c)]. In other where one finds phases that exhibit traveling polar waves, traveling
words, the perturbation with the largest growth rate changes polar wave phase A (TWA) and traveling polar wave phase B (TWB).
from an even orientation in the directions of both types to an White regions indicate regimes showing spatially uniform phases.
orientation only in the a direction. In the unstable region with Regions of linear instability are in yellow-brown (depending on the
upper boundary σ = σtB , δa → 0 for small enough τ , which value of the maximum of the largest real part of the eigenvalue of
means that only the density of species B is unstable. The inset the Jacobian, smax ). Bistable regions at the transitions between phases
of Fig. 4(d) shows the limiting case τ = 0 in the (λ, σ ) plane. are coloured in blue. Blue dots are calculated numerically and indi-
From the color code we read off that, depending on the region, cate transitions between spatially uniform phases, bistable regions
either δb = 0 (yellow region) or δa = 0 (blue region). In other and traveling wave phases. The blue star indicates the vicinity of
words, for two noninteracting species (τ = 0), perturbations a small parameter regime, where the transition between the par-
only grow in either species A or B. Note that in this case, tially polar-ordered uniform phase and the TWA phase becomes
supercritical.
instabilities in the density fields for each species occur at the
respective threshold to macroscopic order: perturbations δa
grow at σ < σtA ; perturbations δb grow at σ < σtB . will study the spatial patterns that emerge in these unstable
There are four more components to the eigenvector, en- regions.
coding the perturbations of the velocity fields. An analogous
analysis as above for the velocity field components par-
allel to macroscopic order, δα1 and δβ1 , yields the same D. Numerical solutions of the hydrodynamic equations
phenomenology as for the densities: For high values of τ , per- The spatially uniform solutions, Eqs. (20) and (22), and
turbations in the velocity fields of both species grow equally their linear stability analysis (Fig. 4) suggest five distinct
fast (δα1 /(δα1 + δβ1 ) → 0.5). For τ → 0, however, pertur- phases. Especially in the regimes where spatially uniform
bations δα1 only grow in the unstable region in the vicinity of ordered states are unstable against spatially nonuniform per-
σtA , while perturbations δβ1 only grow in the unstable region turbations [colored regimes in Figs. 4(b)–4(d)], we expect the
at σtB . These results are discussed in more depth in Appendix formation of spatiotemporal patterns. To study the nonlin-
B 1. Components perpendicular to the macroscopic order are ear dynamics of the hydrodynamic equations, Eqs. (14) and
stable to perturbations. (18), beyond linear stability, we solve these equations numer-
In summary, the main insight gained in this section is that ically. Specifically, we use a finite difference scheme for a
there are regions in parameter space where spatially uniform two-dimensional grid with periodic boundary conditions. We
polar ordered states are linearly unstable against spatially employ an explicit iterative process, the fourth-order Runge-
nonuniform perturbations. These regions are located in the im- Kutta method, to determine the time evolution; for details on
mediate vicinity of the threshold values for the noise strength, the implementation please see Appendix C.
σtA and σtB , that mark the instability of a disordered state to
states with different degrees of spatially uniform polar order. 1. Phase diagram and phase transitions
These threshold values of the noise strength are functions These numerical solutions of the hydrodynamic equations
of the relative fitness, λ. The game strength, τ , affects the confirm the existence of five distinct phases (Fig. 5). We
relative growth rates of perturbations of the two densities, a find three spatially uniform phases, a disordered phase, an
and b, and velocities parallel to macroscopic order, α1 and intermediate phase where species A shows polar order while
β1 : For high values of τ , perturbations in the fields of both species B does not, and a phase where both species exhibit
species grow equally fast; for decreasing τ , growth rates dif- polar order. The parameter regimes determined numerically
fer between the species until for τ → 0 instabilities in the are in accordance with our analytical calculations (described
fields of A and B occur independently. In the next section we in Secs. III B and III C), where we found uniform solutions to
044408-8SNOWDRIFT GAME INDUCES PATTERN FORMATION IN … PHYSICAL REVIEW E 104, 044408 (2021)
be linearly stable against nonuniform perturbations [the three
white regions in Figs. 4(b)–4(d)].
In the parameter regime where the spatially uniform so-
lutions are linearly unstable against spatial perturbations, we
observe the formation of spatial patterns. These patterns are
characterized by density-segregated, polar-ordered bands in
both species that concurrently move through the system; see
Movie 1 in the Supplemental Material [53] for an illustration
(an explanation and the parameter set for the movie can be
found in Appendix E). We will refer to these traveling bands
as polar waves. The wave vector of the traveling patterns
always points in the same direction as the macroscopic polar
order, as suggested by linear stability analysis (see Sec. III C).
When viewed from a comoving frame, the patterns are uni-
form in the direction perpendicular to the wave vector. Similar FIG. 6. Typical wave profiles for the density fields (solid lines), a
density-segregated traveling wave states are known from one- and b, and the velocity fields (dashed lines), |α1 | and |β1 |, in the trav-
species SPP systems [42,54,55]. Interestingly, the domains eling wave phases TWA (a) and TWB (b). Control parameters are set
where traveling wave patterns exist, extends beyond the pa- to relative fitness λ = 0.4, game strength τ = 0.2, and noise strength
rameter regime where the spatially uniform polar-ordered σ = 0.67 in (a), and σ = 0.47 in (b). The insets show how the density
states are unstable to spatial perturbations of finite wave- and velocity fields are related to each other: At every point in space,
length. In the blue shaded regime in Fig. 5 the system exhibits the velocity fields α1 and β1 (dashed lines in the insets) are approx-
bistabilty and subcritcal behavior as discussed in detail below; imately equal to the values |α1 (r, t )| ≈ |α1(0) (r, t )| = μA1 (a, b)/ξ (a)
see Sec. III D 2. We have also investigated phase diagrams for and |β1 (r, t )| ≈ |β1(0) (r, t )| = μB1 (a, b)/ξ (b) corresponding to spa-
different values of τ and found the same topology. tially uniform solutions [see Eqs. (22)] for the respective local
We will see in the next section that the mechanism under- densities a(r, t ) and b(r, t ) (green solid lines in the insets). The
lying pattern formation varies according to the region in the waves in species B in the TWA phase seem to be an exception to
parameter space. A distinction is therefore made between two this observation (inset in the bottom left figure).
different phases with spatial patterns. We refer to the traveling
polar wave state emerging in the unstable region in parameter
space at σtA as traveling polar wave phase A (TWA) and quires that the densities are large enough such that locally
the one in the unstable region at σtB as traveling polar wave one is above the threshold for polar order, i.e., both μA1 (a, b)
phase B (TWB). For both of these phases, examples of the and μB1 (a, b) are positive (see Sec. III B). The only exception
wave profiles for the density fields, a and b, and the velocity are patterns of the B species in the TWA phase in a regime
fields, |α1 | and |β1 |, are shown in Fig. 6; the superscript for where both λ and τ are small enough such that the density
the velocity fields indicates that we consider the component b falls below the threshold value for polar order, μB1 (a, b).
parallel to the direction of macroscopic order, α1(0) and β1(0) , Nevertheless we see order emerge in the high-density wave
and wave vector, q (as mentioned in Sec. III C, the stability peak [see lower inset of Fig. 6(a)]. In summary, the pattern can
analysis showed that instabilities are largest for q α1(0) , β1(0) , segregate the system in regions of high density with macro-
meaning that patterns form along the direction of macroscopic scopic order (|α1 | > 0 or |β1 | > 0) and low density regions
order). Our numerical solutions show that the polar waves for that are disordered (|α1 | = 0 or |β1 | = 0).
species A and B are strongly correlated spatially, meaning that
they follow each other through the system. These findings
are consistent with our results from linear stability analysis, 2. Bistability and subcriticality
where we found that instabilities always occur simultaneously Previous studies on SPP systems indicate that patterns are
for interacting species A and B. stable even in regimes where uniform solutions are linearly
In the three spatially uniform phases, all fields show values stable [47,49]. To identify the full regime in which pattern
that are close to their uniform fixed point values given by solutions exist, we use the following approach (“hysteresis
Eqs. (20) and (22). Although in the TWA and TWB phase, the loop”): We first compute the stationary solution for fixed
fields exhibit spatial patterns and deviate significantly from parameters (λ, τ ) and σ > σtA where there is no macroscopic
their uniform fixed point values at different points in space, order, neither in A nor in B. Then, we reduce the noise strength
the spatial averages ā, b̄, |ᾱ1 | and |β̄1 | are approximately also σ quasistatically in small steps, thereby sweeping through the
given by the corresponding uniform fixed point values. different phases in parameter space. By “quasistatic” we mean
Moreover, we find that at every point in space, the that the system is given enough time to equilibrate between
velocity fields, |α1 (r, t )| and |β1 (r, t )|, are well approxi- successive adjustments in σ . Afterward, we increase the noise
mated by the local values of the spatially uniform solu- strength again quasistatically back to its initial value. During
tions, Eq. (22). For local densities a(r, t ) and √ b(r, t ) with this σ sweep, we monitor the spatial average of the velocity
μA/B
1 > 0, these are
√ given by |α1
(0)
(r, t )| = μA1 (a, b)/ξ (a) fields, |ᾱ1 | and |β̄1 |. To identify the presence of spatial pat-
(0)
and |β1 (r, t )| = μ1 (a, b)/ξ (b), else we have |α1 |, |β1 | = 0.
B terns, we use parameters that we call variation parameters ηA
Hence, nonzero values for the velocity fields α1 and β1 re- and ηB , which measure spatial variations in the densities a and
044408-9MAYER, OBERMÜLLER, DENK, AND FREY PHYSICAL REVIEW E 104, 044408 (2021)
are stable solutions, i.e., the system shows a bistable dynamics
and subcritical behavior.
For a neutral game (λ = 1), species A and B show identical
behavior [Fig. 7(a)]: Quasistatically decreasing (gray arrows)
and then increasing (black arrows) the noise strength σ one
observes a hysteresis loop. Both, the variation parameters, ηA
and ηB , and the average velocities, |ᾱ1 | and |β̄1 |, show dis-
continuous changes (up and down arrows) at the boundaries
of the bistable regime at high noise values (right blue-shaded
region). Hence, in the bistable regime, the disordered phase
and the traveling wave phase (TWA) are possible metastable
states. In contrast, in the bistable regime at low noise strength
(left blue-shaded region) only the spatial variation parameter
exhibits a hysteresis loop while the average velocities change
continuously. Moreover, the spatial averages of the numeri-
cally determined values for the velocities closely follow the
results calculated analytically for a spatially uniform system
[dashed lines in Fig. 7(a)]; compare Eq. (22). This has two
implications. First, a spatially uniform polar-ordered state and
a traveling wave state are metastable solutions (in the left
bistable regime). Second, the spatially uniform and spatially
nonuniform state have the same average velocities. We there-
fore call this transition quasicontinuous.
For small enough values of the relative fitness λ [λ = 0.2 in
Fig. 7(b)], one observes all of the five different phases (com-
FIG. 7. Bifurcations of the spatial average of the velocity fields, pare also Fig. 5) with bistable regions as well as hysteresis
|ᾱ1 | and |β̄1 |, and the spatial variation parameters, ηA and ηB , as a loops at each transition. In addition to the disordered phase
function of the noise strength σ for a fixed game strength τ = 0.2 at high noise strength and the spatially uniform polar-ordered
and for different values of the relative fitness λ: (a) λ = 1, (b) λ = 0.2, phase at low noise strength, in which both species show polar
and (c) λ = 0.5. Blue and red lines consist of sets of dots that each order (|α1 |, |β1 | > 0), there is now an intermediate phase, in
indicate a stable state calculated numerically. The dashed lines in- which only species A shows polar order, but species B does
dicate the analytical results for the spatially uniform amplitudes of
not (|α1 | > 0, |β1 | = 0). Similar as for a neutral game, the
the velocity fields, Eq. (22); compare with Fig. 3. Regions shaded
spatial mean of both velocities is fairly well approximated
in blue and yellow indicate parameter regimes, in which the system
by the analytical results we obtained for a spatially uniform
is bistable or the spatially uniform states are linearly unstable with
respect to spatially heterogeneous perturbations, respectively.
system [dashed lines in Fig. 7(b)], even in the regime where
the system actually shows phase separation into polar bands
b. They are defined as of high density and disordered spatial regions of low density.
For large enough values of the relative fitness λ [λ = 0.5
|a| |b| in Fig. 7(c)], the two traveling polar wave phases, TWA and
ηA := , and ηB := , (25) TWB, merge; compare also Fig. 5. The phase behavior is
x x
similar as in the neutral case with the difference, that the
where |a|
x
is the absolute value of the numerical spatial traveling polar waves in species A and B differ in shape and
derivative of the density field a and the bar indicates the degree of polarization [see the different values of ηA , ηB and
average over all grid points (see Appendix C). For ηA = 0 |α1 |, |β1 | in Fig. 7(c)].
(ηB = 0), species A (B) is in a spatially uniform state, while Last, in a small parameter regime (close vicinity around
a nonzero value ηA > 0 (ηB > 0) indicates that the system the blue star in Fig. 5), the transition between the uniform
exhibits some kind of spatial patterns in A (B). By combining partially polar-ordered phase and the TWA phase becomes
the information about average velocities and spatial variations supercritical, i.e., we do not find jumps in the variation param-
we can infer the phase of the system, as we discuss next. eters and average velocities, but a continuous change between
Figure 7 shows the spatial variation parameters, ηA and zero and nonzero values. Moreover, in this regime the dy-
ηB , and spatial averages of the velocity fields, |ᾱ1 | and |β̄1 |, namics become extremely slow, so that the system takes a
obtained from σ sweeps for different values of the relative long time to reach its spatially nonuniform, stationary state,
fitness λ and a fixed game strength τ = 0.2. A general observa- indicating critical slowing down.
tion from our numerical analysis is that phases with traveling
polar waves exist in parameter regimes wider than the do-
mains predicted by the linear stability analysis of spatially 3. Game-induced pattern formation
uniform states; compare the yellow- and blue-shaded regimes In the last section we found that the two traveling wave
in Fig. 7. While in the yellow-shaded parameter regime only phases, TWA and TWB, exhibit coupled traveling polar wave
the traveling waves are stable solutions, in the blue-shaded pa- patterns. In a next step, we now investigate the relationship
rameter regime both these waves and spatially uniform states between the interaction mediated by the snowdrift game and
044408-10You can also read

















































