Potential of Ultrasound to Control Sesamia cretica (Lepidoptera: Noctuidae)
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Environmental Entomology, 50(6), 2021, 1393–1399
https://doi.org/10.1093/ee/nvab103
Advance Access Publication Date: 1 October 2021
Research
Pest Management
Potential of Ultrasound to Control Sesamia cretica
(Lepidoptera: Noctuidae)
Hamzeh Agah-Manesh,1 Ali Rajabpour,1,3, Fatemeh Yarahmadi,1, and Afrooz Farsi2
Downloaded from https://academic.oup.com/ee/article/50/6/1393/6378662 by guest on 25 December 2021
1
Department of Plant Protection, Faculty of Agriculture, Agricultural Sciences and Natural Resources University of Khuzestan,
Mollasani, Ahvaz, 6341773637, Iran, 2Division of Plant Protraction, Khuzestan Sugarcane & By-products Research & Training
Institute, Ahvaz, 6134814617, Iran, and 3Corresponding author, e-mail: rajabpour@asnrukh.ac.ir
Subject Editor: Punya Nachappa
Received 15 May 2021; Editorial decision 22 August 2021
Abstract
The pink stalk borer, Sesamia cretica Led. (Lepidoptera: Noctuidae), is one of the most important sugarcane pests
in many regions of the world, causing severe damage to sugarcane every year. This insect has a specialized form
of the auditory organ called the tympanal organ, and ultrasound can be employed as a potential tactic employed in
physical control strategy against the pest. The present study evaluates the efficacy of ultrasound in controlling the
pest in laboratory conditions. For this purpose, the repellent properties of various ultrasonic frequencies ranging
from 21 to 100 kHz with 0.5 kHz intervals and wave shapes, including Sin(x), Cos(x) square, and sawtooth, were
studied in choice experiments on the moths.The repellent effects of ultrasonic waves at frequencies 39.5 and 37.5 kHz
were more significant than other frequencies in male and female moths, respectively. Furthermore, there was no
significant difference between the repellent properties of different wave shapes. In non-choice experiments, the
effects of the most repellent ultrasonic treatment, at frequency 37.5 kHz, on biological characteristics of various life
stages and distribution patterns of the moths were investigated. The results showed that the ultrasonic treatment
causes substantial reductions in many biological parameters of the immature life stages of pests, including
longevity, weight, survival rate, and fecundity. Moreover, the pattern indicated that the moths tended to escape
from the ultrasound. The findings of this study can be employed for manufacturing the ultrasonic repeller to be
used in sugarcane fields.
Key words: stalk borer, sugarcane, frequency, repellency, biological parameters
Sugarcane (hybrids of Sacharrum officinarum L.) is an important quality and quantity of sugarcane juice are significantly decreased by
industrial crop cultivated in many tropical and subtropical regions larval infestations. The pest has four generations in Khuzestan prov-
of the world (Hunsigi 2012), providing approximately 70% of sugar ince. The moths are nocturnal and females oviposit behind green
produced worldwide. Moreover, this crop has high potential for pro- sugarcane leaf sheaths (Khanjani 2009).
ducing other products, including bagasse, straw, molasses, filter cake, Because the larvae are active inside sugarcane stems, chemical con-
and ethanol (Paturau 1989). Khuzestan province (Khuzestan means trol is not effective in controlling the pest (Halabian et al. 2013). The
the land of sugar) is the main region of sugarcane cultivation in Iran inoculative release of Telenomus busseolae Gahan (Hymenoptera:
(Hamdi 2016). Scelionidae) is the main strategy for controlling the pest in sugarcane
The corn stem borer, Sesamia cretica Led. (Lepidoptera: fields of Khuzestan province (Khanjani 2009, Nikpay and Goebel
Noctuidae), is one of the most destructive sugarcane pests in 2016). However, it is demonstrated that the parasitic wasp cannot
Khuzestan province, also attacking other cereal plants including sufficiently suppress the pest population in many sugarcane cultivars
maize and rice (Khanjani 2009, Esfandiari 2015). Feeding by the (Halabian et al. 2013).
larvae causes dead heart in young plants. Boring by the larvae in ma- Noctuid moths can detect the ultrasonic pulses of insectiv-
ture plant stems leads to their weakness or plant death. Totally, the orous bats using tympanal organ (Lapshin and Federova 1996), a
© The Author(s) 2021. Published by Oxford University Press on behalf of Entomological Society of America. All rights reserved.
For permissions, please e-mail: journals.permissions@oup.com.
1393
1394 Environmental Entomology, 2021, Vol. 50, No. 6
specialized hearing organ evolved in seven different insect orders for Materials and Methods
hearing their predators, prey, and potential mates and rivals (Hoy
Insect Rearing
and Robert 1996). They show various escape behaviors for avoiding
predators (Zha et al. 2013). Moreover, some of them emit echoloca- The larvae of S. cretica were collected from infested sugarcane shoots
tion signals to determine their spatial directions during night flights in Farabi Agro-industry in Khuzestan province, Iran (30°56′40.6″N,
(Lapshin and Federova 1996) and mating communications (Lapshin 48°33′29.1″E). After the larva identification using Esfandiari et al.
and Vorontsov 2003). (2011) criteria, larvae were reared on the sugarcane shoots (15 cm in
Using ultrasonic signals for lepidopteran pest control (Huang length and 2.2–2.8 cm in diameter), and eight shoots were placed in
et al. 2003, Huang and Subramanyam 2004, Zha et al. 2013, Salehi each rearing cylinder (18 cm in height and 10 cm in diameter). The
et al. 2016) as well as non-lepidopteran insect pests (Kalimuthu et al. rearing was done at 28 ± 1°C, 65 ± 5% RH, and a photoperiod of
2020, Ozkurt 2021) has been suggested by many researchers. There 16:8 (L:D) h in a growth chamber. After the emergence of the moths
are numerous sonic and ultrasonic devices such as insect pest repellers (F2), the species were identified according to the key by Tams and
and pest ultrasonic suppressers claimed to be effective in controlling Bowden (1953). In each rearing cylinder containing eight sugarcane
a wide range of insect pests. Certainly, this claim cannot be scientif- shoots, eight moths (four females and four males) were introduced
Downloaded from https://academic.oup.com/ee/article/50/6/1393/6378662 by guest on 25 December 2021
ically supported. Insect species show different physiological sensitiv- for oviposition. The new shoots were replaced every day, and the
ities to sound or ultrasound waves (Hoy et al. 1989). Therefore, all previous shoots (containing egg masses) were translocated to new
of them cannot respond to a specific ultrasound signal. The physical rearing cylinders. When the eggs hatched, the first instar larvae were
features of sonic/ultrasonic signals such as the frequency and shape reared on small pieces of sugarcane stalks (2 cm in length and 2.2–
of sound waves can influence the efficacy of the devices significantly 2.8 cm in diameter) in plastic Petri dishes (40 pieces per dish). The
(Hoy and Robert 1996, Salehi et al. 2016). Frequency (measured in third instar larvae were translocated to sugarcane shoots (one larva
Hz) is the number of times per second that a sound pressure wave per shoot) by making a small hole at the top of the shoots.
repeats itself. High frequencies produce more oscillations. Humans
with normal hearing can hear sounds between 20 and 20,000 Hz. Generating Various Ultrasonic Waves
Sound with higher frequencies, 20–100 kHz, is considered as ultra- The signal generator device (SGD) and its intermediate software,
sound. The sound wave shape describes a depiction of the pattern of Function Generator version 1.1 (manufactured by Agricultural
sound pressure variation (or amplitude) in the time domain which Sciences and Natural Resources University of Khuzestan, Mollasani,
its main types are sine, triangle, square, and sawtooth (Kruth and Ahvaz, Iran and Ahvaz Robotic Company, Ahvaz, Iran) which was
Stobart 2007). Thus, it is critical to determine the effective features described by Salehi et al. (2016), were used for generating various
of waves emitted from the devices for practical use in integrated pest ultrasonic frequencies (21–100 kHz) and wave shapes at intensity
management (IPM) programs. 50 W/m2.
Thus far no effort has been made to investigate the behavioral
and biological responses of S. cretica and other noctuid species to Repellency
various ultrasonic features. Therefore, the present study aims to The experiment was performed according to the method adopted by
determine the effects of various frequencies and shapes of ultra- Salehi et al. (2016) in which ultrasound and control glass chambers
sonic waves on the repellency and biological characteristics of (0.2 × 0.2 × 0.2 m) were connected using a glass connective tube
the pest. (40 × 5 × 5 cm). The ultrasound emitter was located on the lateral
Fig. 1. The designed repellency test system (A: ultrasonic chamber that its all sides surrounded by soundproofing layers; B: control chamber; C: connecting tube;
D: ultrasonic waves; G: emitter of SGD; E: closure for moth entrance; F: moths; H: SGD; I: operation of intermediate software using laptop computer).
Environmental Entomology, 2021, Vol. 50, No. 6 1395
side of the ultrasound chamber (Fig. 1). Five 1-d-old moths from 2). Similar to the method mentioned in the insect rearing section,
each sex were introduced through a closure in the center of the con- the third, the fourth, and the fifth instar larvae and pupae (nine in-
nective tube. The ultrasonic frequencies ranged from 21 to 100 kHz, sects per RGB) were separately reared on the sugarcane shoots in the
with 0.5 kHz intervals. For each frequency, four different shapes, rearing cylinder as a replication. One cylinder was located in each
i.e., Sin(x), Cos(x), and square and sawtooth wave shapes, were quadrat of the RGB and was exposed to ultrasound. Longevity, sur-
separately generated using the SGD and according to the functions vival rate, and weight of the larvae and the pupae were recorded at
mentioned by Salehi et al. (2016). Each wave shape at a determined the end of each life stage. The experiments were replicated five times
frequency was considered as a treatment which has 10 replications. and, totally, 90 insects of each life stage were used in the experiments.
After each replication, the moths were removed from the chambers After the emergence of the adults, four pairs of moths (four males
and five new moths were introduced. The experimental time for each and four females) treated with ultrasound during their immature life
treatment was 10 min. The number of moths in each chamber, i.e., stages were reared in the rearing cylinder using the methods men-
ultrasound and control, was recorded after the period. The repelled tioned in the insect rearing section. The number of oviposited eggs
moths (RMs) were estimated using the following formula (Salehi and the hatching rates were recorded in daily sampling during 5
et al. 2016): d. Moreover, the sex ratio and survival rates of the moths were esti-
Downloaded from https://academic.oup.com/ee/article/50/6/1393/6378662 by guest on 25 December 2021
mated. The experiment has five replications (totally 40 moths were
(number of released moths/2)
RMs = used in the experiments).
number of moths in ultrasound chamber
In control, the experiments were conducted using the same
method without exposure to ultrasound. Each experiment was rep-
Immature Life Stages licated five times.
For this experiment, when the larvae were collected from the sugar-
cane field, they were reared on sugarcane shoots exposed to the ultra- Effects of Ultrasonic Treatment on Distribution
sonic treatment. The frequency and wave shape of the ultrasound Patterns of S. cretica Moths
treatment were 37.5 kHz and Cos(x), respectively. The frequency and
In non-choice trials, the distribution patterns of the moths under
wave shape were chosen according to the results of the repellency
the most repellent ultrasonic treatment, i.e., 37.5 kHz and Cos(x),
experiments showing the highest repellent effect to the moths. For
and those of control group were evaluated according to the method
this purpose, the infested shoots containing the larvae were placed
employed by Salehi et al. (2016). Twenty-eight pairs of moths, which
in rearing glass boxes (RGBs) (0.6 × 0.6 × 0.6 m). The ultrasound
consisted of 2-d-old males and females, were located in ultrasonic
emitter of the SGD was located at the top of the RGBs. When the
and control chambers. The number of the moths was recorded in
moths (F1) emerged, the females and the males (20 pairs per box)
each quadrat after 12 h.
were mated in another RGB under the ultrasonic treatment. Twenty-
five sugarcane shoots (18 cm in height and 10 cm in diameter) were
located in the RGB in which the honey solution was provided to feed Data Analyses
the moths which were removed after 24 h. After the emergence of For choice test trials, the factorial analysis (80 frequencies × 4
the first instar larvae, as mentioned in the previous section, the larvae wave shapes × 2 sexes) based on a completely randomized design
(the first and the second instar larvae) were reared on small pieces was carried out using the GLM procedure. For mean compari-
of sugarcane stalks in plastic Petri dishes (100 first instar larvae per sons, the least significant difference test was used as a post hoc
dish) under the ultrasonic treatment. To this end, the floor of each test of ANOVA.
RGB was divided into nine smaller quadrats (16 × 16 × 16 cm), and To compare biological parameters (survival rates and weights)
one dish was placed as a replicate in the center of each quadrat (Fig. in the ultrasonic treatment and control, the independent t-test was
Fig. 2. The designed system to evaluate effect of ultrasound on biological characteristics of S. cretica (A: ultrasonic chamber [with the ultrasonic exposure]; B:
control chamber [without the ultrasonic exposure]; C: experimental quadrate; D: sugarcane shoot; F: emitter of SGD; G: ventilation opening [covered with fine
gauze]; H: SGD; I: operation of intermediate software using laptop computer).1396 Environmental Entomology, 2021, Vol. 50, No. 6
done. All statistical analyses were performed using SAS 9.2 (SAS females. The results showed that the oviposition period in ultra-
Institute, Inc., Cary, NC). sound-treated moths (3 d) was shorter than that in control (5 d).
The distribution patterns of the moths in both treatments were Moreover, the total number of oviposited eggs in ultrasound-treated
determined according to the method described by Salehi et al. (2016) moths (45.4 eggs) was notably lower than that in control (76 eggs)
and using GS+ software. In this method, x and y coordinates repre- (Table 6).
sent the position of the insect and z represents the total number of The ultrasonic treatment did not have any significant effects on
moths recorded in each quadrat at a 12-h interval. By interpolating the hatching rates of insect eggs.
the values of the z coordinate using the linear Kriging interpolation
algorithm with a zero nugget, the software produces a grid of values. Distribution Pattern
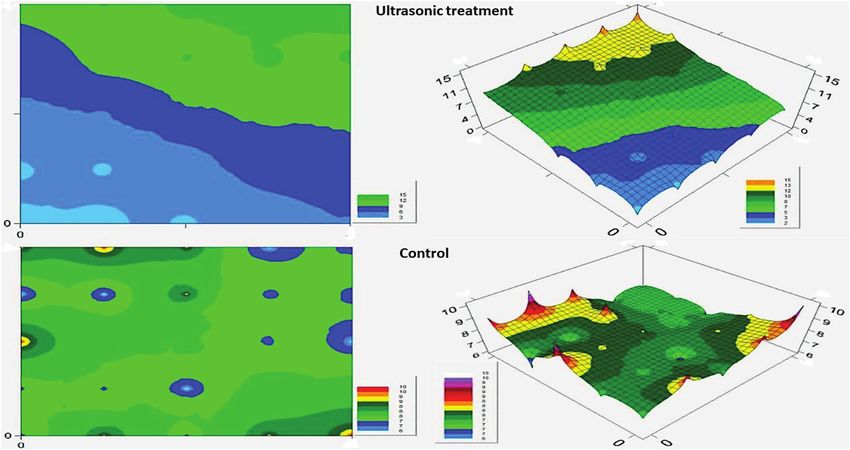
Figure 4 shows the distribution patterns of S. cretica moths under
ultrasonic treatment and control. The patterns indicate that the
Results
moths tended to escape from the ultrasonic emitter, while no
Repellency changes were observed in the distribution patterns of the moths in
The frequency of emitted ultrasound significantly influenced its re- control.
Downloaded from https://academic.oup.com/ee/article/50/6/1393/6378662 by guest on 25 December 2021
pellency to S. cretica moths (df = 77; F = 1.61; P = 0.0007). Moreover,
there was a strong interaction between wave frequency and moth sex
(df = 7; F = 1.89; P = 0.0004) (Table 1). However, wave shape, sex, Discussion
and their interactions had no substantial effects on the repellency
Our findings reveal that ultrasound, at a specific frequency, has
(Table 1).
significant repellent effects on S. cretica moths. Noctuid moths
Figure 3 shows the indices of repelled male and female moths.
can hear a broad range of acoustic signals, particularly those
The repellent effects of ultrasonic waves on male and female moths
whose frequency ranges from 20 to 50 kHz. The behavioral re-
at frequencies 39.5 and 37.5 kHz, respectively, were significantly
sponses of some noctuid moths, including Heliothis spp. (Agee
greater than other frequencies.
1969), Amphipyra perflua F. (Lapshin and Federova 1996),
Enargia paleacea Esp. (Lapshin and Vorontsov 2003), and
Biological Characteristics Spodoptera frugiperda Smith (Cinel and Taylor 2019) to ultra-
Table 2 shows different immature life stage longevities. Among the sound were previously reported.
life stages, the ultrasound significantly reduced the longevity of the Noctuid moths can detect the ultrasonic impulse differently. It
second instar larvae (6.5% lower than control) but did not have is demonstrated that the predation of various noctuid moths by
any marked effects on the longevity of other larval instars as well Hawaiian hoary bat, Lasiurus cinereus semotus Hall and Jones, is
as pupae. related to their different ability to detect the bat echolocation ultra-
In the ultrasonic treatment, there were considerable reductions sonic signals (Fullard 2001).
(6.2–8.1%) in all immature life stage weights, except for female The physical characteristics of ultrasonic signals, including fre-
pupae, in comparison with control (Table 3). quency, wave shape, and initial intensity, can significantly affect the
For immature life stages, the ultrasonic treatment notably de- lepidopteran behavioral responses to an emitted sound (Römer and
creased the survival rate of the third instar larvae (36.5%) (Table 4). Tautz 1992, Zha et al. 2009). Among the abovementioned charac-
However, the ultrasound did not affect the survival rates of other im- teristics, frequency is an important parameter providing the insects
mature life stages noticeably. The survival rates of male and female with special information. The sensitivity of the auditory system of
adults were significantly lowered in the ultrasonic treatment by 23.6 moths is usually maximized at a specific frequency range (Hoy et al.
and 14.2%, respectively (Table 4). 1989). It is demonstrated that the range reflects the frequency range
Furthermore, the sex ratio of the moths did not change radically of the echolocation impulses of sympatric bats (Zha et al. 2009).
in the ultrasound treatment (Table 5). The maximum sensitivity frequency ranges of the moth ears are dif-
Tables 6 and 7 present the number of oviposited eggs and the ferent between various families and species of moths due to their dif-
hatching rates in ultrasonic and control treatments, respectively. ferent types of auditory sensory cells (Göpfert and Wasserthal 1999).
Ovipositions in treated moths were significantly lower than those in No previous studies have focused on determining the responses of
control. The ultrasonic treatment caused early egg-laying. In ultra- Sesamia spp. to various frequencies of ultrasound. Therefore, our
sound-treated females, the maximum number of oviposited eggs was results cannot be compared with those of other studies. Our results
observed on the first day (20.8 eggs), being significantly higher than are in agreement with the findings reported by Zha et al. (2009)
that in control (0.8 eggs). However, the total egg-laying of untreated stating that the optimal frequencies of eared moths, e.g., the family
moths was substantially more than that of ultrasound-treated Noctuidae, range from 20 to 50 kHz.
The most repellent frequency of male and female moths is 39.5
Table 1. General linear model (GLM) parameters for main factors and 37.5 kHz, respectively. A sex-based auditory dimorphism in some
and their interaction for RMs of S. cretica moths, including Lymantria dispar L. (Lepidoptra: Lymantridae),
was distinguished. The maximum sensitivities of the auditory system
Source df F P-value
of male and female moths were observed in the frequency ranges of
Frequency 77 1.61 0.0007 30–50 and 10–20 kHz, respectively. The dimorphism may be related
Wave shape 3 0.51 0.6764 to the exposure of only male species to predatory bat attacks. There
Sex 1 0.02 0.8876 are some differences in the mechanics of the tympanal organ and its
Wave shape × Frequency 231 0.54 1.0000 sensory cells between male and female insects, causing different fre-
Wave shape × Sex 3 1.18 0.3146 quency sensitivities (Sueur et al. 2008).
Frequency × Sex 77 1.89 0.0004
The wave shape of ultrasound did not have any notable effects on
Wave shape × Frequency × Sex 231 0.44 1.0000
the repellency behavior of S. cretica moths. However, it significantlyEnvironmental Entomology, 2021, Vol. 50, No. 6 1397
Male
2
A
1.5
Repelled moths
1
DCB
AB
DCB
DCB
DCB
DCB
DC
DC
CB
DCB
DCB
DC
DCB
DCB
DCB
DCB
0.5
DC
DC
DCB
DCB
DCB
DC
DCB
DCB
DCB
DCB
DC
DCB
DCB
DC
DCB
DCB
DC
DCB
DC
DCB
D
DCB
DCB
DC
DC
DC
DC
DCB
47.5 DCB
DCB
DC
56.5 DCB
DCB
DCB
55.5 DCB
DC
DC
DCB
DCB
DCB
DCB
48.5 DCB
50.5 DCB
DC
DCB
DCB
DC
DCB
DC
DC
DCB
DCB
DCB
46.5 DCB
DCB
DC
DCB
DCB
DCB
53.5 DCB
51.5 DC
0
20.5
21.5
22.5
23.5
24.5
25.5
26.5
27.5
28.5
29.5
30.5
31.5
32.5
33.5
34.5
35.5
36.5
37.5
38.5
39.5
40.5
41.5
42.5
43.5
44.5
45.5
49.5
52.5
54.5
57.5
58.5
-0.5
Frequency
Female
1.5
Downloaded from https://academic.oup.com/ee/article/50/6/1393/6378662 by guest on 25 December 2021
A
CADBE
AB
CAB
FCGADBE
FCGADBE
CADBE
CADB
FCADBE
FCGADBE
FCGADBE
FCGADBE
1
Repelled moths
FCGDBE
FCGDBE
FCGDBE
FCGDBE
FCGDBE
FCGDBE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDE
FCGDBE
FCGDE
FCGDE
FCGDE
FCGDE
0.5 FCGDE
FCGDE
FCGDE
29.5 FCGDE
FCGDE
46.5 FCGDE
45.5 FGDE
FGDE
55.5 FCGDE
FGDE
FGDE
FGDE
FGDE
32.5 FGDE
48.5 FGDE
56.5 FGDE
FGDE
33.5 FGDE
FGDE
FGE
58.5 FGE
52.5 FGE
31.5FGE
FG
FG
0
G
22.5G
FGE
20.5
21.5
23.5
24.5
25.5
26.5
27.5
28.5
30.5
34.5
35.5
36.5
37.5
38.5
39.5
40.5
41.5
42.5
43.5
44.5
47.5
49.5
50.5
51.5
53.5
54.5
57.5
FG
-0.5
Frequuency
Fig. 3. Mean of RM indices of S. cretica at different ultrasonic frequencies (20.5–60 kHz).
Table 2. Mean ± SE S. cretica longevity (day) exposed to ultra- Table 4. Mean ± SE S. cretica survival rate (%) exposed to ultra-
sound (37.5 kHz) sound (37.5 kHz)
Life stage Mean ± SE t(df = 8) P-value Life stage Mean ± SE t(df = 8) P-value
Ultrasonic treatment Control Ultrasonic treatment Control
Larvae first 3.80 ± 0.10 4.04 ± 0.08 −1.81 0.108 Larvae first 100 100 — —
Larvae second 9.30 ± 0.20 9.95 ± 0.03 −3.23 0.012 Larvae second 58.00 ± 5.70 63.00 ± 4.47 −1.54 0.161
Larvae third 10.80 ± 0.37 11.20 ± 0.37 −0.75 0.471 Larvae third 57.25 ± 8.83 82.39 ± 9.35 −4.36 0.002
Larvae fourth 10.80 ± 0.37 11.60 ± 0.24 −1.79 0.111 Larvae fourth 84.64 ± 1.90 88.52 ± 7.19 −1.16 0.276
Larvae fifth 9.20 ± 0.34 10.00 ± 0.31 −1.63 0.141 Larvae fifth 86.95 ± 12.68 88.06 ± 12.52 −0.139 0.89
Pupae 5.00 ± 0.31 5.4 ± 0.24 −1.00 0.347 Pupa (female) 100.00 ± 0.00 100.00 ± 0.00 — —
Pupae (male) 100.00 ± 0.00 100.00 ± 0.00 — —
Adult (female) 4.20 ± 0.50 5.5 ± 0.57 −3.27 0.017
Adult (male) 4.5 ± 0.57 5.25 ± 0.50 −1.96 0.097
Table 3. Mean ± SE S. cretica weight (gram) exposed to ultrasound
(37.5 kHz) Table 5. Female sex ratios (no. females/no. female + males) of
S. cretica in ultrasonic treatment (37.5 kHz) and control
Life stage Mean ± SE t(df = 8) P-value
Sex ratio ± SE t(df = 4) P-value
Ultrasonic treatment Control
Ultrasound 0.43 ± 0.31 −0.48 0.654
Larvae first 0.33 ± 0.01 0.37 ± 0.01 −4.00 0.004
Control 0.46 ± 0.11 −0.65 0.551
Larvae second 0.57 ± 0.01 0.63 ± 0.01 −6.621398 Environmental Entomology, 2021, Vol. 50, No. 6
organ, but immature life stages can hear environmental sounds by Acknowledgments
some types of sensilla (Surlykke et al. 2003, Taylor and Yack 2019).
The authors thank Parian and Khuzestan Sugarcane & By-products Research
Enough devices running the ultrasound in each sugarcane field & Training Institute for their technical assistances. Agricultural Sciences and
can be applied as a suitable push–pull strategy against the pest. Natural Resources University of Khuzestan (grant number 9728401) funded
In the strategy, the insect behavior has been manipulated using this study.
stimuli (the ultrasound in this case) to make the protected resource
unattractive or unsuitable to the pests (push) while luring them
toward an attractive source (pull) from where the pests are sub- References Cited
sequently removed. The strategy has many advantages including: Agee, H. R. 1969. Response of Heliothis spp. (Lepidoptera: Noctuidae) to
1) nontoxic, 2) usually compatible with other IPM strategies, and ultrasound when resting, feeding, courting, mating, or ovipositing. Ann.
3) selectivity. The strategy is a valuable tool for IPM programs re- Entomol. Soc. Am. 62: 1122–1128.
ducing pesticide input or enhancing the efficacy of other control- Cinel, S. D., and S. J. Taylor. 2019. Prolonged bat call exposure induces a
ling strategies by orchestrating a predictable distribution of pests broad transcriptional response in the male fall armyworm (Spodoptera
frugiperda; Lepidoptera: Noctuidae) brain. Front. Behav. Neurosci. 13: 36.
(Cook et al. 2007).
Downloaded from https://academic.oup.com/ee/article/50/6/1393/6378662 by guest on 25 December 2021
Cook, S. M., Z. R. Khan, and J. A. Pickett. 2007. The use of push-pull strat-
In conclusion, the findings of this study showed that the ultra-
egies in integrated pest management. Annu. Rev. Entomol. 52: 375–400.
sound frequencies of 39.5 and 37.5 kHz have the highest repel-
Esfandiari, M., M. S. Mossadegh, and P. Shishehbor. 2011. Sesamia botanephaga
lency to S. cretica moths and that wave shapes do not have any Tams & Bowden, 1953 auct. in Iran, read Sesamia nonagrioides (Lefebvre,
significant effects on this property. Moreover, the ultrasonic treat- 1827) (Lepidoptera: Noctuidae). Munis Entomol. Zool. 6: 400–403.
ment causes considerable reductions in some biological param- Esfandiari, M., N. Mehravar, H. Motamedi, H. R. Memari, and H. Rajaei. 2015.
eters of immature life stages, including longevity, weight, survival Phylogeography of Sesamia nonagrioides (Lepidoptera: Noctuidae) in Iran in-
rate, and fecundity in the pest. Therefore, ultrasound can be em- ferred from mitochondrial DNA. Acta Zool. Acad. Sci. Hung. 61: 341–359.
ployed in pest repellers; however, further field experiments are Fullard, J. H. 2001. Auditory sensitivity of Hawaiian moths (Lepidoptera:
required for evaluating the efficacy of such devices before making Noctuidae) and selective predation by the Hawaiian hoary bat
(Chiroptera: Lasiurus cinereus semotus). Proc. R. Soc. Lond. B: Biol. Sci.
any recommendations.
268: 1375–1380.
Gopfert, M. C., and L. T. Wasserthal. 1999. Auditory sensory cells in hawk-
Table 7. Mean ± SE of egg hatching rate (%) when the moths (male moths: identification, physiology and structure. J. Exp. Biol. 202: 1579–1587.
and female) of S. cretica exposed to ultrasound (37.5 kHz) Halabian, A. H., S. Cheraghi, and J. Pourreza. 2013. Performance evaluation
of biological control of sugarcane stem borers wasp (Telenomus busseo-
Day Mean ± SE t(df = 8) P-value lae). World Appl. Sci. J. 21: 1770–1775.
Hamdi, H. 2016. Sustainability in sugarcane production: opportunities and
Ultrasonic treatment Control
limitations in Southwest of Iran. Sugar Tech. 18: 642–646.
1 21.30 ± 1.5 25.00 ± 5.24 −0.27 0.792 Hoy, R. R., and D. Robert. 1996. Tympanal hearing in insects. Annu. Rev.
2 68.48 ± 15.16 78.99 ± 7.25 −1.39 0.200 Entomol. 41: 433–450.
3 78.73 ± 5.54 79.13 ± 6.76 −0.10 0.922 Hoy, R., T. Nolen, and P. Brodfuehrer. 1989. The neuroethology of acoustic
startle and escape in flying insects. J. Exp. Biol. 146: 287–306.
Fig. 4. Distribution patterns of S. cretica moths in the ultrasonic and control chambers. Spot of 0 and 0 indicated spatial coordinate of the ultrasound outlet.
Different densities of the moths per each quadrat of rearing glass chamber are presented with various colors.Environmental Entomology, 2021, Vol. 50, No. 6 1399
Huang, F., and B. Subramanyam. 2004. Behavioral and reproductive effects of Römer, H., and J. Tautz. 1992. Invertebrate auditory receptors. Comp. Asp.
ultrasound on the Indian meal moth, Plodia interpunctella. Entomol. Exp. Echanorecep. Sys. 10: 185–212.
Appl. 113: 157–164. Salehi, S. S., A. Rajabpour, A. Rasekh, and M. Farkhari. 2016. Repellency and
Huang, F., B. Subramanyam, and R. Taylor. 2003. Ultrasound affects sperm- some biological effects of different ultrasonic waves on Mediterranean
atophore transfer, larval numbers, and larval weight of Plodia interpunc- flour moth, Ephestia kuehniella (Zeller) (Lepidoptera: Pyralidae). J. Stored
tella (Hübner) (Lepidoptera: Pyralidae). J. Stored Prod. Res. 39: 413–422. Prod. Res. 69: 14–21.
Hunsigi, G. 2012 Production of sugarcane: theory and practice, vol. 21. Spangler, H. G. 1984. Responses of the greater wax moth, Galleria mellonella
Springer Science and Business Media, Berlin, Germany. L. (Lepidoptera: Pyralidae) to continuous high-frequency sound. J. Kansas
Kalimuthu, K., L. C. Tseng, K. Murugan, C. Panneerselvam, A. T. Aziz, Entomol. Soc. 57: 44–49.
G. Benelli, and J. S. Hwang. 2020. Ultrasonic technology applied against Sueur, J., J. F. Windmill, and D. Robert. 2008. Sexual dimorphism in audi-
mosquito larvae. Appl. Sci. 10: 3546. tory mechanics: tympanal vibrations of Cicada orni. J. Exp. Biol. 211:
Khanjani, M. 2009. Field crop pests in Iran. Bu-Ali Sina University Press, 2379–2387.
Hamadan, Iran. Surlykke, A., J. E. Yack, A. J. Spence, and I. Hasenfuss. 2003. Hearing in
Kruth, P., and H. Stobart. 2007. Sound. Cambridge University Press, London, hooktip moths (Drepanidae: Lepidoptera). J. Exp. Biol. 206: 2653–2663.
United Kingdom. Tams, W. H. T., and J. Bowden. 1953. A revision of the African species of
Downloaded from https://academic.oup.com/ee/article/50/6/1393/6378662 by guest on 25 December 2021
Lapshin, D. N., and M. V. Federova. 1996. Responses of the tympanic organs Sesamia Guenée and related genera (Agrotidae-Lepidoptera). Bull.
of cutworm moth (Amphipyra perfina: Noctuidae) to ultrasound impulses. Entomol. Res. 43: 645–678.
Sens. Sys. 10: 1–11. Taylor, C. J., and J. E. Yack. 2019. Hearing in caterpillars of the monarch
Lapshin, D. N., and D. D. Vorontsov. 2003. Sensitivity of the noctuid moth butterfly (Danaus plexippus). J. Exp. Biol. 222: 53–63.
Enargia paleceae Esp. (Lepidoptera, Noctuidae) to echolike stimuli. Dokl. Zha, Y. P., and C. L. Lei. 2012. Effects of ultrasound-stress on antioxi-
Biol. Sci. 390: 210–212. dant enzyme activities of Helicoverpa armigera (Hübner) (Lepidoptera:
Nikpay, A., and F. R. Goebel. 2016. Major sugarcane pests and their manage- Noctuidae). J. Agri. Urban Entomol. 28: 34–41.
ment in Iran. Proc. Inter. Soc. Sugarcane Technol. 29: 103–108. Zha, Y., Q. Chen, and C. Lei. 2009. Ultrasonic hearing in moths. Ann. Soc.
Ozkurt, H. 2021. Investigation of some ultrasonic sound frequencies effects on Entomol. Fran. 45: 145–156.
Culex pipiens sensu stricto (Diptera: Culicidae) larvae by using piezoelec- Zha, Y. P., J. Y. Chen, Z. B. Jin, C. B. Wang, and C. L. Lei. 2013. Effects
tric transducer. Inter. J. Trop. Insect Sci. doi:10.1007/s42690-021-00585-6. of ultrasound on the fecundity and development of the cotton bollworm,
Paturau, J. M. 1989. By-products of the cane sugar industry. An introduction to their Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) 1. J. Agri. Urban
industrial utilization. Elsevier Science Publishers BV, Amsterdam, Netherlands. Entomol. 29: 93–98.You can also read

















































