Victorian Naturalist The - Volume 139 (1) - Pronto Marketing
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
The
Victorian
Naturalist
Volume 139 (1) February 2022
Published by The Field Naturalists Club of Victoria since 1884
The Victorian Naturalist
is published six times per year by the
Field Naturalists Club of Victoria Inc
Registered Office: FNCV, 1 Gardenia Street, Blackburn, Victoria 3130, Australia.
Postal Address: FNCV, PO Box 13, Blackburn, Victoria 3130, Australia.
Phone +61 (03) 9877 9860; email: admin@fncv.org.au
www.fncv.org.au
Patron: The Governor of Victoria, the Honorable Linda Dessau AC
Address correspondence to:
The Editors, The Victorian Naturalist, PO Box 13, Blackburn, Victoria 3130, Australia.
Phone: (03) 9877 9860. Email: vicnat@fncv.org.au
The opinions expressed in papers and book reviews published in The Victorian Naturalist are those
of the authors and do not necessarily represent the views of the FNCV. Copyright of all original
material published in The Victorian Naturalist remains with the author.
Yearly Subscription Rates – The Field Naturalists Club of Victoria Inc
(effective from 1 July 2019)
Membership category
Single $ 86 Institutional
Family $111 - within Australia $172
Single Country/Concession $ 65 - overseas AUD186
Family Country/Concession $ 86
*Junior Family $ 52
Junior additional $ 17 Schools/Clubs $99
Student $ 39
* Junior membership is entitled to receive Junior Naturalist only.
Non-member fees: Meetings $3; Excursions $5
All subscription enquiries should be sent to:
FNCV, PO Box 13, Blackburn, Victoria 3130, Australia
Phone 61 3 9877 9860. Email: admin@fncv.org.au
The
Victorian
Naturalist
Volume 139 (1) 2022 February
Editors: Gary Presland, Maria Gibson, Sue Forster
Editorial Assistant: Virgil Hubregtse
Contributions Sassafras Creek Nature Conservation Reserve in the
Dandenong Ranges functions as a wildlife corridor for regionally
significant fauna, by Jasmine Andrews and Alex Maisey......................................4
The impact of Yellow-tailed Black-Cockatoo Calyptorhynchus funereus
(Shaw, 1794) feeding on a Monterey Pine Pinus radiata D.Don.,
by Gregory Moore....................................................................................................13
A case of unsupervised wildlife tourism involving kangaroos on a public
hospital estate, by Matthew Mo.............................................................................21
Book Review Field Guide to the Frogs of Australia, by Michael Tyler and
Frank Knight, reviewed by Nick Clemann ...........................................................31
ISSN 0042–5184
Front cover: Yellow-tailed Black Cockatoo Calyptorhynchus funereus. Photo Sue Forster.
Contribution
Sassafras Creek Nature Conservation Reserve in the Dandenong
Ranges functions as a wildlife corridor for regionally significant fauna
Jasmine Andrews1,3* and Alex Maisey1,2,3
1
Friends of Sassafras Creek, PO Box 20, Sassafras, Victoria 3787;
2
Research Centre for Future Landscapes, La Trobe University, Bundoora, Victoria 3086;
3
Sherbrooke Lyrebird Survey Group, 30 Moores Road, Monbulk, Victoria 3793.
*
Corresponding author: .
Abstract
The Sassafras Creek Nature Conservation Reserve forms a linear wildlife corridor in the Dandenong Ranges,
representing an important link between the Sherbrooke and Olinda units of the Dandenong Ranges National
Park, while on a broad scale linking with the Yarra Ranges via Woori Yallock Creek and the upper reaches
of the Yarra River. This study presents findings from a fauna survey undertaken in the reserve with motion-
sensing cameras, in which regionally significant fauna such as the Mountain Brushtail Possum, Superb Lyre-
bird and Long-nosed Bandicoot were detected in the corridor, providing evidence that the area is functional
for regionally significant fauna. The sites surveyed included an urban environment, typical of land adjacent to
the reserve; revegetated areas restored by local environmental groups; and largely unmodified remnant Cool
Temperate Rainforest with few or no environmental weeds present. Site-level contrasts between these habitat
types are explored (The Victorian Naturalist 139(1), 2022, 4–13)
Keywords: wildlife corridor, peri-urban wildlife, fauna survey, habitat restoration
Introduction
Wildlife corridors are areas of linear vegetation passageway; for example, a corridor that con-
connecting larger patches of wildlife habitat nects gullies and ridges is likely to be used by a
that facilitate the movement of biota through greater suite of species (Lindenmayer and Nix
an unsuitable or inhospitable surrounding 1993). Habitat quality and the degree of distur-
‘matrix’. These features of a landscape may bance from introduced species and weeds may
serve a variety of purposes, including wildlife impair the effectiveness of a wildlife corridor.
conservation, recreation and amenity and the Furthermore, a corridor consisting of remnant
provision of ecosystem services, thus confer- vegetation is likely to be of heightened value to
ring environmental and social value upon a wildlife when located in urban areas, especially
landscape (Bennett 1998). Existing protected where natural features (e.g., large old trees, fall-
areas of habitat may cause the confinement of en timber) remain.
biotic communities and result in negative im- In urban landscapes, riparian zones (veg-
pacts on genetic fitness (Haddad et al. 2003). etated land alongside a body of water) often
To solve this issue, biota need to be able to mi- represent natural wildlife corridors that have
grate through a connected system (Dilkina et remained undeveloped and, therefore, contain
al. 2017). Thus, maintaining wildlife corridors relatively natural vegetation. Riparian veg-
is becoming increasingly recognised as a con- etation is a key element along creek corridors,
servation strategy to enhance the persistence providing a range of ecosystem services, such
of fauna populations (Chetkiewicz et al. 2006). as biofiltration, flood mitigation and erosion
Wildlife corridors may serve as permanent liv- control (Cole et al. 2020). Riparian buffer strips
ing space for some species, while providing a help to maintain a healthy stream and form a
pathway for movement through degraded and protective barrier to surrounding human activ-
often urbanised or agricultural environments ity, which may in turn create a healthy ecosys-
for others. There are numerous aspects that tem (Saunders et al. 2002).
may enhance the function of a corridor. The In the ‘peri-urban’ landscape of the Dande-
landscape context of a corridor has been found nong Ranges, on the fringe of Melbourne, there
to determine the number of species using the is little supporting research concerning whether
4 The Victorian Naturalist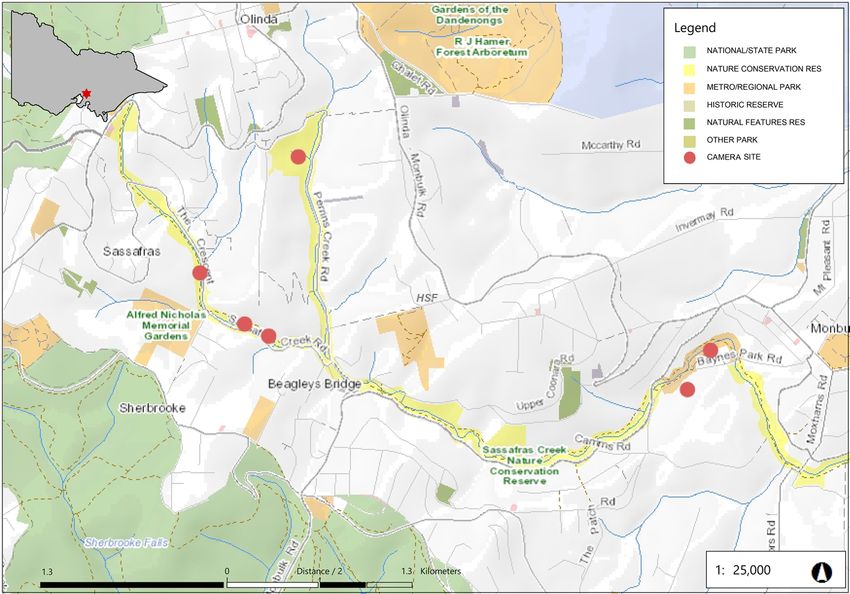
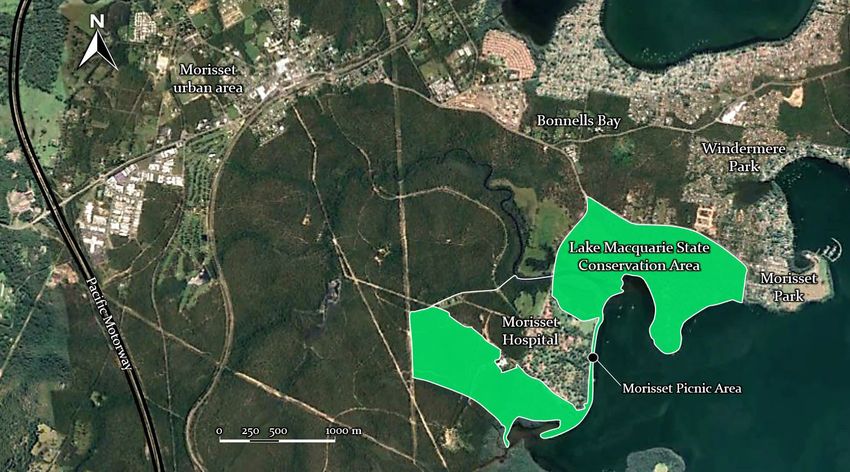
Contribution vegetated areas in riparian corridors are an ef- The community-based volunteer group Friends fective means of supporting wildlife popula- of Sassafras Creek (FOSC) actively works to tions. Sassafras Creek Nature Conservation preserve, protect and improve biodiversity val- Reserve (SCNCR) is a linear reserve situated ues in SCNCR. FOSC has three key aims: 1) on the south-eastern slopes of the Dandenong to work to improve biodiversity and habitat in Ranges (Fig. 1). The reserve area measures 46 the reserve; 2) to educate the community about ha in total, with 27 ha along the Sassafras Creek the local environment; and 3) to form strong Corridor. Sassafras Creek flows from the Sas- working relationships with relevant manage- safras township through Kallista, becoming the ment agencies. Members of the group gather Woori-Yallock Creek south of Monbulk. Veg- for monthly working bees, with a smaller sub- etation along the corridor comprises Wet For- group meeting fortnightly to complete weed- est and Cool Temperate Rainforest in its upper ing and revegetation work. The group is highly reaches, with Damp Forest becoming dominant engaged with the environmental community, downstream from Monbulk (Department of maintaining strong relationships with other lo- Land, Water and Planning [DELWP] 2020b). cal volunteer groups along the creek corridor. In the 1850s, logging occurred throughout the This network facilitates coordinated conserva- townships of Sassafras, Kallista and Olinda, tion actions across the landscape. FOSC works with the harvested timber sent to Melbourne in a systematic way to control weeds and reveg- for construction. Timber harvesting contin- etate areas in the corridor; however, further in- ued in some areas until the 1960s, representing formation is required on the effectiveness of this a period of significant landscape change that work in fostering biodiversity along the corridor. greatly modified the natural landscape (Friends This survey aimed to 1) provide a current of Sherbrooke Forest 2000). camera-trapping fauna survey to record elusive Fig. 1. Map of the Sassafras Creek Nature Conservation Reserve in the Dandenong Ranges. The red dots signify the camera sites. Vol 139 (1) 2022 5

Contribution
and rare species that may still be present in classes relating to the origin of the present veg-
the Dandenong Ranges, and 2) compare the etation: 1) original, represented by areas that
relative activity levels of fauna across areas of comprised remnant vegetation with sparce or
varying habitat quality within and directly absent weed cover (Fig. 2a); 2) revegetated, rep-
adjoining SCNCR. It was expected that sites resented by areas that have been degraded but
within areas of undisturbed habitat should have are now revegetated with native species (Fig.
more recorded native species than those situ- 2b); 3) modified, represented by an environ-
ated within recently revegetated sites and those ment that had been cleared of native vegetation
dominated by environmental weeds. The find- and is presently dominated by exotic species
ings were assessed to gauge whether the creek (Fig. 2c). Vegetation origin was expected to be
corridor is functional as habitat or for movement the most influential driver of faunal activity in
of regionally significant fauna that are likely to comparisons between sites (see below).
be detected by motion-sensing cameras. All footage was captured using the Bushnell
TrophyCam HD (model #119876), inserted
Methods with a 32 GB SD card. Each camera was set to
To undertake this study, six sites were selected record a 60s video clip with a 0.6s re-trigger
for deployment of motion-sensing trail cameras time. Camera sensitivity was programmed to
in the upper reaches of Sassafras Creek (Fig. 1). ‘high’, LED flash intensity at ‘low’, and an image
Overall, the sites were positioned to reflect a gra- resolution of 1920 × 1080 pixels was specified.
dient of habitat quality, from highly urbanised All videos were set to display the time, date
through to weed-free remnant vegetation (Ta- stamp and temperature. The SD cards and
ble 1). The six sites were classified into three
Table 1. Description of the selected study sites including the number of days the camera traps were active and
the vegetation types, including ecological vegetation class (EVC) (DELWP 2020b).
Site description Date surveyed Origin Vegetation type
Site A: An urbanised setting with introduced 28 February Modified Urban
grasses, pines and shrubby cover. Little native –10 May 2020
vegetation.
Site B: Damp fertile soil adjacent to the Sassafras 28 February Original Wet Forest (EVC 30)
Creek. Well-shaded with abundant Soft Tree –10 May 2020
Ferns Dicksonia antarctica comprising the
midstorey.
Site C: A Wet Forest site with a canopy 28 February Revegetation Wet Forest (EVC 30)
of Mountain Ash Eucalyptus regnans and the –10 May 2020
ground cover dominated by Wandering
tradescantia Tradescantia fluminensis.
Revegetation of understorey.
Site D: A revegetated site originally dominated 28 February Revegetation Wet Forest (EVC 30)
by Sycamore Maples Acer pseudoplatanus. The –25 July 2020
site still contains introduced species that
comprise the canopy and midstorey. The ground
cover and understorey primarily comprised
planted native vegetation.
Site E: Moist damp soils adjacent to the creek. 28 February Original Cool Temperate
Southern Sassafras Atherosperma moschatum –25 July 2020 Rainforest (EVC 31)
and fern-dominated ground layer. Few weeds
present.
Site F: A low-light, tree-fern dominated area 28 February Original Cool Temperate
with damp soils adjacent to Sassafras Creek. –25 July 2020 Rainforest (EVC 31)
Few weeds present.
6 The Victorian Naturalist
Contribution Fig. 2. Exemplar images of a) original native vegetation in cool temperate rainforest; b) revegetation site in wet forest; c) modified vegetation in the urbanised site adjacent to the reserve. batteries were renewed fortnightly. Some compared with the number of days where no small adjustments were made to the locations Fox was detected. of the cameras, and the number of trap days A Chi-square test of independence was ap- was recorded for each location. The total num- plied for each contrast of interest, and stand- ber of survey days was 148, within the period ardised residuals with Bonferroni correction 28 February to 25 July 2020. At each of the were used to interpret significant associations sites, the cameras were carefully strapped to a between groups. Comparisons were graphically tree and positioned within 1 m of the ground presented as a trap rate (i.e., number of days a to capture ground-dwelling animals. Cameras species was detected/total days the camera was op- were not baited. erating at a site) between the defined categories. Counts of trigger events were compiled for each species over the duration of the survey. Results If the same species was detected at a site more A total of 292 detection events comprising 32 than once on the same date, it was recorded as a species was detected in the SCNCR between single trigger event (detection event) for analy- 28 February and 25 July 2020. This included 15 sis. This was to limit multiple recordings of the bird species, 16 mammal species and one spe- same animal. cies of crustacean (Table 2; see Fig. 3 for exem- Specific contrasts were investigated relating to plar images). Of these, 24 species were native, vegetation and faunal activity. Firstly, it was of while eight were introduced, including recog- interest to test for an association between the nised pest species such as the Fox, Fallow Deer origin of habitat (i.e., original, revegetation or Dama dama and Sambar Deer Rusa unicolor modified) and origin of species detected (i.e., (Figs 3c and 3d). Regionally significant spe- native or introduced). It was expected that na- cies included the Mountain Brushtail Possum/ tive species would be positively associated with Bobuck Trichosurus cunninghami (Fig. 3a) and the original and revegetated sites, while intro- the Long-nosed Bandicoot Perameles nasuta duced species were expected to be associated (Fig. 4). The Superb Lyrebird Menura novaehol- with the modified site. landiae was detected on eight occasions, while Secondly, activity of the introduced European the Dusky Antechinus Antechinus swainsonii Red Fox Vulpes vulpes was investigated for any and Agile Antechinus Antechinus agilis were association with the origin of vegetation at a detected on four and 10 occasions, respectively. site. Foxes are a key threat to ground-dwelling The origin (native or introduced) of spe- fauna in SCNCR and Fox control is a specific cies captured on cameras was associated with management action, ranked second to removal the different habitat types (χ2 =36.87, df = of weeds, in state Biodiversity Response Plan- 2, p

8
Table 2. The number of detection events of individual species at sites grouped by vegetation origin. Included is the total number of trap days for each vegetation
origin and the total number of trigger events for each species across vegetation groups.
Common name Scientific name Modified Original Revegetation Total
(72 trap days) (283 trap days) (220 trap days)
Contribution
Agile Antechinus Antechinus agilis 9 1 10
Australian Magpie Gymnorhina tibicen 2 2
Bare-nosed Wombat Vombatus ursinus 1 10 1 12
Bassian Thrush Zoothera lunulata 3 3
Black Rat Rattus rattus 13 1 14
Brown Goshawk Accipiter fasciatus 1 1
Brown Thornbill Acanthiza pusilla 1 1
Burrowing Crayfish Sp. Engaeus sp. 1 1
Bush Rat Rattus fuscipes 11 2 13
Cat Felis catus 2 1 4 7
Common Blackbird Turdus merula 1 16 16 33
Common Brushtail Possum Trichosurus vulpecula 1 19 20
Common Myna Sturnus tristis 6 6
Common Ringtail Possum Pseudocheirus peregrinus 2 1 8 11
Crimson Rosella Platycercus elegans 4 2 6
Dog Canis familiaris 1 1
Dusky Antechinus Antechinus swainsonii 4 4
Eastern Yellow Robin Eopsaltria australis 19 6 25
Fallow Deer Dama dama 5 1 6
Laughing Kookaburra Dacelo novaeguineae 2 1 3
Long-nosed Bandicoot Perameles nasuta 1 1
Magpie-lark Grallina cyanoleuca 1 1
Mountain Brushtail Possum/Bobuck Trichosurus cunninghami 3 5 8
Pied Currawong Strepera graculina 1 1
Red Fox Vulpes vulpes 21 16 6 43
Red Wattlebird Anthochaera carunculata 3 3
Rufous Fantail Rhipidura rufifrons 1 1
Sambar Deer Rusa unicolor 2 2
Short-beaked Echidna Tachyglossus aculeatus 1 1
Superb Lyrebird Menura novaehollandiae 1 7 8
Swamp Wallaby Wallabia bicolor 16 20 36
White-browed Scrub-wren Sericornis frontalis 6 2 8
The Victorian Naturalist
Total 59 124 109 292
Contribution
Fig. 3. A selection of species detected on camera traps in Sassafras Creek Nature Conservation Reserve;
a) Mountain Brushtail Possum/Bobuck Trichosurus cunninghami; b) Fallow Deer Dama dama; c) European
Red Fox Vulpes vulpes with a Common Ringtail Possum Pseudocheirus peregrinus in its mouth; d) Short-
beaked Echidna Tachyglossus aculeatus; e) Superb Lyrebird Menura novaehollandiae; f) Bare-nosed Wombat
Vombatus ursinus; g) Agile Antechinus Antechinus agilis; h) Pied Currawong Strepera graculina.
original vegetation. There were no native Bush
Rats Rattus fuscipes or Agile Antechinus An-
techinus agilis identified at the modified site,
possibly reflecting sensitivity to habitat modi-
fication by these two native species. Both were
detected at revegetation sites (2 and 1/220 trap
days, respectively), albeit at a lower rate com-
pared with sites comprising original vegetation
(11 and 9/283 trap days, respectively).
There was a small number of detections of
Mountain Brushtail Possum, or Bobuck, at the
Fig. 4. A still image from video clip of a Long-nosed original (3/283 trap days) and revegetation sites
Bandicoot Perameles nasuta at Site E (Cool Temper- (5/220 trap days), while the Common Brushtail
ate Rainforest). Possum Trichosurus vulpecula was most preva-
lent at the revegetation site (19/220 trap days).
for higher detection rate of native species (std.
residual = 1.55, p>0.05), while native species Discussion
showed an association with revegetation sites There were several significant species recorded
(std. residual = 3.36, p
Contribution Fig. 5. Comparison of the species origin (introduced or native), for the three categories of vegetation origins: modified, original and revegetation. Standard error is included. Fig. 6. Proportional trap days Fox presence at the six sites that are categorised by vegetation origin: modified, original and revegetation. Standard error is included. rainforest habitat. Of all locations surveyed, the proliferum that provide shelter and ample Cool Temperate Rainforest is likely to repre- food resources (e.g., fungi and insects in sent the most suitable habitat for bandicoots, moist litter). Over the 148 days that the rain- primarily due to dense ground vegetation forest camera was active, there was only one such as the Mother Shield Ferns Polystichum detection of a bandicoot. This suggests that the 10 The Victorian Naturalist
Contribution Long-nosed Bandicoot is rare in the corridor. eral road mortalities of Superb Lyrebirds have The last record of Long-nosed Bandicoot in been recorded in the last five years within the the reserve was lodged in the VBA by a FOSC SCNCR (Sherbrooke Lyrebird Survey Group member in 2016. Both Bobuck and Long-nosed unpubl. data, 2021). Dog walking is permitted Bandicoot are likely to be uncommon in the within the reserve, providing these animals are Dandenong Ranges and their population tra- leashed; however, it is common to see walkers jectory in the region is poorly understood. with dogs off-lead (JA, pers. obs. 2021), potential- The records of significant fauna within reveg- ly affecting wildlife behaviour through their direct etation sites suggest there may be positive out- disturbance and scent trails (Lenth et al. 2008). comes stemming from local conservation ef- The removal of ground-smothering weeds forts. The Superb Lyrebird was also recorded and introduced trees that dominate the canopy at revegetation sites. This ground-foraging (e.g., Sycamore Maple Acer pseudoplatnus) is bird has been recognised as an ‘ecosystem en- essential to restoring ecological function in this gineer’ due to its foraging activity (Maisey et peri-urban environment. Because much of the al. 2020), an important process for ecosystem effective habitat of the reserve crosses into pri- function. Therefore, restoration work in the vate properties, landowners play an important reserve considers the habitat requirements for role in maintaining the health of this corridor. lyrebirds (e.g., revegetation with rainforest veg- Collaboration between private landowners, etation to form dense middle-storey). Sightings government stakeholders and environmental of colour-marked individuals from Sherbrooke community groups is essential, as without man- Forest confirm that lyrebirds utilise the reserve agement of invasive species on private property, as a corridor to access the Olinda unit of the the reserve is at risk of ongoing weed incursion. Dandenong Ranges National Park (Sherbrooke Major environmental weeds such as English Ivy Lyrebird Survey Group, unpubl. data 2021). Hedera helix, Sycamore Maple and Wandering Moreover, two active nests were detected in Tradescantia Tradescantia fluminensis smother the reserve while undertaking this survey, native ground vegetation and create a barrier suggesting the corridor also provides suitable between animals and their litter-dwelling prey, nesting sites for lyrebirds. Both were success- likely posing a significant threat to ground- ful to fledging and one has subsequently been foraging fauna detected during this survey. sighted on private property during 2021 (AM, Such weeds are best controlled with a ‘tenure pers. obs. 18 February 2021). Furthermore, the blind’ approach across public and private land. Superb Lyrebird was recorded in revegetated This study has documented regionally signifi- habitat in areas that had previously undergone cant species foraging in areas where introduced treatment of ground-smothering weeds that weeds have been removed, followed by subse- are known to impede foraging. Overall, the quent replanting of native vegetation. presence of the lyrebird within these sites sug- Over the course of the study, eight introduced gests that the weed control and revegetation in species were detected. Unsurprisingly, modi- the corridor is helping to support permanent fied and urbanised environments were found habitat and movement of regionally significant to have the greatest detection rates of intro- fauna throughout the landscape. duced species. The European Red Fox poses While the presence of the Bobuck, Long- serious threat to native species, highlighted by nosed Bandicoot and Superb Lyrebird are en- the image captured of a Fox with a Common couraging indicators of a functional ecosys- Ringtail Possum in its mouth (Fig. 3c). Man- tem, these significant species were detected at agement of this pest animal is likely to have low rates. This may signify that there are many a strong influence over future conservation challenges facing these populations in a peri- of fauna. A Fox control program coordinated urban setting. Fragmented habitat intersected by Parks Victoria throughout the Dandenong by roads and housing adjacent to the corridor Ranges National Park has been credited with present risks to wildlife, while domestic pets the successful regional recovery of the Lyrebird on private properties are also likely to threaten population (Sherbrooke Lyrebird Survey Group these species if they come into contact. Sev- unpubl. data, 2020). While Fox control within Vol 139 (1) 2022 11
Contribution
the SCNCR would likely provide great benefit Acknowledgments
to native species, such programs have not been Camera trapping was undertaken under research
permit 10009650 issued by the Department of En-
possible with current Fox control methods. vironment, Land, Water and Planning in collabora-
However, spill-over effects from the national tion with the Sherbrooke Lyrebird Survey Group.
park baiting program are likely to confer ben- We would like to acknowledge the members of the
efits to native species in the corridors. Friends of Sassafras Creek for their continued ef-
forts to improve the habitat in SCNCR. Many thanks
Two species of deer were also detected within in particular to Sally Bewsher, Jane Hollands and
the reserve. Their presence has many detrimen- Meghan Lindsay for assistance in the field. Finally, we
tal impacts on habitat, including erosion of thank Natalie Andrews for supporting JA throughout
creek banks, caused by their hard hooves; sedi- this study, dedicating much time to help complete
field work.
mentation and increased turbidity in streams
caused by wallows; and mortality of native References
vegetation due to over-browsing (Davis et al. Bennett AF (1998) Linkages in the landscape: the role
of corridors and connectivity in wildlife conservation.
2016). For revegetation efforts to be successful, (IUCN: Gland, Switzerland, and Cambridge, UK).
protection of plants from deer must be consid- Chetkiewicz CLB, St. Clair CC and Boyce MS (2006) Cor-
ered and implemented where possible. An ef- ridors for conservation: integrating pattern and process.
Annual Review of Ecology, Evolution, and Systematics 37,
fective state-wide deer control strategy will be 317–342.
a critical step in maintaining healthy habitats Cole LJ, Stockan J and Helliwell R (2020) Managing riparian
buffer strips to optimise ecosystem services: A review. Ag-
into the future and must be made a priority. riculture, Ecosystems and Environment 296, 106891.
Long-term monitoring projects are vital for Davis NE, Bennett A, Forsyth DM, Bowman DM, Lefroy
EC, Wood SW, Woolnough AP, West P, Hampton JO and
evidence-based conservation action. To meas- Johnson CN (2016) A systematic review of the impacts and
ure the effectiveness of management strategies management of introduced deer (family Cervidae) in Aus-
aiming to restore wildlife populations, surveys tralia. Wildlife Research 43, 515–532.
Department of Land, Water and Planning (DELWP) (2020a)
need to be conducted over prolonged periods. Biodiversity Response Planning Focus Landscapes: Upper
While this study lacks replication in survey Yarra Valley Focus Landscape. [accessed 20 March 2021].
regionally significant species such as the Long- Department of Land, Water and Planning (DELWP) (2020b)
NatureKit. (Victorian Government: Melbourne). [ac-
Over the long-term, this monitoring data may cessed 20 March 2021].
provide a baseline to inform conservation deci- Dilkina B, Houtman R, Gomes CP, Montgomery CA, McK-
elvey KS, Kendall K, Graves TA, Bernstein R and Schwartz
sions and influence the funding prospects for MK (2017) Trade‐offs and efficiencies in optimal budget‐
restoration management, especially when spe- constrained multispecies corridor networks. Conservation
Biology 31, 192–202.
cies of conservation interest are detected (Lin- Friends of Sherbrooke Forest (2000) Sherbrooke Forest: Its
denmayer and Likens 2010). Flora and History. (Friends of Sherbrooke Forest: Belgrave).
Haddad NM, Bowne DR, Cunningham A, Danielson BJ,
This study provides new records of regionally Levey DJ, Sargent S and Spira T (2003) Corridor use by
significant species in the Dandenong Ranges, diverse taxa. Ecology 84, 609–615.
such as Long-nosed Bandicoot and Bobuck Incoll B, Maisey A, and Adam J (2018) Ten years of forest res-
toration in the Upwey Corridor, Dandenong Ranges, Vic-
in both areas of remnant rainforest and areas toria. Ecological Management and Restoration 19, 189–197.
cleared of weeds and revegetated with native Lenth BE, Knight RL and Brennan ME (2008) The effects of
dogs on wildlife communities. Natural Areas Journal 28,
species. The findings paint a positive picture of 218–227.
conservation action by local community groups Lindenmayer DB and Nix HA (1993) Ecological principles
for the design of wildlife corridors. Conservation Biology
and suggest the reserve is functional as both a 7, 627–631.
wildlife corridor as well as permanent habitat Lindenmayer DB, Welsh A, Donnelly CF and Meggs RA
for many species, with potential to support (1996) Use of nest trees by the mountain brushtail pos-
sum (Trichosurus caninus) (Phalangeridae: Marsupialia). 1.
wildlife to a greater degree in future if further Number of occupied trees and frequency of tree use. Wild-
weed and pest animal management strategies life Research 23, 343–361.
Lindenmayer DB and Likens GE (2010) The science and ap-
continue to be implemented and strengthened plication of ecological monitoring. Biological Conservation
throughout the Dandenong Ranges. 143, 1317–1328.
12 The Victorian NaturalistContribution
Maisey AC, Haslem A, Leonard SWJ and Bennett AF (2020) Saunders DL, Meeuwig JJ, and Vincent AC (2002) Freshwater
Foraging by an avian ecosystem engineer extensively mod- protected areas: strategies for conservation. Conservation Bi-
ifies the litter and soil layer in forest ecosystems. Ecological ology 16, 30–41.
Applications 31. Victorian Biodiversity Atlas (VBA) (2020) Victorian Biodi-
Martin JK and Handasyde KA (2007) Comparison of bobuck versity Atlas. [accessed
(Trichosurus cunninghami) demography in two habitat 27 September 2020].
types in the Strathbogie Ranges, Australia. Journal of
Zoology 271, 375–385. Received 22 April 2021; accepted 26 June 2021
The impact of Yellow-tailed Black-Cockatoo Calyptorhynchus
funereus (Shaw, 1794) feeding on a Monterey Pine Pinus radiata D.Don.
Gregory Moore
The University of Melbourne, Burnley College, 500 Yarra Boulevard, Richmond, Victoria 3121.
Correspondence: .
Abstract
The Yellow-tailed Black-Cockatoo Calyptorhynchus funereus consumes seeds and wood-boring insect larvae
from woody-fruited native species such as Hakea, Casuarina, Allocasuarina, Eucalyptus and Banksia and from
Acacia as well as seeds from the cones of introduced pine species. While its feeding on pine seeds is well known,
there have been few studies describing the details of this feeding behaviour or its impact on the trees.
An opportunity to collect such data arose in 2020 when a single Monterey (Radiata) Pine Pinus radiata
was visited on three occasions by small flocks of Yellow-tailed Black-Cockatoos. The behaviour of these birds
during feeding sessions was observed over three days and the fallen debris of small and large needle-bearing
shoots and cones was collected, sorted and categorised. Large and short needle-bearing shoots were counted
and weighed. Eight birds visited the tree on the first day, two fed on the second day and four on the final day
of feeding. The Cockatoos harvested 721 large and short needle-bearing shoots with a total mass of 19 552 g
and 392 cones weighing nearly 17 kg. Only 134 cones (34.18%) with damage greater than 51% had all seeds
removed. (The Victorian Naturalist 139(1), 2022, 13–20)
Keywords: harvesting of seeds from pine cones, feeding session, Black-Cockatoos, defoliation
Introduction
The Yellow-tailed Black-Cockatoo Calypto- There has been interest in the feeding and
rhynchus funereus (Shaw, 1794) is unusual damage done to plantation forestry species such
among cockatoos in that it consumes seeds as Monterey Pine and Flooded Gum Eucalyptus
and large numbers of wood-boring insect lar- grandis W.Hill by Australian birds, including
vae (McInnes and Carne 1978; Higgins 1999). the Yellow-tailed Black-Cockatoo (Graham
It is Australia’s longest Black-cockatoo, reach- 1928; Pawsey 1966; Gepp 1976; McInnes and
ing 55–65 cm in length (Higgins 1999; Forshaw Carne 1978; Way and van Weenen 2008). The
2002; Harris 2007; Way and van Weenen 2008) Cockatoos peel off large strips of bark and then
and is known to feed on the seeds of both native excavate wood to extract the larvae from the
and exotic trees. The seeds of, and wood-boring heartwood, not only damaging the timber, but
larvae in, Acacia, Hakea, Casuarina, Allocasu- causing trees to snap in storms, with up to 40 %
arina, Eucalyptus and Banksia are consumed, of trees in some plantations being lost due to
but the species also feeds on seeds in cones of Cockatoo damage (McInnes and Carne 1978).
exotic species such as Aleppo Pine Pinus ha- Similar damage has been reported for native
lepensis Mill, Monterey (Radiata) Pine Pinus trees and shrubs, such as eucalypts, wattles and
radiata D.Don. and Bhutan Cypress Cupressus hakeas, by birds excavating the heartwood for
torulosa D.Don. (Higgins 1999; Forshaw 2002; larvae (Way and van Weenen 2008).
Way and van Weenen 2008).
Vol 139 (1) 2022 13Contribution Yellow-tailed Black-Cockatoos are known to by a cacophony of shrieks mixed with the calls revisit feeding sites, not only day after day but of the Ravens. The Ravens soon depart, leaving also year after year (Noske 1980; Way and van the Cockatoos to feed in peace; however, they Weenen 2008). Cockatoo flock sizes vary with usually return within 10 minutes of the Cocka- species, seasonally and with the availability toos’ departure, so they must be keeping watch of food (Higgins 1999; Cameron 2005). Over over the tree. the past 10–15 years, small flocks of up to 30 Yellow-tailed Black-Cockatoos, while not as birds have been observed moving along the loud or raucous as the Sulphur-crested Cocka- Maribyrnong River banks, west of Melbourne, too Cacatua galerita (Latham, 1790), make often feeding from wattle and eucalypts, and their presence known with a series of calls ultimately arriving at Brimbank Park, a large and whistles (Higgins 1999). They are diur- metropolitan park in Melbourne’s west. They nal and noisy, and, with the loud responses of would then spend a number of days (often 10– the Ravens, their arrival at the tree would be 28 days) feeding within the park, but also ven- heard during the Covid-19 lockdown period turing from the park to feed from pine species in Victoria, when people were restricted to in the nearby village of Keilor and on individual their homes. It would provide an opportunity Monterey Pines growing in suburban Kealba. to collect data on the removal of cones and There is a single large Monterey Pine growing needle-bearing shoots. in a vacant suburban block in Kealba, adjacent to Brimbank Park. In the past, this tree has been Methods visited regularly by Yellow-tailed Black-Cocka- Yellow-tailed Black-Cockatoos arrived at the toos. When the birds were observed feeding in tree in early October 2020. Their feeding ses- Brimbank Park and, later, in Keilor, plans were sions were timed from the beginning until the made to monitor their behaviour should they last bird departed the tree. Birds of a flock ar- visit the Kealba tree. rived within seconds of each other and stopped The cones of Monterey Pine are serotinous feeding within seconds of each other. The and often require a trigger such as heat or fire numbers of birds present at each feeding ses- for the release of seeds. The scales are capable sion were counted. Despite the birds being dif- of remaining closed for several years, so a suc- ficult to see in the canopy, small flock sizes of cession of cones can result in cumulative seed two to eight birds and lengthy feeding sessions crops being stored in the canopy for years made accurate counts possible along with ob- (Centre for Agriculture and Bioscience In- servations of different aspects of their feeding ternational [CABI] 2020). The scales can also behaviour. open and then close again depending on envi- All small and large needle-bearing shoots ronmental conditions, releasing small numbers and cones dropped by the birds during feed- of viable seeds over a couple of decades. Old ing sessions were collected separately after the cones can remain attached to a tree for many final feeding session. Prior to the arrival of the years after they have opened and all seed within flocks, the ground around the tree was tidied has been shed (Cope 1993). Monterey Pine is so that freshly fallen material could be readily monoecious, with the small male cones occur- identified. The fine debris on the ground fol- ring lower on the tree than the female cones, lowing feeding sessions was not collected as it and only the female cones are retained (Walden was too small and scattered more widely. The et al. 1999). site was inspected daily for ten days after the The Monterey Pine in the vacant block at departure of the Yellow-tailed Black-Cockatoos Kealba is a large, forty-year-old tree that is usu- in order to collect any cones or needle-bearing ally the domain of a family of Little Ravens Cor- shoots that had been snagged in the tree and vus mellori Matthews, 1912, which have both fallen to the ground later. roosted and perched there at various times The number and weight of the cones that during the day, but have never nested in it. had been removed were determined. Cones The Ravens are no match for the Yellow-tailed were categorised according to the degree of Black-Cockatoos, whose arrival is announced discernible damage done using a seven-point 14 The Victorian Naturalist
Contribution
Table 1. Feeding hours of Yellow-tailed Black-Cockatoos feeding on a Monterey Pine.
Feeding session Number of birds Duration (hours) Feeding hours
1 8 1.5 12.0
2 2 1.25 2.5
3 4 1.0 4.0
Total feeding hours 18.5
Table 2. Number and mass of needle-bearing shoots removed from Monterey Pine by Yellow-tailed Black-Cockatoos.
Number Weight (g) Average weight (g) Number of cones
on shoots
Small shoots (100g) 16 1755 109.7 0
Large shoots with undamaged cones 37 7629 206.2 70
TOTAL 721 19 552 70
scale: 0 (no damage); + (evidence of minor Yellow-tailed Black-Cockatoos fed on the Mon-
damage), 1(71% damage). rived at the tree on 1 October 2020; they re-
Needle-bearing shoots were separated into three turned on the next two days. On the first day,
groups: small, large, and those with undamaged when eight birds were present, one bird kept up
cones. For each group, the shoots were counted a low almost croaking/growl call for the whole
and weighed to determine the mass of material time that the birds were there and was probably
removed. After the final feeding session, the a juvenile bird. There was no such calling when
tree was scanned with Tasco 10 × 25 binoculars the smaller numbers of birds were feeding on
to determine whether any cones of the same days 2 and 3; these latter sessions occurred in
comparative size and age of those removed had near silence except for the occasional low call
been left on the tree. Scanning was done from and the cracking of woody shoots and cones
the ground, but since most of the canopy of the and the thuds as cones landed. In every case
tree could be seen, a reasonably accurate count that could clearly be observed, the Cockatoos
could be made. used their left foot to manipulate the cones
To allow quantification of feeding sessions while feeding. The beak was used to dismantle
that involved different flock numbers and dif- the cones and to sever shoots and cones from
ferent durations, a concept of bird-feeding- the tree.
hours was utilised, where the number of feed- Over these three days, the Cockatoos re-
ing birds was multiplied by the feeding time moved 668 small shoots from the tree (Table 2),
in hours. This enabled the rates of removal of ranging in weight from 1 to 100 g but averag-
pine cones and large and small needle-bearing ing 15 g. Fifty-three large needle-bearing shoots
shoots and their respective masses to be cal- were removed (Table 2), each weighing more
culated. The average rates of removal per bird than 100 g; the largest weighed 898 g. Some of
were also estimated. these larger shoots were over 600 mm in length
(Fig. 1). The total weight of large shoots (9384
Results g) was less than that of small shoots (10 168 g)
Three feeding sessions were observed (Table 1). (Table 2). Thirty-seven of the larger shoots also
Eight birds fed for 1.5 hours in the first session, had untouched, generally small cones attached
two birds fed for 1.25 hours in the second ses- (Table 2). No damage was done to the tree or
sion and four birds fed for 1.0 hour in the final detached shoots by birds in search of larvae.
session. Using the feeding hour concept, the
Vol 139 (1) 2022 15Contribution
Fig. 1. Examples of small shoots (left) and large shoots greater than 60 cm long (right) removed from Monterey
Pine by Yellow-tailed Black-Cockatoos.
Table 3. Number, mass, size and level of damage exhibited by cones removed from Monterey Pine by Yellow-
tailed Black-Cockatoos.
Level of damage Number of Closed cones Status of seed Mass of cones (g) Ave. mass of
open cones cones (g)
0 None 34 None removed 1816 53.41
0 None 6 None present 392 65.33
+ Minor 151 0 None removed 8023 53.13
1 (71%) 108 0 All removed 2426 22.46
TOTAL 358 34 16 803 42.86
The most obvious result of the Cockatoos the damage was to the smallest scales at the tip
feeding on the tree was the large number of of the cone (114 cones), to the tip and mid-
cones (392) found on the ground after the feed- dle scales (28 cones) and rarely at the base of
ing sessions (Table 3). These were cones only, cones (nine cones). On these cones, none of the
without any foliage attached. All were a light scales had been completely removed, as was
brown or tan colour except for six, which were the case for the more significantly damaged
grey and typical of older cones. It was interest- cones (Fig. 2). There were 201 dropped cones
ing to note that many of the cones opened by that showed more significant damage (>10%)
the birds had approximately the same width from Cockatoo feeding (Table 3; Fig. 3). Once
and length. damage to the cones exceeded 50%, no seed re-
Of the cones removed, 40 (10.0%) showed no mained within them (Table 3).
damage from feeding (Table 3; Fig. 2) and, of The average weight of cones declined as the
these, 34 had not opened at all (Table 3). The damage to cones increased (Table 3). This was
other six cones were older fully open cones expected as greater damage to the cone resulted
containing no seeds (Table 3). Another 151 in a greater amount of missing cone structure.
(38.5%) had very little damage and no evidence After the feeding sessions, there was a lot of
of seed removal (Table 3). At first glance these fine debris on the ground consisting of scales
appeared to have no damage, but closer inspec- and bits of scales that had been removed from
tion showed very slight damage caused by the cones, and some winged seeds (Fig. 4). Four
beaks of the Cockatoos. Usually, but not always, days after the last feeding session, pieces of
16 The Victorian NaturalistContribution Fig. 2. Examples of Monterey Pine cones with closed scales (left) and cones showing a range of damage caused by Yellow-tailed-Cockatoos (right). Fig. 3. Cones of Monterey Pine showing greater levels of damage after seed harvesting by Yellow-tailed Black- Cockatoos. The most severe damage is shown on the right and is very similar to that found in Aleppo Pine by Way and van Weenen (2008). Fig. 4. Pieces of cones and cone scales (left) and the viable winged seeds (right) left after harvesting of seeds from a Monterey Pine by Yellow-tailed Black-Cockatoos. Vol 139 (1) 2022 17
Contribution
Table 4. Calculated feeding rates on cones and needle-bearing shoots of Monterey Pine by Yellow-tailed
Black-Cockatoos.
Needle-bearing shoots Cones
Number Mass (g) Number Mass (g)
Per feeding hour 38.97 1056.86 21.19 908.27
Feeding session 1 467.64 12 682.32 254.28 10 899.24
Feeding session 2 97.43 2642.15 52.98 2270.68
Feeding session 3 155.88 4227.44 84.76 3633.08
TOTAL 721.00 19 552.00 392.00 16 803.00
cone scales could still be found, but no pine spent about 20 minutes dissecting a cone and
seeds remained. had a preference for green cones but, in this
Scanning the tree with binoculars after the study, the Cockatoos harvested an average of
feeding sessions revealed that hundreds of the 21.19 cones in an hour, so were spending an av-
old, grey, open cones remained. There were 12 erage of less than three minutes on each cone.
large (estimated as > 8 × 8 cm), green, closed- Older birds are also known to remove cones
cones still attached to needle-bearing shoots and more rapidly than younger birds (Cameron
at least 100 small cones (estimated as < 3 × 3 cm) 2005). This difference in feeding time may be
attached to shoot apices. It is likely that there explained partly by the fact that nearly half the
were more of these smaller cones hidden by fo- cones were removed with little, if any, damage.
liage that could not be seen from the ground. Only 134 cones (34.18 %) with damage greater
No light brown cones of the size or age collect- than 51 % had all seeds removed, so removal
ed in the dropped material could be seen. of scales and seeds would have taken longer.
The Yellow-tailed Black-Cockatoos spent a Cones that suffered greater than 51 % damage
total of 18.5 feeding hours at the Monterey were very similar to those illustrated for Alep-
Pine. They removed 721 foliage-bearing shoots po Pine by Way and van Weenen (2008). Green
with a mass of 19 552 g (Table 2) and 392 cones cones were left largely intact.
with a mass of 16 803 g (Table 3). Dividing In contrast to the cones with little damage, the
these totals by the feeding hours (18.5), the rate pattern of the most severe damage done usually
of removal of cones and long and short needle- saw the smaller scales toward the apex left in-
bearing shoots per hour and per feeding ses- tact (Fig. 3). The scales toward the centre of the
sions were calculated (Table 4). cones were often damaged and, in many cases,
removed from the axis of the cone altogether,
Discussion as were the scales towards the base of the cone.
The impact of a flock of Yellow-tailed Black- This pattern was consistently displayed in cones
Cockatoos feeding on a tree is significant. Even with more than 51 % damage, but was particu-
a small flock of birds can remove large numbers larly evident when damage exceeded 71 %. It
and masses of both cones and needle-bearing seems likely that no seeds occurred in these
shoots. The birds remove cones and feed on apical scales, so the birds did not waste time or
the winged seeds within, using their left foot to energy in damaging them further. This is con-
manipulate the twigs and cones and their beaks sistent with the efficient foraging strategies of
and tongues to sever the plant parts and extract other cockatoos (Clout 1989).
the seeds. All birds observed clearly used the The Cockatoos removed over 36 kg of ma-
left foot for manipulation, which is consistent terial from the tree. Removal of cones to feed
with other reports that cockatoos are predomi- on the winged seeds within is well-known,
nantly, if not exclusively, left footed (Brown but 40 (10.0%) of the cones were undamaged
2011; Brown and Magat 2011). and mostly closed, and another 151 (38.52%)
As previously reported by Dawson (1994), the had very little damage and no evidence of seed
birds systematically lifted and often removed retrieval. Given that nearly 50% of the cones
each scale. Dawson (1994) estimated that birds
18 The Victorian NaturalistContribution removed from the tree yielded no seed, this for weeks and became part of the site mulch. removal seems to be inefficient, as does the However, within four days of the last feeding removal of large shoots bearing cones that were session there were no pine seeds remaining. It all closed and undamaged by the birds. would seem that these were eaten by other birds None of the large, old, open cones on the tree and, while it is uncertain which birds consumed were touched by the Cockatoos. This was to be the seeds, up to four Spotted Doves Spilopelia expected as there would be no seeds in them chinensis, (Scopoli, 1786) were observed feeding and, so, there would be no incentive for explo- under the Monterey Pine. This ground-feeding ration by the birds. There were so few cones of could further reduce the seed bank and dimin- the type harvested by the Yellow-tailed Black- ish the tree’s weed potential. The feeding-debris Cockatoos remaining that it would not be deposited under the tree contributed substan- worth their while returning to feed from that tially to the organic matter, adding to the quan- tree for some years. The tree has been visited on tity and the particle size diversity of the litter, three occasions over the past decade. A feed- which previously consisted almost entirely of ing-visit every three years or so seems likely to pine needles. Such a change could impact the yield an appropriate number of cones of suit- biodiversity of the organisms present in the litter. able size and age to make the visit productive The removal of large and small needle-bear- for the birds. ing shoots without cones raises questions. Is The cones of the tree are known to contain this material removed with a purpose? What viable seed as it has been collected and propa- benefit do birds derive from its removal? The gated in the past. However, there have been few removal could be aimed at improving access to occurrences of self-seeding in the vicinity of and manipulation of the cones and so, may be the tree and no known occurrences in nearby considered incidental damage. It may also as- Brimbank Park. Yellow-tailed Black-Cockatoos sist birds to clean their beak of resin encoun- have been reported as spreading Monterey Pine tered during chewing of cones. The removal seeds and contributing to the distribution of of so many needle-bearing shoots has a visible the species as a weed (Gill and Williams 1996; impact on the canopy of a tree, but is still a rela- Williams and Wardle 2005). In this study, the tively small proportion of a large canopy. How- birds were not seen carrying any shoots, cones ever, it opens up the canopy and the removed or other material from the site. All material that shoots tend to bear the younger greener foliage; was dropped by the birds fell in the immediate as a result, this tree did not look as healthy as vicinity of the tree and usually immediately be- it did before the feeding sessions. While the low the canopy. overall effect is likely to be a slight reduction in Furthermore, feeding by Yellow-tailed Black- photosynthetic activity, the longer-term impli- Cockatoos significantly reduced the number of cations remain unknown and would be worthy cones and seeds held on the plant; cones with of further study. damage greater than 51 % contained no seed While this study involved only a single Mon- at all. While the harvesting does not eliminate terey Pine and small flocks of Yellow-tailed the risk that the tree might self-seed and pre- Black-Cockatoos, it revealed some interesting sent as a weed, it may significantly lessen its aspects of the birds’ behaviour. It also provided weed potential by reducing the seed bank, es- some insight into the rate of feeding and the pecially when the tree is regularly and repeat- amounts of material that Yellow-tailed Black- edly grazed. Larger flocks feeding on plantation Cockatoos can remove during a feeding ses- populations of Monterey Pine on a regular basis sion. Further research on larger numbers of could significantly reduce the seed bank held trees and bigger flocks would provide valuable within the canopy of the trees, reducing their information concerning not only damage done weed potential. The influence of Yellow-tailed to trees, but also the effects on seed banks held Black-cockatoos on the seed stored in trees in tree canopies and the reduction in potential they harvest warrants further research. weediness of harvested species. The fine debris consisting of cone scales re- mained on the ground after the feeding sessions Vol 139 (1) 2022 19
Contribution
Acknowledgements Gill AM and Williams JE (1996) Fire regimes and biodiver-
My colleague John Delpratt is sincerely thanked for sity: the effects of fragmentation of south-eastern Austral-
his encouragement to publish and his close reading ian Eucalypt forests by urbanisation, agriculture and pine
plantations. Forest Ecology and Management, 85, 261–278.
of, and many helpful suggestions for improving, the Graham G (1928) The Yellow-tailed Black Cockatoos. The
manuscript. Emu 28, 154.
Higgins P (1999) (Ed) Handbook of Australian, New Zealand
References and Antarctic Birds. Volume 4, Parrots to Dollarbird. (Ox-
Brown C and Magat M (2011) Cerebral lateralization de- ford University Press: Melbourne).
termines hand preferences in Australian parrots. Biology Harris L (2007) Husbandry manual for Yellow-tailed Black
Letters 7, 496–8. . Cockatoo, Calyptorhynchus funereus. (Western Sydney In-
Brown C (2011) News in Science, ABC Science [accessed 12 August South Wales. Australian Wildlife Research 5(1), 101–121.
2020]. Noske S (1980) Aspects of the Behaviour and Ecology of
Centre for Agriculture and Bioscience International (2020) the White Cockatoo (Cacatua galerita) and Galah (C. ro-
Invasive Species Compendium. (CAB International: Wall- seicapilla) in Croplands in North-East New South Wales.
ingford, UK). (Unpublished MSc thesis, University of New England, Ar-
[accessed 8 October 2020]. midale). .
Cameron M (2005) Group size and feeding rates of Glossy Pawsey CK (1966) Birds in relation to the pine forests of the
Black Cockatoos in central New South Wales. Emu 105, south east of South Australia. The South Australian Orni-
299–304. thologist 24, 93–95. [accessed 9 October
toos. Australian Wildlife Research 16, 467–473. 2020].
Cope AB (1993). Pinus radiata. In: Fire Effects Information Walden AR, Walter C and Gardner RC (1999) Genes ex-
System, U.S. Department of Agriculture, Forest Service, pressed in Pinus radiata male cones include homologs to
Rocky Mountain Research Station, Fire Sciences Labora- anther-specific and pathogenesis response genes. Plant
tory (Producer). [accessed 13 October 2020]. Way SL and van Weenen J (2008) Eyre Peninsula Yellow-
Dawson J (1994) The Yellow-tailed Black Cockatoo, Calypto- tailed Black Cockatoo (Calyptorhynchus funereus whitei)
rhynchus funereus funereus and Calyptorhynchus funereus Regional Recovery Plan. Department for Environment and
xanthanotus: Report on the Yellow-tailed Black Cockatoo Heritage, South Australia.
Survey, 1983–1988. (Bird Observers Club of Australia: Williams MC and Wardle GM (2005) The invasion of two
Nunawading). native Eucalypt forests by Pinus radiata in the Blue Moun-
Forshaw J (2002) Australian Parrots revised 3rd edition, (Al- tains, New South Wales, Australia. Biological Conservation
exander Editions: Brisbane). 125, 55–56.
Gepp BC (1976) Bird Species Distribution and Habitat Di-
versity in an Exotic Forest in South Australia, Australian
Forestry 39, 269–287. Received 8 December 2020; accepted 13 January 2022
Fifty-eight Years Ago
Black Cockatoos and Grass-trees
By K. G. Simpson
A paper by P. A. Gilbert (1935) records the Yellow-tailed Black Cockatoo (Calyplorhyncus funereus Shaw)
extracting beetle larvae (Cerambycidae, Coleoptera) from dead flower spikes of the Grass-tree or Black-
boy (Xanthorrhoea spp.) on the central New South Wales coastal strip (“Movements of birds, Pt. II —The
seasonal movements and migrations of birds in eastern N.S.W.”, Emu 34: 205). In June 1963, after reading
the above record, I examined a large patch of X. australis R.Br., situated on the northern slope of a hill at
Tidbinbilla, 25 miles west of Canberra. Evidence was found that extensive damage to flower spikes had oc-
curred within the last two or three years. The damage was somewhat similar in appearance to that caused
by C. funereus in extracting large moth larvae from several species of Acacia in the same district, a descrip-
tion of which is to be published shortly (Simpson, K. G., in press—Emu).
From The Victorian Naturalist 81, p. 217, December 10, 1964
20 The Victorian NaturalistContribution
A case of unsupervised wildlife tourism involving kangaroos on a
public hospital estate
Matthew Mo
Correspondence:
Abstract
There is a strong interest in wildlife tourism; however, some human-animal interactions may be unsupervised
and lack management controls. The ethics of tourists using the estate of a psychiatric hospital to interact with
(including feeding) wild Eastern Grey Kangaroos Macropus giganteus were debated. There was considerable
argument. Some considered the hospital estate a convenient location to see kangaroos habituated to close
encounters. Others objected, citing potential implications of tourist numbers on psychiatric patients and po-
tential harm to kangaroos from feeding them artificial foods, and stress caused by some tourist behaviours. The
issue gained widespread political and media attention in May 2018 following reports of tourists being injured.
This eventuated in the land manager closing the site to the general public. Tourist interaction with kangaroos is
not a new issue; however, a psychiatric hospital is an unlikely tourist attraction, making this case study unique.
(The Victorian Naturalist 139(1), 2022, 21–30)
Keywords: human-animal interactions, food provisioning, wildlife management, wildlife tourism
Introduction
Globally, there is insatiable interest in the po- knowing of locations frequented by certain ani-
tential of encountering wildlife in situ. This is mals and travelling there to interact with them
embedded in wildlife tourism (Mutanga et al. (Newsome et al. 2004; Pocock 2006). In these
2017; Newsome et al. 2019). Human-animal scenarios, it is difficult for wildlife managers
interactions in wildlife tourism range from to regulate activities or implement controls to
strictly observational activities (Cerri et al. address risks and impacts. Some examples of
2019) to experiences involving physical con- unsupervised wildlife tourism include visits to
tact with animals (e.g. Blewitt 2008; Newsome bays to feed fish (Semeniuk et al. 2009), leaving
and Rodger 2013). Wildlife tourism contrib- out carcasses to attract carnivores to particular
utes to conservation objectives by providing sites (Markwell 2015) and feeding birds and mon-
tourists with interpretive experiences that pro- keys to encourage close interactions (Robb et al.
mote public familiarity and learning about the 2008; Sengupta and Radhakrishna 2020).
animals (Ballantyne et al. 2011) or establishing Kangaroos (family Macropodidae) are some
economic alternatives to the commercial har- of the world’s most recognisable tourism spe-
vesting of some species (Garrod and Wilson cies (Higginbottom et al. 2004; Andrew 2015).
2003; Cisneros-Montemayor et al. 2013). ‘Eco- Kangaroos are entrenched in the cultural iden-
tourism’ is a concept formed in the late 20th tity of Australia, as demonstrated by their ap-
Century. It prioritises the operation of tourism pearance on the Commonwealth Coat of Arms;
in a manner that is sustainable and does not marketing emphasis by the tourism sector; nu-
damage natural values (Blumstein et al. 2017; merous kangaroo-related souvenir products;
Worrell et al. 2017). and their presence in almost every zoological
Despite the benefits of wildlife tourism to facility in Australia. Conveniently for tourists,
conservation objectives, there are many tourist some species are abundant in many rural and
activities that attract concerns about the risks urban areas of Australia (Fig. 1). Populations
and impacts on animal welfare and conserva- of Eastern Grey Kangaroos Macropus giganteus
tion, animal behaviours and ecosystem health in some parts of capital cities, such as Can-
(Brookhouse et al. 2013; Trave et al. 2017). In berra (Fletcher 2007) and Sydney (Colgan et al.
particular, some human-wildlife encounters are 2019), are a case in point. However, kangaroos
unsupervised or unstructured (see Newsome are also the focus of contentious management
and Rodger 2008), characterised by people issues (Descovich et al. 2016). In heavily developed
Vol 139 (1) 2022 21You can also read

















































