Population status and habitat preference of Crocodylus moreletii Duméril & Bibron, 1851 (Crocodilia: Crocodylidae) within the limits of two ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Herpetology Notes, volume 14: 55-62 (2021) (published online on 09 January 2021)
Population status and habitat preference of Crocodylus moreletii
Duméril & Bibron, 1851 (Crocodilia: Crocodylidae) within the
limits of two protected natural areas in southeastern Mexico
C. Iván Flores-Escalona1, Pierre Charruau1,*, Marco A. López-Luna2, Claudia E. Zenteno-Ruiz2,
Judith A. Rangel-Mendoza2, and Candelario Peralta-Carreta1
Abstract. Morelet’s crocodile (Crocodylus moreletii) has recovered from past overexploitation but it is necessary to continue
monitoring its status in the context of the sustainable harvest (ranching) planned for the species in Mexico. We obtained
new information on the population status of C. moreletii at the boundaries of two protected natural areas in southeastern
Mexico that serve as the basis for long-term monitoring. From September 2014–March 2018 we carried out nocturnal sampling
and captured crocodiles along four selected routes representing different water body types. For each crocodile observed we
collected information on the localization, environment and the individual. We observed a mean of 0.8 ± 0.3 crocodiles/km,
with significant differences detected among routes. We captured 54 crocodiles (sex ratio: 1.7 ♂ : 1 ♀). Except for subadults,
crocodiles were more frequently associated with medium flooded forest than with other types of vegetation. Adults were
observed at deeper sites than other age classes. Observation sites of yearlings have higher salinity than those of neonates,
juveniles, and adults. The results show a dynamic population with all size classes in a suitable habitat for its growth.
Keywords. Morelet’s crocodile, conservation, population ecology, environmental variables, Pantanos de Centla, Laguna de
Términos
Introduction al., 2010). Currently, wild populations are considered
stable in Mexico, and in 2010 the species was transferred
Morelet’s crocodile (Crocodylus moreletii Duméril
to Appendix II of the Convention on International Trade
& Bibron, 1851; Fig. 1) is one of three species of
in Endangered Species of Wild Fauna and Flora (CITES;
crocodilians present in Mexico. It is distributed on the
Sánchez-Herrera et al., 2011). In addition, this species is
coast of the Gulf of Mexico, from the low coastal areas
classified as low risk in the Red List of threatened species
of Tamaulipas State to the Yucatan Peninsula, Belize,
of the International Union for Conservation of Nature
and northern Guatemala (Platt et al., 2010). It is mainly
(IUCN; Platt et al., 2010) and it is a subject of special
associated with shallow freshwater bodies and the
protection within the Norma Oficial Mexicana NOM-
presence of abundant aquatic vegetation. Its importance
059-SEMARNAT-2010. Based on the recovery of the
is recognized at the ecological, economic, and cultural
populations in Mexico, a ranching protocol for Morelet’s
level (Casas-Andreu, 2002; Casas-Andreu et al., 2013).
crocodile in Mexico was developed, which aims to carry
The main threats to C. moreletii conservation are habitat
out the sustainable use of eggs of the species (Barrios
degradation and loss, as well as illegal hunting (Platt et
and Cremieux, 2018). However, it is important to
continue generating information about the populations
and monitor them over time to determine appropriate
measures for management plans (Sánchez-Herrera et al.,
1
Centro del Cambio Global y la Sustentabilidad, A.C., Calle
2011; Barrios and Cremieux, 2018).
Centenario del Instituto Juárez SN, Col. Reforma, 86080
Villahermosa, Tabasco, México.
The Pantanos de Centla Biosphere Reserve (PCBR)
2
Universidad Juárez Autónoma de Tabasco, División and the Laguna de Terminos Flora and Fauna Protection
Académica de Ciencias Biológicas, Carretera Villahermosa- Area (LTFFPA) are two important protected natural areas
Cárdenas km. 0.5 SN, Entronque a Bosques de Saloya, (PNA) in southeastern Mexico (Fig 2) with a combined
86150 Villahermosa, Tabasco, México. area of just
over one million hectares (Instituto Nacional
*
Corresponding author. E-mail: charruau_pierre@yahoo.fr de Ecología [INE], 1997; 2000). This large area allows for
© 2020 by Herpetology Notes. Open Access by CC BY-NC-ND 4.0. the protection of a wide diversity of plant communities,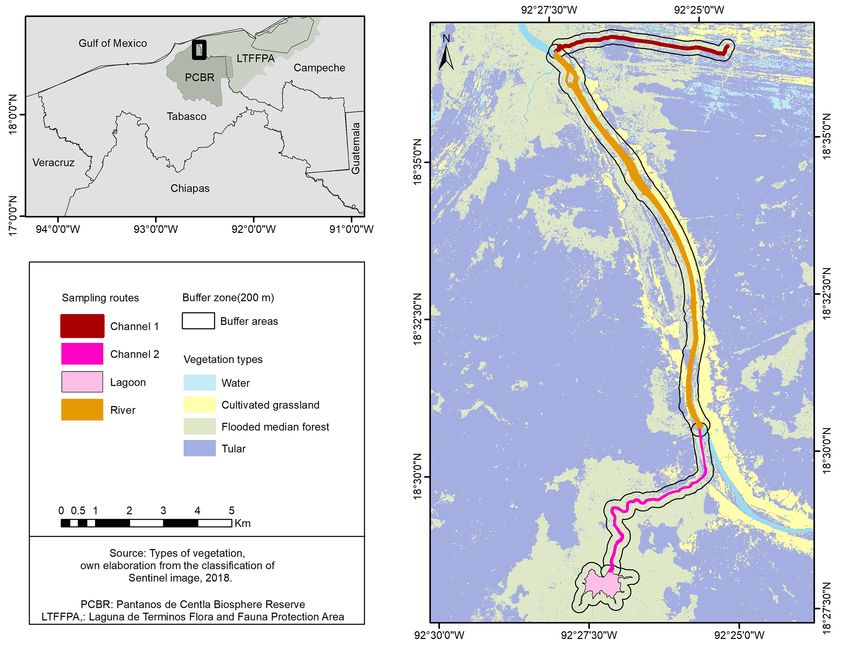
56 C. Iván Flores-Escalona et al.
Figure 1. An adult Crocodylus moreletii from Cometa Lagoon (Lagoon route). Photo by Pierre Charruau.
which favour the absorption of considerable amounts Surveys. We conducted nine fieldtrips to the study
of nitrogen and phosphorous. These areas also serve as sites between September 2014 and March 2018.
refuges for various vertebrates, such as migratory and Sampling was conducted along four routes representing
native birds (Vaga-Moro, 2005; Barba-Macías et al., three different bodies of water, including River (10 km),
2015). However, various threats affect the PCBR and Channel 1 (5.5 km), Channel 2 (6.7 km), and Lagoon
LTFFPA, including livestock activities, the oil industry, (9.9 km), in order to identify the similarities and
deforestation, urbanization, the introduction of exotic differences among the different bodies of water. The
species, and fires, causing the loss and fragmentation routes (Fig. 2) were marked with a global positioning
of habitats and affecting the integrity of the reserves system (GPS, datum WGS84, Garmin eTrex 20).
(Zenteno et al., 2010; Barba-Macías et al., 2015). On each route, night counts were carried out from
The habitats of the PCBR and the LTFFPA represent a flat-bottom aluminium swamp boat, powered by a
great potential for the sustainable use of C. moreletii 15-hp outboard motor, beginning at 21:00 h and with
through a ranching program. Furthermore, as a top a maximum duration of 5 h, performing one route
predator, crocodiles also serve as an indicator species per night. The mean ± SD (minimum–maximum) of
for the conservation of their environment (Mazzotti sampling hours by routes was of 1.5 ± 0.4 (0.9–2.0) at
et al., 2009; Fujisaki et al., 2012). For these reasons, Channel 1, 3.7 ± 1.3 (2.0–5.0) at River, 2.5 ± 1.1 (1.3–
the objective of this work was to obtain information 4.1) at Channel 2, and 1.6 ± 1.3 (0.5–4.1) at Lagoon.
on the population status of C. moreletii within the Crocodiles were located by reflecting the light from
limits of the PCBR and the LTFFPA, to serve as the hand and mining lamps on the tapetum lucidum of their
basis for long-term monitoring of the population. Such eyes. The crocodile encounter rate was determined
information will help create appropriate strategies for along each route, dividing the number of registered
the conservation and protection of C. moreletii and crocodiles by the distance of the routes (crocodiles/
PNA. km). Each individual observation was geolocated with
GPS. In addition, the fishing nets were counted on the
Materials and Methods different routes during the nocturnal surveys.
Crocodile assessment. After locating crocodiles, we
Study area. The study was carried out within the
approached as close as possible to capture them by hand
limits of the PCBR and the LTFFPA, 11.6 km from the
or with a rope tied to a pole, depending on the size of the
Gulf of Mexico. The PCBR is located in Tabasco State
individual. Once captured, the total length (TL) of the
and has an area of 302,706
ha and features 110 bodies of
individual was measured dorsally from the tip of the snout
fresh or brackish water, both temporary and permanent
to the tip of the tail using a flexometer (± 0.1 cm). The
(INE, 2000). The LTFFPA is located in Campeche State,
sex of the individuals was determined by cloacal touch
with an area of 706,148
ha (INE, 1997; Fig. 2).Population Status and Habitat Preferences of Crocodylus moreletii, Mexico 57 Figure 2. Map of the study area between the Pantanos de Centla Biosphere Reserve and the Laguna de Términos Flora and Fauna Protection Area. or by using a rhinoscope to open the cloaca in smaller observation point: air and water temperature (°C) with a individuals. Crocodiles were categorized according mercury thermometer, salinity (ppm) with a hand-held to their TL as neonates (< 30 cm), yearlings (30.1–50 refractometer, and depth (± 0.5 cm) using a rope with a cm), juveniles (50.1–100 cm), subadults (100.1–150 weight that could reach the bottom of the body of water cm), and adults (> 150 cm) (Platt and Thorbjarnarson, and measuring the length of the submerged rope with a 2000). Individuals that were not captured were classified flexometer. according to their estimated TL or as “eyes only” if their Analysis. Statistical analyses were carried out using size could not be estimated. The population structure was Statgraphics centurion XVI Software. The normality of discerned by size classes for both observed and captured the data was evaluated with the Kolmogorov-Smirnov individuals, and by sex for captured individuals. test. A Kruskal-Wallis (KW) test was used to assess Habitat. Habitat characterization was performed differences in encounter rates among sampling routes. A using a supervised classification of the study area with Chi-square test (c2) was used to assess differences in the the help of ArcGIS 10.5 Software and Centinel 2 images general population structure (size and sex) and by size of 10 x 10 m. A buffer zone of 200 m was generated class and route. A G-test was used to assess differences in along the banks of sampling routes, determined based abundance of individuals by vegetation type and among on the maximum reported distance between the shore size classes, sexes, and sampling routes. To identify and a C. moreletii nest (123 m; Pérez-Flores, 2018). The statistical differences in environmental variables (i.e., percentages of vegetation types found within this buffer water and air temperatures, depth and salinity) between zone were calculated. Each crocodile sighting point was sexes and among size classes, Student’s t-test and KW assigned the closest type of vegetation. The following were used, according to the normality of the data. The environmental data were taken at each crocodile results were considered significant with P < 0.05.
58 C. Iván Flores-Escalona et al.
Results to herbaceous plants between 1 and 3 m high rooted in
the bottom of the water and forming a dense mass of
We observed 208 crocodiles during the samplings
difficult access; and the cultivated grassland in which
with an average of 0.8 ± 0.3 crocodiles/km. We obtained
the herbaceous vegetation predominates with the
the highest average encounter rate in Channel 2 with
main use for livestock and agriculture. The median
1.6 ± 0.7 crocodiles/km, while the lowest was obtained
floodplain forest presents the highest percentage of
in Lagoon (0.2 ± 0.2 crocodiles/km). In Channel 1 and
vegetation cover (60%), followed by the tular (22%),
River, we obtained intermediate rates of 0.5 ± 0.3 and
and the cultivated grassland (18%). The same trend can
1.03 ± 0.9 crocodiles/km, respectively. There was a
be observed on each route (Fig. 3). In general, for the
significant difference in the kilometric indices between
study site, we found a significantly higher number of
the different routes (KW; E = 13.8; df = 3; P = 0.003).
crocodiles in flooded median forest compared to the
The encounter rate of Channel 2 is equal to that of
other vegetation types (G = 153; df = 2; P < 0.001). The
River, but higher than the other routes. An average of
same result was obtained for the different size classes
10 ± 3.5 fishing nets was recorded in the river, and no
and sexes, except for the subadult class (Tables 2,3).
nets were found on the other routes.
No significant differences were observed in the
Size class could be determined for 81.3% (n = 169) of the
averages of the environmental variables between sexes
observed/captured crocodiles. Significant differences in
(Table 3). The only significant differences observed at
abundance of individuals were found between size classes
the level of environmental variables among size classes
for Channel 1, Channel 2, and River, where yearlings and
were at the depth and salinity levels. Adults were
juveniles were more abundant than other classes (Table 1).
observed at deeper sites than the other size classes, and
It was possible to capture 26.0% (n = 54) of the crocodiles
yearlings were found at sites significantly more salty
observed and determine the sex of 51 individuals, of
than sites where neonates, juveniles, and adults were
which 37.2% (n = 19) were female and 62.7% (n = 32)
found (Table 2).
male. The results suggest a balanced sex ratio (1.7 ♂ : 1 ♀;
c2 = 3.3, df = 1, P = 0.06). A significantly higher number
Discussion
of males than females was found for Channel 2 (c2 = 4.5,
df = 1, P = 0.03) and juveniles (c2 = 4.8; df = 1; P = 0.02). The average crocodile encounter rate at the study site is
Three types of vegetation were determined in the similar or lower than those reported in areas close to the
established buffer zone: the flooded median forest that study area (Rivera-Téllez et al., 2017; Rodas-Trejo et al.,
is dominated by mangrove vegetation, with the presence 2018; Corado-García et al., 2020). The rates recorded are
of shrub vegetation and palms; the tular that corresponds similar between Channel 2 and River, compared to the
Table 1. Number (percentage) of Crocodylus moreletii observed and captured by age classes recorded in the different
sampling routes.
Neonates Yearlings Juveniles Subadults Adults Eyes Only Statistical Tests
Observations
Channel 1 2 (6.9) 14 (48.3) 8 (27.6) 2 (6.9) 2 (6.9) 1 (3.4) c2 = 23.2; df = 4; P < 0.001
River 4 (4.7) 18 (21.2) 28 (32.9) 13 (15.3) 4 (4.7) 18 (21.2) c2 = 15.2; df = 4; P = 0.004
Channel 2 6 (7.2) 16 (19.3) 33 (39.8) 6 (7.2) 5 (6.0) 17 (20.5) c2 = 10.4; df = 4; P = 0.03
Lagoon 0 (0.0) 3 (27.3) 4 (36.4) 0 (0.0) 1 (9.1) 3 (27.3) c2 = 1.8; df = 2; P = 0.3
Total 12 (5.8) 51 (24.5) 73 (35.1) 21 (10.1) 12 (5.8) 39 (18.8) KW; E = 8.0; df = 4; P = 0.09
Captures
Channel 1 0 (0.0) 10 (52.5) 7 (36.8) 1 (5.3) 1 (5.3) 0 (0.0) c2 = 11.6; df = 3; P = 0.008
River 0 (0.0) 3 (14.2) 14 (66.7) 4 (19) 0 (0.0) 0 (0.0) c2 = 12.1; df = 2; P = 0.002
Channel 2 0 (0.0) 4 (28.6) 10 (71.4) 0 (0.0) 0 (0.0) 0 (0.0) c2 = 2.5; df = 1; P = 0.1
Total 0 (0.0) 17 (31.5) 31 (57.4) 5 (9.3) 1 (1.9) 0 (0.0) KW; E = 5.3; df = 3; P = 0.1Population Status and Habitat Preferences of Crocodylus moreletii, Mexico 59
classes is likely due to size-specific needs in terms of
protection, foraging, and reproduction, and the effects of
the vegetation on their physiology and communication
(Somaweera et al., 2019; Corado-García et al., 2020).
The sex ratio obtained does not show bias towards
any particular sex and coincides with data from other
populations of C. moreletii in Mexico (Casas-Andreu
et al., 2013). The size-class structure shows a dynamic
population with a majority of yearlings and juveniles,
similar to that reported at other sites for this species
(Leyte-Manrique and Ramírez-Bautista, 2005; Cedeño-
Vázquez et al., 2006; Rueda-Cordero et al., 2017). Some
Figure 3. Percentages of vegetation types present in the
different sampling routes in the study area. studies report populations dominated by neonates and
yearlings (Rivera-Téllez et al., 2017) and others report
populations with a higher proportion of adults (Cedeño-
Vázquez and Pérez-Rivera, 2010). According to Llobet
other two routes, despite showing differences in hydric, and Seijas (2003), a population structure dominated
geomorphic and topographic conditions. River has greater mainly by juveniles may be an indicator of the recovery
availability of habitats, as well as banks used for livestock of the population from events of, for example, past
(paddocks) which offers sunbathing areas for crocodiles. overexploitation.
Despite having a similar habitat composition on their The low numbers of subadults and adults in this study
banks, Channel 1 and Channel 2 do not have the same can be related to their more reserved behaviour and the
width, with Channel 1 narrower than Channel 2. In presence of vegetation on the banks, which reduces the
Channel 1 the mangrove coverage reduces the entry of detection of crocodiles (Kushlan and Mazzotti, 1986).
sunlight, and the presence of mangrove roots makes it This could be similar to that observed by Ouboter and
difficult for the boat to move. In contrast, in Channel 2 Nanhoe (1988) in Crocodylus acutus, where subadults
the entry of sunlight is greater, increasing the sunbathing are usually in areas with denser vegetation for their
areas for crocodiles. In addition, access by boat to Channel protection. Other factors, such as illegal hunting and
2 and Lagoon is more controlled by the local fishing fishing with nets, could affect the behaviour of these
cooperative compared to the other two routes, reducing individuals, making their location more difficult. This
human pressure within these two routes. However, has been observed in Caiman crocodilus fuscus, where
the presence of flooded median forest dominated by individuals in constant contact with fishing and hunting
mangrove vegetation on both routes may disadvantage activity are more elusive and difficult to spot than
the observation of crocodiles, since they may be interred alligators with little or no contact with these activities
between the roots out of the reach of the lamp light, (Ron et al., 1998). The illegal fishing nets observed at
making detection impossible. This can be seen in the a relatively high frequency in River represent a threat
Lagoon data, where the smallest number of individuals to the survival of crocodiles at the site, since they are
was recorded and where the shore is dominated by a cause of death cited in several works on crocodiles
mangroves. These differences could condition access to in the region (e.g., Platt and Thorbjarnarson, 2000;
different feeding, coupling, and nesting areas between Cedeño-Vázquez et al., 2006). The main factor affecting
the climatic seasons. Vegetation seems to be the main the capture of neonates at the study site is the presence
environmental variable that generates differences in the of roots as well as floating and shrub vegetation on the
abundance of crocodiles in this study. A recent study banks, which made it impossible to capture many of
on C. moreletii in Guatemala, south of our study area, the sighted crocodiles and to access certain areas on the
showed certain preferences of habitat by size classes different routes of sampling. On tours in areas close to
(Corado-García et al., 2020). Hatchlings, juveniles and the sampling routes, it was possible to observe and hear
subadults seems to prefer forest habitat and reed bed the vocalization of different groups of hatchlings, but
while adults present higher abundance in grassland and access was very limited by the dense vegetation and
tular (Corado-García et al., 2020). Vegetation plays an swampy soil. Hatchlings likely select those areas with
important but poorly understood role in crocodilian life difficult access to be out of reach of predators.
(Somaweera et al., 2019). Habitat preference of size60 C. Iván Flores-Escalona et al.
Table 2. Number of Crocodylus moreletii observed by vegetation type and mean ± SD (N, range) of environmental variables
by size class.
Condition Neonates Yearlings Juveniles Subadults Adults Statistical tests
G = 605.7, df = 3
Flooded median forest 8 47 48 10 10
P < 0.001
G = 3.9, df = 1
Cultivated grassland 1 0 6 0 0
P = 0.04
G = 15.2, df = 1
Tular 2 5 14 7 2
P = 0.004
G = 7.4, df = 2 G = 39.1, df = 1 G = 42.5, df = 2 G = 0.5, df = 1 G = 5.8, df = 1
Statistical test –
P = 0.02 P < 0.05 P < 0.001 P = 0.4 P = 0.01
29.3 ± 1.3 27.5 ± 2.4 27.7 ± 3.1 27.9 ± 2.7 27.0 ± 3.4 E = 7.6, df = 4
Water temperature (°C)
(12, 27–31) (53, 22–31) (65, 11–34) (18, 20–30) (11, 19–30) P = 0.1
Air temperature 26.9 ± 1.2 25.9 ± 2.0 25.7 ± 2.8 26.1 ± 2.2 25.3 ± 2.3 E = 2.6, df = 4
(°C) (12, 26–29) (53, 20–29) (65, 11–29) (18, 22–29) (11, 22–28) P = 0.6
Depth 76.2 ± 53.6 113.6 ± 136.9 91.6 ± 79.2 117.5 ± 73.1 302.4 ± 236.5 E = 13.7, df = 4
(cm) (12, 2.4–150) (52, 5–1000) (62, 10–438) (19, 10–276) (11, 65–850) P = 0.008
Salinity 0.0 ± 0.0 0.7 ± 1.5 0.1 ± 0.6 0.3 ± 1.2 0.0 ± 0.0 E = 13.7, df = 4
(ppm) (12, 0–0) (52, 0–5) (60, 0–5) (16, 0–5) (10, 0–0) P = 0.01
Table 3. Number of Crocodylus moreletii observed by vegetation type and mean ± SD (N, range) of
environmental variables by sex.
Condition Females Males Total Statistical tests
G = 0.04, df = 1
Flooded median forest 15 19 34
P = 0.4
Cultivated grassland 0 3 3 -
G = 2.7, df = 1
Tular 4 6 10
P = 0.2
G = 6.7; df = 1 G = 14.9, df = 2 G = 33.7, df = 2
Statistical test –
P = 0.009 P = 0.0005 P < 0.05
Water temperature 27.6 ± 2.4 27.1 ± 3.1 t = 0.5, df = 16
27.4 ± 0.4
(°C) (17, 23–31) (30, 20–30) P = 0.6
Air temperature 24.6 ± 6.1 25.7 ± 20 t = 0.7, df = 16
25.2 ± 0.8
(°C) (17, 20–29) (30, 20–29) P = 0.4
Depth 48 ± 53.0 125 ± 176 t = 0.1, df = 15
128.9 ± 5.5
(cm) (16, 10–85) (30, 0–1000) P = 0.8
Salinity 0.2 ± 0.7 0.8 ± 1.8 t = –1.3, df = 16
0.5 ± 0.4
(ppm) (17, 0–3) (30, 0–5) P = 0.1Population Status and Habitat Preferences of Crocodylus moreletii, Mexico 61
The recorded air and water temperatures are within Acknowledgments. We thank the Consejo Nacional de Ciencias y
the averages reported for the species, and so are depth Tecnología of Mexico for granting a Master’s support scholarship,
and the Crocodile Specialist Group of the Species Survival
and salinity (Casas-Andreu et al., 2013). The set of
Commission of the International Union for Conservation of Nature
environmental variables such as depth and temperature,
for awarding a student research support program scholarship, to the
can directly influence crocodiles, which they can be first author. This study was financed by project “Fordecyt 273646:
using to regulate their temperature, staying in warmer Cambio global y sustentabilidad en la cuenca del Usumacinta y
waters in cold periods and submerging in colder zona marina de influencia: Bases para la adaptación al cambio
waters during the hot season (Pérez and Rodríguez, climático desde la ciencia y la gestión del territorio.” The
2005). These results, together with the large number of Dirección General de Vida Silvestre of the Secretaría de Medio
Ambiente y Recursos Naturales of Mexico granted permits for
registered yearlings and juveniles of crocodiles, show
the capture and handling of wild crocodiles with official numbers
that the study area maintains adequate conditions for
SGPA/DGVS/03484/14, 01009/16, 08912/17, and 002467/18.
the crocodile population. Therefore, the conservation
of the study area is important to sustain a population
References
of C. moreletii that can be maintained or increased over
time. The study area shows salinity values of 0–5 ppm Barba-Macías, E., Valadez-Cruz, F., Pinkus, M., Pinkus, M., Juárez
Flores, J. (2015): Reserva de la Biosfera Pantanos de Centla:
due to its proximity to the coastline but this is not a
aspectos socio-ambientales prioritarios. In: La Investigación
problem for C. moreletii since the species, considered a
Científica y las Áreas Naturales Protegidas en México, p. 11–29.
freshwater species, survives very well in brackish water Ortega-Rubio, A., Pinkus-Rendón, M.J., Espitia-Moreno, I.C.,
conditions (Casas-Andreu et al., 2013). Eds., Centro de Investigaciones Biológicas del Noroeste S.C. La
The vegetation cover present in the study area is Paz B.C.S., Universidad Autónoma de Yucatán, Mérida, Yucatán
dominated by flooded median forest, which could and Universidad Michoacana de San Nicolás de Hidalgo,
indicate that it was the type of vegetation present in Morelia, Michoacán, México.
Barrios, G., Cremieux, J. (Eds.) (2018): Protocolo de Rancheo
almost the entire area before undergoing changes due to
para Cocodrilo de Pantano (Crocodylus moreletii) en México.
human activity. Some areas have been transformed into
México, D.F., Comisión Nacional para el Conocimiento y Uso
cultivated grassland, used mainly for livestock. Despite de la Biodiversidad.
the fact that the study area does not present a great variety Casas-Andreu, G. (2002): Hacia la conservación y manejo sustentable
of vegetation types available for crocodiles, it is evident del lagarto o cocodrilo de pantano (Crocodylus moreletii)
that they have a greater preference towards areas with en México. In: La Conservación y el Manejo de Caimanes y
presence of flooded median forest. A greater number Cocodrilos de América Latina. Volume 2, p. 27–45. Verdade, L.M.,
Larriera, A., Eds., Piracicaba, São Paulo, Brail, C.N. Editoria.
of crocodiles were recorded in the River route in the
Casas-Andreu, G., Barrios-Quiroz, G., Escobedo-Galván, A.H.,
flooded median forest, leaving the cultivated grassland
Aguilar-Miguel, X. (2013): Sinopsis de Datos Biológicos y
last, even though the percentages of habitat between Ecológicos del Cocodrilo de Pantano (Crocodylus moreletii).
the flooded median forest and the cultivated grassland México, D.F., Instituto de Biología, Universidad Autónoma de
are very similar. This highlights the importance of the México.
flooded median forest for crocodiles by providing them Cedeño-Vázquez, J.R., Pérez-Rivera, S.D. (2010): El cocodrilo de
with refuge, feeding and nesting areas, as observed in pantano (Crocodylus moreletii) en laguna esmeralda, Quintana
Roo, México. Revista Latinoamericana de Conservación 1: 91–98.
other studies (Corado-García et al., 2020).
Cedeño-Vázquez, J.R., Ross, J.P., Calmé, S. (2006): Population
The present work contributes to the population
status and distribution of Crocodylus acutus and C. moreletii
knowledge of C. moreletii in an area between the PCBR in southeastern Quintana Roo, Mexico. Herpetological Natural
and the LTFFPA. Maintaining control over human History 10: 53–66.
activities in certain areas directly benefits crocodile Corado-García, V.A., Cajas-Castillo, J.O., Benítez-Moreno, J.A.
populations, as observed along the Channel 2 route (2020): Estado poblacional y distribución del cocodrilo de
where the greatest crocodile abundance was recorded. pantano (Crocodylus moreletii) en regiones prioritarias de Petén,
Guatemala. Revista Latinoamericana de Herpetología 3(2): 23–33.
The study shows a dynamic population in recovery and
Fujisaki, I., Mazzotti, F.J., Hart, M.K., Rice, G.K., Ogurcak, D.,
a suitable habitat for its growth. However, it is important
Rochford, M., et al. (2012): Use of alligator hole abundance and
to continue monitoring the population in order to detect occupancy rate as indicators for restoration of a human-altered
effects on its population parameters due to possible wetland. Ecological Indicators 23: 627–633.
environmental or anthropogenic factors. The presence Instituto Nacional de Ecología (1997): Programa de Manejo de
of illegal fishing nets and the change in land use are the la Zona de Protección de Flora y Fauna Laguna de Términos.
main threats detected in the study area for C. moreletii. México, D.F., Secretaria del Medio Ambiente, Recursos
Naturales y Pesca.62 C. Iván Flores-Escalona et al.
Instituto Nacional de Ecología (2000): Programa de Manejo de Rivera-Téllez, E., López, S.G., Antaño, L.A., Benítez, D.H.
la Reserva de la Biosfera Pantanos de Centla. México, D.F., (2017): Informe del Programa de Monitoreo del Cocodrilo de
Secretaría del Medio Ambiente, Recursos Naturales y Pesca. Pantano en México, Temporadas 2014 a 2015 y Análisis de
Kushlan, J.A., Mazzotti, F.J. (1986). Population biology and Tendencias del 2011 al 2015. México, D.F., Comisión Nacional
status of the American Crocodile in South Florida. In: para el Conocimiento y Uso de la Biodiversidad.
Proceedings of the 7th Working Meeting of the Crocodile Rodas-Trejo, J., Ocampo-González, P., Mandujano-Camacho,
Specialist Group, p. 184–206. Gland, Switzerland, IUCN. H., Grajales-Zepeda, R., Hernández-Nava, J. (2018): Estado
Leyte-Manrique, A., Ramírez-Bautista, A. (2005): Contribución a poblacional del Crocodylus moreletii (Reptilia: Crocodylidae)
la ecología de Crocodylus moreletii en el sistema Yucalpeten- en el Área Natural Protegida Laguna de Términos, Campeche,
Progreso-Chicxulub, Yucatán, México. Boletín de la Sociedad México. Quehacer Científico en Chiapas 12: 41–46.
Herpetológica Mexicana 13: 69–77. Ron, R.S., Vallejo, A., Andanza, E. (1998): Human influence on
Llobet, A., Seijas, A.E. (2003): Estado poblacional y lineamientos the wariness of Melanosuchus niger and Caiman crocodilus in
de manejo del caimán del Orinoco (Crocodylus intermedius) en Cuyabeno, Ecuador. Journal of Herpetology 32: 320–324.
el río Capanaparo, Venezuela. In: Manejo de Fauna Silvestre Rueda, B.A., López, M.A., Olivera, L.D. (2017): Uso de hábitat
en Amazonia y Latinoamérica. Selección de Trabajos, p. 117– del cocodrilo de pantano Crocodylus moreletii en una laguna
129. Polanco-Ochoa, R., Ed., Bogotá, Colombia, V Congreso urbanizada en México. Quehacer Científico en Chiapas 12: 35–40.
Internacional, CITES, Fundación Natura. Sánchez-Herrera, O., López, S.G., García, N.A., Benítez, D.H.
Mazzotti, F.J., Ronnie, B.G., Brandt, A.L., Cherkiss, S.M., (2011): Programa de Monitoreo de Cocodrilo de Pantano
Jeffery, M.A., Rice, G.K. (2009): Alligator and crocodiles as (Crocodylus moreletii) México-Belice-Guatemala. México,
indicators for restauration of Everglades ecosystem. Ecological D.F., Comisión Nacional para el Conocimiento y Uso de la
Indicators 9: 37–49. Biodiversidad.
Ouboter, P.E., Nanhoe, M.R. (1988): Habitat selection and Somaweera, R., Brien, M.L., Platt, S.G., Manolis, C., Webber,
migration of Caiman crocodilus crocodilus in a swamp B.L. (2019): Direct and indirect interactions with vegetation
and swamp-forest habitat in Northern Suriname. Journal of shape crocodilian ecology at multiple scales. Freshwater
Herpetology 22: 283–294. Biology 64: 257–268.
Pérez, A.T., Rodríguez, R.J. (2005): Influencia de la temperatura Vega-Moro, A. (2005): Plan de Conservación para la Reserva
del aire y del agua en el crecimiento de Crocodylus intermedius de la Biosfera Pantanos de Centla y el Área de Protección
en dos condiciones de cautiverio. Boletín del Centro de de Flora y Fauna de Laguna de Términos. México, D.F.,
Investigaciones Biológicas 39: 15–26. Pronatura, The Nature Conservancy, Comisión Nacional de
Pérez-Flores, J. (2018): Predation of adult female Morelet’s áreas Naturales Protegidas.
crocodile (Crocodylus moreletii) by a jaguar (Pantera onca) in Zenteno, C.E., Barba, M.E., Bello-Gutiérrez, J., Ochoa-Gaona, S.
the Calakmul region, México. Herpetology Notes 11: 613–616. (2010): Caracterización espacio-temporal del hábitat y presencia
Platt, S.G., Thorbjarnarson, J.B. (2000): Population status and de Dermatemys mawii (Testudines: Desmatemydae) en la
conservation of Morelet’s crocodile, Crocodylus moreletii, in cuenca del Grijalva-Usumacinta, Tabasco, México. Revista de
northern Belize. Biological conservation 96: 21–29. Biología Tropical 58: 1247–1260.
Platt, S.G., Sigler, L., Rainwater, T.R. (2010): Morelet’s Crocodile
Crocodylus moreletii. In: Crocodiles, Status Survey and
Conservation Action Plan, p. 79–83. Third Edition. Manolis,
S.C., Stevenson, C., Eds., Darwin, Northern Territory, Australia,
IUCN Crocodile Specialist Group.
Accepted by Graham WaltersYou can also read

















































