Desmosomes and Intermediate Filaments: Their Consequences for Tissue Mechanics - Cold Spring Harb ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
Desmosomes and Intermediate Filaments:
Their Consequences for Tissue Mechanics
Mechthild Hatzfeld,1 René Keil,1 and Thomas M. Magin2
1
Institute of Molecular Medicine, Division of Pathobiochemistry, Martin-Luther-University Halle-Wittenberg,
06114 Halle, Germany
2
Institute of Biology, Division of Cell and Developmental Biology and Saxonian Incubator for Clinical
Translation (SIKT), University of Leipzig, 04103 Leipzig, Germany
Correspondence: thomas.magin@uni-leipzig.de; mechthild.hatzfeld@medizin.uni-halle.de
Adherens junctions (AJs) and desmosomes connect the actin and keratin filament networks of
adjacent cells into a mechanical unit. Whereas AJs function in mechanosensing and in
transducing mechanical forces between the plasma membrane and the actomyosin cyto-
skeleton, desmosomes and intermediate filaments (IFs) provide mechanical stability required
to maintain tissue architecture and integrity when the tissues are exposed to mechanical
stress. Desmosomes are essential for stable intercellular cohesion, whereas keratins deter-
mine cell mechanics but are not involved in generating tension. Here, we summarize the
current knowledge of the role of IFs and desmosomes in tissue mechanics and discuss
whether the desmosome–keratin scaffold might be actively involved in mechanosensing
and in the conversion of chemical signals into mechanical strength.
he majority of tissues are constantly exposed epithelia that line organ and body surfaces to
T to external forces, such as mechanical load,
stretch, and shear stress, in addition to intrinsic
provide structural support and serve as barriers
against diverse external stressors such as me-
forces generated by contractile elements inside chanical force, pathogens, toxins, and dehydra-
tissues. Both extrinsic and intrinsic forces con- tion. Epithelia contain two types of intercellular
tribute to tissue morphogenesis, homeostasis, adhesion complexes: adherens junctions (AJs)
and regeneration and affect cell shape, prolifer- and desmosomes, connected to the actin and
ation, and migration (Evans et al. 2013). Sensing keratin cytoskeleton, respectively (Fletcher and
and transmitting forces depend to a large extent Mullins 2010).
on tight interactions between cell adhesion The detailed molecular mechanisms that
complexes and the cytoskeleton. Mechanosens- underlie mechanotransduction are complex
ing and mechanotransduction can be defined as and only partially understood. Recent data in-
cellular processes that convert mechanical cues dicate that mechanosensor proteins can under-
into intracellular signaling (Furuse et al. 2002; go force-induced conformational changes that,
Huveneers and de Rooij 2013; Janmey et al. in turn, induce changes in their activity or affin-
2013). These processes are exemplified by ity for binding partners (Yonemura et al. 2010;
Editors: Carien M. Niessen and Alpha S. Yap
Additional Perspectives on Cell –Cell Junctions available at www.cshperspectives.org
Copyright # 2017 Cold Spring Harbor Laboratory Press; all rights reserved
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157
1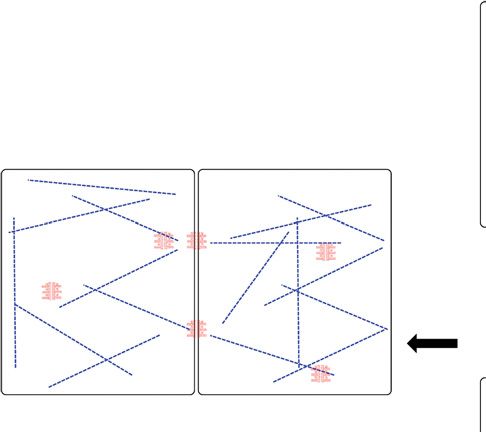
Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
M. Hatzfeld et al.
Huveneers and de Rooij 2013). This can finally referred to recent reviews (Meens et al. 2013;
lead to the activation of chemical signaling Patel and Green 2014).
cascades. As discussed in Yap (2017), AJs func-
tion as mechanosensors (Huveneers and de
Rooij 2013; Yao et al. 2014; Ladoux et al. 2015; FUNCTION OF DESMOSOMES IN
Muhamed et al. 2016), whereas a role in force CONFERRING MECHANICAL STABILITY
sensing has so far not been attributed to desmo-
Composition and Structure of Desmosomes
somes. At the same time, desmosome-mediated
intercellular adhesion is much stronger than AJ- Desmosomes are intercellular junctions essen-
mediated cohesion as shown by the epithelial tial for mediating strong intercellular cohesion
sheet assay: Whereas depletion of the desmoso- (Garrod 2010; Green et al. 2010; Kowalczyk and
mal plaque component plakophilin 1 (PKP1) in Green 2013). They are composed of three pro-
keratinocytes disrupts epithelial cohesion on tein families. The desmosomal cadherins, des-
application of mechanical stress, knockdown mogleins (DSGs), and desmocollins (DSCs),
of the corresponding components from AJs, are transmembrane proteins whose extracellular
p120, or p0071/PKP4, has no immediate effect domains form the adhesive interface of the des-
on intercellular cohesion (Fig. 1). Thus, this mosome, whereas their cytoplasmic tails anchor
suggests that in tissues in which both junctions the armadillo proteins, plakoglobin (PG/JUP),
are present, AJs are important in mechanosens- and plakophilins 1 – 3 (PKPs) to the desmo-
ing, whereas desmosomes are crucial for provid- somal plaque. The armadillo proteins, in turn,
ing mechanical stability under force. bind to desmoplakin (DSP), a member of the
Here, we will review the contribution of the plakin family of cytoskeleton-associated pro-
desmosome – keratin complex to mechanical teins. DSP links the desmosome to the keratin
integrity of epithelial barriers, in particular of filament network, which is essential to provide
the epidermis, and discuss their potential func- tensile strength (Fig. 2). The importance of des-
tion in sensing and transmission of forces. For mosomes for tissue integrity is highlighted by
the role of intercellular contacts and intermedi- the severe skin and cardiac defects that arise in
ate filaments (IFs) of the heart, the reader is autoimmune and genetic diseases.
A Control-si p0071-si p120-si PKP1-si PKP3-si B
Control-si
p0071-si
PKP1-si
PKP3-si
p120-si
rotation
Before
p0071
p120
rotation
PKP1
5 min
PKP3
α-Tubulin
rotation
20 min
Figure 1. Dispase-based dissociation assay highlights the importance of desmosomes for intercellular cohesion.
Only the knockdown of the desmosomal plaque protein plakophilin 1 (PKP1) severely disturbed intercellular
cohesion of mouse keratinocytes grown for 24 h in a medium containing 1.2 mM Ca2þ. The knockdown of the
corresponding proteins from adherens junctions (AJs), p120, or p0071/PKP4 did not interfere with mechanical
resistance of mouse keratinocytes (A), although the respective protein amounts were considerably decreased as
shown by western blot (B).
2 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
Desmosomes and Keratins in Tissue Mechanics
The desmosome–keratin scaffold in cell mechanics
C
B
A
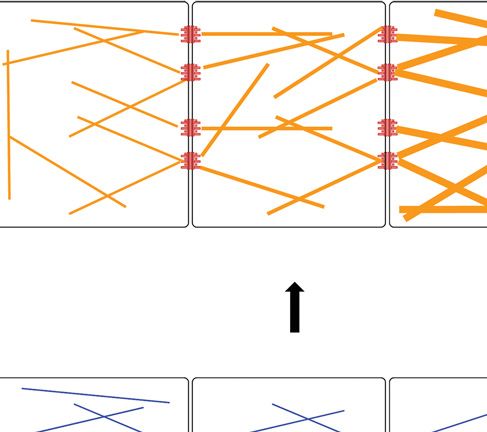
Figure 2. The desmosome –keratin complex as a micromechanical scaffold during epidermal differentiation.
(A) Expression of keratins K5/14 and interaction with desmosomal protein isoforms forms stable cohesion
among cells and protects basal keratinocytes against mechanical stress. Under conditions of tissue homeostasis,
stable desmosome –keratin scaffolds prevail. (B) Cells react to wounding (activated keratinocytes) by
modulating their micromechanical properties through altering adhesion and the cytoskeleton. Underlying
mechanisms involve altered expression of isotype proteins and posttranslational modifications that can dimin-
ish adhesion and render cells more migratory. Expression of K6/16/17 coincides with dynamic desmosomes.
(C ) To withstand increased mechanical stress, for example, in upper strata of the epidermis, number and size of
desmosomes are increased and keratin filaments become more abundant, bundled by associated proteins and
possibly elevated interkeratin Cys crosslinks. Gray arrow in (C) indicates tissue differentiation. Desmosomes are
depicted to indicate stable adhesion (red), to demarcate dynamic, less adhesive (light red), and hyperadhesive
complexes (dark red). Keratin filaments are drawn to indicate stable networks (straight blue lines), dynamic,
less stable networks (dashed green lines) or to indicate highly resilient, bundled networks (orange thick lines).
The above properties result from expression of distinct isotypes, relative abundance, and posttranslational
modifications.
Desmosomal cadherins are divided into whereas DSG2, -3, and DSC2, -3 occur primar-
four DSGs and three DSCs. The DSG – DSC het- ily in the lower layers of the epidermis (Green
erodimers represent the basic adhesive unit of and Gaudry 2000).
desmosomes (Harrison et al. 2016). All DSGs Despite their critical roles in maintaining
form adhesive dimers with all DSC isoforms epidermal adhesion and integrity, desmosomes
with affinities characteristic of each DSG – DSC are highly dynamic entities and undergo con-
pair. In contrast, homophilic DSG – DSG and stant remodeling to allow for plasticity and cell
DSC – DSC trans-interactions were suppressed migration within the epidermis during epider-
by charged amino acids. In multilayered epithe- mal differentiation and regeneration (Green
lia such as the epidermis, desmosomal cadher- and Gaudry 2000; Garrod 2010; Kowalczyk
ins reveal differentiation-dependent expression. and Green 2013). Desmosome adhesion can
DSG1, -4, and DSC1 are expressed in flattened be regulated at several levels: Their composi-
cells of the upper granular and cornified layers, tion, as well as size and number, vary among
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157 3Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
M. Hatzfeld et al.
tissues and among the individual layers of the somes (Gallicano et al. 1998). In an epidermis-
epidermis, and are controlled at the transcrip- specific DSP KO, intercellular separations were
tional, posttranscriptional, and posttransla- observed as expected (Vasioukhin et al. 2001).
tional levels. Surprisingly, desmosome number was unal-
tered although the lack of keratin association
compromised their function. PKPs were previ-
Evidence from Mouse Models and Human
ously considered as nonessential plaque pro-
Diseases
teins. However, PKP2 KO mice died around
Gene ablation in mice has established the con- day 11.5 of embryonic development because of
tribution of individual desmosomal proteins heart defects, indicating that at least one PKP
to mechanical stability and tissue integrity in is required for stable intercellular adhesion
vivo. These studies show (a) that many of the (Grossmann et al. 2004). In contrast, PKP32/2
desmosomal proteins are required for organis- mice were viable but developed hair abnormal-
mal survival and (b) that isotypes of DSGs, ities and increased inflammation of the skin,
DSCs, and PKPs have distinct functions in manifest in mice kept in a nonpathogen-free
vivo. For example, phenotypes observed after environment (Sklyarova et al. 2008). We have
ablation of the three DSC genes differ dramati- recently shown that PKP1 is essential for epider-
cally in their severity: Whereas a DSC2 knock- mal integrity in vivo. PKP1 KO mice died
out (KO) did not result in any obvious pheno- shortly after birth with defects in epidermal co-
type (Rimpler 2014), ablation of DSC1 led to hesion and barrier formation (Rietscher et al.
epidermal fragility with hyperproliferation and 2016). Desmosomes were small and sparse in
dermatitis, but mice were viable and fertile the skin, as well as in cultured keratinocytes de-
(Chidgey et al. 2001). In contrast, DSC3 abla- rived from these mice, and cell separation oc-
tion resulted in preimplantation lethality, sug- curred in the granular layers.
gesting a desmosome-independent role during Additional support for the importance of
early development (Den et al. 2006). Similarly, desmosomal adhesion in tissue integrity comes
ablation of DSG isotypes revealed distinct func- from monogenetic human diseases. Ectoder-
tions in vivo: Whereas the DSG2 KO was em- mal dysplasia-skin fragility syndrome (EDSFS,
bryonic lethal (Eshkind et al. 2002), DSG3 KO MIM604536), a genetic disease caused by PKP1
mice showed weakened desmosomal adhesion, mutations (McGrath et al. 1997), is character-
leading to the separation of keratinocytes (Koch ized by skin fragility with generalized superficial
et al. 1997). The loss of DSG1 function has so far erosions and chronic inflammatory plaques and
not been analyzed in mice as it requires deletion pruritus. Additional abnormalities include alo-
of all three DSG1 genes. pecia and nail dystrophy (McGrath et al. 1997;
Desmosomal plaque proteins are equally Sprecher et al. 2004; McGrath 2005; McGrath
important for intercellular cohesion and me- and Mellerio 2010). Desmosomes in the skin
chanical stability: PG/JUP KO embryos died of patients were generally small with perturbed
as a result of severe heart defects with reduced desmosome – keratin interactions. Severe der-
and structurally altered desmosomes (Bierkamp matitis, multiple allergies and metabolic wast-
et al. 1996; Ruiz and Birchmeier 1998). Al- ing (sinobronchial allergic mycosis, SAM) syn-
though b-catenin, the homologue of PG in drome (MIM615508) is a recently described
AJs, localized to desmosomes and could substi- genodermatosis caused by homozygous muta-
tute for PG in cadherin clustering, it failed to tions in DSG1 or DSP. Dermatologic manifesta-
recruit normal levels of PKP1 and DSP to the tions comprise congenital erythroderma, striate
plaque (Bierkamp et al. 1999; Acehan et al. palmoplantar keratoderma (SPPK), skin ero-
2008). DSP KO embryos did not survive beyond sions, scaling, and hypotrichosis. Patients devel-
embryonic day 6.5 and displayed severe defects oped severe allergies, recurrent skin, and respi-
in tissue architecture, shaping of the embryo, ratory tract infections, indicating that loss of
and in anchoring keratin filaments to desmo- DSG1 interferes with skin barrier function (Sa-
4 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
Desmosomes and Keratins in Tissue Mechanics
muelov et al. 2013; Has et al. 2015). More re- and DSCs. Using analytical ultracentrifugation
cently, a point mutation in the keratin-binding and plasmon plasmon surface resonance reso-
domain of DSP was identified as causing SAM nance, these investigators determined KD-values
syndrome (McAleer et al. 2015). Nonsense, as for DSG and DSC homodimers, as well as het-
well as splice site mutations, have been observed erodimers. Whereas homodimers were very
in PG causing skin fragility, generalized epider- weak with KD values .400 mM, heterodimeric
molysis, palmoplantar keratoderma, and wool- pairs revealed KDs ranging from 3.6 to 43.9 mM,
ly hair (Pigors et al. 2011; Li et al. 2012). Ar- indicative of stronger adhesion than provided by
rhythmogenic cardiomyopathy (AC) is a rare E-cadherin (Harrison et al. 2016). These data
disease of the heart characterized by progres- correlate well with the role of AJs in mechano-
sive myocardial dystrophy with fibro-fatty re- sensing and one for desmosomes in conferring
placement. In the majority of cases, dominant stability. Interestingly, the strongest-binding
mutations have been identified in desmosomal pairs were Dsg1:Dsc1 and Dsg4:Dsc1, which
genes including DSP: ARVD8 (MIM607450); are expressed in the outermost layers of the epi-
PKP2: ARVD9 (MIM609040); DSG2: ARVD10 dermis, whereas the basally expressed cadherins
(MIM610193); DSC2: ARVD11 (MIM610476); Dsg3:Dsc3 formed the weakest adhesive pair.
JUP: ARVD12 (MIM611528) (Al-Jassar et al. This correlates with the force-protective func-
2013; Cerrone and Delmar 2014; Calore et al. tion of the outer epidermal layers, although the
2015; Pilichou et al. 2016). basal layers must allow for remodeling and re-
Desmosomes can also be affected in ac- generation, which is facilitated by weaker adhe-
quired diseases, which comprise epidermal au- sion. In agreement, preliminary experiments
toimmune disorders and infections (Stahley suggest an up-regulation of DSG1 on applica-
and Kowalczyk 2015). Pemphigus is a family tion of stretch to cultured keratinocytes (our
of diseases characterized by circulating autoan- own unpublished results).
tibodies that target desmosomal proteins and Similarly, PKPs -1 and -3 contribute differ-
compromise cell – cell adhesion (Amagai 2010; entially to intercellular cohesion. A comparison
Jennings et al. 2011; Amagai and Stanley 2012). of keratinocytes derived from the correspond-
Pemphigus autoantibodies are directed against ing KO mice confirmed that the loss of PKP1
the extracellular domains of desmosomal cad- interfered with formation of a stable epithelial
herins and are sufficient to cause the loss of sheet, whereas adhesion was not compromised
keratinocyte adhesion. in PKP3 KO keratinocytes. Surprisingly, desmo-
Taken together, these studies clearly indicate somal PKP3 was not only much more dynamic
that desmosomes are essential for robust inter- compared with PKP1, but was able to destabi-
cellular cohesion and tissue integrity under me- lize PKP1-dependent desmosomes when over-
chanical strain. expressed (Keil et al. 2016). Collectively, these
data indicate that isoform expression has a con-
Why is Desmosomal Adhesion Stronger siderable influence on desmosome stability and
than AJ-Mediated Adhesion? resistance to force and appears well suited to
adapt desmosomes to mechanical stresses.
Isoform-Dependent Composition
of Desmosomes Determines Adhesive
Strength Hyperadhesion is a Unique Feature
of Desmosomes
Although the differential expression of desmo-
somal cadherins and PKPs has been known for a In contrast to AJs, desmosomes can occur in
long time, experiments addressing their indi- two functionally distinct adhesive states, which
vidual and distinct contributions to desmoso- are distinguished on Ca2þ-depletion. In normal
mal adhesion have only recently emerged. Har- tissues, desmosomes adopt a Ca2þ-indepen-
rison et al. (2016) have systematically analyzed dent state, also referred to as hyperadhesion,
the structural basis of adhesive binding by DSGs which is crucial for stable intercellular cohesion
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157 5Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
M. Hatzfeld et al.
and resistance to mechanical strain (Garrod a role of DSG2 in desmosome maturation and
2010; Thomason et al. 2010). In contrast, dur- in the switch to Ca2þ-independent adhesion
ing regeneration and wound healing, desmo- (Lowndes et al. 2014).
somes become Ca2þ-dependent resulting in Because an ordered intercellular zone re-
weaker intercellular cohesion, which allows for quires an ordered plaque, desmosomal plaque
tissue remodeling (Wallis et al. 2000; Kimura proteins are also important in regulating desmo-
et al. 2012). In cultured keratinocytes, desmo- somal hyperadhesion. PKP1 overexpression in-
somes depend on extracellular Ca2þ after their duced Ca2þ-independent hyperadhesive des-
formation but undergo a maturation process to mosomes. This prevented Pemphigus vulgaris
become hyperadhesive. Although the mecha- (PV)-IgG-mediated desmosome disruption
nism controlling formation of Ca2þ- insensitive (Tucker et al. 2014), indicative of a very stable
desmosomes is incompletely understood, one adhesive state. Moreover, PKP1 enhances des-
model predicts that a highly organized arrange- mosome size and number by laterally interact-
ment of the DSG and DSC extracellular do- ing with DSP in agreement with a postulated role
mains is induced (Thomason et al. 2010), which of plaque proteins in mediating cadherin spac-
correlates with a wider intercellular space and ing (Kowalczyk et al. 1999; Hatzfeld et al. 2000;
the presence of a midline observed in electron Bornslaeger et al. 2001; Hatzfeld 2007). Thus,
microscopic images, whereas Ca2þ-dependent the desmosomal cadherins, as well as the plaque
desmosomes appear to lack this midline (Gar- protein PKP1, cooperate to provide strong ad-
rod 2010). hesion and stability under mechanical strain.
Based on structural analyses of DSG2, Tariq
et al. (2015) proposed that the flexibility of des- What are the Upstream Signals that Modulate
mosomal cadherin ectodomains allows them to Desmosomal Adhesion?
adopt a conformation that facilitates desmo-
Cross Talk Desmosomes-AJ
some plasticity. The desmosomal plaque would
contribute to plasticity by introducing spacings Epithelial junction assembly, maturation, and
between desmosomal cadherins and preventing maintenance occur through sequential process-
ectodomain cis-interactions. The distance be- es that are differentially regulated (Keil et al.
tween cadherins and the lack of cis-interactions 2016). Earlier work suggested a hierarchical
facilitates conformational changes needed to process in which AJs form before desmosomes.
adopt the ordered architecture in the Ca2þ-in- However, increasing evidence indicates that
dependent state. Depletion of DSC2 prevented newly formed contacts contain components of
the acquisition of hyperadhesion in keratino- AJs and desmosomes, and sorting into distinct
cytes (Kurinna et al. 2014), although it is not junctional complexes occurs on junction mat-
known whether this is a unique feature of DSC2 uration in epithelial cells (Green et al. 2010; Keil
or shared by all DSCs and DSGs. The fact that a et al. 2016).
DSC2 mouse KO did not reveal any phenotypic PG was the first protein identified in desmo-
abnormalities (Rimpler 2014) suggests that hy- somes and AJs based on colocalization studies
peradhesion does not critically depend on the and its coimmunoprecipitation with E-cadherin
DSC2 isoform. In an attempt to characterize the (Cowin et al. 1986; McCrea et al. 1991). Gosavi
combinatorial roles of DSCs and DSGs in des- et al. (2011) showed that PKP3, PG, and E-cad-
mosome assembly, micropatterned substrates herin are present at the cell border when cells are
were used to uncouple desmosome assembly grown in media with low concentrations of Ca2þ
from other cell contacts (Lowndes et al. 2014). and suggested a model in which PG and E-cad-
In this assay, DSC2, but not DSG2, formed herin recruit PKP3 to the cell border to initiate
Ca2þ-dependent homophilic bonds, indicating desmosome formation. A role of PKP3 in coor-
that DSC2 was required for desmosome assem- dinating AJ and desmosome formation was fur-
bly. In contrast, DSG2 formed Ca2þ-indepen- ther supported by the finding that PKP3 locali-
dent heterophilic bonds with DSC2, suggesting zation at the plasma membrane occurs together
6 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
Desmosomes and Keratins in Tissue Mechanics
with E-cadherin and precedes other desmo- Mechanosensitive Signaling Pathways in the
somal proteins, including PKP1 and DSP (Keil Control of Desmosomes
et al. 2016). At the molecular level, PKP3 forms a
complex with the Rap1 GTPase, promoting its So far, little is known about how desmosome
activation and facilitating desmosome assembly gene transcription and isotype expression are
(Todorovic et al. 2014). Moreover, PKP3 was controlled and how mechanical stimuli direct
required for AJ maturation and sealing. These these processes. Several recent studies have
findings reveal PKP3 as a coordinator of desmo- addressed the question how external mechani-
some assembly and AJ maturation through its cal forces are transmitted into chemical signals.
association with Rap1 (Todorovic et al. 2014). To induce changes in the cell’s gene expression
program, signaling molecules regulated by
external forces must be translocated from the
Actin and Desmosomes cytoplasm into the nucleus and nucleocytoplas-
Formation of AJs but not desmosomes coin- mic shuttling has been suggested as a common
cides with an increase in the apparent stiffness theme in mechanotransduction (Sharili and
of cell monolayers reflecting the generation of a Connelly 2014). The SRF (serum-response fac-
tissue-level tension (Harris et al. 2014). Tension tor) and YAP (Yes-associated protein)/TAZ
rapidly increases with a maximum at 150 min, (transcriptional coactivator with PDZ-binding
which correlates with the initiation of desmo- motif ) pathways are known mediators of this
some assembly. Although desmosomes are as- process in multiple cell types, including kerati-
sociated with IFs and according to current views nocytes.
are unable to generate tension, their assembly SRF transcriptional activity is regulated by
requires actin microfilaments (Pasdar and Li actin and RhoA and its cofactors MRTF (myo-
1993). It is, therefore, possible that AJ-associat- cardin-related transcription factor) -A and -B.
ed actomyosin-dependent force generation is an Monomeric actin sequesters MRTFs, which are
important signal for desmosome formation al- released on actin polymerization and accumu-
though direct experimental evidence remains to late in the nucleus where they interact with SRF
be adduced. to stimulate target gene expression. Intercellular
The AJ protein p120 plays an essential role tension can increase actin polymerization at AJs
in limiting actomyosin-dependent tension gen- and drive MRTF-A translocation into the nu-
erated at AJ as shown by uncoupling p120’s cleus (Gomez et al. 2010). SRF and MRTFs co-
cadherin-stabilizing and RhoA-suppressing ac- activate immediate early genes, such as JunB,
tivities. Removing p120’s Rho-suppressing ac- and adhesion-related genes, including vinculin.
tivity lead to excessive actomyosin contractility Mice with a keratinocyte-specific SRF-KO re-
along the vertical axis of cells and disrupted the vealed severe intercellular gaps between kerati-
integrity of the apical surface, irrespective of E- nocytes as the most striking pathophysiology
cadherin stability (Yu et al. 2016). Interestingly, (Koegel et al. 2009). The number of desmo-
p120 has been described to associate with des- somes was significantly lower in mutant com-
mosomes during their formation in a Ca2þ- pared with control mouse skin, whereas the size
shift experiment (Kanno et al. 2008). of individual desmosomes was unaltered. How-
Desmosomes can also influence actin orga- ever, a direct regulation of desmosomal genes
nization: Loss of any of the PKPs from human was not supported by transcription profiles of
or mouse keratinocytes results in changes in SRF KO skin samples compared with control
cortical actin organization (Godsel et al. 2010; skin (Koegel et al. 2009). In contrast, loss of
Keil et al. 2016). However, it is not clear whether MRTF-A reduced the levels of DSG1 messenger
PKPs regulate RhoA activity and stress fiber for- RNA (mRNA) in keratinocytes, suggesting that
mation directly or indirectly, by influencing the DSG1 is directly regulated by the SRF/MRTF-A
localization or activity of a Rho GEF or a Rho complex (Dubash et al. 2013). Another experi-
GAP (Godsel et al. 2010). ment showed that MRTF-A and SRF were
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157 7Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
M. Hatzfeld et al.
recruited to cis-regulatory elements of the PKP2 also respond to the rigidity of the extracellular
gene to regulate its expression (Leitner et al. matrix, cell geometry, cell density, cell polarity,
2011). Taken together, these data suggest that and the status of the actin cytoskeleton. Sensing
RhoA/MRTF-A signaling in keratinocytes af- of substrate stiffness by YAP/TAZ depends on
fects the expression of desmosomal proteins di- the tension of the actin cytoskeleton (Zhang
rectly and indirectly although the regulation via et al. 2011; Elbediwy et al. 2016). The AJ protein
SRF/MRTF-A in response to mechanical strain a-catenin limits YAP activity by modulating its
has not been directly shown. interaction with 14-3-3 (Kanai et al. 2000; Schle-
The transcription factor AP-1 mediates gene gelmilch et al. 2011; Sambandam et al. 2015).
expression in response to a variety of extracel- YAP/TAZ mediate their function in controlling
lular stimuli and is activated by SRF/MRTF sig- gene expression through interaction with TEAD
naling (Wang et al. 2015). Microarray analysis transcription factors, which drive the expression
performed on mechanical stretched and normal of proliferative genes (Zhang et al. 2011). Al-
human skin revealed an up-regulation of AP-1 though YAP activation accelerates proliferation
in mechanically stretched skin (Yang et al. and suppresses differentiation of mouse kerati-
2011). Other stimuli of AP-1 include growth nocytes, which correlates with altered desmo-
factor signaling via mitogen-activated protein some number, size, and composition, a role of
(MAP) kinases, which are also activated in re- YAP/TAZ/TEAD in the transcriptional regula-
sponse to mechanical stress (Kippenberger et al. tion of desmosomal genes has not been directly
2000). AP-1 acts as a homodimeric or hetero- addressed. However, a large scale ChIP-seq anal-
dimeric transcription factor composed of mem- ysis of Tead4 target genes in ECC1 endometrial
bers of the Fos (c-Fos, FosB, Fra-1, and Fra-2) cells identified DSCs 1-3, DSG1, DSP, PG, and
and Jun (c-Jun, JunB, and JunD) families. Jun is PKPs -1 and -2 as putative Tead4 target genes
regarded as a positive regulator of keratinocyte (Liu et al. 2016).
proliferation/differentiation during develop-
ment and in skin cancer through its direct tran-
scriptional effect on epidermal growth factor ROLE OF IFs IN MECHANICAL STABILITY
receptor (EGFR) expression. In contrast, JunB
Composition, Assembly, and Organization
can antagonize proliferation of keratinocytes.
of Keratin Filaments
AP-1 target genes include keratins (Yates and
Rayner 2002). IFs can be assembled from a superfamily of
The Hippo signaling pathway is another reg- approximately 70 proteins that form cell-type-
ulator of mechanotransduction. It regulates or- specific cyto- and nucleoskeletal arrays in the
gan size, tissue regeneration, and stem cell re- majority of multicellular animal species. Based
newal (Zhang et al. 2011; Barry and Camargo on sequence homology and expression, IFs are
2013). Recently, mechanical signals were found grouped into six classes, all of which except lam-
to be transduced by the two mediators of the ins form predominantly cytoplasmic assemblies
Hippo pathway, the transcriptional coactivators (Schweizer et al. 2006; Hobbs et al. 2016). Here,
YAP and TAZ (Dupont et al. 2011). In the skin, we focus on epithelial keratins to examine their
YAP is required for epidermal barrier formation, contribution to the mechanical resilience of ep-
hair follicle development, and maintenance of ithelia in conjunction with desmosomes, al-
the epidermal stem cell compartment (Zhang though desmin and vimentin also interact with
et al. 2011; Barry and Camargo 2013; O’Neill desmosomes in cardiac tissue, meningiomal,
2015). Overexpression of YAP in transgenic and arachnoidal cells in which they fulfill anal-
mice promotes squamous cell carcinoma for- ogous functions (Kartenbeck et al. 1983, 1984).
mation (Chan et al. 2011; Jia et al. 2016), where- Keratin assembly begins with heterodimeri-
as its knockdown inhibits cutaneous wound zation of type I and II keratin molecules, fol-
healing (Lee et al. 2014), suggesting a central lowed by formation of antiparallel tetramers.
role in regulating proliferation. YAP and TAZ The lateral association of four tetramers gener-
8 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
Desmosomes and Keratins in Tissue Mechanics
ates so-called unit-length filaments (ULFs), follicle, and are characterized by the expression
which longitudinally coalesce into mature fila- of KRT15 (Lyle et al. 1998; Watt 1998; Goldstein
ments with a propensity to bundle and organize and Horsley 2012). The vibrissae bulge harbors
into three-dimensional (3D) networks (Herr- two types of slow cycling stem cells, character-
mann et al. 2009; Koster et al. 2015; Herrmann ized by the expression of KRT5/15/17/19 and
and Aebi, 2016). Keratins show extensive se- KRT5/17, respectively, which display loose ker-
quence diversity, a feature that discriminates atin bundles in the former and tight bundles
them from actins and tubulins (Schweizer et in the latter configurations (Larouche et al.
al. 2006; Fletcher and Mullins 2010). This se- 2008). Notably, in both subpopulations lacking
quence diversity is responsible for their relative KRT14, keratin network organization is differ-
affinities to each other and to associated pro- ent. The complexity of keratin expression sug-
teins, and enables isotype-specific posttrans- gests that keratins contribute to an intracellular
lational modifications (Hatzfeld and Franke epithelial niche, analogous to a stem cell niche
1985; Hofmann and Franke 1997; Snider and (Tumbar et al. 2004), that endows distinct epi-
Omary 2014). In humans and the mouse, 28 thelial cells with unique micromechanical prop-
type I keratins (KtyI) and 26 type II keratins erties through formation of specifically tailored
(KtyII) genes are predominantly expressed as keratin-desmosome networks (Fig. 2).
pair-specific combinations of types I and II ker-
atin proteins, respectively, in epithelial cells and
Human Disease and Mouse Models
tissues. The major impact of keratins on cell
integrity and adhesion is most obvious in the The importance of keratins for providing me-
epidermis, in addition to embryonic epithelia chanical stress resilience in the epidermis is best
(Simpson et al. 2011; Bouameur and Magin documented by blistering and hyperkeratotic
2017). The basal compartment expresses the skin disorders exemplified by Epidermolysis
keratin pair KRT5/KRT14, organized in loose bullosa simplex (EBS; OMIM #131900), caused
bundles that extend from hemidesmosomes by mutations in KRT5 and KRT14 (Szeverenyi
and desmosomes. On terminal differentiation, et al. 2008). EBS is characterized by cytoplasmic
these keratinocytes disconnect from the ECM keratin aggregates, cytolysis of basal keratino-
accompanied by replacement of KRT5/KRT14 cytes, and bullous lesions following mild trau-
with KRT1/KRT10. Depending on regional dif- ma to the skin. Although it is recognized that
ferences in the epidermis, additional keratins the pathomechanisms contributing to EBS and
become expressed. At sites of high mechanical additional keratinopathies are more complex
strain, such as palms and soles, the keratins than originally considered (Coulombe and Lee
KRT1/KRT10 are supplemented by keratins 2012; Roth et al. 2012; Bohnekamp et al. 2015;
KRT2e and KRT9 (Moll et al. 2008). Analogous Hobbs et al. 2016; Kumar et al. 2016), it is evi-
to desmosomes, keratin concentration increases dent that loss of an intact keratin cytoskeleton
from 40 mg/ml in basal keratinocytes to prob- renders keratinocytes fragile on mild physical
ably twice as much in terminally differentiated stress, shown by KRT5 and KRT14 KO mice
keratinocytes (Sun and Green 1978; Feng et al. (Lloyd et al. 1995; Peters et al. 2001). Of note,
2013). The above keratin expression pattern var- even mutations causing severe disease do not
ies, for example, during epidermal injury, which prevent formation of long keratin intermediate
triggers the rapid induction of KRT6, -16, and filaments (KIFs) in vitro (Herrmann et al.
-17 at the wound edge at the expense of KRT1/ 2002), suggesting that mutations and physical
10, accompanied by fewer and less adhesive des- stress act at the level of keratin bundling, net-
mosomes (Garrod and Chidgey 2008; Simpson work organization, dynamics, or by affecting
et al. 2011). KRT6, KRT16, and KRT17 are also association with other proteins. Indeed, the
expressed in hair follicles and the nails (Moll et most frequent KRT14 Arg125 mutation com-
al. 2008). Epidermal stem cells are located in promises desmosome adhesion (Russell et al.
protected niches, such as the bulge of the hair 2004; Homberg et al. 2015).
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157 9Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
M. Hatzfeld et al.
Deletion and/or mutation of other keratin (Gallicano et al. 1998; Vasioukhin et al. 2001;
isotypes expressed in different epidermal com- Sumigray and Lechler 2012). Moreover, DSP
partments, either in the mouse or in humans, and DSG1 mutations can give rise to SPPK
did not cause cytolysis and skin blistering to the (OMIM #612908, #148700, respectively), a hy-
same extent as KRT5 and KRT14, but was typ- perkeratotic skin condition with fewer or small-
ified by altered cell and tissue growth, barrier, er desmosomes in the suprabasal epidermis and
and immune defects (Reichelt et al. 2001; perinuclear accumulation of KIF in DSP-asso-
McGowan et al. 2002; Lessard and Coulombe ciated SPPK (Wan et al. 2004). Conversely, de-
2012; Roth et al. 2012; Fischer et al. 2014, 2016; letion of keratins reduced DSP deposition at the
Fu et al. 2014; Kumar et al. 2016; Bouameur and plasma membrane (Loranger et al. 2006; Vi-
Magin 2017). Transgenic expression of a bun- jayaraj et al. 2009). Distinct steps during desmo-
dling-competent KRT5/KRT8 chimaera par- some assembly also require a transient but stable
tially rescued KRT52/2 mice, whereas expres- interaction between DSP and KIF in the cyto-
sion of bundling-deficient KRT8 did not, plasm (Godsel et al. 2005; Hobbs and Green
providing strong evidence that, in addition to 2012; Albrecht et al. 2015).
keratin abundancy, isotype-specific properties Keratins interact with the plectin and DSP
and keratin network organization are crucial carboxy termini through segments of their rod
contributors to keratin-mediated stress resil- domain (Meng et al. 1997; Fontao et al. 2003;
ience (Alvarado and Coulombe 2014). Bouameur et al. 2014). Association with kera-
The known redundancy of keratin ex- tins is regulated by serine phosphorylation and
pression in part explains lack of similarly severe arginine methylation of the DSP tail, involving a
defects in other epithelial compartments and coordinated activity of glycogen synthase kinase
complicates the analysis of isotype-specific 3 (GSK3) and protein arginine methyltransfer-
functions (Reichelt et al. 2001; McGowan et al. ase 1 (PRMT1). Inhibition of GSK3 or PRMT1
2002; Lessard and Coulombe 2012; Roth et al. delayed desmosome assembly and enhanced
2012). To overcome the latter, mice lacking the DSP–KIF interactions in the cytoplasm (Al-
entire KtyI and KtyII genes in their epidermis brecht et al. 2015). In the absence of keratins,
were generated, in addition to mice lacking both using keratinocytes from keratin-deficient mice
KRT1 and KRT10. Such mice developed a fully (Vijayaraj et al. 2009), DSP phosphorylation
stratified epidermis but died perinatally because was elevated and desmosomes were endocy-
of extensive epidermal damage (Wallace et al. tosed at accelerated rates. Such desmosomes
2012; Bar et al. 2014; Kumar et al. 2015). Most were unable to render epithelial sheets stable
importantly, these mice showed diminished in- on rotational stress (Kroger et al. 2013). Reex-
tercellular adhesion and significantly smaller pression of KRT5 and KRT14 reconstituted
desmosomes. This was accompanied by accu- properly formed desmosomes and shear-resis-
mulation of desmosomal proteins in the cyto- tant intercellular adhesion. Preliminary evi-
plasm, highlighting that in vivo interaction with dence indicates that sequestration of protein ki-
keratins was required for the maintenance of nase C-a (PKCa) by the scaffold protein Rack1
functionally intact desmosomes. and KRT5/14 contribute to the stabilization of
desmosomes (Kroger et al. 2013), whereas PKP3
may recruit PKCa to abrogate hyperadhesion
Interdependence of Keratins and
(Keil et al. 2016). In agreement, loss of PKCa
Desmosomes
in mice delays reepithelialization following
Desmosomes have been recognized early on as wounding and is accompanied by maintenance
important sites for KIF formation and organi- of hyperadhesive desmosomes (Thomason et al.
zation in vivo (Jackson et al. 1980; Bologna et al. 2012).
1986; Schwarz et al. 2015), and genetic deletion Reepithelialization requires not only altered
of DSP can cause extensive reorganization or adhesion of keratinocytes at the wound edge
collapse of KIF, depending on the cell type (Shaw and Martin 2009) but also diminished
10 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
Desmosomes and Keratins in Tissue Mechanics
contacts of keratins to desmosomal proteins, well accepted, the contribution of keratins to the
along with elevated expression of KRT6/16/17 resilience of epithelia against various types of
and a decrease in KRT1/10 (Patel et al. 2006). In deformation remained unknown. Unlike other
agreement, cells expressing KRT6 or KRT17 cytoskeletal elements, IF networks are less rigid
show elevated, PKCa-mediated desmosome at low shear strain but harden at high strains and
disassembly and subsequent destabilization of resist breakage, indicating that they are crucial
epithelial sheets. In contrast, KRT5 or KR14 for the maintenance of tissue integrity (Janmey
supported stable desmosomes, suggesting that et al. 1991, 2013). Using atomic force microsco-
expression of “wound healing” keratins weakens py (AFM)-based single-cell compression to ex-
intercellular adhesion. Further studies suggest amine resistance against external pressure and
that the type II keratin KRT5 is a major deter- global rupturing forces, keratinocytes were 6 –
minant of desmosome stability (Loschke et al. 70 times stiffer than most other cell types. Selec-
2016). In addition to PKCa, p38 MAP kinase tive disruption of actin filaments and microtu-
(MAPK) signaling downstream from the serine bules did not significantly alter keratinocyte
protease inhibitor SPINT1 participates in the mechanics (Lulevich et al. 2010). However, ap-
interaction of keratins and desmosomal pro- plication of AFM to keratin-deficient individual
teins (Kawaguchi et al. 2015). keratinocytes revealed a significant softening
Thus, available data provide strong evidence of keratin-deficient cells (Ramms et al. 2013).
for an interdependence of keratins and DSP Magnetic tweezer experiments additionally
regulated by posttranslational modifications. showed a major contribution of keratins to vis-
The casein kinase I (CK-1a) scaffold protein coelastic cell properties. Similar conclusions
FAM83H regulates keratin networks by recruit- were reached using a microfluidic optical
ing CK-1a directly to keratins (Kuga et al. stretcher (Seltmann et al. 2013). To relate these
2013). It may be additionally involved in mem- findings to functional consequences, invasion
brane localization of desmosomal proteins and and 3D growth assays were performed with nor-
in regulating desmosome – keratin interactions mal and keratin-deficient keratinocytes, reveal-
(Kuga et al. 2016). Whether FAM83H activity ing higher invasiveness in the latter. However,
toward keratins or desmosomal proteins is this came at the price of increased disruption
coordinated with force-dependent conforma- of cells during invasion, suggesting that certain
tional changes in desmosomal proteins remains levels of keratins or other IF proteins are re-
currently unknown. In Xenopus mesendoderm quired to sustain the invasive process (Cheung
cells, increased tension at P-cadherin-contain- et al. 2013; Seltmann et al. 2013).
ing junctions recruits PG/JUP to these sites. In Beyond single cells, very little is known
turn, PG/JUP recruits KIF to these junctions to about the contribution of keratin architecture
reorganize and reinforce the keratin cytoskele- to tissue mechanics. A recent AFM-based study,
ton. Through this sequence of events, local forc- using the human hair follicle as a model system,
es from neighboring cells mediate keratin reor- showed stiffening of the soft keratinocyte matrix
ganization required for coordinated cell at the base of the hair follicle 360-fold, from
behavior (Weber et al. 2012). 30 kPa to 11 MPa along the first millimeter of
the follicle. This stiffening coincided with an
increased thickness of keratin macrofibrils and
Keratins as Main Determinants for the
their orientation. The continued stiffening like-
Mechanical Integrity of Keratinocytes and
ly is because of increasing network orientation,
Tissues
compaction, and mechanical reinforcement of
The mechanical properties of epithelial cells the keratin macrofibrils by disulfide cross-links.
largely depend on actin filaments, microtubules, This study links changes in the architecture and
and KIF (Fletcher and Mullins 2010; Koster et al. molecular structure of keratin macrofibrils to
2015). Although the contribution of actin fila- the local mechanical behavior at a tissue scale
ments and microtubules to these properties is level (Bornschlogl et al. 2016).
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157 11Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
M. Hatzfeld et al.
Impact of Mechanical Stretch on the response of keratins, affecting predominantly
Desmosome –Keratin Complex the peripheral keratin network (Fois et al.
2013). A set of studies has investigated bio-
Exposure of keratinocytes to stretch results physical and physiological consequences of
in rapid and transient induction of MAPK hyperphosphorylation on normal and mutant
ERK1/2 and SAP kinases (Kippenberger et al. keratins. The bioactive lipid sphingosylphos-
2000; Yano et al. 2004), accompanied by altered phorylcholine (SPC) activates JNK and Erk ki-
keratin expression, increased keratin phosphor- nases, mediating keratin reorganization through
ylation and reorganization of keratin networks phosphorylation of KRT8 at Ser431 and KRT18
(Yano et al. 2004; Snider and Omary 2014). At at Ser52 in pancreatic tumor cells. The resulting
least in keratinocytes, mechanical stretching perinuclear keratin reorganization affects the
acts via induction of calcium influx, EGFR viscoelasticity of metastatic cancer cells and pro-
phosphorylation, and ERK1/2 activation motes tumor invasion (Beil et al. 2003). Me-
(Yano et al. 2004). In this setting, the established chanically stretching of keratinocytes expressing
role of DSG1 in EGFR signaling has not yet been EBS-like keratin mutations triggered a progres-
examined, thus, it remains unknown whether sive disassembly of desmosomes and weakened
keratin reorganization is mediated by desmo- intercellular adhesion (Russell et al. 2004; Hom-
somes or affects them (Harmon et al. 2013). berg et al. 2015). Thus, keratin reorganization
On mechanical stretch, keratinocytes expressing on posttranslational modifications might con-
the disease mutation KRT10 Arg156His show tribute to the invasive properties of metastatic
stronger activation of p38 MAPK compared tumor cells and to altered adhesion in skin dis-
with control transfectants, which ultimately orders (Beil et al. 2003; Russell et al. 2004; Hom-
might drive hyperproliferation typical of the berg et al. 2015; Loschke et al. 2016).
corresponding keratinopathy (Obarzanek-Fojt
et al. 2011). In lung epithelial cells, shear stress,
but not stretch, caused hyperphosphorylation Intrinsic Keratin Properties and Associated
and disassembly of KRT8 and KRT18-contain- Proteins as Determinants of Network
ing IF, regulated by PKC-d (Ridge et al. 2005). Organization and Stress Resilience
Blocking the shear stress – mediated keratin hy- The organization of cytoskeletal networks de-
perphosphorylation and reorganization de- pends on their intrinsic properties and their
creased cell viability and increased apoptosis. associated proteins, and determines the com-
Notably, shear stress induced bundling of KIF prehensive array of functions that they perform
into thick fibrils. In this study, shear stress – me- (Fletcher and Mullins 2010). Bundling of indi-
diated phosphorylation of KRT18 at Ser33 was a vidual filaments into fibers, an intrinsic keratin
prerequisite for keratin network reorganization property (Lee and Coulombe 2009), is seen in
to alter cells’ mechanical properties and sustain most epithelia, evident in the epidermis and in
their integrity (Flitney et al. 2009; Sivaramak- hair follicles, whereas simple epithelia show less
rishnan et al. 2009). dense arrays. Here, an interesting contributor to
To understand how phosphorylation affects IF organization lies in their capacity to form
keratin reorganization, select phosphomutants disulfide bonds. With the exception of KRT8,
of KRT8 and KRT18 were characterized for their KRT18, and KRT19, all other human and
network mechanical properties using rheology mouse keratins contain variable numbers of cys-
and electron microscopy. Phosphokeratin-con- teine residues. Following the observation that a
taining networks showed reduced intraconnec- subset of keratins becomes disulfide-cross-
tivity and resulted in mechanically weaker and linked on terminal differentiation in vivo, ho-
more deformable networks in vitro. This ap- motypic disulfide bonds involving three Cys res-
peared to result from the formation of shorter idues in KRT14 and one in K10 were identified
mutant filaments (Deek et al. 2016). In cultured in cultured keratinocytes and mouse epidermis,
cells, hyperphosphorylation alters the stretch in addition to interkeratin disulfide-bonded
12 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
Desmosomes and Keratins in Tissue Mechanics
KRT5/14 species (Lee et al. 2012; Feng and Cou- domains may uncover the hidden SH3 domain
lombe 2015; Bunick and Milstone, 2016). Avail- (Osmani and Labouesse 2015).
able data point to multiple roles of disulfide- Very little is known about the interaction of
bonding on keratin assembly, dynamics, and keratins with actin and actomyosin, which is
network organization. Intriguingly, disulfide crucial during morphogenesis, cell polarity,
bonding of K14 coincided with formation of cell migration, and invasion. In a study aimed
networks enriched in the nuclear periphery, a to normalize mutation-disrupted keratin net-
region reactive to mechanical cues (Feng and works in hepatocytes, the multikinase inhibitor
Coulombe 2015; Wallrath et al. 2016). Disulfide PKC412 was found to perform this function by
bonding may also alter bending and buckling of enhancing nonmuscle myosin heavy chain-IIA
keratin networks, which occur on compressive (NMHC-IIA) interaction with KRT8 and KRT
intracellular forces (Nolting et al. 2014). Finally, 18 through inhibiting NMHC-IIA phosphory-
disulfide bonds in keratins not only alter keratin lation (Kwan et al. 2015). Solo (ARHGEF40) is a
organization and dynamics on mechanical sig- RhoA-targeting guanine nucleotide exchange
nals but could tie them to the highly dynamic factor (GEF) involved in cyclical stretch-induced
cellular redox network controlled by the antiox- human endothelial cell reorientation and con-
idant transcription factor Nrf2 (Schafer et al. vergent extension cell movement in zebrafish
2012). gastrula and binds to KRT8/18 IF. Knockdown
In addition to posttranslational modifica- of Solo suppresses tensile force– induced stress
tions, the small heat shock protein Hsp27 and fiber reinforcement and RhoA activation. Thus,
proteins of the plakin and S100 families regulate the interplay between Solo and KRT8/18 plays a
keratin assembly, bundling, and network orga- crucial role in tensile force– induced RhoA acti-
nization (Windoffer et al. 2011; Kayser et al. vation and consequent actin cytoskeletal rein-
2013; Bouameur et al. 2014; Lesniak and Grac- forcement (Fujiwara et al. 2016).
zyk-Jarzynka 2015). Plectin is a plakin protein
that links keratin bundling to MAPK signaling.
In contrast to prediction, genetic deletion of CONCLUDING REMARKS
plectin rendered cells more susceptible to me-
Desmosomes as Mechanosensors?
chanical stress, despite an increase in keratin
bundling (Osmanagic-Myers et al. 2006). The As mentioned above, AJs are considered as the
latter is consistent with unaltered mechani- mechanosensing unit in intercellular force
cal properties of plectin knockdown cells as transduction and distribution in multicellular
assessed by indentation analyses using AFM tissues. It remains an open question whether or
and by displacement analyses of cytoplasmic not desmosomes can function as mechanosen-
superparamagnetic beads using magnetic twee- sors. Although desmosomes are composed of
zers (Moch et al. 2016). To investigate the trans- structurally related sets of proteins, including
mission of forces to the nucleus, which is crucial cadherin and armadillo family members, a di-
for nuclear function, a recent study examined rect homologue of the key mechanosensor in
plectin’s role in controlling nuclear morphology AJs, a-catenin is lacking. Instead, desmosomes
via keratins. On plectin knockdown, actomyo- contain the cytoskeletal linker protein DSP with
sin-dependent nuclear deformation occurred, an important role in lateral clustering of cadher-
whereas direct interactions between keratins ins and in linking KIFs to the junction. Its ami-
and the nuclear envelope were not required. In- no-terminal domain interacts with PG/JUP
stead, plectin down-regulation reduced KIF and PKPs, and is essential for DSP clustering
density in the nuclear perimeter (Almeida and determining desmosome size. This domain
et al. 2015). Possibly, plectin acts as a mechano- contains spectrin repeats homologous to the
sensor as it contains a cryptic SH3 domain in the spectrin repeats found in the related plakin fam-
fifth spectrin repeat of its plakin domain. Fol- ily protein, plectin (Choi and Weis 2011). Spec-
lowing mechanical stress, unfolding of spectrin trin repeat domains undergo only moderate
Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157 13Downloaded from http://cshperspectives.cshlp.org/ on January 22, 2022 - Published by Cold Spring Harbor Laboratory Press
M. Hatzfeld et al.
bending under mechanical stress, but repeats conserved in epithelial cells and between PKP
were shown to unfold individually at low pull- isoforms because the majority of published YAP
ing forces (Rief et al. 1999). Thus, it is possible targets from mouse keratinocytes (Liu et al.
that the DSP-amino-terminal spectrin repeat 2016) appeared down-regulated in PKP1 KO
domain undergoes force-induced unfolding of keratinocytes (our own unpublished results).
individual spectrin repeats leading to confor- Taken together, these data indicate that desmo-
mational changes that might alter DSP– protein somes might regulate YAP/TAZ signaling. Be-
interactions. In such a scenario, DSP would act cause YAP and PG/JUP were coimmunopreci-
directly as a mechanosensor. Interestingly, sev- pitated from human heart protein extracts,
eral disease-causing mutations cluster in this desmosomes could function along with a-cat-
region and are potentially linked to structural enin to recruit YAP to cell–cell contacts and
destabilization (Ortega et al. 2016). A central, limit its transcriptional activity.
a-helical domain of DSP forms coiled coils
leading to homodimerization. In contrast to
the spectrin repeat domain, coiled-coils bend ACKNOWLEDGMENTS
easily but are rather resistant to stretching (Ada-
movic et al. 2008). The carboxy-terminal do- Deutsche Forschungsgemeinschaft (DFG) fund-
main of DSP interacts with keratins and consists ing to the Hatzfeld and Magin laboratories is
of three plakin repeat domains. Whereas the gratefully acknowledged (Ha1791/8-1, Ha1791/
first plakin repeat domains are linked by just 10-1, MA1316-15, MA1316-17, MA1316-19,
four amino acids, plakin repeat domains 2 and MA1316-21, INST 268/230-1).
3 are separated by a 154 amino acid linker (Kang
et al. 2016). Decoupling of the plakin domains
suggests that the hinge region might be extend- REFERENCES
ed and strained in an initial step, before the
Reference is also in this collection.
amino-terminal spectrin repeats undergo
force-induced conformational changes (Ortega Acehan D, Petzold C, Gumper I, Sabatini DD, Muller EJ,
Cowin P, Stokes DL. 2008. Plakoglobin is required for
et al. 2016). Therefore, the plakin domain may effective intermediate filament anchorage to desmo-
work as a molecular shock absorber that dissi- somes. J Invest Dermatol 128: 2665–2675.
pates elastic energy when cells are subjected to Adamovic I, Mijailovich SM, Karplus M. 2008. The elastic
properties of the structurally characterized myosin II S2
external forces (Ortega et al. 2016). subdomain: A molecular dynamics and normal mode
analysis. Biophys J 94: 3779– 3789.
Albrecht LV, Zhang L, Shabanowitz J, Purevjav E, Towbin JA,
Desmosomes in Mechanotransduction Hunt DF, Green KJ. 2015. GSK3- and PRMT-1-depen-
dent modifications of desmoplakin control desmopla-
As mentioned before, the Hippo mediators kin-cytoskeleton dynamics. J Cell Biol 208: 597–612.
YAP/TAZ may be involved in regulating desmo- Al-Jassar C, Bikker H, Overduin M, Chidgey M. 2013.
somes via the transcription factor Tead4 (Liu et Mechanistic basis of desmosome-targeted diseases. J
Mol Biol 425: 4006– 4022.
al. 2016). Recently, desmosome mutations in
Almeida FV, Walko G, McMillan JR, McGrath JA, Wiche G,
AC were linked to modulation of Hippo/YAP Barber AH, Connelly JT. 2015. The cytolinker plectin
signaling, suggesting that there might be a feed- regulates nuclear mechanotransduction in keratinocytes.
back/feedforward mechanism (Chen et al. J Cell Sci 128: 4475– 4486.
2014). In AC, levels of phospho-YAP were in- Alvarado DM, Coulombe PA. 2014. Directed expression of a
chimeric type II keratin partially rescues keratin 5-null
creased compared with normal human hearts, mice. J Biol Chem 289: 19435– 19447.
and phospho-YAP was recruited to the junc- Amagai M. 2010. Autoimmune and infectious skin diseases
tions. In agreement, RNA sequencing, quanti- that target desmogleins. Proc Jpn Acad Ser B Phys Biol Sci
86: 524– 537.
tative polymerase chain reaction, and reporter
Amagai M, Stanley JR. 2012. Desmoglein as a target in skin
assays all showed suppressed TEAD activity in disease and beyond. J Invest Dermatol 132: 776– 784.
HL-1 myocytes with a PKP2 knockdown (Chen Bar J, Kumar V, Roth W, Schwarz N, Richter M, Leube RE,
et al. 2014). We suppose that this mechanism is Magin TM. 2014. Skin fragility and impaired desmo-
14 Advanced Online Article. Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a029157You can also read

















































