SLCO4A1-AS1 mediates pancreatic cancer development via miR-4673/KIF21B axis
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Open Medicine 2022; 17: 253–265
Research Article
Jianxin Zhang#, Yanbing Shen#, Dandan Ma*, Zhonghu Li, Zhiyong Zhang, Weidong Jin*
SLCO4A1-AS1 mediates pancreatic cancer
development via miR-4673/KIF21B axis
https://doi.org/10.1515/med-2022-0418 Keywords: SLCO4A1-AS1, miR-4673, KIF21B, pancreatic
received July 9, 2021; accepted December 7, 2021 cancer
Abstract: In this study, we intended to figure out the
biological significance of long non-coding RNAs (lncRNAs)
solute carrier organic anion transporter family member 4A1
antisense RNA 1 (SLCO4A1-AS1) in pancreatic cancer (PC). 1 Introduction
Cell counting kit-8, colony formation, wound healing, trans-
well, and flow cytometry experiments were performed to Pancreatic cancer (PC) is known as a prevailing malig-
reveal how SLCO4A1-AS1 influences PC cell proliferation, nancy in digestive system worldwide, accounting for
migration, invasion, and apoptosis. Thereafter, bioinfor- approximately 56,770 new cases and 45,750 deaths in
matics analysis, RNA immunoprecipitation assay, luciferase America in 2019 [1]. Due to the aggressive malignant
reporter assay, and RNA pull-down assay were applied for behaviors at early stage, the shortage of early diagnostic
determining the binding sites and binding capacities markers, the lack of effective therapeutic strategies, and
between SLCO4A1-AS1 and miR-4673 or kinesin family the speedy disease progression, the prognosis of patients
member 21B (KIF21B) and miR-4673. The results depicted with PC is extremely unsatisfactory [2,3]. Surgical resec-
that SLCO4A1-AS1 was upregulated in PC, and SLCO4A1- tion and systemic chemo-radiotherapy are considered as
AS1 knockdown suppressed PC cell growth, migration, the primary choices for PC treatment [4]. Despite great
invasion, and induced cell apoptosis. Furthermore, SLCO4A1- advances and innovations during the past decades [5,6],
AS1 was verified to modulate the expression of KIF21B by the underlying mechanisms of the initiation and progres-
binding with miR-4673. SLCO4A1-AS1 exerted an oncogenic sion of PC remain obscure. Therefore, it is urgent to
function in PC. The overexpression of SLCO4A1-AS1 aggra- develop a further understanding of the molecular mecha-
vated the malignant behaviors of PC via the upregulation of nisms underlying PC and identify novel effective biomarkers
KIF21B by sponging miR-4673. Our findings revealed a novel of PC.
molecular mechanism mediated by SLCO4A1-AS1, which Accumulating evidence has suggested that long non-
might play a significant role in modulating the biological coding RNAs (lncRNAs) are a type of RNA transcripts in
processes of PC. eukaryotic cells longer than 200 nucleotides defined by a
lack of protein-coding potential [7]. More and more stu-
dies have revealed the regulation of lncRNAs in a range of
# These authors contributed equally to this work. biological processes, involving cell growth, apoptosis,
migration, and invasion in almost all human cancers,
including gastric cancer, hepatocellular carcinoma, and
* Corresponding author: Dandan Ma, Department of General prostate cancer [8–11]. LncRNA solute carrier organic anion
Surgery, General Hospital of Central Theater Command, 627 Wuluo transporter family member 4A1 antisense RNA 1 (SLCO4A1-
Road, Wuchang District, Wuhan 430070, Hubei, China,
AS1) was verified to be an oncogene in some cancers [12].
e-mail: Doctor_madandan@163.com,
tel: +86-027-50772353 For example, in non-small-cell lung cancer, SLCO4A1-AS1
* Corresponding author: Weidong Jin, Department of General facilitates cellular processes by mediating the miR-223-3p/
Surgery, General Hospital of Central Theater Command, 627 Wuluo IKKα/NF-κB signaling [13]. In colorectal cancer, SLCO4A1-
Road, Wuchang District, Wuhan 430070, Hubei, China, AS1 accelerates cell growth and migration via β-catenin-
e-mail: ptwkhero@163.com,
dependent Wnt pathway [14]. SLCO4A1-AS1 binds with
tel: +86-027-50772353
Jianxin Zhang, Yanbing Shen, Zhonghu Li, Zhiyong Zhang:
miR-335-5p to elevate organic cation/carnitine transporter4
Department of General Surgery, General Hospital of Central Theater (OCT4) expression, thus aggravating malignant phenotypes
Command, Wuhan 430070, Hubei, China of bladder cancer [15]. The GEPIA database indicates that
Open Access. © 2022 Jianxin Zhang et al., published by De Gruyter. This work is licensed under the Creative Commons Attribution 4.0
International License.
254 Jianxin Zhang et al.
SLCO4A1-AS1 is upregulated in 179 pancreatic adenocarci- PCR was carried out with SYBR Green PCR Kit (Takara,
noma (PAAD) tissue samples compared with 171 non-tumor Dalian, China) on ABI 7500 Fast Real-Time PCR system
tissue samples. Nevertheless, the function and underlying (Applied Biosystems, Foster City, CA, USA) with particular
mechanism of SLCO4A1-AS1 in PC remain unclear. primers according to the recommendations of the manu-
Therefore, this study investigated the regulatory mecha- facturer. GAPDH and U6 served as the internal controls.
nism of SLCO4A1-AS1 in PC cells. Additionally, the role of The relative expression of RNAs was quantified and ana-
the SLCO4A1-AS1/miR-4673/KIF21B regulatory network in lyzed with the 2−ΔΔCt method. Primer sequences are listed
the malignant progression of PC was explored, providing a in supplementary Table 1.
better understanding of the pathogenesis of PC.
2.4 Cell counting kit-8 (CCK-8) assay
2 Materials and methods Cell viability was assessed by CCK-8 assay. Briefly, cells
were seeded into 96-well plates (1 × 104 cells/well). Each
well was added with 200 µL medium containing CCK-8
2.1 Cell culture
PC cell lines (CAPAN-1, SW1990, CFPAC-1, and Panc
03.27) and human pancreatic duct epithelial cell line
(H6C7) used in the study were purchased from American Table 1: Primer sequences used for qRT-PCR
Type Culture Collection (ATCC, Manassas, VA, USA). SW1990
and CAPAN-1 cells were incubated in Dulbecco’s Modified Gene Sequence (5′ → 3′)
Eagle’s Medium (DMEM; Gibco, NY, USA) containing 10% fetal SLCO4A1-AS1 CTGGGCTGACAAGTGAGGAG
bovine serum (FBS, Gibco), and Panc 03.27, CFPAC-1, and SLCO4A1-AS1 GGGTTCAAGTCAGCGACACT
H6C7 cells were incubated in RPMI 1640 medium containing miR-4673 ACACTCCAGCTGGGTCCAGGCAGGAGCCGG
miR-4673 TGGTGTCGTGGAGTCG
10% FBS (Gibco). All cells were maintained at 37°C in a humid-
miR-7855-5p ACACTCCAGCTGGGTTGGTGAGGACCCCAA
ified atmosphere with 5% CO2. miR-7855-5p TGGTGTCGTGGAGTCG
miR-876-3p CTGTGGTGGTTTACAAAGTAATT
miR-876-3p GTGCAGGGTCCGAGGT
2.2 Cell transfection miR-6783-3p ACACTCCAGCTGGGTTCCTGGGCTTCTCCT
miR-6783-3p TGGTGTCGTGGAGTCG
Short-hairpin RNAs (shRNAs) for the knockdown of SLCO4A1- miR-6514-3p ACACTCCAGCTGGGCTGCCTGTTCTTCCA
AS1 (sh-SLCO4A1-AS1#1/2) were constructed by GenePharma miR-6514-3p TGGTGTCGTGGAGTCG
(Shanghai, China), with sh-NC as the negative control miR-5001-3p ACACTCCAGCTGGGTTCTGCCTCTGTCCAG
miR-5001-3p TGGTGTCGTGGAGTCG
(NC). To upregulate SLCO4A1-AS1 and KIF21B, pcDNA3.1/
FBXL18 GATGATGACATGCACCCTGC
SLCO4A1-AS1 and pcDNA3.1/KIF21B were obtained from FBXL18 AGCTGCCTCACTTTGTCCTC
GenePharma, respectively. The empty pcDNA3.1 plasmid GPR37L1 TCTTGCTGTGATTTTGGCTGTG
was regarded as NC. Meanwhile, miR-4673 mimics, miR- GPR37L1 CTCTTGGATCGGCTCTGCTG
4673 inhibitors, and the negative control (NC mimics and CACNG8 ATCATTTCCCGGAGGACACG
CACNG8 CGCCGAAGTAGAAGGACCAG
NC inhibitors) were synthesized by Thermo Fisher Scientific
KIAA1614 CAGCATGAAGCATGGGCATC
(Carlsbad, CA, USA). Cell transfection was conducted uti- KIAA1614 GGTGCTCATACCAGGCACTT
lizing Lipofectamine 3000 (Invitrogen, Carlsbad, CA, USA) KREMEN1 ACAGTCTGAAATACCCCAACG
for 24 h according to the specification of the manufacturer. KREMEN1 GTTTCCATGATCCTTGTAGCAG
CYB561A3 CACTTCGTCCCACTGTGGTT
CYB561A3 AGGAGGAAGACAGCAAAGCC
KIF21B ACAGCGACTCCTCTTTGTCG
2.3 Quantitative real-time PCR (qRT-PCR) KIF21B TTGTGCGCATTGGGGATTTG
AP5S1 AACTACAACCCCCGAAGTGC
Total RNA was isolated and extracted from treated PC cells AP5S1 ATCAGAGCTGCTGGGTAGGA
using TRIzol Reagent (Invitrogen) under the guidelines of GAPDH forward TATGATGATATCAAGAGGGTAGT
the manufacturer. Moreover, complementary DNA (cDNA) GAPDH reverse TGTATCCAAACTCATTGTCATAC
U6 forward CTCGCTTCGGCAGCACA
was obtained by reverse transcription with extracted RNA
U6 reverse AACGCTTCACGAATTTGCGT
using PrimeScript RT Reagent Kit (Invitrogen). Later, qRT-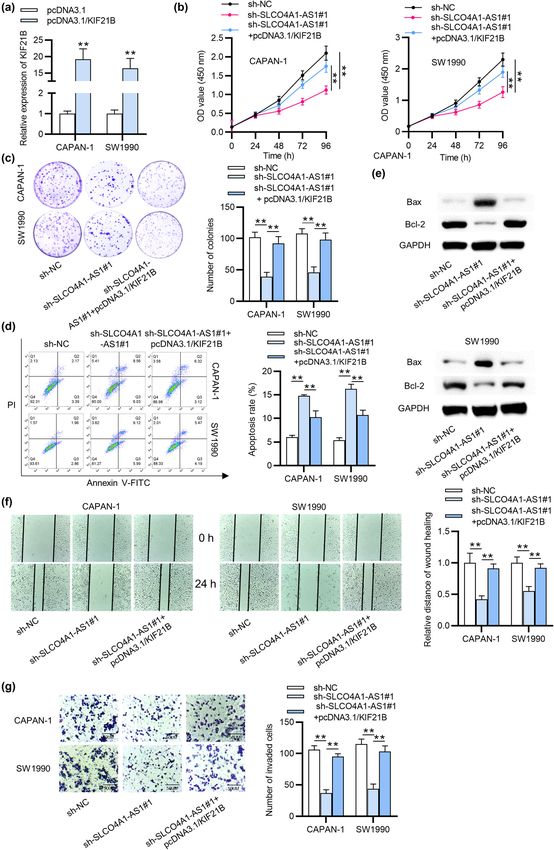
Role of SLCO4A1-AS1 in pancreatic cancer 255
reagent (Beyotime, Shanghai, China) after incubation for 2.8 Wound healing assay
0, 24, 48, 72, and 96 h, as instructed by the manufacturer.
Next, a microplate reader (Thermo Fisher Scientific) was The migratory capacity of the transfected PC cells was
used for detecting the absorbance at 450 nm of each determined by wound healing assay. Briefly, cells were
well. implanted into 6-well plates until 80% confluence. Then,
a 10 µL pipette tip was applied to scratch a straight line in
the center of the plates. Besides, the plates were cleaned
three times using phosphate-buffered saline to remove
2.5 Colony formation assay cell debris. Subsequently, the cells were incubated in
RPMI 1640 medium without serum for another 24 h.
For detecting cell proliferation, transfected PC cells were Finally, the recovery state of the wounds at 0 and 24 h
seeded in 6-well plates (1 × 103 cells/well), followed by was observed under a light microscope (Olympus).
2-week incubation in DMEM containing 10% FBS. Thereafter,
cells were stained with crystal violet (Sigma-Aldrich St.
Louis, MO, USA) for 30 min after mounted with paraformal- 2.9 Transwell invasion assay
dehyde (Sigma-Aldrich) for 15 min. Colony numbers were
calculated and analyzed after imaged with a microscope After transfection, PC cells were resuspended in serum-
(Olympus, Tokyo, Japan). free DMEM and plated into the upper chamber of 24-well
transwell (pore size 8 µm; Corning Costar) coated with
Matrigel (BD Biosciences). The lower chamber was filled
2.6 Flow cytometry analysis with DMEM containing 10% FBS. After incubation for 24 h,
noninvaded cells were removed with cotton swabs, while
Flow cytometry was performed to examine cell apoptosis invaded cells were fixed with 4% paraformaldehyde, and
with the utilization of Annexin V-FITC/PI Apoptosis Detection then stained with 0.1% crystal violet for 20 min. The invaded
Kit (KeyGEN, Nanjing, China). The harvested cells were cells were imaged with an inverted microscope (Olympus).
suspended in binding buffer and dyed with annexin
V-Fluorescein Isothiocyanate (V-FITC) solution and pro-
pidium iodide (PI) solution for 15 min. Subsequently, a 2.10 Subcellular fractionation assay
flow cytometer AccuriC6 (BD Biosciences, Franklin Lakes,
NJ, USA) was utilized to observe the dyed cells. According to the recommendations of the manufacturer,
a PARIS™ Kit (Invitrogen) was employed to perform the
subcellular fractionation assay. SW1990 and CAPAN-1
2.7 Western blot analysis cell lines placed into cell fractionation buffer were sub-
jected to centrifugation. Moreover, the supernatant was
48 h after transfection, aggregate proteins were lysed collected, and the remaining lysates were rinsed with cell
from PC cells utilizing RNA immunoprecipitation assay fractionation buffer. Next cell nuclei were lysed by cell
(RIPA) buffer (Beyotime). Equal amounts of proteins were disruption buffer. Thereafter, the lysate was incubated
loaded and segregated via 10% sodium dodecyl sulfate with the supernatant, ethanol, and 2 × lysis/binding solu-
polyacrylamide gel electrophoresis (SDS-PAGE), followed tion. Lastly, cytoplasmic and nuclear RNAs were investi-
by transferring to polyvinylidene fluoride (PVDF) mem- gated through qRT-PCR.
branes (Millipore, Boston, MA, USA). Later, the PVDF mem-
branes were blocked with 5% non-fat milk for 2 h, and
then incubated with primary antibodies overnight at 4°C, 2.11 Bioinformatics analysis
including anti-Bcl-2 (1:2,000, ab182858, Abcam), anti-Bax
(1:2,000, ab182733, Abcam), and anti-GAPDH (1:10,000, SLCO4A1-AS1 expression profile in PAAD is shown at
ab181603, Abcam). Then, after washing three times with GEPIA database (http://gepia.cancer-pku.cn/detail.php?
Tris-buffered saline with Tween-20 buffer, the membranes gene=&clicktag=boxplot). This dataset contains 179 PAAD
were incubated with HRP-conjugated secondary antibodies samples and 171 normal samples. DIANA tools – LncBase
at room temperature for 2 h. GAPDH served as a loading Predicted v2 (http://carolina.imis.athena-innovation.gr/diana_
control. The bands were analyzed using ECL detection tools/web/index.php?r=lncbasev2%2Findex) [16] was applied
system (Thermo Fisher Scientific). to search for downstream miRNAs of SLCO4A1-AS1 with the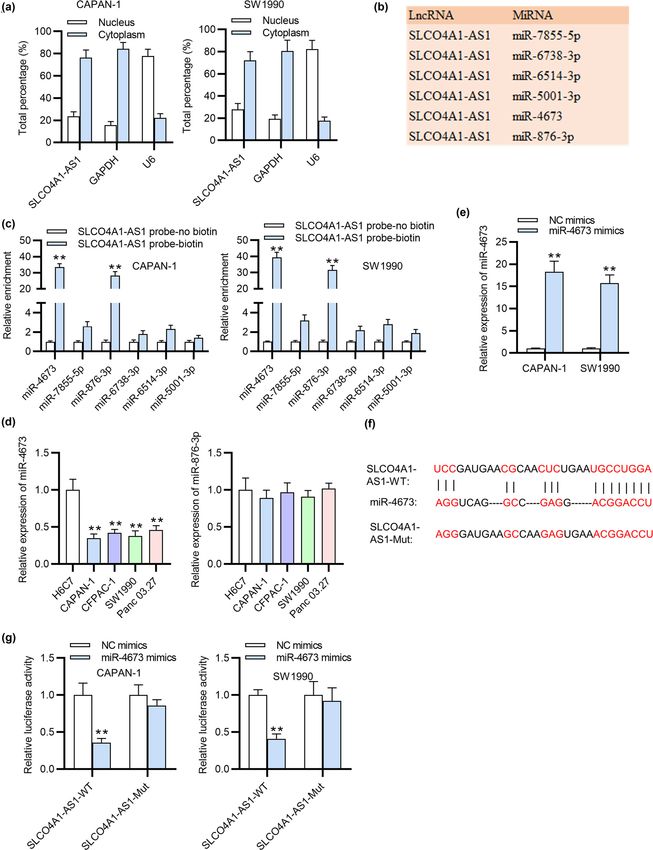
256 Jianxin Zhang et al.
screening condition of binding score more than 0.85 and pre- and analyzed using SPSS 18.0 software (SPSS, Chicago, IL,
dict the binding site between SLCO4A1-AS1 and miR-4673. After USA). The differences of two or more groups were assessed
screening out the target gene of miR-4673 with the screening by Student’s t test or one-way analysis of variance (ANOVA)
condition of cumulative weighted context++ score > −0.86, the followed by Tukey’s post hoc analysis, with p < 0.05 repre-
binding site between miR-4673 and KIF21B was analyzed with senting as statistically significant.
the utilization of TargetScan database (http://www.targetscan.
org/vert_71/) [17].
3 Results
2.12 RNA pull-down assay
3.1 Silencing SLCO4A1-AS1 inhibits the
Biotinylated SLCO4A1-AS1 and a control probe purchased
from Genechem Biotech (Shanghai, China) were trans- cellular processes of PC
fected into SW1990 and CAPAN-1 cells. Afterwards, cell
lysates were harvested and incubated with Dynabeads M- To examine whether SLCO4A1-AS1 was dysregulated in
280 Streptavidin (Sigma-Aldrich) in line with the manu- PC, we first explored the expression profile of SLCO4A1-
facturer’s protocols. After washing with wash buffer AS1 in PAAD from GEPIA database, and found that
(Invitrogen), RNA complexes were separated and puri- SLCO4A1-AS1 was upregulated in PAAD tissues compared
fied with lysis buffer and proteinase K. Finally, purified to normal tissues (Figure 1a). Additionally, SLCO4A1-AS1
RNA was analyzed by qRT-PCR. was overexpressed in PC cells in comparison to the normal
H6C7 cells, as shown by qRT-PCR analysis (Figure 1b).
Among the PC cell lines, SLCO4A1-AS1 expression in SW1990
2.13 Luciferase reporter assay and CAPAN-1 cells was the highest, thus they were singled out
for the following experiments. To figure out the function of
The fragments of wild-type (WT) and mutant (MUT) SLCO4A1-AS1 in PC, SW1990 and CAPAN-1 cells were
SLCO4A1-AS1 or KIF21B containing putative binding site transfected with sh-SLCO4A1-AS1#1/2 or sh-NC to measure
of miR-4673 were synthesized and subcloned into pmirGLO the efficiency of SLCO4A1-AS1 knockdown. We discovered
vectors (Promega, Madison, WI, USA) to generate the recom- that SLCO4A1-AS1 expression displayed a significant decrease
binant reporters including SLCO4A1-AS1-WT, SLCO4A1-AS1- in selected cells in contrast to NC group (Figure 1c). Next we
MUT, KIF21B-WT, and KIF21B-MUT. Next the reporters were carried out CCK-8 assay and found that the silencing of
co-transfected together with miR-4673 mimics, NC mimics, SLCO4A1-AS1 repressed PC cell viability (Figure 1d). The
or miR-4673 mimics + pcDNA3.1/SLCO4A1-AS1 into PC cells. findings of the colony formation assay also elucidated that
The luciferase activity after 48 h was evaluated by the Dual- SLCO4A1-AS1 downregulation attenuated cell proliferation
Luciferase Reporter Assay System (Promega). (Figure 1e). Furthermore, the influence of SLCO4A1-AS1
depletion on cell apoptosis examined by flow cytometry
and western blot analysis indicated that SLCO4A1-AS1
2.14 RIP assay downregulation induced apoptosis in PC cells (Figure 1f
and g). Subsequently, we conducted wound healing assay
Magna RNA immunoprecipitation Kit (Millipore) was used and discovered that SLCO4A1-AS1 depletion repressed cell
for RIP assay. All the cell lysates were incubated with RIPA migratory potential (Figure 1h). Later, the transwell assay
lysis buffer containing magnetic beads (Invitrogen) conju- demonstrated the repressed invasive ability of PC cells
gated to antibodies of human anti-Ago2 (1:30, ab186733, after the silencing of SLCO4A1-AS1 (Figure 1i). These find-
Abcam) or control anti-IgG (1:16, ab182931, Abcam). The ings elucidated that SLCO4A1-AS1 works as a cancer-pro-
coprecipitated RNA was eluted from the beads and ana- moting gene in PC cells.
lyzed by PCR.
3.2 SLCO4A1-AS1 acts as a molecular
2.15 Statistical analysis sponge of miR-4673
All experiments in the present study were repeated in Increasing studies have illustrated that lncRNAs exert
triplicate. The data are expressed as the mean value ± SD regulatory function in the malignant processes of tumors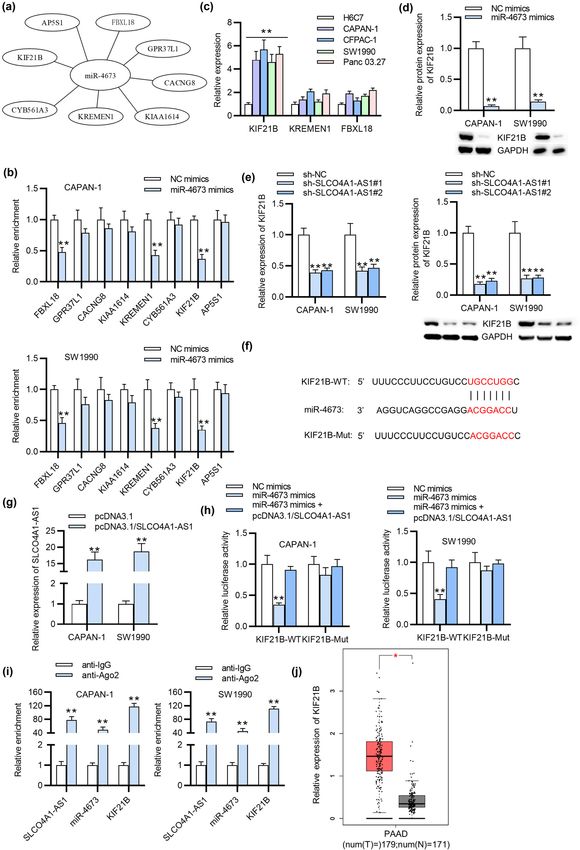
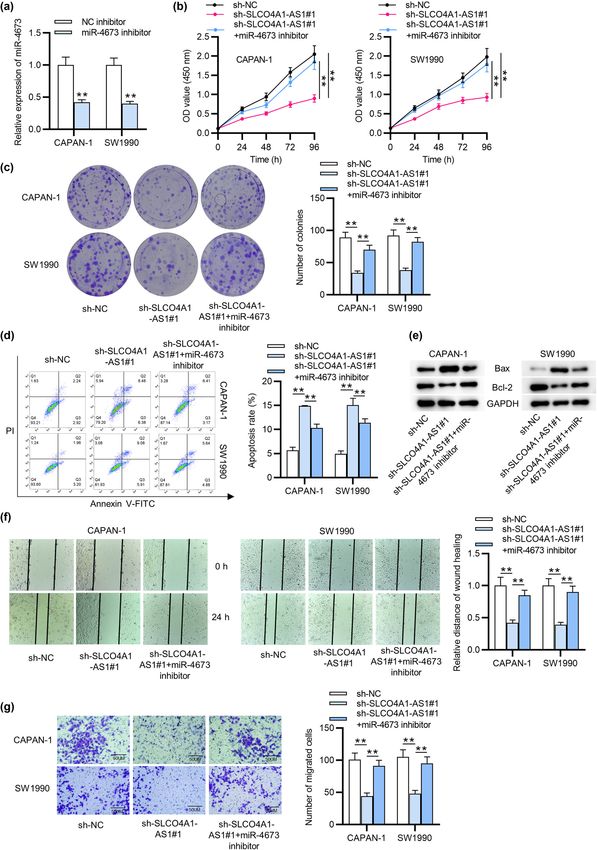
Role of SLCO4A1-AS1 in pancreatic cancer 257 Figure 1: Silencing SLCO4A1-AS1 inhibits the cellular processes of PC. (a) SLCO4A1-AS1 expression pattern in PAAD was revealed in the boxplot from GEPIA. (b) SLCO4A1-AS1 expression in PC cell lines in contrast with normal H6C7 cells was examined by qRT-PCR. (c) qRT-PCR investigated the interfering efficiency of SLCO4A1-AS1 in PC cells. (d) CCK-8 assay was used to detect PC cell viability affected by SLCO4A1- AS1 knockdown. (e) The effect of SLCO4A1-AS1 knockdown on CAPAN-1 and SW1990 cell proliferation was investigated by colony formation assay. (f and g) The apoptosis of PC cells transfected with sh-SLCO4A1-AS1 was analyzed by flow cytometry and western blot analysis. (h) Wound healing assay measured how downregulated SLCO4A1-AS1 affected cell migration. (i) Transwell assay detected the invasive cap- ability of PC cells after SLCO4A1-AS1 depletion. All experiments were performed in triplicate. More images for replicates are provided in a supplementary file named Replicate images. *p < 0.05 and **p < 0.01.
258 Jianxin Zhang et al.
by acting as competing endogenous RNAs (ceRNAs) to abilities of PC cells which was caused by SLCO4A1-AS1
sponge miRNAs [18]. Considering that the nuclear-cyto- silencing, as shown by wound healing and transwell
plasmic localization of lncRNAs plays an important part assays (Figure 3f and g). Hence, inhibition of miR-4673
in the molecular mechanism, we first conducted the sub- can reverse SLCO4A1-AS1 knockdown-mediated suppres-
cellular fractionation assay, which suggested that SLCO4A1- sive effects on the malignant phenotypes of PC cells.
AS1 was mainly located in cytoplasm of PC cells (Figure 2a).
Moreover, six putative downstream miRNAs interacting
with SLCO4A1-AS1 were selected from database of DIANA
(Figure 2b). Thereafter, the result of RNA pull-down sug- 3.4 KIF21B is a target gene for miR-4673
gested that miR-4673 and miR-876-3p were enriched in PC
cells treated with SLCO4A1-AS1 probe-biotin (Figure 2c). We next screened out eight candidate messenger RNAs
Next the expression of miR-4673 and miR-876-3p in PC cells (mRNAs) targeted by miR-4673 with TargetScan database
and normal cells was assessed by qRT-PCR, suggesting (Figure 4a). PCR analysis demonstrated that three mRNAs
that miR-4673 was significantly downregulated in PC cells (KIF21B, KREMEN1, and FBXL18) were markedly down-
(Figure 2d). In addition, the results of qRT-PCR demon- regulated in PC cells after upregulating miR-4673 (Figure 4b).
strated that miR-4673 expression was elevated in PC cells Next the level of KIF21B was enhanced in PC cells among
transfected with miR-4673 mimics (Figure 2e). Later, we three mRNAs, as illustrated by qRT-PCR (Figure 4c). The
analyzed the binding site of miR-4673 on SLCO4A1-AS1 protein level of KIF21B was decreased in PC cells upon
with the use of DIANA database (Figure 2f). Furthermore, miR-4673 overexpression, as indicated by western blot ana-
the binding relation between SLCO4A1-AS1 and miR-4673 lysis (Figure 4d). Additionally, silencing SLCO4A1-AS1 led to
was further verified by luciferase reporter assay. The find- a significant reduction in both mRNA and protein levels of
ings depicted that upregulated miR-4673 attenuated the KIF21B in PC cells (Figure 4e). Afterwards, the binding site of
luciferase activity of SLCO4A1-AS1-WT reporter vector, miR-4673 on KIF21B 3′-UTR was predicted by TargetScan
while the luciferase activity of SLCO4A1-AS1-MUT remained database (Figure 4f). Moreover, qRT-PCR showed that
almost unchanged with miR-4673 overexpression (Figure 2g). SLCO4A1-AS1 was upregulated in CAPAN-1 and SW1990 cells
Collectively, SLCO4A1-AS1 can directly sponge miR-4673. after transfection of pcDNA3.1/SLCO4A1-AS1 (Figure 4g).
Later, the relationship among SLCO4A1-AS1, miR-4673, and
KIF21B was detected by luciferase reporter assay, which
demonstrated that the luciferase activity of KIF21B-WT was
3.3 Inhibition of miR-4673 reverses the attenuated by miR-4673 upregulation and then enhanced by
effects of SLCO4A1-AS1 silencing on the SLCO4A1-AS1 overexpression in SW1990 and CAPAN-1 cell
phenotypes of PC cells lines, while the luciferase activity of KIF21B-MUT witnessed
no significant change (Figure 4h). Besides, the RIP assay
Subsequently, we investigated whether miR-4673 inhibi- examined the interaction of SLCO4A1-AS1, miR-4673, and
tion had an effect on the carcinogenic role of SLCO4A1- KIF21B, indicating their co-existence in the RNA-induced
AS1 in PC. As shown by qRT-PCR, miR-4673 was significantly silencing complex (RISC) (Figure 4i). Additionally, the data
downregulated in miR-4673 inhibitors-transfected CAPAN-1 from GEPIA database reveal the upregulation of KIF21B in PAAD
and SW1990 cells (Figure 3a). The results from CCK-8 tissues (Figure 4j). These findings elucidated that KIF21B is the
assay demonstrated that miR-4673 inhibitors significantly downstream target for miR-4673, and SLCO4A1-AS1 upregulates
attenuated SLCO4A1-AS1 knockdown-induced suppres- KIF21B expression at a miR-4673 dependent way.
sion of PC cell viability (Figure 3b). Similarly, colony
formation assay showed that SLCO4A1-AS1 knockdown
led to a significant reduction in the number of colo-
nies of CAPAN-1 and SW1990 cells; however, treat- 3.5 SLCO4A1-AS1 positively modulates the
ment of miR-4673 inhibitors was shown to alleviate this malignant progression of PC cells via
effect (Figure 3c), indicating that miR-4673 inhibitors might upregulating KIF21B
have a promotive effect on cell proliferation. As displayed
by Figure 3d and e, SLCO4A1-AS1 silencing-induced cell To further expose the functional mechanism of SLCO4A1-
apoptosis was partially reversed by miR-4673 inhibitors in AS1 in PC cells, rescue assays were carried out. First, qRT-
CAPAN-1 and SW1990 cells. Furthermore, miR-4673 inhibi- PCR depicted that KIF21B expression was increased in PC
tion rescued the decrease in the migratory and invasive cells transfected with pcDNA3.1/KIF21B (Figure 5a). NextRole of SLCO4A1-AS1 in pancreatic cancer 259 Figure 2: miR-4673 is directly absorbed by SLCO4A1-AS1. (a) Subcellular fractionation assay determined the localization of SLCO4A1-AS1 in PC cells. (b) We searched the downstream miRNAs of SLCO4A1-AS1 from the DIANA database. (c) RNA pull-down investigated the expression of candidate miRNAs pulled down by SLCO4A1-AS1 probe-biotin in PC cells. (d) The levels of selected miRNAs in PC cells were measured through qRT-PCR. (e) The overexpression efficiency of miR-4673 was subjected to qRT-PCR. (f) The binding site of SLCO4A1-AS1 on miR-4673 is revealed from DIANA database. (g) Luciferase reporter assay was employed to examine the luciferase activity of SLCO4A1-AS1-WT and SLCO4A1-AS1-MUT in PC cells transfected with miR-4673 mimics. All experiments were performed in triplicate. **p < 0.01.
260 Jianxin Zhang et al. Figure 3: miR-4673 inhibition reverses SLCO4A1-AS1 knockdown-induced suppressive effects on the phenotypes of PC cells. (a) qRT-PCR analysis for detecting the transfection efficiency of miR-4673 inhibitors in CAPAN-1 and SW1990 cells. (b) The viabilities of CAPAN-1 and SW1990 cells transfected with sh-NC, sh-SLCO4A1-AS1#1, or sh-SLCO4A1-AS1#1 + miR-4673 inhibitors were analyzed by CCK-8 assay. (c) Colony formation assay was used to examine the proliferation of PC cells with above transfection. (d and e) The apoptosis of transfected PC cells was assessed by flow cytometry and western blot analysis. (f and g) Wound healing and transwell assays were performed to evaluate the migration and invasion of PC cells with above transfection. All experiments were performed in triplicate. More images for replicates are provided in a supplementary file named Replicate images. **p < 0.01.
Role of SLCO4A1-AS1 in pancreatic cancer 261 Figure 4: KIF21B is targeted by miR-4673. (a) The eight candidate mRNAs of miR-4673 were selected from TargetScan database. (b) qRT-PCR analyzed the expression of these miRNAs in PC cells transfected with miR-4673 mimics. (c) The expression of selected mRNAs in PC cells were detected through qRT-PCR. (d) The protein level of KIF21B in PC cells after miR-4673 upregulation was analyzed by qRT-PCR. (e) qRT-PCR assessed both the mRNA and protein levels of KIF21B in PC cells after silencing SLCO4A1-AS1. (f) We searched the binding site of KIF21B on miR-4673 from TargetScan database. (g) The overexpression efficiency of SLCO4A1-AS1 in PC cells was measured through qRT-PCR. (h) Luciferase reporter assay detected the luciferase activity of KIF21B-WT and KIF21B-MUT in PC cells transfected with miR-4673 mimics or miR-4673 mimics + pcDNA3.1/ SLCO4A1-AS1. (i) RIP assay was applied to confirm the co-existence of SLCO4A1-AS1, miR-4673, and KIF21B in RISC. (j) The expression of KIF21B in PAAD and normal tissues is shown by GEPIA database. All experiments were performed in triplicate. *p < 0.05 and **p < 0.01.
262 Jianxin Zhang et al. Figure 5: SLCO4A1-AS1 accelerates the cellular process of PC through regulating KIF21B. (a) qRT-PCR detected the overexpression efficiency of KIF21B in PC cells. (b) CCK-8 assay was carried out to investigate the impact of upregulated KIF21B on suppressed cell viability caused by depleting SLCO4A1-AS1. (c) The SLCO4A1-AS1 silence-reduced proliferation capacity of PC cells was affected by KIF21B overexpression, as indicated by colony formation assay. (d and e) The apoptosis of PC cells transfected with appointed plasmids was analyzed by flow cytometry and western blot analysis. (f) The influence of SLCO4A1-AS1 knockdown on KIF21B overexpression-enhanced migrated capability of PC cells was determined by wound healing assay. (g) The change in invasive capability of PC cells with designated transfections was investigated by Transwell assay. All experiments were performed in triplicate. More images for replicates are provided in a supplementary file named Replicate images. **p < 0.01.
Role of SLCO4A1-AS1 in pancreatic cancer 263
as indicated by CCK-8 assay, SW1990 and CAPAN-1 cell It has been elucidated that lncRNAs serve as ceRNAs
viabilities reduced by the depletion of SLCO4A1-AS1 were by performing as molecular sponges of miRNAs, thereby
reversed by overexpressed KIF21B (Figure 5b). In addi- regulating the specific target genes for miRNAs [25]. For
tion, the results of colony formation assay depicted that instance, lncRNA LINC01559 promotes the cellular pro-
the SLCO4A1-AS1 silence-mediated decrease in prolifera- cess of PC by sponging miR-1343-3p to upregulate Raf-1
tive ability of PC cells was countervailed by the upregula- proto-oncogene, serine/threonine kinase level [26]. In
tion of KIF21B (Figure 5c). Flow cytometry and western our study, the specific miR-4673 sponged by SLCO4A1-
blot analysis elucidated that enhanced KIF21B expression AS1 was selected with bioinformatic tools. miR-4673 was
offset the apoptosis rate of PC cells formerly increased by reported as a tumor suppressor in hepatocellular carci-
SLCO4A1-AS1 downregulation (Figure 5d and e). Further- noma [27]. Presently, we verified that the miR-4673 was
more, the suppressive influence of SLCO4A1-AS1 down- downregulated in PC cells. Moreover, the binding relation
regulation on PC cell migratory capacity was rescued by between SLCO4A1-AS1 and miR-4673 was detected. Inhi-
the overexpression of KIF21B as shown by wound healing bition of miR-4673 was shown to reverse SLCO4A1-AS1
assay (Figure 5f). Later, the result of transwell assay indi- knockdown-mediated inhibitory effects on the malignant
cated the counteracting impact of upregulated KIF21B on behaviors of PC cells. Therefore, miR-4673 can directly
the inhibited cell invasion of SW1990 and CAPAN-1 cells interact with SLCO4A1-AS1.
caused by the knockdown of SLCO4A1-AS1 (Figure 5g). These Subsequently, KIF21B, as the downstream gene for
data illustrated that SLCO4A1-AS1 upregulates KIF21B expres- miR-4673, was screened out from TargetScan database.
sion by competitively interacting with miR-4673, thereby KIF21B belongs to the motor of Kinesin-4 motor and is
modulating the malignant behaviors of PC cells. located at 1q32.1 [28]. Previous studies validated that
KIF21B, a classic kinesin protein, was able to regulate
microtubule dynamics [29]. It is widely known that kinesin
proteins and microtubules exert pivotal functions in tumor
4 Discussion progression intercellular and signal transduction [30]. It
was reported that KIF21B exhibited a carcinogenic function
As one prevalent in digestive malignancy with high in non-small cell lung cancer; silencing KIF21B represses
morbidity and mortality, PC has imposed tremendous cell growth and induces apoptosis [28]. Furthermore, KIF21B
pressure on public health [19]. Moreover, despite new tech- is overexpressed and facilitates cellular behaviors in hepa-
nologies and therapies for early diagnosis, most patients tocellular carcinoma [31]. In the current study, we verified
are still diagnosed at an advanced stage for the insidious the overexpression of KIF21B and the binding capacity
onset of this disease [20]. Unfortunately, these patients between miR-4673 and KIF21B in PC cells. Furthermore,
are unlikely subject to radical resection, and adjuvant the repressed cellular processes of PC by depleting SLCO4A1-
chemotherapy has limited influence because of the type AS1 was rescued with the upregulation of KIF21B.
and efficacy [21]. Hence, there is an exigent need to figure In summary, we identified SLCO4A1-AS1 as a cancer-
out novel and effective biomarkers to improve the prognosis causing gene, exerting critical functions in cellular pro-
of PC. Previous studies have revealed that the aberrant cesses of PC. Mechanistically, SLCO4A1-AS1, as a ceRNA,
expression of lncRNAs plays a pivotal role in regulating upregulates KIF21B to positively modulate the molecular
the malignant properties of PC. For instance, lncRNA processes of PC via sponging miR-4673. These results
nuclear paraspeckle assembly transcript 1 facilitates the showed that SLCO4A1-AS1/miR-4673/KIF21B axis might
tumorigenesis and growth of PC via stabilizing E74 like be an effective biomarker for the prognosis of PC, dee-
ETS transcription factor 3 [22]. LncRNA 00976 upregulates pening the understanding of its pathogenesis. Other
OTU deubiquitinase 7B to enhance PC cell growth and inva- possible molecules involved in SLCO4A1-AS1-mediated
sion by binding with miR-137 [23]. LINC01559 facilitates PC ceRNA network require further investigation. For example,
cell proliferation and migration through Yes1 associated miR-223-3p and miR-335-5p were reported to a down-
transcriptional regulator-mediated pathway [24]. Neverthe- stream molecule of SLCO4A1-AS1 involved in non-small-
less, the role of SLCO4A1-AS1 in the biological mechanism cell lung cancer and bladder cancer, respectively [13,15].
of PC remains unclear. Herein we confirmed that SLCO4A1- Additionally, miR-223-3p was reported to be upregulated
AS1 was overexpressed in PC. Additionally, SLCO4A1-AS1 in the serum of PC and miR-335-5p was shown to be
downregulation repressed cell growth and motion, and sti- sponged by LINC00941 and involved in the progression
mulated cell apoptosis of PC, indicating that SLCO4A1-AS1 of PC [32,33]. More studies are needed to be carried out
exerted oncogenic functions in PC. in the future.264 Jianxin Zhang et al.
Abbreviations [4] Fogel EL, Shahda S, Sandrasegaran K, DeWitt J, Easler JJ,
Agarwal DM, et al. A Multidisciplinary approach to pancreas
CCK-8 cell counting kit-8 cancer in 2016: a review. Am J Gastroenterol. 2017;112(4):537–54.
[5] Zhan HX, Xu JW, Wu D, Zhang TP, Hu SY. Pancreatic cancer
cDNA complementary DNA
stem cells: new insight into a stubborn disease. Cancer Lett.
ceRNA competing endogenous RNA 2015;357(2):429–37.
DMEM Dulbecco’s modified eagle’s medium [6] Manji GA, Olive KP, Saenger YM, Oberstein P. Current and
KIF21B kinesin family member 21B emerging therapies in metastatic pancreatic cancer. Clin
lncRNAs long non-coding RNAs Cancer Res. 2017;23(7):1670–8.
[7] Quinn JJ, Chang HY. Unique features of long non-coding RNA
NC negative control
biogenesis and function. Nat Rev Genet. 2016;17(1):47–62.
mRNA messenger RNA [8] Li J, Meng H, Bai Y, Wang K. Regulation of lncRNA and its role in
miR-4673 microRNA-4673 cancer metastasis. Oncol Res. 2016;23(5):205–17.
MUT mutant [9] Liu J, Xu R, Mai SJ, Ma YS, Zhang MY, Cao PS, et al. LncRNA
PAAD pancreatic adenocarcinoma CSMD1-1 promotes the progression of hepatocellular carci-
noma by activating MYC signaling. Theranostics.
PC pancreatic cancer
2020;10(17):7527–44.
PI propidium iodide
[10] Zhang W, Shi X, Chen R, Zhu Y, Peng S, Chang Y, et al. Novel
PVDF polyvinylidene fluoride long non-coding RNA lncAMPC promotes metastasis and
qRT-PCR quantitative real-time PCR immunosuppression in prostate cancer by stimulating LIF/LIFR
RIPA RNA immunoprecipitation assay expression. Mol Ther. 2020;28(11):2473–87.
SDS-PAGE sodium dodecyl sulfate polyacrylamide [11] Wang L, Xiao B, Yu T, Gong L, Wang Y, Zhang X, et al. lncRNA
PVT1 promotes the migration of gastric cancer by functioning
gel electrophoresis
as ceRNA of miR-30a and regulating Snail. J Cell Physiol.
shRNAs short-hairpin RNAs 2021;236(1):536–48.
SLCO4A1-AS1 lncRNA solute carrier organic anion [12] Wei Y, Wei L, Li J, Ma Z, Zhang Q, Han Z, et al. SLCO4A1-AS1
transporter family member 4A1 anti- promotes cell growth and induces resistance in lung adeno-
sense RNA 1 carcinoma by modulating miR-4701-5p/NFE2L1 axis to activate
V-FITC V-fluorescein isothiocyanate WNT pathway. Cancer Med. 2020;9(19):7205–17.
[13] Li Q, Jiang B, Qi Y, Zhang H, Ma H. Long non-coding RNA
WT wild type
SLCO4A1-AS1 drives the progression of non-small-cell lung
cancer by modulating miR-223-3p/IKKα/NF-κB signaling.
Acknowledgements: Not applicable. Cancer Biol Ther. 2020;21(9):806–14.
[14] Yu J, Han Z, Sun Z, Wang Y, Zheng M, Song C. LncRNA SLCO4A1-
AS1 facilitates growth and metastasis of colorectal cancer
Funding information: This work was supported by National
through β-catenin-dependent Wnt pathway. J Exp Clin Cancer
Natural Science Foundation of China (81902501). Res: CR. 2018;37(1):222.
[15] Yang Y, Wang F, Huang H, Zhang Y, Xie H, Men T. lncRNA
Conflict of interest: The authors declare that there is no SLCO4A1-AS1 promotes growth and invasion of bladder cancer
conflict of interests. through sponging miR-335-5p to upregulate OCT4.
OncoTargets Ther. 2019;12:1351–8.
[16] Paraskevopoulou MD, Vlachos IS, Karagkouni D,
Data availability statement: The datasets used or ana- Georgakilas G, Kanellos I, Vergoulis T, et al. DIANA-LncBase
lyzed during the current study are available from the v2: indexing microRNA targets on non-coding transcripts.
corresponding author on reasonable request. Nucleic Acids Res. 2016;44(D1):D231–8.
[17] Agarwal V, Bell GW, Nam JW, Bartel DP. Predicting effective
microRNA target sites in mammalian mRNAs. eLife.
2015;4:e05005.
[18] Cheng D, Fan J, Ma Y, Zhou Y, Qin K, Shi M, et al. LncRNA
References SNHG7 promotes pancreatic cancer proliferation through ID4
by sponging miR-342-3p. Cell & Biosci. 2019;9:28.
[1] Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA: [19] Dauer P, Nomura A, Saluja A, Banerjee S. Microenvironment in
Cancer J Clinicians. 2019;69(1):7–34. determining chemo-resistance in pancreatic cancer: neigh-
[2] Neuzillet C, Tijeras-Raballand A, Bourget P, Cros J, borhood matters. Pancreatology. 2017;17(1):7–12.
Couvelard A, Sauvanet A, et al. State of the art and future [20] Zhu H, Li T, Du Y, Li M. Pancreatic cancer: challenges and
directions of pancreatic ductal adenocarcinoma therapy. opportunities. BMC Med. 2018;16(1):214.
Pharmacol Therapeutics. 2015;155:80–104. [21] Pan S, Shen M, Zhou M, Shi X, He R, Yin T, et al. Long non-
[3] Miller KD, Nogueira L, Mariotto AB, Rowland JH, Yabroff KR, coding RNA LINC01111 suppresses pancreatic cancer aggres-
Alfano CM, et al. Cancer treatment and survivorship statistics, siveness by regulating DUSP1 expression via microRNA-3924.
2019. CA: Cancer J Clinicians. 2019;69(5):363–85. Cell Death Dis. 2019;10(12):883.Role of SLCO4A1-AS1 in pancreatic cancer 265
[22] Feng Y, Gao L, Cui G, Cao Y. LncRNA NEAT1 facilitates pan- [28] Sun ZG, Pan F, Shao JB, Yan QQ, Lu L, Zhang N. Kinesin
creatic cancer growth and metastasis through stabilizing ELF3 superfamily protein 21B acts as an oncogene in non-small cell
mRNA. Am J Cancer Res. 2020;10(1):237–48. lung cancer. Cancer Cell Int. 2020;20:233.
[23] Lei S, He Z, Chen T, Guo X, Zeng Z, Shen Y, et al. Long non- [29] Muhia M, Thies E, Labonté D, Ghiretti AE, Gromova KV,
coding RNA 00976 promotes pancreatic cancer progression Xompero F, et al. The kinesin KIF21B regulates microtubule
through OTUD7B by sponging miR-137 involving EGFR/MAPK dynamics and is essential for neuronal morphology, synapse
pathway. J Exp Clin Cancer Res: CR. 2019;38(1):470. function, and learning and memory. Cell Rep.
[24] Lou C, Zhao J, Gu Y, Li Q, Tang S, Wu Y, et al. LINC01559 2016;15(5):968–77.
accelerates pancreatic cancer cell proliferation and migration [30] He M, Subramanian R, Bangs F, Omelchenko T, Liem Jr. KF,
through YAP-mediated pathway. J Cell Physiol. Kapoor TM, et al. The kinesin-4 protein Kif7 regulates mam-
2020;235(4):3928–38. malian Hedgehog signalling by organizing the cilium tip
[25] Militello G, Weirick T, John D, Döring C, Dimmeler S, Uchida S. compartment. Nat Cell Biol. 2014;16(7):663–72.
Screening and validation of lncRNAs and circRNAs as miRNA [31] Zhao HQ, Dong BL, Zhang M, Dong XH, He Y, Chen SY, et al.
sponges. Brief Bioinforma. 2017;18(5):780–8. Increased KIF21B expression is a potential prognostic bio-
[26] Chen X, Wang J, Xie F, Mou T, Zhong P, Hua H, et al. Long marker in hepatocellular carcinoma. World J Gastrointest
noncoding RNA LINC01559 promotes pancreatic cancer pro- Oncol. 2020;12(3):276–88.
gression by acting as a competing endogenous RNA of miR- [32] Wang J, He Z, Xu J, Chen P, Jiang J. Long noncoding RNA
1343-3p to upregulate RAF1 expression. Aging. LINC00941 promotes pancreatic cancer progression by com-
2020;12(14):14452–66. petitively binding miR-335-5p to regulate ROCK1-mediated
[27] Pu J, Wei H, Tan C, Qin B, Zhang Y, Wang A, et al. Long non- LIMK1/Cofilin-1 signaling. Cell Death Dis. 2021;12(1):36.
coding RNA SNHG14 facilitates hepatocellular carcinoma pro- [33] Zou X, Wei J, Huang Z, Zhou X, Lu Z, Zhu W, et al. Identification
gression through regulating miR-4673/SOCS1. Am J Transl of a six-miRNA panel in serum benefiting pancreatic cancer
Res. 2019;11(9):5897–904. diagnosis. Cancer Med. 2019;8(6):2810–22.You can also read

















































