Resolved-Editorial Expression of Concern: Visualizing active viral infection reveals diverse cell fates in synchronized algal bloom demise - PNAS
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Resolved—Editorial Expression of Concern: Visualizing active viral infection
reveals diverse cell fates in synchronized algal bloom demise
MICROBIOLOGY
PNAS is publishing an Editorial Expression of Concern regarding the
following article: “Visualizing active viral infection reveals diverse cell
fates in synchronized algal bloom demise,” by Flora Vincent, Uri
Sheyn, Ziv Porat, Daniella Schatz, and Assaf Vardi, which was first
published March 11, 2021; 10.1073/pnas.2021586118 (Proc. Natl.
Acad. Sci. U.S.A. 118, e2021586118). The editors are issuing a note to
alert readers of a potential problem with data in this study. The au-
thors contacted PNAS ahead of publication to request a delay, not-
ing, “We recently received Pacbio sequences of Emiliania huxleyi
(long-read nuclear and chloroplast transcriptome sequencing). A first
annotation of this data revealed a mistake in the previous annotation
of the psbA gene, which we used to design our smFISH probes
MICROBIOLOGY
presented in this study. The original probes we used in the paper were
designed based on a Emiliania huxleyi transcriptome that predicted
the gene structure using Illumina short-read RNA sequencing, thus
our probes are only partially covering the psbA gene. We are pres-
ently revalidating our results to assess the possible implications of the
previous misannotation of the E. huxleyi transcriptome. Importantly,
the viral probes are all validated, thus all main findings concerning
viral infection dynamics in the lab and in the field are fully con-
firmed.” Unfortunately, the request was not caught in our production
process in time to stop publication. We will update readers as soon as
the authors have completed their reanalysis.
As of June 22, the authors have rerun their experiments and
confirmed the initial findings. See the correction to the article
for additional information.
May R. Berenbaum
Editor-in-Chief
Published under the PNAS license.
Published April 7, 2021.
www.pnas.org/cgi/doi/10.1073/pnas.2105198118
Downloaded by guest on February 20, 2022
PNAS 2021 Vol. 118 No. 15 e2105198118 https://doi.org/10.1073/pnas.2105198118 | 1 of 1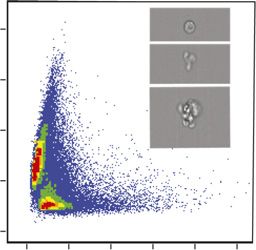
Correction
MICROBIOLOGY
Correction for “Visualizing active viral infection reveals diverse
cell fates in synchronized algal bloom demise,” by Flora Vincent,
Uri Sheyn, Ziv Porat, Daniella Schatz, and Assaf Vardi, which
published March 11, 2021; 10.1073/pnas.2021586118 (Proc. Natl.
Acad. Sci. U.S.A. 118, e2021586118).
The authors note, “The reference sequence used to design the
psbA smFISH probes presented in the manuscript was based on
RNAseq Illumina short read sequencing. We recently produced
an updated E. huxleyi transcriptome obtained with PacBio long
read sequencing. Therefore, we constructed a new set of psbA
smFISH probes and validated all the conclusions that are pre-
sented in the manuscript. Dataset S2 contains the updated psbA
reference sequence, individual smFISH probes, and corre-
sponding location on the psbA transcript.” The new dataset has
been added to the article’s Supporting Information.
CORRECTION
Published under the PNAS license.
Published June 21, 2021.
www.pnas.org/cgi/doi/10.1073/pnas.2110243118
PNAS 2021 Vol. 118 No. 26 e2110243118 https://doi.org/10.1073/pnas.2110243118 | 1 of 1
Visualizing active viral infection reveals diverse cell
fates in synchronized algal bloom demise
Flora Vincenta, Uri Sheyna,1, Ziv Poratb, Daniella Schatza, and Assaf Vardia,2
a
Department of Plant and Environmental Sciences, Weizmann Institute of Science, 7610001 Rehovot, Israel; and bFlow Cytometry Unit, Life Sciences Core
Facilities, Weizmann Institute of Science, 7610001 Rehovot, Israel
Edited by James L. Van Etten, University of Nebraska, Lincoln, NE, and approved February 1, 2021 (received for review October 15, 2020)
Marine viruses are the most abundant biological entity in the diverse infection states and cell fates of the virocells. Single-
ocean and are considered as major evolutionary drivers of micro- Molecule mRNA Fluorescent In situ Hybridization (smFISH)
bial life [C. A. Suttle, Nat. Rev. Microbiol. 5, 801–812 (2007)]. Yet, enables detection, enumeration, and localization of single mes-
we lack quantitative approaches to assess their impact on the ma- senger RNA (mRNA) molecules within morphologically intact
rine ecosystem. Here, we provide quantification of active viral in- individual cells (23). In this approach, mRNAs are fluorescently
fection in the bloom forming single-celled phytoplankton Emiliania labeled whereby probe fluorescence intensity becomes a proxy
huxleyi infected by the large virus EhV, using high-throughput for gene expression levels (24). Therefore, smFISH represents a
single-molecule messenger RNA in situ hybridization (smFISH) of both tool to visualize transcripts involved in specific host–virus inter-
virus and host transcripts. In natural samples, viral infection reached actions within microbial consortia.
only 25% of the population despite synchronized bloom demise ex- Here, we provide unprecedented quantification of active viral
posing the coexistence of infected and noninfected subpopulations. infection and host response of the cosmopolitan unicellular alga
We prove that photosynthetically active cells chronically release viral
Emiliania huxleyi both in the laboratory and during algal bloom
particles through nonlytic infection and that viral-induced cell lysis can
succession in the natural environment. Demise of E. huxleyi’s
MICROBIOLOGY
occur without viral release, thus challenging major assumptions re-
massive oceanic blooms is attributed to infection by its large
garding the life cycle of giant viruses. We could also assess active
infection in cell aggregates linking viral infection and carbon export
double-strand DNA (dsDNA) virus EhV (3, 25, 26), a member of
to the deep ocean [C. P. Laber et al., Nat. Microbiol. 3, 537–547 (2018)]
the nucleo-cytoplasmic large DNA viruses (27). We used smFISH
and suggest a potential host defense strategy by enrichment of in- to quantify host and virus mRNA and tracked infection dynamics
fected cells in sinking aggregates. Our approach can be applied to at a single-virocell level by high-throughput imaging flow cytom-
diverse marine microbial systems, opening a mechanistic dimension etry. Our study provides unique insights into the life cycle and
to the study of biotic interactions in the ocean. ecology of an ecologically important giant virus.
Results
giant virus | algal blooms | viral life cycle | single cell | smFISH In order to quantify dynamics of active viral infection, a time
course of E. huxleyi infection at high virus:host ratio was per-
M icrobial communities play a crucial role in shaping marine
ecosystems and biogeochemical cycles (1–3). In particular,
abundant photosynthetic microeukaryotes and cyanobacteria
formed, comparing infected and noninfected E. huxleyi cultures.
(phytoplankton) contribute to half of the total primary produc- Significance
tion on Earth and form the basis of the marine food web (4–6).
Phytoplankton blooms can undergo synchronized demise fol- Despite years of research in aquatic virology, we remain unable
lowing infection by viruses, present in up to 108 viruses per liter to estimate viral-induced mortality in the ocean and, conse-
of seawater (3, 7). One of the main challenges in aquatic virology quently, to resolve viral impact on nutrient fluxes and micro-
is to quantify how viruses, through control of the host metabo- bial dynamics. Here, we assess active infection in algal single
lism and abundance, remodel nutrient fluxes within major bio- cells by subcellular visualization of virus and host transcripts,
geochemical cycles and impact the microbial community via the revealing the coexistence of infected and noninfected sub-
viral shunt (8–10). This challenge meets the critical need to as- populations. We revisit major assumptions of a giant virus’ life
sess active viral infection among host cells (virocells) within a cycle: cells can produce virions without lysing and can lyse
complex microbial consortium in the marine environment. without producing virions. In a natural algal bloom, only 25%
Approaches such as bulk RNA sequencing (RNA-Seq) (11, of cells were infected, highlighting the importance of other
12), Digital Droplet PCR (13), polony (14), viral-BONCAT (15), mortality agents. Enrichment of infected cells in cell aggregates
phageFISH (16), and single-cell genomics (17, 18) have consid- suggests potential host defense strategies. Our approach
erably expanded the viral ecology toolbox and provided insights opens a mechanistic dimension to the study of marine microbial
into viral diversity and virus-encoded auxiliary metabolic genes interactions.
that remodel the cell metabolism (19). However, these methods
Author contributions: F.V. and A.V. conceptualized and designed all aspects of the re-
do not report active viral infection and its heterogeneity among search; F.V., U.S., and D.S. performed research; Z.P. contributed analytic tools; F.V. ana-
individual virocells, which are defined as “the living form of the lyzed data; and F.V. and A.V. wrote the paper.
virus [...] that is, in its intra-cellular form” (ref. 20, p. 233). Recent The authors declare no competing interest.
advances in single-cell transcriptomics have greatly improved our This article is a PNAS Direct Submission.
understanding of host–virus interactions by uncovering diverse This open access article is distributed under Creative Commons Attribution License 4.0
infection states and dynamics within individual virocells (21, 22). (CC BY).
However, these methods remain expensive and restricted to lab- 1
Present address: Research and Education, Daniel K. Inouye Center for Microbial Ocean-
oratory cultures and lack the ability to visualize infection on the ography, University of Hawai’i at Manoa, Honolulu, HI 96822.
subcellular level that can provide fresh insights into the complex 2
To whom correspondence may be addressed. Email: assaf.vardi@weizmann.ac.il.
life cycle of viruses and the interplay with their host. Furthermore, This article contains supporting information online at https://www.pnas.org/lookup/suppl/
we are still missing the ability to quantify active infection at the doi:10.1073/pnas.2021586118/-/DCSupplemental.
single-cell level in natural blooms that will enable the definition of Published March 11, 2021.
PNAS 2021 Vol. 118 No. 11 e2021586118 https://doi.org/10.1073/pnas.2021586118 | 1 of 8
Samples were collected to count cell and viral abundances as well active infected cells (mcp+/psbA+) are responsible for early viral
as for smFISH probing. To track the metabolic state of E. huxleyi release, indicating a nonlytic infection phase. This was supported
during viral infection, we monitored the mRNA expression of by transmission electron microscopy (TEM) of infected cells at 4
psbA, a chloroplast-encoded gene of the D1 protein. D1 is a major hpi, displaying viral egress from undamaged cells that display
component of the photosystem II complex involved in the first intact chloroplast morphology and thylakoid stacking (Fig. 2E).
step of the light reaction during photosynthesis. D1 has a rapid mcp+/psbA+ cells had an average DAPI intensity four times
turnover, requiring constant transcription of psbA (28), and is higher than other subpopulations and showed strong positive
essential for optimal viral infection of E. huxleyi (29). Concomi- correlation between the mcp signal and DAPI intensity (SI Ap-
tantly, we followed the expression of the viral mcp gene that en- pendix, Fig. S3). This is likely a consequence of the virus-induced
codes the major capsid protein expressed at the late stage of the de novo nucleotide synthesis required to meet the high metabolic
viral infection (22) (Fig. 1A). Epifluorescence microscopy was demand of giant viruses (30). Utilizing de novo nucleotide syn-
used to analyze the samples at 1 and 24 h postinfection (hpi) and thesis rather than host DNA degradation or nucleotide recycling,
showed an increase in the fraction of cells that express mcp at which would compromise cell viability, may serve as a viral hijacking
24 hpi compared to 1 hpi, in concert with a profound loss of psbA strategy at early stages of infection (Fig. 2F and SI Appendix, Fig.
expression, thus validating our approach (Fig. 1 B and C). Host- S4B). Among mcp+/psbA– cells at 24 hpi, mcp and DAPI signals
cell abundance of infected cultures remained relatively stable at a were colocalized, suggesting overlap between viral genome dsDNA
cell density of 5 × 105 cells per milliliter before decreasing at and transcripts of viral structural proteins (SI Appendix, Fig. S4D).
48 hpi during the onset of the lytic phase (Fig. 1D). The first in- Average psbA intensity in the mcp+/psbA+ subpopulation was
crease in extracellular concentration of virions occurred at 8 hpi higher than in mcp–/psbA+ cells at 8 and 24 hpi (SI Appendix, Fig.
before reaching a plateau of 5.3 × 108 virions per milliliter at S5). This could reflect either enhanced psbA transcription in in-
56 hpi (Fig. 1E). In order to quantify infection dynamics at a fected cells or down-regulation of psbA translation leading to ac-
single-cell resolution in a high-throughput manner, smFISH cumulation of mRNA, as shown for some plant viruses (31),
samples were probed for mcp and psbA, stained with DAPI, and although transcription and translation can be decoupled (32). At
acquired using a multispectral imaging flow cytometer (Image- the population level in the first 12 h, despite identical cell counts
StreamX). In the infected culture, mcp+ cells represented 19% of between noninfected and infected cultures, mean cell chloro-
the population at 4 hpi, 75% at 24 hpi, and plateaued at 60% phyll intensity decreased in the infected culture suggesting
between 32 and 48 hpi before declining (Fig. 1F). Host psbA ex- chlorophyll degradation, coherent with the general loss of pho-
pression showed that 98% of the cells in the noninfected culture tosynthetic transcripts (SI Appendix, Fig. S6). Higher psbA ex-
remained psbA+, in contrast with a sharp drop to 12% in the pression in the mcp+ population emphasizes the interdependence
infected cultures between 8 and 24 hpi (Fig. 1F). Infected cells, between optimality of infection (mcp expression) and host pho-
defined by their mcp+ signal, showed large variability in the tosynthetic regulation (psbA expression).
amounts of mcp mRNA per cell, reflected by a wide dynamic Based on intracellular virocell dynamics, we quantified fun-
range of intensity values of the mcp probe per cell (from 103 to 105 damental parameters in the life cycle of a giant marine virus. We
arbitrary units [a.u.] of fluorescence), suggesting cell-to-cell het- assumed that all cells encounter an infectious viral particle
erogeneity in levels of viral infection (Fig. 1G). Simultaneously, we within 30 min after infection as experiments were conducted at
detected a bimodal response in the infected culture with high and high virus:host ratio (SI Appendix, Table S1). We further con-
low psbA expression (threshold at 1.1 × 103 a.u. fluorescence) sider the mcp mRNA as a strong signal for active viral infection.
indicating the loss of photosynthetically active cells in a distinct We estimate that the eclipse phase, the time elapsed between
subpopulation (Fig. 1H). smFISH therefore enables dual quanti- successful cell infection and the initiation of virus produc-
fication of host and viral genes during infection dynamics across tion (33), occurs within the first 4 h, based on the induction of
thousands of single cells. mcp+/psbA+ cells, without an increase in virion concentration (Fig.
To investigate E. huxleyi virocell heterogeneity in infected 2C). The onset of the maturation phase, during which viral progeny
cultures, we plotted host and virus mRNA expressions in parallel are released, occurs within 8 h as suggested by previous studies
and revealed four distinct coexisting subpopulations (Fig. 2A) (34). The distinct infection states can be associated with unique cell
that we quantified throughout the course of infection (Fig. 2B fates: photosynthetically active infected cells (mcp+/psbA+) are
and SI Appendix, Fig. S1A). At 0 and 1 hpi, most of the cells were responsible for early viral release, most likely through budding (34,
mcp–/psbA+ (green gate), indicating photosynthetically active 35), while mcp+/psbA– likely contribute less to this process. Vi-
cells (Fig. 2A, 0 hpi). Cells from noninfected cultures appeared rion egress from the host cells without induction of cell death may
in this gate at all time points. At 4 hpi, 24% of the cells were both provide an optimal coexistence mechanism during bloom initia-
mcp+ and psbA+ (red gate, Fig. 2A, 4 hpi), coexpressing viral tion phase (36). The mcp–/psbA– subpopulation correlated with
and host genes. Two new subpopulations were clearly distin- Sytox Green staining at later time points (SI Appendix, Fig. S1B),
guishable at 24 hpi: 60% of the cells were mcp+/psbA– (yellow suggesting that, after 56 hpi, a substantial fraction of the cells
gate) as a consequence of psbA transcription shutdown or rapid contain compromised cell membranes and induce cell death
psbA mRNA degradation; 20% of the cells were mcp–/psbA– without releasing viruses. These cells can die from accumulation
(gray gate, Fig. 2A, 24 hpi). This subpopulation can originate of DNA damage (37), exposure to cytotoxic viral glycosphingolipids
from mcp–/psbA+ cells experiencing gradual photosynthetic (38), or abortive infection. The mcp–/psbA– and mcp–/psbA+ cells
shutdown, illustrated by the large heterogeneity in psbA intensity might serve as the seed for the resistant phenotypes, often ob-
within the mcp–/psbA+ subpopulation or from mcp+/psbA– cells served in a small subpopulation that recovers from viral infection
that interrupted mcp transcription. We further compared these in the so-called “Cheshire cat” escape strategy (39). The transition
four subpopulations with single-cell RNA-Seq data (22) and between different phenotypic states could be mediated by pro-
show that they corresponded to distinct stages of viral progression, duction of infochemicals, such as viral glycosphingolipids (38, 40),
as defined by the temporal succession of viral gene expression (SI or by extracellular vesicles (41) (Fig. 2G).
Appendix, Fig. S2). Intriguingly, viral release increased between The consequences of viral infection in the ocean are not
4 and 8 hpi, when infected cells were observed only in the limited to the fate of the individual host cell. Viral infection can
mcp+/psbA+ subpopulation (Fig. 2C). In parallel, algal cell abun- also lead to aggregate formation, mediated by production of trans-
dance remained stable, and only a small fraction of cells displayed parent exopolymer particles (TEP) consisting of acidic polysaccha-
membrane permeability based on Sytox+ staining, indicating the rides, which are classically quantified in bulk by staining with Alcian
absence of cell lysis (Fig. 2D). This suggests that photosynthetically Blue (42). Aggregates accelerate sinking of phytoplankton biomass,
2 of 8 | PNAS Vincent et al.
https://doi.org/10.1073/pnas.2021586118 Visualizing active viral infection reveals diverse cell fates in synchronized algal bloom
demise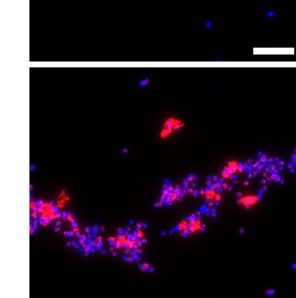
A Algae sampling Probing mRNA in single cells using smFISH markers Quantitative vizualisation
psbA mRNA A
AAA
AAAAA A A
Epifluorescence
psbA microscopy
Vacuole
Lab cultures Host chloroplast mRNA
EhV
+
AA
Fixation
A
mcp Nucleus
AA
mcp mRNA AA
A A
AA
Late viral mRNA Imaging
100 km flow-cytometry
Natural algal Chloroplast
bloom
B C D
107
Cell abundance
Early Infection Late Infection
(cells mL-1)
1 hpi 24 hpi 106
****
105
Non infected culture
104 Infected culture
103
102
0 12 24 36 48 60 72 84 96 10 8 12 0
psbA
Time (hpi)
E 8 Non infected culture
(108 virions mL-1)
Infected culture
6
****
EhV 4
MICROBIOLOGY
2
0
0 12 24 36 48 60 72 84 96 10 8 12 0
Time (hpi)
F
psbA+ or mcp+ cells
100
mcp
80 mcp+ cells (%)
**** Non infected
60 Infected
(%)
40 psbA+ cells (%)
**** Non infected
20
Infected
0
0 8 16 24 32 40 48 56 64 72 80
Time (hpi)
G H
56 hpi 56 hpi
DAPI
48 hpi 48 hpi
Normalized cell frequency
Normalized cell frequency
32 hpi 32 hpi
24 hpi 24 hpi
8 hpi 8 hpi
4 hpi 4 hpi
All
1 hpi 1 hpi
0 hpi 0 hpi
mcp- mcp+ psbA- psbA+
101 103 105 101 103 105
mcp fluorescence intensity (a.u.) psbA fluorescence intensity (a.u.)
1 10 102103104105 1 10 102103104105
Fig. 1. High-throughput visualization of active viral infection in algal virocells using mRNA smFISH. (A) Simplified workflow of sample processing and data
acquisition. After initial fixation, samples were hybridized to custom-made fluorescent probes targeting the host (psbA) and the virus (mcp) mRNA and were
subjected to epifluorescence microscopy (high resolution) and imaging flow cytometry (high throughput) analyses. (B and C) Epifluorescence images of an
infected culture of E. huxleyi cells at 1 and 24 hpi (early and late infection, respectively) in each channel. (Scale bar: 20 μm.) In the DAPI channel, the Inset
images depict segmented nuclei (see SI Appendix, Fig. S12 for a full picture). (D and E) Cell and virion concentrations, respectively, of infected and noninfected
cultures during infection of E. huxleyi cells by EhV. Infection was quantified using flow cytometry at a high virus:host ratio. (F) Fraction of mcp+ and psbA+
cells in infected (dashed line) and noninfected (solid line) cultures. Values are presented as the mean ± SD, n = 3. ****P < 0.0001 tested with linear mixed
model fit by REML. T-tests use Satterthwaite’s method. (G and H) Distribution of mcp and psbA fluorescence intensity values in E. huxleyi single cells acquired
by ImageStreamX during a time course of viral infection. The dashed line depicts the intensity threshold used to define mcp+ and psbA+ cells according to the
fluorescent intensity (103 a.u.).
Vincent et al. PNAS | 3 of 8
Visualizing active viral infection reveals diverse cell fates in synchronized algal bloom https://doi.org/10.1073/pnas.2021586118
demise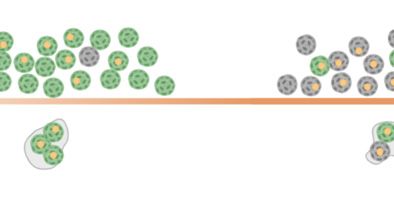
A 0 hpi 4 hpi 8 hpi 24 hpi B
0.0% 0.0% 0.2% 23.9% 5.0% 48.8% 60.1% 13.6%
mcp intensity (a.u.)
Population (%)
104 75
104 104 104 mcp-/psbA-
50 mcp-/psbA+
'
mcp+/psbA-
103 103 103 103 mcp+/psbA+
25
0 0 0 0
8.5% 91.5% 5.3% 70.6% 7.4% 38.8% 19.8% 6.5%
0
0 103 104 105 0 103 104 105 0 103 104 105 0 103 104 105 0 1 4 8 24 32 48 56 72 80
psbA intensity (a.u.) Time (hpi)
C mcp+/psbA-
D Chl+ Cells
E F G mcp-/psbA+ mcp+/psbA+
mcp-/psbA- mcp+/psbA-
mcp+/psbA+ Sytox+ Cells Photosynthetic Photosynthetic
mcp-/psbA+ mcp+/psbA+
60 4 100 Non infected Infected
24 hpi AAAAAAAAAA
A
AAAAAAAAAA
A Virions
Normalized cell frequency
A
AAAAAAAAAA
**** psbA mRNA
EhV(108 virions mL-1)
50
80 V AAAAAAA AA
Population (%)
AA
3 mcp mRNA AAA
AA
Population (%)
AAAAA
A
V
40 Permeable
60 8 hpi
V membrane
30 2 V
****
40
20 AAAAAAA AA
4 hpi AA
Viral lysis
1 20
10 C ****
0 0 0 mcp-/psbA- mcp+/psbA-
0 4 8 12 0 4 8 12 3*103 3*104 3*105 Non photosynthetic Non photosynthetic
Time (hpi) Time (hpi) DAPI Intensity (a.u.) Infected
Fig. 2. Heterogeneity in transcriptional states during viral infection reveals distinct potential cell fates. (A) To investigate the heterogeneity of E. huxleyi
virocells during infection, we plotted host and virus mRNA coexpression per cell at different time points of hours postinfection. The x axis represents the value
of the probe intensity (in fluorescent arbitrary units) targeting the psbA host gene, and the y axis represents the value of the probe intensity targeting the
viral mcp gene. Using a threshold of ∼10^3 a.u. of fluorescence to define both mcp+ (y axis) and psbA+ (x axis) cells, we define four subpopulations as a
combination of mcp and psbA signals: mcp–/psbA+ (green gate), mcp+/psbA+ (red gate), mcp+/psbA– (yellow gate), and mcp–/psbA– (gray gate) (B) Relative
abundance of the four subpopulations throughout the course of infection. (C) Virion production (blue) and mcp+ dynamics in the first 12 hpi (mcp+/psbA+ in
red, mcp+/psbA– in yellow). (D) Fraction of cells in the population with high chlorophyll signal (Chl+) and percentage of cells with permeable membranes
(Sytox+). Values are presented as the mean ± SD; n = 3. (E) Transmission electron microscopy of an infected E. huxleyi cell at 4 hpi. White arrowheads indicate
budding viruses. C: chloroplast; V: immature intracellular viruses. (F) Comparison of DNA content based on DAPI intensity between the four subpopulations
(with more than 25 events per population) throughout infection. Comparison of DAPI intensity was performed with a linear mixed model fit by REML. T-tests
use Satterthwaite’s method. ****P < 0.0001. (G) Conceptual scheme of the potential cell fates of different subpopulations.
contribute to “marine snow,” and enhance carbon export to the experiment in a fjord near Bergen, Norway, and collected sam-
deep sea (3). In order to assess the contribution of viral infection ples for smFISH analysis across different phases of an E. huxleyi
to aggregate formation, we quantified aggregated E. huxleyi cells bloom. We quantified viral infection specifically in E. huxleyi host
and characterized the infection state of individual cells within the cells by using a 28S ribosomal probe unique to that alga, a psbA
aggregates using the smFISH approach (Fig. 3 A and B). Fol- probe as a proxy for host photosynthetic state, and a probe tar-
lowing infection, up to 40% of the algal cells (DAPI-positive geting viral mcp transcripts (Fig. 4A). smFISH analysis conducted
fraction) were found within aggregates (Fig. 3C). At the early by imaging flow cytometry successfully revealed the fraction of
stage of infection, less than 10% of the cells are found in ag- infected E. huxleyi virocells during bloom succession, based on
gregates, mostly infected (75% of the aggregates contained at over 5,000 cells at each time point (SI Appendix, Fig. S7). From
least one infected cell thereafter named mcp+) and photosyn- the initiation of the bloom to the peak of E. huxleyi abundance
thetically active (100% were psbA+) (Fig. 3 D and E). At the on day 17 (Fig. 4B), only a slight viral increase was observed
onset of the lytic phase (48 hpi), aggregates were highly abun- (Fig. 4C), and infection level was minimal (
A B BF mcp SSC DAPI psbA
C Aggregates (%)
D mcp+ aggregates (%)
E psbA+ aggregates (%)
50 100
Single cells
20 40 75 ****
75
30
Doublets **** 50 ****
15 20 50
DAPI circularity
25
10
25
10 Non-infected aggregate 0 0
BF mcp SSC DAPI psbA 0 8 16 24 32 40 48 56 64 72 80 0 8 16 24 32 40 48 56 64 72 80 0 8 16 24 32 40 48 56 64 72 80
Aggregates
Time (hpi) Time (hpi) Time (hpi)
5
Non infected culture Infected culture
0
0 50 100 150 200 250 Infected aggregate
DAPI area
F
Fig. 3. Quantification of viral infection within aggregates formed during infection. (A) Based on the DAPI mask area and circularity (the degree of the mask’s
deviation from a circle), three populations are identified as single cells, doublets, and aggregates (aggregates are defined by a DAPI area above 60 μm2). A
representative image of each population is presented in the Inset. (B) Imaging flow-cytometry images of noninfected (Top) and infected (Bottom) aggregates
MICROBIOLOGY
in the brightfield (BF), mcp (yellow), Side Scatter (SSC as a proxy for calcification, pink), DAPI (blue), and psbA (red) channels. (C) Fraction of aggregates of all
DAPI+ events in infected (dashed line) and noninfected (solid line) cultures. (D and E) Quantification of mcp+ and psbA+ aggregates, respectively, throughout
the time course of infected (dashed line) and noninfected (solid line) cultures. In C–E, values are presented as the mean ± SD; n = 3; ****P < 0.0001 (t test). (F)
Schematic representation of active infection in single cells versus aggregates throughout progression of viral infection. Illustration produced with Biorender.
bloom demise at a later stage. These top-down regulators may meet the high demand for nucleotides by the giant viruses and
include pathogenic bacteria (46), eukaryotic parasites, and grazers enhance the antioxidant capacity under oxidative stress (52).
(3, 47, 48). Alternatively, highly synchronized viral infection and Thus, a functional convergence arises among viruses infecting pho-
release might have occurred in a large fraction of the E. huxleyi totrophs from diverse evolutionary origin, whereby cyanophages (53),
population in a narrow time frame between days 17 and 18. In- small RNA plant viruses (54), and large algal dsDNA viruses
deed, recent reports demonstrate that viruses are intrinsically share the ability to manipulate the metabolism of their photosyn-
synchronized with the daily rhythms of their photosynthetic host thetic host. Quantifying the virocell states, through their diverse
both in controlled laboratory experiments and in natural pop- phenotypes, will help integrate viral-dependent processes into eco-
ulations (49, 50). Additionally, we probed a late viral gene and system models by providing necessary parameters of infection dy-
potentially might detect a higher fraction of infected cells by namics, such as the fraction of actively infected cells, resistant cells,
using an early viral gene probe. Furthermore, the synchronized and the nature of sinking biomass (55). Finally, this quantitative
bloom demise can be explained by increasing cell aggregation at smFISH approach can be expanded to diverse host-pathogen and
later infection stages (Fig. 3F). host-symbiont systems with important ecological significance, add-
ing a mechanistic dimension to the field of microbial ecology.
Discussion
Our approach opens up avenues to study specific host–virus in- Materials and Methods
teractions among a complex microbial consortium in the marine Culture Growth and Viral Infection. The noncalcifying E. huxleyi strain CCMP
environment. It combines high-throughput morphological and 2090 was used for this study. Cells were cultured in K/2 medium with anti-
single-molecule transcriptional data, thus providing quantitative biotics (ampicillin and kanamycin) and incubated at 18 °C with a 16:8 h
subcellular localization of ribosomal and messenger RNA, of light–dark illumination cycle. A light intensity of 100 μmol photons m−2·s−1
was provided by cool-white light-emitting diode lights. All experiments started
both hosts and viruses simultaneously, at single-virocell resolu-
with exponential phase cultures (5 × 105 cells mL−1). The virus used for this
tion. Our approach further enables detection of aggregates and
study is EhV201 propagated on CCMP2090 with antibiotics. Five days before
small subpopulations, connects intra- and extracellular dynamics, infection, most probable number (MPN) assays were performed to assess the
and can be adapted to target viruses and virocells in mixed fraction of infectious viruses in the stock (48). E. huxleyi was infected with 5:1
natural populations by designing probes based on the wealth of multiplicity of infection (MOI) ratio of infectious virus per cell, thereby
available environmental genomics data. guaranteeing that all cells encountered an infectious particle 30 min postin-
In this study, we unravel fundamental aspects of the life cycle fection. The time courses of infected and noninfected cultures were sampled
of a large virus by linking scales between single-cell and population- simultaneously in triplicates. A second MPN experiment was performed on
level dynamics, showing viral increase without cell lysis in the the day of the experiment with the same virus and same algae to assess the
mcp+/psbA+ subpopulation, and cell lysis without viral increase in exact MOI at the beginning of the experiment (SI Appendix, Table S1).
the mcp–/psbA– subpopulation. The high-resolution examination
Enumeration of Algal Cell Abundance, Cell Death, and Viral Abundance. Cells
of host and virus transcriptional states revealed cell-to-cell het-
were monitored and quantified using an Eclipse (iCyt) flow cytometer. Cells
erogeneity in viral infection, mediated by the dependence on the
were identified by plotting the chlorophyll fluorescence (excitation [ex]:
host photosynthetic state, despite synchronized bloom demise. 488 nm and emission [em]: 663 to 737 nm) versus side scatter. For extracellular
Indeed, viral replication may require photosynthetic activity in viral counts, samples were stained with SYBR gold (Invitrogen) that was di-
order to energize the redirection of carbon flux from the Calvin luted 1:10,000 in Tris–ethylenediaminetetraacetic acid buffer, incubated for
cycle to the pentose phosphate pathway. This will increase pro- 20 min at 80 °C, and cooled to room temperature (RT). Samples were ana-
duction of NADPH required for de novo DNA synthesis (51) to lyzed by an Eclipse flow cytometer (ex: 488 nm; em: 500 to 550 nm), and a
Vincent et al. PNAS | 5 of 8
Visualizing active viral infection reveals diverse cell fates in synchronized algal bloom https://doi.org/10.1073/pnas.2021586118
demise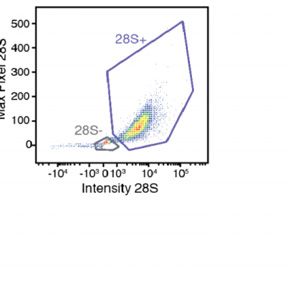
A Tracking viral infection dynamics in mixed natural B C D
phytoplankton populations using smFISH 20 30
E.huxleyi cells (104 cells mL-1)
8
Infected cells (% mcp +)
Virions (106 VLP mL-1)
16
6 20
12
4
8
10
2 4
E.huxleyi ribosomal probe
0 0 0
11 13 15 17 19 21 23 11 13 15 17 19 21 23 11 13 15 17 19 21 23
Time (days) Time (days) Time (days)
E mcp+ cells F H
BF 28S mcp SSC DAPI psbA Combined mcp+/psbA+ mcp+/psbA-
mcp- cells
mcp-/psbA- mcp-/psbA+ mcp-/psbA-
Day 22
mcp+/psbA- 100
Normalized Cell Frequency
Day 21 mcp+/psbA+
80
Population (%)
Day 20 mcp-/psbA+
60
Day 19
G 40
BF 28S mcp SSC DAPI psbA Combined
Day 18
Non infected 20
Day 17 aggregate
0
Infected 11 12 13 14 15 16 17 18 19 20 21 22 23
3*104 105 3*105 106
aggregate
DAPI intensity (a.u.) Time (days)
Fig. 4. Visualization of active viral infection during a natural E. huxleyi bloom. (A) Applying smFISH to track E. huxleyi virocells within mixed natural mi-
crobial populations. A probe specifically targeting the 28S ribosomal RNA of E. huxleyi was used to identify our host of interest, based on the intensity and
maximum pixel of the probe signal in flow-cytometry plots, along with the mcp and psbA probes. (B and C) Abundance of calcified E. huxleyi cells and EhV-
like particles (VLP), respectively, measured by flow cytometry throughout bloom succession in a mesocosm experiment. (D) Fraction of infected (mcp+) E.
huxleyi virocells, measured by smFISH analysis throughout bloom succession. (E) Comparison of DAPI intensities between infected and noninfected E. huxleyi
populations throughout bloom succession. (F and G) Visualizing subpopulations of E. huxleyi single cells and aggregates, respectively, using ImageStreamX
(SSC: side scatter). (H) Quantification of the relative abundance of the four subpopulations in single cells throughout the bloom succession by coprobing of
host and viral mRNA (psbA and mcp, respectively).
minimum of 50,000 events was collected. To analyze cell permeability, solution (prepared in 1× phosphate-buffered saline containing 4% para-
samples were stained with a final concentration of 1 μM Sytox Green (Invi- formaldehyde and 30% sucrose), transferred to a 1.7 mL Eppendorf tube,
trogen), incubated in the dark for 1 h at RT, and analyzed by an Eclipse flow and incubated for 1 h at 4 °C with agitation. Tubes were then centrifuged
cytometer (ex: 488 nm; em: 500 to 550 nm). An unstained sample was used as for 2 min at 4 °C and 3,000 × g, the supernatant was removed and stored at
a control to eliminate the background signal. The data were exported and 80 °C until hybridization.
analyzed in R. A day before ImageStreamX acquisition, selected samples were thawed
and chlorophyll was extracted by a first wash using 900 μL of 70% ethanol
mcp and psbA Probe Design and Conjugation for smFISH. The smFISH tech- applied for 3 min, followed by centrifugation for 3 min at 3,000 × g and
nique identifies a single mRNA based on the binding of multiple small probes removal of the supernatant. A second wash was performed with 900 μL of
targeted to different locations to the mRNA of interest. Fasta sequences of 100% ethanol applied for 3 min, followed by centrifugation for 3 min at
the target genes mcp and psbA were first submitted to Stellaris Probe De- 3,000 × g and removal of the supernatant. Samples were treated with 500 μL
signer to obtain potential probes. We designed 48 probes per gene with a Proteinase K at 10 μg/mL final concentration (Ambion #AM2546) for 10 min
probe length of 20 nucleotides. Probes with more than 70% guanine– at room temperature and washed with 3 min at 3,000 × g centrifugation.
cytosine content were discarded. To discard off-targets and decrease non- Samples were then resuspended in 50 μL of hybridization buffer (17.5%
specific signal, each probe sequence was blasted against the E. huxleyi formamide concentration) containing equal concentrations of the different
transcriptome and EhV201 genome. Probes with an off-target gene matching target genes at a 0.1 ng/mL final concentration. Mcp probes were conju-
over 17 nucleotides were discarded. A minimum of 20 probes per gene is gated with TMR, and psbA probes were coupled to Cy5. Hybridization was
necessary to detect a single molecule. Validated probes were ordered with 3′ performed overnight in 30 °C shakers.
amine groups through the Custom Oligo Service of BioSearch Technologies The day of ImageStreamX acquisition, hybridization buffer was washed
(Probe_Sequences.xlsx). Fluorophores with succinimidyl ester group were or- away by centrifugation for 3 min at 3,000 × g. Samples were stained for
dered from Click Chemistry Tools: Tetramethylrhodamine (TMR) and sulfo- dsDNA with 500 μL of GLOX buffer (prepared in nuclease-free water with
Cyanine5 (Cy5). Probes were coupled to fluorophores and purified as de- 0.4% final concentration of glucose, 2× final concentration sodium
scribed in ref. 23. All the conjugated probes of a given gene were used as a chloride–sodium citrate (SSC) Ambion #AM9765, and 10 mM final concen-
mixture each time that we targeted the expression of that gene. Therefore, tration of Tris, pH 8.0) with DAPI in 10 μg/mL final concentration (except the
the mcp mRNA was detected using 47 probes of 20 nucleotides each, and the single stains). DAPI staining was done for 30 min in 30 °C, followed by
psbA mRNA was detected using 48 probes of 20 nucleotides each. The position centrifugation for 3 min at 3,000 × g, removal of supernatant, and resus-
of the probes on the reference gene are shown in SI Appendix, Fig. S9 A and B, pension in 40 μL of GLOX buffer before being acquired in the Image-
and their respective positions and sequences are indicated in Dataset S1. StreamX. DAPI was used for several reasons. The first one is technical: DAPI is
required for the fluorescent microscope to localize cells when the brightfield
Sample Fixation and Hybridization for smFISH in Laboratory Samples. At each channel is not available. We used DAPI to draw the contour of the nucleus
time point, 50 mL of each flask was fixed in a cold 1% paraformaldehyde final using Cell Profiler in Fig. 1 and SI Appendix, Fig. S12. DAPI is also important
concentration and incubated for 1 h at 4 °C with gentle agitation. Each for gating of the cells in the imaging flow cytometer. Gating on brightfield
sample was then centrifuged for 2 min at 4 °C and 3,000 × g. The super- signal and on the DAPI achieves better detection of the cells and thus better
natant was discarded, the pellet resuspended in 1 mL of cryopreservant downstream statistics. The second major reason to use DAPI is that it
6 of 8 | PNAS Vincent et al.
https://doi.org/10.1073/pnas.2021586118 Visualizing active viral infection reveals diverse cell fates in synchronized algal bloom
demise
provides important information regarding the general DNA content of the washed in ASW and post fixed in 2% osmium tetroxide, 0.5% potassium
cells, its cell-cycle phase, and is a nucleus marker that serves as a reference dichromate, and 0.5% potassium hexacyanoferrate in ASW for 1 h at room
point in our cells. temperature, washed again, and stained in bloc with 2% aqueous uranyl
The following day, samples were resuspended in 300 μL of GLOX with the acetate for 1 h followed by ethanol dehydration. Samples were infiltrated
3,3′-dihexyloxacarbocyanine iodide membrane stain at final concentration with increasing concentrations of Epon EMBED 812 (EMS) and polymerized
of 1 μM (ThermoFisher, catalog number D273). Excessive dye was removed at 60 °C. Thin sections (∼70 nm) obtained with an Ultracut UCT microtome
by centrifugation for 3 min at 3,000 × g and removal of supernatant, fol- (Leica Microsystems) were poststained with 2% uranyl acetate and Reynold’s
lowed by resuspension in 4 μL GLOX. A total of 2 μL was deposited on a lead citrate and examined using an FEI Tecnai T12 TEM operating at 120 kV.
microscope slide with 2 μL of antibleach solution, and the sample was im- Images were recorded on an FEI Eagle 2Kx2K CCD camera.
aged with the epifluorescent microscope.
Single-Cell Transcriptomics Comparison. Based on single-cell dual transcriptomics
Imaging Flow Cytometry Acquisition, Compensation, and Analysis for Laboratory during E. huxleyi infection by EhV201 (22), we plotted gene expression of psbA
Samples. Samples were acquired using the ImageStreamX MarkII machine (ISX, (SI Appendix, Fig. S13A) and mcp (SI Appendix, Fig. S13B) in available cells (n =
Amnis, Luminex). Three excitation wavelengths were used: 405 nm (DAPI: 384) across different time points postinfection. For each gene, we plotted the
channel 7 to 50 mW), 561 nm (TMR: channel 3 to 200 mW), and 642 nm (Cy5: abundance of the gene’s mRNA molecules per cell (Left), followed by the rel-
channel 11 to 120 mW). For each sample, at least 50,000 cells were acquired and ative expression of that gene with respect to the total number of mRNA
imaged with 60× magnification. Single stained samples were acquired using molecules in the cell (Center), and, finally, the percentage of cells expressing
the compensation wizard of the ImagestreamX. On average, less than 10 μL of that specific gene (Right). Furthermore, we were able to map the single-cell
2 × 106 cell stock concentration was necessary to collect the optimal amount of transcriptomics infection states (metacell) into each of the smFISH transcriptional
cells, except toward the end of the experiment where cells were scarce. states that were based on psbA versus mcp coexpression on a single-cell resolution
Data were analyzed using IDEAS6.2 (Amnis, Luminex). The compensation (SI Appendix, Fig. S2). The MetaCell method uses the k-nearest neighbor graph
matrix was built using the IDEAS wizard and manually checked before being partitions (60) for organizing single cells into cohesive groups of cells (metacells)
applied to all the acquired files. Based on the area (the number of microns with similar viral gene expression profiles and is defined in our system in ref. 22.
squared in a mask) and circularity (the degree of the mask’s deviation from a
circle) of DAPI, three populations were identified as single cells (mainly, DAPI Mesocosm Experimental Setup. The mesocosm experiment—Aquacosm-viral-induced
area 60 a.u.). Single
microbial succession (Aquacosm-VIMS)—was carried out over 24 d (May 24 to June
cells were additionally selected in the same focal plane using the BF gradient
MICROBIOLOGY
17, 2018) in Raunefjorden at the University of Bergen’s Marine Biological
and contrast (both gradient and contrast measure the sharpness quality of
Station at Espegrend, Norway (60.27° N; 5.22° E). The experiment consisted
an image by detecting large changes of pixel values in the image). All gates
of seven enclosure bags made of transparent polyethylene (11 m3, 4 m deep,
were defined on a single file before being applied to the total data set (SI
and 2 m wide; 90% photosynthetically active radiation) mounted on floating
Appendix, Fig. S10). Each file was then manually inspected to check the
frames and moored to a raft in the middle of the bay. Each bag was filled
accuracy of single-cell and aggregate gating. All the data (fluorescent in-
with surrounding fjord water and supplemented with nutrients at days 0 to
tensities, morphological features, populations) were then exported for each
7 and 13 to 17 at a nitrogen:phosphorous ratio of 16:1 (1.6 μM NaNO3, 0.1
cell of each file for analysis in R. ImagestreamX source data are available on
μM KH2PO4). On days 6, 7, and 13 only NaNO3 was added. The water in each
Dryad (https://doi.org/10.5061/dryad.h44j0zpjc) (56). Compensation matrixes
bag was continuously mixed by pumping air to the bottom of each bag.
are available in .cif files, and analysis of each time point in .daf files.
Samples for flow cytometric counts were taken twice a day, morning (7 AM)
Similarity was calculated in the IDEAs software. As defined in the IDEAs
and evening (8 to 9 PM), using 50 mL centrifugal tubes and following fil-
User Manual (ref. 57, p. 215), “The Similarity feature is the log-transformed
tration using a 40 μm cell strainer (61). Calcified E. huxleyi were identified
Pearson’s correlation coefficient and is a measure of the degree to which
using the Eclipse flow cytometer based on high side scattering and high
two images are linearly correlated within a masked region.” In SI Appendix,
Fig. S4D, we correlate DAPI and mcp masks for each cell of two subpopu- chlorophyll content (SI Appendix, Fig. S14). All bags converged to a similar
lations (mcp+/psbA+, mcp+/psbA–) and plot its distribution. The threshold pattern of an E. huxleyi bloom and demise, and the current study focuses on
used to declare colocalization was similarity >1.2. bag 4.
RNase Treatment. E. huxleyi cultures were infected with EhV201 at low MOI smFISH Specificities for Field Samples. Sample fixation, storage, and initial
and fixed in 1% paraformaldehyde for smFISH 24 hpi. Two samples were chlorophyll washes of field samples were identical to laboratory samples. In
used to compare mcp signal with and without RNase treatment. The sample order to identify E. huxleyi cells, we designed a single probe specific to the
with RNase treatment was treated with RNase before the Proteinase K 28S region of E. huxleyi (62) (EG28‐03, 5′‐TAAAGCCCCGCTCCCGGGTT‐3′,
treatment, with RNase A from Thermo Fisher (catalog no. EN0531) at a final bound to C3-Fluorescein (ex/em = 490/525 nm). Two helper probes were used
concentration of 10 μg/mL in a reaction volume of 200 μL of infected E. (Helper A, 5′-GCCAGGACGGGAGCTGGCCG-3′ and Helper B, 5′-GAGGCGCGG-
huxleyi culture for 20 min at 37°. Both samples were then processed as usual CGCCGAGGCGC-3′) to facilitate unfolding of the 28S rRNA molecule by
for mcp staining and acquired in the ImageStreamX. Application of RNase binding to the flanking regions of the probe of interest EG28; 28S, HelperA,
treatment on infected samples shows heavily reduced mcp signal, confirm- HelperB were used at a final concentration of 0.5 μM at the same time as the
ing that the probes bind to RNA and not to DNA (SI Appendix, Fig. S11) mcp and psbA probes at 0.1 ng/mL final concentration. We further checked
the efficiency of our mcp probes to detect the most abundant EhV isolated in
Epifluorescence Microscope Acquisition and Analysis. Slides were acquired the mesocosm, EhVM1. Specifically, of the 47 mcp probes designed to target
using a Nikon inverted fluorescence microscope Eclipse Ti2 Series and imaged the laboratory viral strain EhV201, 34 probes had a 100% match to EhVM1,
with a Ixon Ultra 888 camera with 100× magnification, using the Nikon NIS which is sufficient to obtain high fluorescent signal. The positions of the mcp
Element Advanced Research Software. Illumination time for each excitation probes on the mcp gene of EhVM1 are shown in SI Appendix, Fig. S9C. EhV201
was adjusted at the beginning of each batch acquisition and not modified and EhVM1 shared above 96% identity over the 1,491-bp alignment, sug-
after that. Each image is composed of 15 0.3-μm stacks. Images were exported gesting that our probes are efficient in capturing the most abundant EhV of
in .nd2 format and inspected in Fiji 2 (58). Stacking projection was performed our natural samples. In order to test whether or not our EhV mcp probes could
for each file (Image > Stacks > Z-Project > MaxIntensity) on a maximum of 10 bind against the mcp sequence of another phycodnaviridae large virus
stacks. Mean intensity in the background was measured for each channel infecting microeukaryotic algae, we mapped the probes against the mcp gene
(Analyze > Measure) and subtracted (Process > Math > Subtract). of the Ostreococcus tauri giant virus OtV (SI Appendix, Fig. S9D). Only four
To estimate the fraction of infected cells, we used CellProfiler (59) using probes were mapped, with less than 17 nucleotides identity. All samples were
the PercentPositive pipeline separately for each probe after a maximum probed in 50 μL of 40% formamide hybridization buffer, incubated at 37°
projection and background subtraction for each channel separately (SI Ap- overnight. Samples were acquired using the ImageStreamX MarkII machine
pendix, Fig. S12). (ISX, Amnis, Luminex). Four excitation wavelengths were used: 405 nm for the
DAPI (DAPI: channel 7 to 50 mW), 488 nm for 28S rRNA (AF488: channel 2 to
TEM. E. huxleyi CCMP2090 was infected with 1:50 volumetric ratio of viral 200 mW), 561 nm for mcp (TMR: channel 3 to 200 mW), and 642 nm for psbA
lysate to culture a MOI of 1:1 viral particles per cell. At 4 hpi, 500 mL of (Cy5: channel 11 to 120 mW).
culture was collected (8,000 × g, 10 min, 20 °C), resuspended in fixation
media (2% glutaraldehyde, 4% paraformaldehyde, 2% acrolein in artificial Data Availability. All ImagestreamX data are available on Dryad (https://doi.
sea water [ASW]), and fixed for at least 24 h at 4 °C. The cells were then org/10.5061/dryad.h44j0zpjc).
Vincent et al. PNAS | 7 of 8
Visualizing active viral infection reveals diverse cell fates in synchronized algal bloom https://doi.org/10.1073/pnas.2021586118
demise
ACKNOWLEDGMENTS. We thank S. Itzkovitz, L. Farack, and S. Ben-Moshe for reading; and R. Avraham for useful critical feedback. This work was supported
guidance in smFISH protocols, analysis, and fruitful discussions; D. Hirsch for by the European Research Council CoG VIROCELLSPHERE Grant 681715 (to
training on the epifluorescence microscope; the viral induced microbial A.V.) and the Israeli Academy for Science and Humanities and Weizmann Dean
succession–Emiliania huxleyi (VIMS-EHUX) consortium for the mesocosm ex- of Faculty (F.V.). The mesocosm experiment was supported by AQUACOSM
periment; all A.V. laboratory members for discussion; G. Schleyer for critical (VIMS-EHUX, 2018). A.V. is The Bronfman Professorial Chair of Plant Science.
1. F. Azam, Microbial control of oceanic carbon flux: The plot thickens. Science 280, 32. Y. Taniguchi et al., Quantifying E. coli proteome and transcriptome with single-
694–696 (1998). molecule sensitivity in single cells. Science 329, 533–538 (2010).
2. C. A. Suttle, Marine viruses: Major players in the global ecosystem. Nat. Rev. Micro- 33. S. J. Flint, V. R. Racaniello, L. W. Enquist, A. M. Skalka, Principles of Virology: Mo-
biol. 5, 801–812 (2007). lecular Biology (Wiley, 2015).
3. C. P. Laber et al., Coccolithovirus facilitation of carbon export in the North Atlantic. 34. L. C. M. Mackinder et al., A unicellular algal virus, Emiliania huxleyi virus 86, exploits
Nat. Microbiol. 3, 537–547 (2018). an animal-like infection strategy. J. Gen. Virol. 90, 2306–2316 (2009).
4. T. Fenchel, Marine plankton food chains introduction: The classical view of plankton 35. B. Knowles et al., Temperate infection in a virus-host system previously known for
food chains. Annu. Rev. Ecol. Syst. 19, 19–38 (1988). virulent dynamics. Nat. Commun. 11, 4626 (2020).
5. C. B. Field, M. J. Behrenfeld, J. T. Randerson, P. Falkowski, Primary production of the 36. S. Yau et al., Virus-host coexistence in phytoplankton through the genomic lens. Sci.
biosphere: Integrating terrestrial and oceanic components. Science 281, 237–240 Adv. 6, eaay2587 (2020).
(1998). 37. K. D. Bidle, L. Haramaty, J. Barcelos E Ramos, P. Falkowski, Viral activation and re-
6. Y. M. Bar-On, R. Phillips, R. Milo, The biomass distribution on Earth. Proc. Natl. Acad. cruitment of metacaspases in the unicellular coccolithophore, Emiliania huxleyi. Proc.
Sci. U.S.A. 115, 6506–6511 (2018).
Natl. Acad. Sci. U.S.A. 104, 6049–6054 (2007).
7. Y. Lehahn et al., Decoupling physical from biological processes to assess the impact of
38. A. Vardi et al., Viral glycosphingolipids induce lytic infection and cell death in marine
viruses on a mesoscale algal bloom. Curr. Biol. 24, 2041–2046 (2014).
phytoplankton. Science 326, 861–865 (2009).
8. S. W. Wilhelm, C. A. Suttle, Viruses and nutrient cycles in the sea. Bioscience 49,
39. M. Frada, I. Probert, M. J. Allen, W. H. Wilson, C. de Vargas, The “Cheshire Cat” escape
781–788 (1999).
strategy of the coccolithophore Emiliania huxleyi in response to viral infection. Proc.
9. J. S. Weitz, S. W. Wilhelm, Ocean viruses and their effects on microbial communities
Natl. Acad. Sci. U.S.A. 105, 15944–15949 (2008).
and biogeochemical cycles. F1000 Biol. Rep. 4, 17 (2012).
40. G. Schleyer et al., In plaque-mass spectrometry imaging of a bloom-forming alga
10. J. R. Brum et al., Patterns and ecological drivers of ocean viral communities. Science
during viral infection reveals a metabolic shift towards odd-chain fatty acid lipids.
348, 1261498 (2015).
Nat. Microbiol. 4, 527–538 (2019).
11. E. T. Sieradzki, J. C. Ignacio-Espinoza, D. M. Needham, E. B. Fichot, J. A. Fuhrman, Dy-
41. D. Schatz et al., Communication via extracellular vesicles enhances viral infection of a
namic marine viral infections and major contribution to photosynthetic processes shown
by spatiotemporal picoplankton metatranscriptomes. Nat. Commun. 10, 1169 (2019). cosmopolitan alga. Nat. Microbiol. 2, 1485–1492 (2017).
12. M. Moniruzzaman et al., Virus-host relationships of marine single-celled eukaryotes 42. A. Vardi et al., Host-virus dynamics and subcellular controls of cell fate in a natural
resolved from metatranscriptomics. Nat. Commun. 8, 16054 (2017). coccolithophore population. Proc. Natl. Acad. Sci. U.S.A. 109, 19327–19332 (2012).
13. A. D. Tadmor, E. A. Ottesen, J. R. Leadbetter, R. Phillips, Probing individual environ- 43. X. Mari, M. E. Kerros, M. G. Weinbauer, Virus attachment to transparent exopoly-
mental bacteria for viruses by using microfluidic digital PCR. Science 333, 58–62 meric particles along trophic gradients in the southwestern lagoon of New Caledonia.
(2011). Appl. Environ. Microbiol. 73, 5245–5252 (2007).
14. N. Baran, S. Goldin, I. Maidanik, D. Lindell, Quantification of diverse virus populations 44. M. J. L. Coolen, 7000 years of Emiliania huxleyi viruses in the black sea. Science 333,
in the environment using the polony method. Nat. Microbiol. 3, 62–72 (2018). 451–452 (2011).
15. A. L. Pasulka et al., Interrogating marine virus-host interactions and elemental transfer 45. J. E. Lawrence, A. M. Chan, C. A. Suttle, Viruses causing lysis of the toxic bloom-
with BONCAT and nanoSIMS-based methods. Environ. Microbiol. 20, 671–692 (2018). forming alga Heterosigma akashiwo (Raphidophyceae) are widespread in coastal
16. E. Allers et al., Single-cell and population level viral infection dynamics revealed by sediments of British Columbia, Canada. Limnol. Oceanogr. 47, 545–550 (2002).
phageFISH, a method to visualize intracellular and free viruses. Environ. Microbiol. 15, 46. N. Barak-Gavish et al., Bacterial virulence against an oceanic bloom-forming phyto-
2306–2318 (2013). plankter is mediated by algal DMSP. Sci. Adv. 4, eaau5716 (2018).
17. J. M. Labonté et al., Single-cell genomics-based analysis of virus-host interactions in 47. A. Chambouvet et al., Diverse molecular signatures for ribosomally ‘active’ Perkinsea
marine surface bacterioplankton. ISME J. 9, 2386–2399 (2015). in marine sediments. BMC Microbiol. 14, 110–120 (2014).
18. D. M. Needham et al., A distinct lineage of giant viruses brings a rhodopsin photosystem 48. K. M. J. Mayers et al., Growth and mortality of coccolithophores during spring in a
to unicellular marine predators. Proc. Natl. Acad. Sci. U.S.A. 116, 20574–20583 (2019). temperate shelf sea (Celtic Sea, April 2015). Prog. Oceanogr. 177, 101928 (2019).
19. B. L. Hurwitz, S. J. Hallam, M. B. Sullivan, Metabolic reprogramming by viruses in the 49. R. Liu, Y. Liu, Y. Chen, Y. Zhan, Q. Zeng, Cyanobacterial viruses exhibit diurnal
sunlit and dark ocean. Genome Biol. 14, R123 (2013). rhythms during infection. Proc. Natl. Acad. Sci. U.S.A. 116, 14077–14082 (2019).
20. P. Forterre, The virocell concept and environmental microbiology. ISME J. 7, 233–236 50. F. O. Aylward et al., Diel cycling and long-term persistence of viruses in the ocean’s
(2013). euphotic zone. Proc. Natl. Acad. Sci. U.S.A. 114, 11446–11451 (2017).
21. S. Rosenwasser et al., Unmasking cellular response of a bloom-forming alga to viral 51. R. J. Puxty, A. D. Millard, D. J. Evans, D. J. Scanlan, Viruses inhibit CO2 fixation in the
infection by resolving expression profiles at a single-cell level. PLoS Pathog. 15, most abundant phototrophs on Earth. Curr. Biol. 26, 1585–1589 (2016).
e1007708 (2019). 52. U. Sheyn, S. Rosenwasser, S. Ben-Dor, Z. Porat, A. Vardi, Modulation of host ROS
22. C. Ku et al., A single-cell view on alga-virus interactions reveals sequential tran- metabolism is essential for viral infection of a bloom-forming coccolithophore in the
scriptional programs and infection states. Sci. Adv. 6, eaba4137 (2020).
ocean. ISME J. 10, 1742–1754 (2016).
23. A. Raj, P. van den Bogaard, S. A. Rifkin, A. van Oudenaarden, S. Tyagi, Imaging in-
53. L. R. Thompson et al., Phage auxiliary metabolic genes and the redirection of cyano-
dividual mRNA molecules using multiple singly labeled probes. Nat. Methods 5,
bacterial host carbon metabolism. Proc. Natl. Acad. Sci. U.S.A. 108, E757–E764 (2011).
877–879 (2008).
54. J. Zhao, X. Zhang, Y. Hong, Y. Liu, Chloroplast in plant-virus interaction. Front. Microbiol.
24. S. O. Skinner, L. A. Sepúlveda, H. Xu, I. Golding, Measuring mRNA copy number in
7, 1565 (2016).
individual Escherichia coli cells using single-molecule fluorescent in situ hybridization.
55. N. R. Record, D. Talmy, S. Våge, Quantifying tradeoffs for marine viruses. Front. Mar.
Nat. Protoc. 8, 1100–1113 (2013).
Sci. 3, 251 (2016).
25. W. H. Wilson et al., Isolation of viruses responsible for the demise of an Emiliania
56. F. Vincent, U. Sheyn, Z. Porat, D. Schatz, A. Vardi, Visualizing active viral infection
huxleyi bloom in the English Channel. J. Mar. Biol. Assoc. 82, 369–377 (2002).
reveals diverse cell fates in synchronized algal bloom demise. Dryad. https://doi.org/
26. U. Sheyn et al., Expression profiling of host and virus during a coccolithophore bloom
10.5061/dryad.h44j0zpjc. Deposited 15 February 2021.
provides insights into the role of viral infection in promoting carbon export. ISME J.
57. Luminex Corporation, IDEASÒ image data exploration and analysis software user’s
12, 704–713 (2018).
27. E. V. Koonin, N. Yutin, Origin and evolution of eukaryotic large nucleo-cytoplasmic manual. https://www.luminexcorp.com/download/amnis-ideas-software-user-man-
DNA viruses. Intervirology 53, 284–292 (2010). ual/. Accessed 8 March 2021.
28. A. K. Mattoo, J. B. Marder, M. Edelman, Dynamics of the photosystem II reaction 58. J. Schindelin et al., Fiji: An open-source platform for biological-image analysis. Nat.
center. Cell 56, 241–246 (1989). Methods 9, 676–682 (2012).
29. K. Thamatrakoln et al., Light regulation of coccolithophore host-virus interactions. 59. H. Hennig et al., An open-source solution for advanced imaging flow cytometry data
New Phytol. 221, 1289–1302 (2019). analysis using machine learning. Methods 112, 201–210 (2017).
30. S. Rosenwasser, C. Ziv, S. G. V. Creveld, A. Vardi, Virocell metabolism: Metabolic in- 60. Y. Baran et al., MetaCell: Analysis of single-cell RNA-seq data using K-nn graph par-
novations during host-virus interactions in the ocean. Trends Microbiol. 24, 821–832 titions. Genome Biol. 20, 206 (2019).
(2016). 61. F. Vincent et al., Data from “AQUACOSM VIMS-Ehux: Core data.” Dryad. https://doi.
31. K. Lehto, M. Tikkanen, J. B. Hiriart, V. Paakkarinen, E. M. Aro, Depletion of the org/10.5061/dryad.q573n5tfr. Accessed 5 March 2021.
photosystem II core complex in mature tobacco leaves infected by the flavum strain of 62. M. J. Frada, K. D. Bidle, I. Probert, C. de Vargas, In situ survey of life cycle phases of the
tobacco mosaic virus. Mol. Plant Microbe Interact. 16, 1135–1144 (2003). coccolithophore Emiliania huxleyi (Haptophyta). Environ. Microbiol. 14, 1558–1569 (2012).
8 of 8 | PNAS Vincent et al.
https://doi.org/10.1073/pnas.2021586118 Visualizing active viral infection reveals diverse cell fates in synchronized algal bloom
demiseYou can also read

















































