PROBOSCIDEA-HOMO INTERACTIONS IN OPEN-AIR LOCALITIES DURING THE EARLY AND MIDDLE PLEISTOCENE OF WESTERN EURASIA: A PALAEONTOLOGICAL AND ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

3. PROBOSCIDEA-HOMO INTERACTIONS IN OPEN-AIR LOCALITIES
DURING THE EARLY AND MIDDLE PLEISTOCENE OF WESTERN
EURASIA: A PALAEONTOLOGICAL AND ARCHAEOLOCIGAL
PERSPECTIVE
George E. Konidaris1,*, Vangelis Tourloukis1
1
Palaeoanthropology, Eberhard Karls University of Tübingen, Senckenberg Centre for Human Evolution and
Palaeoenvironment, Rümelinstr. 23, 72070 Tübingen, Germany
*
georgios.konidaris@uni-tuebingen.de
https://dx .doi.org/10.15496/publikation-55599
KEYWORDS | Palaeoloxodon; Mammuthus; carnivore guild; lithic technology; human evolution
ABSTRACT havioral approach, we examine the large carnivore
community structure and dynamics, with empha-
In recent years, a significant number of Pleistocene sis in the hunting strategies of large predators. Ad-
localities with evidence of proboscidean exploita- ditionally, we aim to infer their possible role in the
tion by humans has been discovered, substantial- changes of early human subsistence strategies fo-
ly enriching our knowledge on Homo subsistence cusing on proboscidean acquisition, and to explore
strategies and megafauna acquisition. In this study, the role of humans within the predatory guild. The
we provide a synthesis of the evidence for Pro- ecological adaptations of the two common Middle
boscidea-Homo interactions in Early and Middle Pleistocene proboscideans in Europe, the Europe-
Pleistocene open-air sites of western Eurasia with an straight-tusked elephant Palaeoloxodon antiquus
direct (e.g., presence of cut marks, proboscidean and the steppe mammoth Mammuthus trogonthe-
bone artifacts, fractures for marrow extraction) and rii, are also evaluated. Finally, we discuss various
indirect (e.g., association and refitting of lithic ar- aspects of the Homo bio-cultural evolution during
tifacts, use-wear analysis) evidence of exploitation. the period under study, including developments in
Sex and ontogenetic age of butchered proboscide- material culture and relevant inferences about hu-
ans are recorded, in order to assess possible human man social behavior.
preferences. Furthermore, we investigate the role
of large carnivores focusing on important renew-
als in the carnivore guilds, and their significance 3.1 INTRODUCTION
in terms of carrion availability for scavenging and
human-carnivore competition for access to food The last decades, a significant number of sites
resources. By applying an ecomorphological/be- with evidence of anthropogenic exploitation of
Konidaris, G. E., Barkai, R., Tourloukis, V., Harvati, K. (Eds.), Human-elephant interactions: from past to present.
Tübingen University Press, Tübingen 2021. http://dx.doi.org/10.15496/publikation-55604
68 GEORGE E. KONIDARIS, VANGELIS TOURLOUKIS
proboscideans has been discovered in western from ~1.3 Ma to ~120 ka (Eemian Interglacial),
Eurasia, dating to the Early and Middle Pleisto- thus covering the Lower and the early Middle Pa-
cene, considerably increasing our knowledge on laeolithic periods. We selected proboscidean single-
Homo subsistence strategies. Proboscideans, the and multi-carcass localities, as well as key locali-
largest terrestrial animals during the Pleistocene, ties where proboscideans constitute an important
were widely distributed on all continents (apart faunal element of the large mammal assemblage,
from Antarctica and Australia). Although they in which human presence is indicated by the oc-
constituted an ideal food package and therefore currence of human fossils, lithic/wooden artifacts
an attractive target for early humans (Ben-Dor and/or anthropogenic bone modifications. Their
et al., 2011; Reshef and Barkai, 2015; Agam and geographic position and chronology are shown
Barkai, 2016), their enormous size would have de- in Figures 3.1 and 3.2, and Appendix 3.1. For
manded the employment of special obtainment/ each locality the parameters that we recorded are:
exploitation methods and coordinated effort by a Marine Isotope Stage (MIS; glacial/interglacial);
group of people, as well as investment of energy proboscidean taxon; direct evidence of processing
and time (Lupo and Schmitt, 2016). Assessing (cut marks, breakage for brain/marrow extraction,
the human agency in proboscidean-bearing faunal proboscidean bone tools, weapons associated with
assemblages is not always straightforward. First, proboscidean skeleton); cut-marked/fractured
the stratigraphic association of proboscidean and skeletal element and purpose of modification;
cultural remains does not in itself necessarily im- indirect evidence (presence of lithic/wooden arti-
ply anthropogenic processing of carcasses and the facts, refitting of lithics, use-wear analysis, human
verification of their functional relation requires fossils); lithic technology; bone artifacts; gender
taphonomic analysis (e.g., Giusti et al., 2018; and ontogenetic age of the proboscidean individ-
Giusti, this volume). Second, hominin exploita- ual; presence of large carnivores; occurrence of
tion of carcasses can be difficult to demonstrate, carnivore gnawing and coprolites; and associated
because bone modifications can result from other large mammals (Appendix 3.1). In examining the
(non-human) agents, human-induced cut marks sites, we do not directly compare data on mortal-
are only rarely preserved, and possible subsequent ity patterns, body part representation, site tapho-
weathering or other natural processes may delete nomy and lithic or bone assemblages, because of
direct evidence on bone surfaces (e.g., Haynes and discrepancies and/or information shortage in the
Klimowicz, 2015). Despite these impediments, published datasets.
the Proboscidea-Homo interactions are relatively Several studies analyzing the ecomorphology
well recorded in the Pleistocene of western Eur- and guild structure of carnivores have been con-
asia. The objective of this study is to evaluate the ducted, mainly as a tool to infer palaeoenviron-
Proboscidea-Homo record in open-air continental mental conditions (Morlo et al., 2010 and refer-
localities during the Early and Middle Pleistocene ences therein), but also to investigate carnivore
of western Eurasia, and to assess emerging patterns communities during the Plio-Pleistocene and/
between ecological, ethological, environmental or correlate them with early human settlements
and cultural parameters. in Europe (e.g., Turner, 1992; Croitor and Bru-
gal, 2010; Palombo, 2016; Rodríguez-Gómez et
al., 2017). Here, we introduce a modified ver-
3.2 METHODS sion of the three-dimensional ecomorphological
approach of Morlo et al. (2010) in order to: 1)
We included in our study 35 open-air sites from examine the community structure and dynamics
western Eurasia, dated from the Early until the (with emphasis on hunting strategy) of the large
Middle/Late Pleistocene boundary, and ranging predatory guild of Europe during the Early and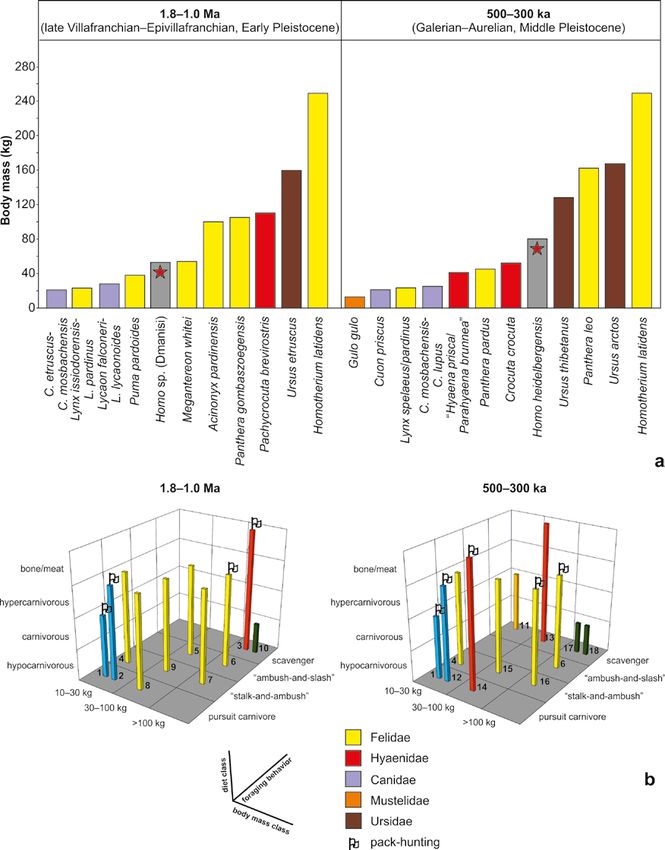
PROBOSCIDEA-HOMO INTERACTIONS IN OPEN-AIR LOCALITIES 69 Figure 3.1: Map showing the location of the studied open-air Early–Middle Pleistocene sites of western Eurasia with Proboscidea- Homo interactions (made with Natural Earth, naturalearthdata.com). Middle Pleistocene, 2) infer the possible role of larger than its size (MacDonald, 2009). We ex- large carnivores in the changes of early human cluded small-sized mustelids, felids (Felis) and ca- subsistence strategies (passive/active scavenging nids (Vulpes), with a weight ~20 kg (Carbone et al., 2007), that prac- thibetanus) and brown bear (U. arctos), because tice hunting and/or scavenging on large prey, with their diet includes ungulates, and both species are only exception the large-sized mustelid Gulo gulo reported to exhibit hunting and more commonly (wolverine), which although its average body mass scavenging behavior (MacDonald, 2009; Saladié is 10 kg), it preys on mammals much et al., 2013; Pappa et al., 2019 and references
70 GEORGE E. KONIDARIS, VANGELIS TOURLOUKIS
therein). We separated carnivores into two chro- 3.3.1. LATE EARLY PLEISTOCENE
nofaunas: 1.8–1.0 Ma (within Early Pleistocene) (~1.8–0.78 MA)
and 500–300 ka (within Middle Pleistocene); the
former includes the carnivores that were present The oldest-known Proboscidea-Homo event in Eu-
during the first human colonization of Europe, rope is attested at the Upper Archaeological Lev-
and the latter involves a time when human pres- el of Fuente Nueva-3 (Spain), with an estimated
ence is well recorded in various localities almost age of ~1.3 Ma (1.19 ± 0.21 based on U-series/
throughout the continent. The estimated body ESR dating; 1.50 ± 0.31 Ma from cosmogenic
mass (BM) of carnivores is taken from Meloro et nuclides; Espigares et al., 2013; this volume and
al. (2007), Palmqvist et al. (2011), Hemmer et al. references therein). The discovery of a partial
(2011), Van Valkenburgh et al. (2016) and Rodrí- skeleton of the southern mammoth Mammuthus
guez-Gómez et al. (2017). Three BM classes were meridionalis, in association with lithic artifacts of
defined, keeping the large-sized classes of Morlo “Mode 1” character and coprolites of the hyena
et al. (2010): 1. 10–30 kg, 2. 30–100 kg, and 3. Pachycrocuta brevirostris was interpreted as pos-
>100 kg. Diet was classified into four categories sible competition for scavenging between Homo
based on Van Valkenburgh (1988) and Morlo et and Pachycrocuta (Espigares et al., 2013; this vol-
al. (2010): 1. hypocarnivorous (includes the om- ume). At Barranc de la Boella (Spain; 960–781
nivores, 70% meat), and 4. bone/meat senting a butchering event (Vallverdú et al., 2014;
(>70% meat with the addition of bones). Diet Mosquera et al., 2015).
data were modified from Palombo (2016). Four
foraging behavior groups (carcass acquisition strat-
egy) have been distinguished following Werdelin 3.3.2. MIDDLE PLEISTOCENE (0.78–0.12 MA)
(1996): 1. pursuit carnivore, 2. “stalk-and-am-
bush”, 3. “ambush-and-slash”, and 4. scavenger. In Gesher Benot Ya’aqov (Israel; 780 ka, MIS 19)
Hunting strategy data were modified from Palom- a cranium of Palaeoloxodon antiquus (or perhaps P.
bo (2016). Sociality is distinguished into: 1. social recki) was discovered in association with Acheule-
(group/pack-hunting), and 2. solitary, acknowl- an lithic artifacts; a basalt core, a boulder and an
edging, however, the flexibility of fission/fusion oak log that were found below the cranium, were
sociality. Sociality data were acquired from Treves possibly used to invert it. The upside-down posi-
and Palmqvist (2007) and Palombo (2016). The tion of the cranium, the missing basicranial and
dataset is given in Table 3.1. palatal regions, and the damage below the nasal
opening, were attributed to possible deliberate
brain extraction and trunk removal (Goren-Inbar
3.3 THE RECORD OF PROBOSCIDEA- et al., 1994). A similar case may also be represent-
HOMO INTERACTIONS ed in Notarchirico (Italy; 670–610 ka, MIS 16).
In this locality, a cranium of P. antiquus was lying
In this section we briefly present some key localities, in an overturned position, lacking the masticato-
where more systematic studies on Proboscidea-Ho- ry apparatus and the occipital, and the mandible
mo interactions have been conducted, but the read- was found some meters away, indicating possible
er is referred to Appendix 3.1 and the references anthropogenic utilization of the brain, the tongue
cited therein for more detailed accounts, as well as and the trunk (Piperno and Tagliacozzo, 2001).
for the whole set of sites considered in this study. The lithic assemblage includes bifaces, however,PROBOSCIDEA-HOMO INTERACTIONS IN OPEN-AIR LOCALITIES 71
Figure 3.2: Chronology of the
studied localities, biostratigra-
phic range of Homo, carnivores
and proboscideans, and miles-
tones in human evolution and
Proboscidea-Homo interactions
in western Eurasia.72 GEORGE E. KONIDARIS, VANGELIS TOURLOUKIS
the association between lithic artifacts and bones mals (Villa et al., 2005). In Southfleet Road (En-
is not clearly demonstrated (Moncel et al., 2019). gland; ~425–375 Ma, MIS 11), a P. antiquus par-
In Ficoncella (Italy; ~500 ka, MIS 13), the dis- tial skeleton was found without direct evidence of
covery of a P. antiquus partial skeleton bearing car- anthropogenic activity. However, the tight spatial
nivore gnawing and a small-tool lithic assemblage association between lithic artifacts and elephant
(with refits), indicates human occupation and in bones, as well as the lithic refits and edge dam-
situ knapping events, as well as carnivore access to age on some lithics, which probably resulted from
the carcass (Aureli et al., 2015). In Marathousa-1 on-site production and subsequent use as butch-
(Greece; 500–400 ka, probably close to MIS 12/11 ery tools, altogether indicate butchering activities
transition; Panagopoulou et al., 2018 and referenc- (Wenban-Smith, 2013). In Castel di Guido (Italy;
es therein), a partial skeleton of P. antiquus, and iso- ~412 ka, MIS 11), abundant remains of P. antiquus
lated elephant and other vertebrate remains were and other mammals (notably the auroch Bos pri-
found in spatial and stratigraphic association with migenius) were associated with Acheulean lithic
a small-tool lithic assemblage and bone artifacts artifacts; human bones were also discovered at the
(Konidaris et al., 2018; Tourloukis et al., 2018). site. The locality represents a complex palimpsest
Cut marks on the tibia and astragalus of the ele- formed by natural processes, human activities and
phant skeleton, as well as on isolated elephant ribs minor carnivore involvement (Boschian and Saccà,
(accompanied with peeling) and on other mammal 2010; Saccà, 2012). In addition to the exploitation
bones, indicate butchering activities (Konidaris of elephants for meat and marrow, elephant bones
et al., 2018). A bone percussor, most likely made served as raw material for shaping tools, including
from an elephant limb bone, is also documented at bifaces. In the levels of Terra Amata (France; MIS
this site (Tourloukis et al., 2018). 11), several P. antiquus remains were discovered
In Áridos 2 (Spain; ~380 ka, MIS 11), there is in association with lithic artifacts; in addition to
a strong human exploitation signal on the elephant red deer hunting and carcass transport, young el-
bones: here, remains of a P. antiquus were associat- ephant remains were also transported to the site
ed with lithic tools, and the presence of cut marks (Valensi et al., 2011).
on the scapula and on one rib suggest butchering In La Polledrara di Cecanibbio (Italy; ~325
activities. Their location on the bones indicates ka, MIS 9), a great amount of P. antiquus remains
defleshing and evisceration, the latter pointing to were discovered, accompanied by a diverse mam-
early access to the carcass by humans before car- mal fauna rich in Bos primigenius; additionally, a
nivores accessed it, leaving tooth marks and fur- deciduous premolar is attributed to Homo heidel-
rowing on the bones (Yravedra et al., 2010). In bergensis (Anzidei et al., 2012). Of great interest
Ambrona (Spain; >350 ka, MIS 11), large mam- are the remains of an elephant, which was possibly
mals, most notably P. antiquus, were identified in trapped in muddy sediments and exploited by hu-
several stratigraphic units. Of particular interest mans (Santucci et al., 2016). A rich lithic industry
is the AS3 unit, where several elephant specimens was produced at the site (refitting, knapping de-
were discovered, including a partial elephant skel- bris) and the use-wear analysis indicates butcher-
eton. Human modifications were not detected on ing activities. Human modifications on elephant
the skeleton; however, a cut mark was identified bones include intentional fractures and removals,
on another cranium, and two femoral shafts show as well as production of bone tools (Anzidei et al.,
anthropogenic fractures (Villa et al., 2005). The 2012; Santucci et al., 2016). In Revadim Quar-
taphonomic analysis indicated that Ambrona rep- ry (Israel; ~500?–300? ka) several specimens of P.
resents a combination of natural accumulations antiquus were found together with other faunal
and activities of humans, who regularly visited remains and a rich lithic assemblage. Among the
the site for exploiting elephants and other mam- elephant bones, one scapula and two ribs bear cutPROBOSCIDEA-HOMO INTERACTIONS IN OPEN-AIR LOCALITIES 73
marks indicative of filleting, and others have been and unique craniodental features, the giant hyena
shaped to tools (Rabinovich et al., 2012). The in- P. brevirostris was well adapted for dismembering
terpretation of butchering activities at Revadim is carcasses and consuming bones, and was the most
further supported by use-wear and fat residue anal- direct competitor of Homo for scavenging large
yses (Solodenko et al., 2015). mammal carcasses during the Early Pleistocene
In Poggetti Vecchi (Italy; MIS 7/6), wooden (Martínez-Navarro, 2010; Palmqvist et al., 2011).
(digging sticks), bone (including elephant ones) Pachycrocuta and Homo were highly dependent on
and lithic artifacts were found in association with flesh-eating predators, such as the saber-toothed
vertebrate remains; several P. antiquus individuals cats Megantereon whitei and Homotherium latidens,
possibly died due to a natural cause and were sub- which were well adapted to hunt, but possibly ate
sequently exploited by humans (Aranguren et al., mainly the soft parts of their prey (especially of
2019). The Layers 3 and 6.1 of La Cotte de St Bre- large carcasses like proboscideans), leaving behind
lade (England; MIS 6) preserve evidence for mega- food resources (leftovers) that could be afterwards
fauna exploitation (wooly mammoths and rhinos). scavenged (Turner, 1992; Martínez-Navarro, 2010;
Some mammoth bones show cut marks, while Palmqvist et al., 2011; see also Blumenschine, 1987
there exist also indications for brain extraction (rib for large herbivores and for proboscideans in par-
driven into the cranium) (Smith, 2015). In PRE- ticular). In particular, Homotherium groups were
RESA (Spain; MIS 6), bones of an elephantid in- able to hunt (having cursorial adaptations), dis-
dividual show, in addition to cut marks, green frac- articulate, transport and deflesh very large prey
tures and percussion damage indicative for marrow (~5,700 kg) compared to their own size, includ-
extraction (Yravedra et al., 2012, 2019). The ear- ing juveniles, adult female and young adult male
liest so far known evidence for the use of wooden proboscideans (Rawn-Schatzinger, 1992; Marean
weapons in proboscidean hunting dates close to and Ehrhardt, 1995; Hemmer, 2001; Palmqvist
the Middle/Late Pleistocene boundary and is at- et al., 2003, 2011; Van Valkenburgh et al., 2016;
tested at Lehringen (Germany; ~120 ka, MIS 5e), Barnett et al., 2020). But the large carnivore guild
where a wooden lance was discovered within a P. of the European Early Pleistocene was much more
antiquus skeleton, associated also with lithic arti- diversified, and apart from Pachycrocuta, Megan-
facts (Weber, 2000). tereon and Homotherium, included wolves (Canis
etruscus1-C. mosbachensis), wild dogs (Lycaon fal-
coneri-L. lycaοnoides), bears (Ursus etruscus), lynxes
3.4 DISCUSSION (Lynx issiodorensis), jaguars (Panthera gombaszoe-
gensis), giant cheetahs (Acinonyx pardinensis) and
3.4.1. CARNIVORE GUILDS AND HOMININS puma-like cats (Puma pardoides)2, each of them
equipped with great hunting, killing or scavenging
Shortly after the first “out of Africa” human disper-
sal, documented at Dmanisi (Georgia, ~1.8 Ma),
1 In Dmanisi the recently described Canis borjgali.
and contemporaneous with the first appearance of
2 The sympatry of these 10 large carnivores is recorded
humans in Europe (~1.3 Ma), the first association at Untermassfeld (Germany; Ursus cf. dolinensis instead of U.
of a proboscidean skeleton with lithic artifacts in etruscus) and these are present collectively in the various sedi-
mentary units of Pirro Nord (Italy). Other localities rich in large
western Eurasia is documented at Fuente Nueva-3 carnivores (≥7 species) include Dmanisi (Georgia), Apollonia-1
(~1.3 Ma), where a possible competition between (Greece), Venta Micena, Cueva Victoria, Vallparadís Estació (all
Spain), Ceyssageut and Vallonnet (both France); Cueva Victoria,
humans and Pachycrocuta brevirostris for scavenging Vallparadís Estació and Vallonnet with U. deningeri instead of
a mammoth skeleton was suggested (Espigares et U. etruscus. All sites include Pachycrocuta and saber-toothed
felids, and yielded also Mammuthus meridionalis; Pirro Nord in-
al., 2013). With a powerfully built body, mass near- cludes lithic artifacts; Dmanisi both lithic artifacts and human
ly twice that of the spotted hyena Crocuta crocuta remains.74 GEORGE E. KONIDARIS, VANGELIS TOURLOUKIS
capabilities, and dental specializations related to carnivores discovering them, and in lack of strong
their diet preferences; some of them were are also evidence for elaborate hunting weaponry or tech-
characterized by social foraging behavior (Fig. 3.3b, niques, we can assume, in particular for megafauna
Table 3.1). Therefore, as a member of the predato- carcasses, that: a) access of early Homo to carcasses
ry guild3, encompassing 10 large carnivores, early would have been possible mainly through passive
Homo (estimated BM: 40–65 kg, stature: 145–155 scavenging of an already consumed and abandoned
cm; based on Dmanisi specimens; Gallagher, 2013) carnivore kill, and perhaps more possibly through
would have been positioned below the median active (confrontational/kleptoparasitic) scavenging
of body mass of the carnivores (Fig. 3.3a), much (Blumenschine, 1987; Espigares et al., 2013; Madu-
smaller or nearly equal to 5 felids and Pachycrocuta, rell-Malapeira et al., 2017), and b) food acquisition
and only larger than the lynx and the canids; the and exploitation, carcass- and self-defense especial-
latter, however, are pack-hunters (allowing them to ly in the shadow of the fierce scavenger P. breviros-
kill prey much larger than their own size and larger tris would have been impossible without a certain
than what a single individual would succeed) and, degree of cooperation, social intelligence, and use
like the other large carnivores, also faster-running of “weapons”, even if these were still relatively sim-
than Homo. This means that early humans had to ple (see also Agam and Barkai, 2018). Indeed, the
confront and compete constantly, if they were to scenario of stone-throwing for intimidating other
regularly exploit animal resources. Although the carnivores was proposed for Dmanisi and Fuente
carnivores of that period occurred at low densities Nueva-3, based on the abundant cobbles associated
in Southern Europe, based on the low number of with mammal fossils (Espigares et al., 2013).
prey species (moderate herbivore biomass; Rodrí- From the end of the Early Pleistocene/begin-
guez and Mateos, 2018), we can assume that prime ning of Middle Pleistocene and until ~0.6 Ma, an
and undefended carcasses would have been rather important faunal turnover took place, which in-
rare (in particular the small- to medium-sized ones; volved the disappearance of several Villafranchian/
Blumenschine, 1987), considering also the high Epivillafranchian taxa and the arrival of new im-
diversity of large carnivorous/hypercarnivorous/ migrants (Galerian fauna), including also carni-
bone-cracking predators, particularly with Pachy- vores and proboscideans (Palombo, 2014). In pro-
crocuta (the most important agent of bone accu- boscideans, the latest appearance of Mammuthus
mulations during the Early Pleistocene of Eurasia; meridionalis is documented at ~900–800 ka, while
Martínez-Navarro, 2010) being present at the kill around that time the steppe mammoth M. trogon-
sites soon after the event, as it happens with recent therii and the European straight-tusked elephant
hyenas (Domínguez-Rodrigo, 2001; Van Valken- Palaeoloxodon antiquus appear in Europe, increas-
burgh, 2001). This would result in relatively high ing the number of proboscidean representatives.
competition for carcass acquisition, both among In carnivores, the most prominent event is the
carnivores, and between carnivores and humans. extinction of Pachycrocuta and the arrival of Cro-
Therefore, although early Homo could have taken cuta crocuta (“Crocuta crocuta event”). The last ap-
advantage of naturally died proboscideans before pearance of P. brevirostris is traced at ~800 ka, while
the oldest European records of C. crocuta are docu-
mented at ~900–800 ka (Palombo, 2014). The ex-
3 In the European Early Pleistocene, percussion and cut
marks, although occasional, thus suggesting a more opportu- tinction of P. brevirostris and the survival of C. cro-
nistic behavior, prove that meat, fat and marrow were integral cuta are possibly attributed to ecological reasons:
part of early human diet. Human modifications are recorded on
bones from a wide range of animal body sizes, including both the super specialist scavenger P. brevirostris could
slow and fast prey, which indicates a generalist behavior. In the not survive in the changing climatic conditions of
discussion that follows, our premise is that Early Pleistocene
Homo can be considered as a member of the large predatory
the Middle Pleistocene, in which the more general-
guild, to which we also refer as “carnivore guild”. ist and social hunting/scavenging C. crocuta couldPROBOSCIDEA-HOMO INTERACTIONS IN OPEN-AIR LOCALITIES 75 Figure 3.3: a, body mass distribution of large carnivores and Homo and, b, 3D guild structure diagrams of large carnivores, for 1.8–1.0 Ma and 500–300 ka in Europe. The numbering and the groups in (b) is according to Table 3.1.1, Canis etruscus-C. mosba- chensis-C. lupus; 2, Lycaon falconeri-L. lycaonoides; 3, Pachycrocuta brevirostris; 4, Lynx issiodorensis-L. pardinus; 5, Megantereon whitei; 6, Homotherium latidens; 7, Panthera gombaszoegensis; 8, Acinonyx pardinensis; 9, Puma pardoides; 10, Ursus etruscus; 11, Gulo gulo; 12, Cuon priscus; 13, “Hyaena prisca/Parahyaena brunnea”; 14, Crocuta crocuta; 15, Panthera pardus; 16, Panthera leo; 17, Ursus thibetanus; 18, Ursus arctos.
76 GEORGE E. KONIDARIS, VANGELIS TOURLOUKIS
Family Species Body mass Diet class Hunting Sociality
(kg) behavior
1.8–1.0 Ma (late Villafranchian–Epivillafranchian, Early Pleistocene)
1. Canidae Canis etruscus-C. mosba- 10–30 carnivorous pursuit carnivore pack
chensis
2. Canidae Lycaon falconeri-L. lycao- 10–30 hypercarnivorous pursuit carnivore pack
noides
3. Hyaenidae Pachycrocuta brevirostris >100 bone/meat scavenger pack
4. Felidae Lynx issiodorensis-L. 10–30 hypercarnivorous stalk-and-ambush solitary
pardinus
5. Felidae Megantereon whitei 30–100 hypercarnivorous ambush-and-slash solitary
6. Felidae Homotherium latidens >100 hypercarnivorous ambush-and-slash pack
7. Felidae Panthera gombaszoegensis >100 hypercarnivorous stalk-and-ambush solitary
8. Felidae Acinonyx pardinensis 30–100 hypercarnivorous pursuit carnivore solitary
9. Felidae Puma pardoides 30-100 hypercarnivorous stalk-and-ambush solitary
10. Ursidae Ursus etruscus >100 hypocarnivorous scavenger solitary
500–300 ka (Galerian–Aurelian, Middle Pleistocene)
11. Mustelidae Gulo gulo 10–30 carnivorous scavenger solitary
1. Canidae C. mosbachensis-C. lupus 10–30 carnivorous pursuit carnivore pack
12. Canidae Cuon priscus 10–30 hypercarnivorous pursuit carnivore pack
13. Hyaenidae “Hyaena prisca/Parahyaena 30–100 bone/meat scavenger solitary
brunnea”
14. Hyaenidae Crocuta crocuta 30–100 bone/meat pursuit carnivore pack
4. Felidae Lynx pardinus 10–30 hypercarnivorous stalk-and-ambush solitary
6. Felidae Homotherium latidens >100 hypercarnivorous ambush-and-slash pack
15. Felidae Panthera pardus 30–100 hypercarnivorous stalk-and-ambush solitary
16. Felidae Panthera leo >100 hypercarnivorous stalk-and-ambush pack
17. Ursidae Ursus thibetanus >100 hypocarnivorous scavenger solitary
18. Ursidae Ursus arctos >100 hypocarnivorous scavenger solitary
Table 3.1: Large carnivores with their ecomorphological/behavioral parameters included in the guild structure analysis (see Figu-
re 3.3b). Data were acquired from references cited in “3.2 Methods”.
adapt (Martínez-Navarro, 2010). The extinction of nas and hominins (Palmqvist et al., 2011). On the
Pachycrocuta might be additionally correlated with other hand, the disappearance of Pachycrocuta pos-
the disappearance of Megantereon, which resulted sibly offered the opportunity for larger amounts of
in a decrease of carrion available for scavengers. At available carrion, and for easier and early access to
~600–500 ka Panthera gombaszoegensis and Aci- carcasses by hominins, who would have been most
nonyx pardinensis also disappear, while the modern possibly outcompeted during the Early Pleistocene
pantherine cats, Panthera leo and Panthera pardus by the fierce giant hyena.
occur in Europe (Fig. 3.2). The two latter species When we compare the Early and the Mid-
exploited carcasses more intensively than Megante- dle Pleistocene carnivore guilds (Fig. 3.3a, b, Ta-
reon, which possibly resulted in the decrease of the ble 3.1), we see that: 1) their structure and dy-
carrion that would be left available for both hye- namics are different, and 2) the carnivore diversityPROBOSCIDEA-HOMO INTERACTIONS IN OPEN-AIR LOCALITIES 77
slightly increased to 11 species during the Middle there were less hypercarnivorous predators to com-
Pleistocene. It should be noted, however, that in pete with, and, on the other hand, less available
contrast to the Early Pleistocene, the sympatry of carrion; the latter would have resulted in fewer
all (or most of ) these predators is not recorded so scavenging opportunities and thereby could have
far anywhere during the Middle Pleistocene; on offered an ecological incentive towards more regu-
the contrary, the number of both predator species4 lar hunting. In contrast, the number of the bone/
and predator specimens in the archaeo-palaeonto- meat eating (bone-cracking) hyenas increased to
logical sites is rather low (see also Martínez-Navar- two (however, both of them were smaller than
ro, 2018); this is the case also for the proboscidean Pachycrocuta); as this was accompanied with the
localities examined here, for which the predator as- inclusion of the wolverine Gulo gulo and the in-
semblage is poor in most of them (Appendix 3.1). crease of the mostly scavenging ursids (although
In canids, the only difference involves the their diet relied only little on meat), the scavenging
“replacement” of Lycaon lycaonoides by the slight- behavior is altogether reinforced. The so far rare
ly smaller Cuon priscus (still within the same BM “Hyaena prisca/Parahyaena brunnea” (or Pliocrocuta
group), but with the same diet type and hunting perrieri; the taxonomy of this taxon is debated; see
strategy. The number of the hypocarnivorous ur- Palombo, 2014) takes over the scavenging niche
sids increased with the “more omnivorous” Ursus previously occupied by Pachycrocuta. Crocuta cro-
arctos and the “more herbivorous” U. thibetanus cuta employs cooperative strategies, and consider-
(van Heteren, 2011; Pappa et al., 2019). There are ing also the inclusion of Panthera leo (as well as of
two main changes in the Middle Pleistocene car- Homo; see below), cooperative foraging is in gen-
nivore guild, compared to that of the Early Pleis- eral reinforced during the Middle Pleistocene (see
tocene (Fig. 3.3b): 1) the decline of taxa with a also Croitor and Brugal, 2010).
hypercarnivorous diet (the bulk of carrion provid- Notwithstanding the complexities surround-
ers) and an ambush hunting strategy, and 2) the ing the Middle Pleistocene hominin phylogeny
increase in the number of taxa with a bone/meat and nomenclature (e.g., Roksandic et al., 2018),
diet, as well as in the representation of pack-hunt- for the sake of our discussion we use H. heidelber-
ing and scavenging behavior. With the disappear- gensis (sensu lato) as the (more or less) “represen-
ance of the solitary species Megantereon whitei, tative” taxon of early-middle Middle Pleistocene
Acinonyx pardinensis, Puma pardoides (all from hominins. When comparing body mass values in
the middle-sized group 30–100 kg) and Panthera the two carnivore guilds (Fig. 3.3a), the increased
gombaszoegensis, the number of felids significant- BM and stature of Homo heidelbergensis (estimated
ly decreased (overall from six to four), along with BM of males: 70–90 kg, stature of males: 170–180
the demise of “ambush-and-slash” hunters. Most cm; Gallagher, 2013) places humans at a higher
importantly, all these carnivores were hypercar- rank than that in the Early Pleistocene: besides the
nivorous and carcass providers (hunters), having hypocarnivorous ursids, only Panthera leo and Ho-
primary access to and control of carcasses; conse- motherium latidens lie above Homo (Fig. 3.3a). The
quently, their disappearance resulted in a decrease potential for successful confrontational scaveng-
of available carrion for scavengers. For hominins, ing, early access to and defense of carcasses against
this change would have a dual effect: on one hand, predators, would have been much higher than in
the Early Pleistocene, considering also some shifts
in hominin technology and subsistence strategies
4 Among the richest localities of this period in terms of large
predators are Lunel-Viel (France) with 7 species, Taubach (Ger- (including regular hunting from ~400–300 ka on-
many) with 6 species, and Arago III and Orgnac 3 (both France) wards) and an inferred increase in human social
with 5 species, all with human presence, and Taubach and Ara-
go III additionally with Palaeoloxodon and Mammuthus remains,
cooperation (see below). Therefore, within the car-
respectively. nivore guild, an overall more privileged position78 GEORGE E. KONIDARIS, VANGELIS TOURLOUKIS
can be assumed for hominins in the Middle as op- the exception perhaps of weakened individuals. It
posed to the Early Pleistocene. follows that humans would not regularly acquire
Large predators (skeletal remains, carnivore access to proboscidean carcasses from felid kills.
bone modifications or carnivore coprolites, the lat- Rather, they would likely take advantage of indi-
ter in most cases attributable to hyenas) are present viduals already dead or caught in natural traps, in
in 29 of the herein studied sites (83%), and 12 which case they would exercise either passive or
out of those (41%) preserve also carnivore mod- active scavenging, according to the degree of carni-
ifications on proboscidean bones (34% of all the vore interference. Alternatively, humans acquired
35 sites). Moreover, 7 out of these 12 sites (58%) carcasses by hunting, employing tactics such as
preserve also direct evidence (or possible direct ev- ambush hunting, hunting with traps or confron-
idence) of human exploitation (Appendix 3.1), in- tational encounters. However, we should note that
dicating a certain degree of carnivore-human com- humans take a high risk when approaching and try
petition for early access to proboscidean carcasses, killing an elephant, which renders proboscidean
and highlighting the important interference and hunting a challenging and dangerous procedure
crucial role of carnivores in the human-probosci- (Lewis, this volume).
dean interactions. At least in two cases, humans In this light, we suggest the following, as a
acquired early access: at Áridos 2, where cut marks working hypothesis. In the Early Pleistocene pred-
on the ventral side of a rib was attributed to evis- atory guild (Fig. 3.3b), humans would occupy the
ceration (which takes place at the early stages of ecological space that was “available” for a preda-
carcass consumption; see e.g., Blumenschine; 1986 tor with a 30–100 kg BM and a (mostly?) scav-
and Potts, 1988), and at Marathousa 1, where cut enging behavior, perhaps with a carnivorous or
and scrape marks on the ventral side of a rib are hypocarnivorous diet according to ecological cir-
accompanied by classical peeling (fresh breakage). cumstances and geographic setting. In the Middle
The vertebral column and the rib cage seem to Pleistocene guild, humans would occupy the niche
be the proboscidean skeletal locations, which are that was previously held by the saber-toothed cat
mostly gnawed by carnivores (Appendix 3.1). Car- Megantereon (see also Werdelin and Lewis, 2013
nivore marks in the thorax region in particular are and Egeland, 2014 for African examples) in the
related to its opening for the consumption of intes- group of predators with 30–100 kg BM. Similar to
tines and inner organs, which happens at the ini- Megantereon, humans could have a carnivorous to
tial feeding stages of recent and extinct lions and hypercarnivorous diet, but unlike the large solitary
spotted hyenas (Haynes, 2005; Diedrich, 2014). felid, the biological, technological, cultural and so-
Recent lions (Panthera leo), and presumably cial developments would have allowed humans to
also Homotherium, prey on proboscideans, but employ not only the “ambush-and-slash” hunting
preferentially on young individuals (MacDonald, strategy, for instance modified into a cooperative
2009; Power and Compion, 2009; van Valken- “ambush-and-spear” strategy (in accordance with
burgh et al., 2016). Considering the much larger the evidence for use of hunting spears during this
size of Palaeoloxodon antiquus and Mammuthus period), but also a number of other hunting tac-
trogontherii compared to recent Loxodonta afri- tics, including for example prey stalking or prey
cana, Homotherium would selectively target young impediment by driving animals into natural or an-
individuals in the Middle Pleistocene. However, thropogenic traps. The seizing of a niche previously
during this period, the Proboscidea-Homo record occupied by a large felid such as Megantereon and
is dominated by adult proboscidean individuals the incorporation of such hunting behavior made hu-
(see below); it can be safely assumed that felids mans fairly independent of erratic food sources from
would have managed to kill such large adult prey scavenging carnivore kills and allowed the provision-
only occasionally and with great difficulties, with ing of animal resources on a more regular basis.PROBOSCIDEA-HOMO INTERACTIONS IN OPEN-AIR LOCALITIES 79
3.4.2. PROBOSCIDEAN EXPLOITATION AND one could counter-argue that an annual catch rate
HOMININ BEHAVIOR of “only” one or two proboscideans, such as that
observed for the Mbuti hunter-gatherers [Ichika-
Rather than regular hunting of proboscideans, a wa (this volume); of the African forest elephant
more opportunistic subsistence behavior of Early Loxodonta cyclotis, which is much smaller than Pa-
Pleistocene European Homo is suggested also by re- laeoloxodon; see also Lewis (this volume) and Ya-
cent studies, based on estimates of carrying capac- suoka (this volume) for similar catch rates among
ity, resource availability, the competition intensity the BaYaka and Baka) provides an ethnographic
within the carnivore guild and the network anal- example of a “regular and systematic” foraging of
ysis of food webs (Rodríguez-Gómez et al., 2016; proboscideans, and then the question is whether a
Lozano et al., 2016; Rodríguez and Mateos, 2018; comparable rate (in the Early Pleistocene) would
see also Palombo and Cerilli, this volume; Rosell still create a signal that is archaeologically detect-
and Blasco, this volume). Human presence togeth- able today. Factors related to the structure of the
er with Mammuthus meridionalis is documented carnivore community (e.g., the ratio of flesh-eaters
at a number of localities during this period, such to carcass-destroyers, which largely determines car-
as Dmanisi, Pirro Nord, Barranco León and Sima cass availability; Turner, 1992), habitat traits, and
del Elefante (Spain). The latter sites lack so far evi- climatic parameters such as seasonality, would have
dence of proboscidean processing and it is not clear altogether conditioned the scale to which hominins
whether this is a real lack of human exploitation, would employ any of the strategies included with-
absence of cut marks due to the thickness of perios- in their range of capabilities: from opportunistic,
teal connective tissue and cartilage of the bones, or non-confrontational scavenging as an effective,
a result of low archaeological resolution (e.g., due low-risk and low-cost means of food gathering, to
to taphonomy and preservation). Thus, on the ba- more active scavenging, hunting, and any combi-
sis of the evidence at hand, during the Early Pleisto- nation of tactics within this spectrum of foraging
cene, the acquisition of megafauna in general, and options.
of proboscideans in particular, appears to have been Whatever the means of procurement in the
mainly occasional and sporadic. Aside from issues Early Pleistocene, carcasses were processed by
of differential preservation and research intensity, use of apparently non-specialized, core-and-flake
which undoubtedly mask our archaeological visi- tool-kits with few retouched blanks and an overall
bility, we identify two main reasons to explain the expedient character. Nevertheless, at Barranc de
low archaeological signal for Proboscidea-Homo la Boella, an Acheulean-like pick that was prob-
interactions in the Early Pleistocene: 1) The large ably shaped off-site and transported to the mam-
carnivore guild remained stable during 1.8–0.8 Ma moth-butchery locality, points to some degree of
and all these powerful carnivores (especially P. brevi- tool curation and foresight. Notably, at Fuente
rostris), constituted a prohibiting factor for human Nueva-3, one of the main aims of the debitage
access to large carcasses, including proboscideans, was the production of small flakes (flakes >30 mm
until ~0.8 Ma, when most of them became extinct are rare). The association of proboscidean carcass-
(Martínez-Navarro, 2010; Madurell-Malapeira et es with small-sized, simple flakes is observed also
al., 2017). 2) The social structure of human groups, in the Middle Pleistocene (see below) and the evi-
their technological means and the behavioral solu- dence from Fuente Nueva-3 may be reflecting the
tions that they employed to mitigate the risks from roots of a long-lasting trend, which remains largely
competition with other members of the predatory unexplored and unexplained. Barranc de la Boella
guild, were not converging towards the inclusion of (Pit 1 level 2) is so far the oldest locality, where
proboscidean exploitation as a regular and system- possibly cut-marked proboscidean bones are re-
atic component of their subsistence strategies. Yet, ported, as well as the oldest locality with possible80 GEORGE E. KONIDARIS, VANGELIS TOURLOUKIS
human modifications on proboscidean remains tools in proboscidean butchery experiments (e.g.,
outside of Africa. Schick and Toth, 1993; Gingerich and Stanford,
In the Middle Pleistocene, lithic technology 2018). In carcass processing activities, simple and/
associated with proboscidean carcass processing or small flakes appear to entail two main advan-
mainly involved possible heavy-duty (core-)tools, tages over bifaces: 1) they allow for better preci-
such as choppers, chopping tools and bifacial im- sion (e.g., Venditti et al., 2019) and 2) they yield
plements (including handaxes and other Large higher return rates when raw material economics
Cutting Tools), as well as flake-based retouched are considered, mainly because a smaller mass of
tools, but also minimally modified blanks (usu- raw material is needed for their production; as
ally flakes) and cores (Appendix 3.1). However, they also provide more cutting edge per unit of
the artifact class most commonly found together mass, tool-kits based on small-sized blanks offer
with proboscidean remains is simple, unretouched economic advantages, especially to groups that
flakes of variable sizes and morphologies, together are highly mobile (Galán and Domínguez-Rodri-
with flake fragments and debris. Use-wear and res- go, 2014; Pargeter and Shea, 2019). Taphonomic
idue data show that both large(r) tools, such as bi- biases aside, those advantages alone could partly
faces or core-tools, and retouched or unretouched explain the higher frequency of small flake blanks,
flakes, often of small size (400 kg and an even higher weight is ex-
from experimental studies, which have shown that, pected for P. antiquus, its full exploitation should
while handaxes may be more efficient at specific have required from the Lower Palaeolithic humans
tasks such as defleshing, simple or retouched flakes the investment of a significant amount of time and
can be equally efficient at other tasks, such as disar- energy (Reshef and Barkai, 2015), knowledge of
ticulation (Galán and Domínguez-Rodrigo, 2014; its edible components, as well as social skills and
see also Jones, 1980). Moreover, both unmodified constructive cooperation among the group mem-
flakes and bifaces have been reported as efficient bers. Gesher Benot Ya’aqov is the oldest knownPROBOSCIDEA-HOMO INTERACTIONS IN OPEN-AIR LOCALITIES 81
locality with possible intentional breakage of a not discuss any possible functional use of blanks
proboscidean cranium. and tools made on bone fragments: depending on
Intentional bone fracturing for marrow ex- ecological and cultural contexts, some of them,
traction is reported much later, during MIS 11, al- such as percussors, scrapers or unmodified flakes,
though older assemblages with proboscidean bones could have been produced to serve functional
should be reassessed for this kind of exploitation. goals, while others were possibly manufactured
A recent study on P. antiquus limb bones from for non-utilitarian purposes. Zutovski and Barkai
Castel di Guido, showed that, although marrow (2016) proposed that proboscidean bone artifacts
cavities are proportionally small compared to the might additionally hint to cosmological, cultural
bones’ size, they do exist, and the fracturing of pro- and symbolic relations between proboscideans and
boscidean bones at this site involved not only bone humans. Fontana Ranuccio and Malagrotta (Italy;
tool fashioning, but also consumption of marrow both MIS 13) currently represent the oldest sites
for nutritional purposes (Boschian et al., 2019; see with evidence of bone tools made on proboscidean
also Anzidei et al., 2012: fig. 16e). Bone fractur- bones in western Eurasia (Fig. 3.2); other younger
ing for marrow extraction usually takes place at a localities include Castel di Guido, La Polledrara,
second stage of carcass exploitation, after the strip- Revadim, Casal de’ Pazzi, and perhaps Marathou-
ping of meat and the removal of fat, and possibly sa-1, Biltzigsleben and Vértesszölös (Hungary; but
when bones are still fresh, producing thus green see Fluck, 2011) (Appendix 3.1).
bone fractures (e.g., EDAR Culebro 1, PRERE- Overall, considering the limitations of preser-
SA); however, the time interval between these stag- vation of human-induced modifications in probos-
es is not easy to define archaeologically and would cidean bones, as already stated in the introduction,
depend on the environmental conditions (for the the presence in 22 (63%) of the studied localities
preservation of fresh bone and of edible marrow) of direct evidence of human exploitation can be
and on food availability (seasonality) (see also considered a relatively high number, almost dou-
Boschian et al., 2019). Some localities evidence a ble than that of direct carnivore gnawing in pro-
third processing stage, namely the bone tool pro- boscidean bones (34%; higher also than 41%, the
duction, which experimentally has proved to be percentage in sites with carnivore presence; see
feasible on both fresh and drier bones (Stanford et above), indicating the significant contribution of
al., 1981; Backwell and d’Errico, 2004); again, the humans in the accumulation and modification of
time interval from the previous stage is difficult to the bones, and overall in the formation and tapho-
ascertain. Taken together, brain exploitation and nomic history of the localities.
fracturing of bones are evident in 8 sites (23%), in-
dicating that proboscidean carcasses were import-
ant sources not only for meat and fat. 3.4.3. REMARKS ON PROBOSCIDEAN
Full exploitation of carcasses becomes evident PALAEOECOLOGY
with the manufacturing of proboscidean bone
artifacts. Indeed, proboscidean bone artifacts (or The Middle Pleistocene Proboscidea-Homo records
possibly attributed to proboscideans) are present are far from being equally divided between P. an-
in 11 sites (31%) —many of which yield also bone tiquus and Mammuthus (in particular M. trogon-
artifacts made from other mammal bones (e.g., therii, the widely distributed mammoth during the
equids, bovids). The use of proboscidean bones as Middle Pleistocene), and there is a clear dominance
raw material for the production of cultural objects of P. antiquus (26 sites in total5; 81% of the Middle
suggests that the exploitation of carcasses involved
behavioral aspects beyond those related to subsis- 5 In Stanton Harcourt the mammoth accounts for more than
half of the faunal assemblage and therefore the locality is inclu-
tence. In lack of micro-wear investigations, we can- ded here in the Mammuthus-bearing sites.82 GEORGE E. KONIDARIS, VANGELIS TOURLOUKIS Pleistocene ones with identification at genus lev- ponents under (at least relatively) temperate con- el), as well as of localities correlated with intergla- ditions. Importantly, climatic conditions in these cial stages (Appendix 3.1). The latter correlation settings would never, or only rarely, reach freezing can be attributed to the different ecological adap- temperatures, as indicated by the presence of bea- tations of these proboscideans, the environmental vers and hippos in the faunal lists. Thus, it seems preferences or tolerances of hominins, as well as that the environments that P. antiquus inhabited preservation biases. Mammuthus trogontherii is were also favorable settings for human occupation generally considered a steppe dweller and was a and subsistence (hunting/scavenging). Almost common faunal element of the glacial stages in all of the sites examined here occur in fluvial or Central Europe, adapted to open landscapes and lacustrine environments; these are known to be aridity, being less abundant in the more temperate nutritionally advantageous locations, but also conditions of Mediterranean Europe (Athanas- depositional regimes that foster archaeological siou, 2012). Recent dental micro- and macrowear preservation. studies indicate grass-dominated mixed-feeding preferences (Rivals et al., 2019 and references therein). Although European sites with skeletons 3.4.4. EXPLORING ASPECTS OF of M. trogontherii do exist during the Middle PROBOSCIDEAN ETHOLOGY AS Pleistocene, there is hardly any evidence of human INTERPRETATIVE TOOLS presence, which can be attributed to preservation biases, local extinctions of (small) human popula- Mammuthus meridionalis, M. trogontherii and P. tions, lack of adequate procurement strategies, or antiquus with mean BMs >9 tones (Larramendi, to the less favorable habitats. On the other hand, P. 2016) were by far the largest terrestrial animals of antiquus had wide and flexible ecological adapta- the Pleistocene terrestrial ecosystems of Europe, tions, as it was an inhabitant of mild humid, warm clearly surpassing other megaherbivores, such as to warm-temperate and moderately wooded to Hippopotamus and the rhino Stephanorhinus. How- wooded environments, but also of wooded grass- ever, direct evidence (in the form of cut marks) for lands or even rather arid grasslands (Palombo et the exploitation of these latter taxa in Europe is al., 2010). Palaeodietary studies indicate a dietary so far limited (Appendix 3.2), while although 22 plasticity, which included browsing, grazing and (63%) of the studied proboscidean localities yield- mixed feeding (Rivals et al., 2019 and references ed also hippos and/or rhinos, only 2 of them (9%) therein). In Northern and Central Europe, it oc- preserve also cut marks on these megaherbivores. curred during interglacial phases and apart from Moreover, in contrast to the 12 localities bearing some exceptions, it was generally absent from the cut marks on proboscidean bones, cut marks in intervening cold stages of open habitats, when it hippo bones exist so far only in 4 localities and was contracted to Southern Europe, which acted in rhino bones in 8 (including both open-air and as a refugium (Lister, 2004). In particular, most cave ones). For H. antiquus this could be possi- of the Proboscidea-Homo localities have yielded a bly attributed to its strongly aquatic life habits diversified fauna rich in medium- to large-sized going usually only sporadically outside of water herbivores, offering a wide prey spectrum for large bodies and feeding mainly on aquatic vegetation carnivores and humans (Appendix 3.1). A lot of (Palmqvist et al., 2003; Martínez-Navarro, 2010). them include Castor (beaver) and Hippopotamus, Even if we consider an occasional nocturnal feed- as well as a diversity of cervids and some also the ing activity on land, like the recent H. amphibius, rarer primate Macaca (macaque), indicating the its tracking down and hunting in the dark would presence of permanent freshwater bodies (river be particularly difficult and challenging for hu- or lake settings) and substantial woodland com- mans, especially considering that H. amphibius
PROBOSCIDEA-HOMO INTERACTIONS IN OPEN-AIR LOCALITIES 83
is a dangerous animal, responsible for numerous of other animals, and they are not aggressive ex-
human deaths annually (Eltringham in Hutchins cept when males are in musth (Lister in Hutchins
et al., 2004). Similarly, rhinos show aggressive be- et al., 2004).
havior and can be frightening animals to encoun- In light of the above, there are two addition-
ter, often chasing human intruders (especially the ally interesting outcomes from the assessment
black rhino Diceros bicornis); rhinos have also an of the Proboscidea-Homo open-air Early–Mid-
acute sense of smell, detecting human scent even at dle Pleistocene localities in western Eurasia: 1)
a distance of ~800 m, alarming them to ran away males prevail in the record, and 2) most of the
(Owen-Smith in Hutchins et al., 2004). sites involve subadult/adult proboscidean indi-
In contrast to the scanty evidence for hippo viduals6 (Appendix 3.1), corresponding to the
and rhino exploitation, the proboscidean ex- Type C (“selective mortality”) of Haynes (2017).
ploitation record is rather abundant (Appendix Despite the fact that adult males had markedly
3.1), even though the enormous size of elephants more robust body size and more powerful tusks
and the fact that they live in flocks constituted compared to females and juveniles, and thus
probably a prohibiting factor for the direct con- were more deterring, there are some possible ex-
frontation and hunting by both large carnivores planations of their higher percentage in butcher-
and humans. Firstly, it seems possible that human ing sites, related mainly to the elephants’ social
exploitation involved individuals that were al- organization: 1) The fact that elephants live in
ready killed by other predators or died by natural flocks is a prohibiting factor for predators (both
causes, or individuals that were vulnerable and/or carnivores and humans), which would have to
weakened by e.g., diseases, injuries or malnutri- face the protection and defensive behavior of
tion; the latter would usually stay close to water adults (MacDonald, 2009). Indeed, young el-
sources, abandoned from their flock (Cannell, ephants stay closely dependent on their mother
2014). Secondly, certain aspects in the behavior of in the first ten years, being additionally protect-
proboscideans and the way they modify the land- ed by the whole female-dominant group (Mac-
scape would have served to the advantage of hu- Donald, 2009). 2) In contrast to females, males
man foraging tactics (Haynes, 2006, 2012). For on puberty leave or are forced out of the family;
example, elephants repeatedly use known paths fully-grown adult males acquire a more nomad-
leading to water sources; this habit would have ic and solitary life, roaming either alone or in
allowed hominins to practice particular hunting loose groupings (Moss, 1988; Lister in Hutchins
strategies, including the use of natural traps, am- et al., 2004; MacDonald, 2009). Consequently,
bushes, use of spears and inflicting injuries, espe- it becomes more possible for males to enter an
cially on their vulnerable cushioned feet (Haynes, unfamiliar landscape, get into more difficult or
2006, 2012; Cannell, 2014; Agam and Barkai, dangerous situations and take higher risks when
2018; Lewis, this volume). Thirdly, male individ- roaming a more adventurous terrain, increas-
uals acquire a more solitary life after puberty (see ing also the proneness to be caught, injured or
below), which makes them more vulnerable and die in natural traps (Moss, 1988; see also Lister
perhaps an easier target, while specifically during and Agenbroad, 1994; Álvarez-Lao et al., 2009;
the musth period (when testosterone levels are Haynes, 2017; Pečnerová et al., 2017 for mam-
increased) combats between males (accompanied moth analogies). Studies on extant African ele-
by loud vocalizations and thus easy to be located phants show that particularly during dry seasons,
by humans) can potentially end with the death of adult males frequent more types of habitats than
one of the individuals (Lister in Hutchins et al.,
6 When the approximate ontogenetic age is known, we infer
2004). Lastly, elephants are not territorial, they do that these individuals did not die due to advanced age, consi-
not defend their range, which overlaps with that dering that P. antiquus lifespan was ~75 years.You can also read

















































