Molecular mechanisms and highly functional development for stress tolerance of the yeast Saccharomyces cerevisiae - Oxford Academic Journals
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Bioscience, Biotechnology, and Biochemistry, 2021, Vol. 85, No. 5, 1017-1037
doi: 10.1093/bbb/zbab022
Advance access publication date: 12 February 2021
AWARD REVIEW
AWA R D R E V I E W
Molecular mechanisms and highly functional
development for stress tolerance of the yeast
Downloaded from https://academic.oup.com/bbb/article/85/5/1017/6134096 by guest on 14 December 2021
Saccharomyces cerevisiae
1,∗,†
Hiroshi Takagi
1
Division of Biological Science, Graduate School of Science and Technology, Nara Institute of Science and
Technology, Nara, Japan
∗
Correspondence: Hiroshi Takagi, hiro@bs.naist.jp
†
This review was written in response to the author’s receipt of the JSBBA Award in 2020.
ABSTRACT
In response to environmental stress, microorganisms adapt to drastic changes while exerting cellular functions by
controlling gene expression, metabolic pathways, enzyme activities, and protein–protein interactions. Microbial cells
that undergo a fermentation process are subjected to stresses, such as high temperature, freezing, drying, changes in pH
and osmotic pressure, and organic solvents. Combinations of these stresses that continue over long terms often inhibit
cells’ growth and lead to their death, markedly limiting the useful functions of microorganisms (eg their fermentation
ability). Thus, high stress tolerance of cells is required to improve productivity and add value to fermented/brewed foods
and biofuels. This review focuses on stress tolerance mechanisms, including l-proline/l-arginine metabolism, ubiquitin
system, and transcription factors, and the functional development of the yeast Saccharomyces cerevisiae, which has been
used not only in basic science as a model of higher eukaryotes but also in fermentation processes for making alcoholic
beverages, food products, and bioethanol.
Received: 6 January 2021; Accepted: 25 January 2021
© The Author(s) 2021. Published by Oxford University Press on behalf of Japan Society for Bioscience, Biotechnology, and Agrochemistry. This is
an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0/),
which permits unrestricted reuse, distribution, and reproduction in any medium, provided the original work is properly cited.
1017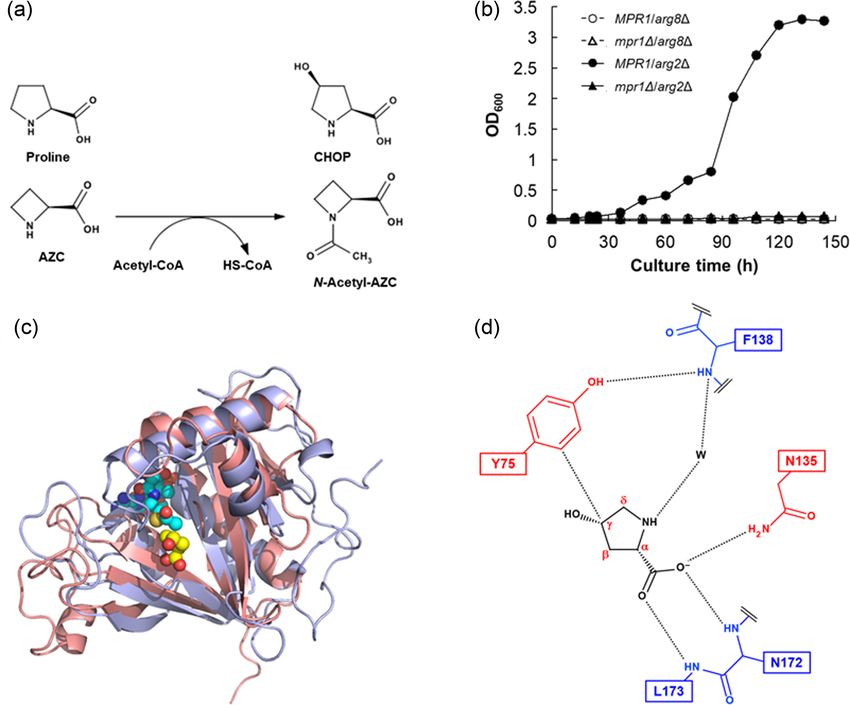
1018 Bioscience, Biotechnology, and Biochemistry, 2021, Vol. 85, No. 5
Graphical Abstract
Downloaded from https://academic.oup.com/bbb/article/85/5/1017/6134096 by guest on 14 December 2021
Stress tolerance mechanisms found in Saccharomyces cerevisiae can contribute to breeding of industrial yeast strains with
improved fermentation productivity.
Keywords: yeast stress tolerance, proline/arginine metabolism, nitric oxide, ubiquitin system, transcription factor
Abbreviations: AcCoA: acetyl-CoA; Arg: l-arginine; Art: arrestin-related trafficking adaptor; AVT: amino acid vacuolar
transport; AZC: l-azetidine-2-carboxylate; CHOP: cis-4-hydroxy proline; Cit: l-citrulline; DARP:
2,5-diamino-6-(5-phospo-d-ribosylamino)-pyrimidin-4(3H)-one; GK: γ -glutamyl kinase; GM: genetically modified; GSA:
l-glutamate-γ -semialdehyde; GSH: glutathione; GSNO: S-nitrosoglutathione; GSNOR: S-nitrosoglutathione reductase;
GTPCH2: GTP cyclohydrolase II; HPLC/FLD: high-performance liquid chromatography with fluorescence detection; MRC:
mitochondrial respiratory chain; NIR: nitrite reductase; NO: nitric oxide; NOD: NO dioxygenase; NOS: NO synthase; NR:
nitrate reductase; Orn: l-ornithine; P5C: l-1 -pyrroline-5-carboxylate; PCR: polymerase chain reaction; Pro: l-proline;
ROS: reactive oxygen species; RSNO: S-nitrosothiols; SC: self-cloning; α-syn: α-synuclein; TCA: tricarboxylic acid; Ub:
ubiquitin
Environmental stresses, such as nonoptimal growth tempera- For example, the 2016 Nobel Prize in Physiology or Medicine
tures, prolonged nutrient starvation, osmotic pressure and pH was awarded to Yoshinori Ohsumi for his discoveries of the
imbalances, and exposure to toxic molecules and free radicals, autophagy mechanisms in yeast cells (Długońska 2017). Yeast
continuously challenge all livings cells (Mager and De Kruijff microbes have been also used in fermentation processes to
1995). Microbial cells, which are present in and on humans and make alcoholic beverages, bread, and fermented food products
other animals and in soil, water, and the air, have developed a for hundreds of years. More recently, yeast has been used in
series of stress-responsive systems to withstand such adverse the production of bioethanol. Our research group recently iso-
environmental conditions (Estruch 2000). These systems exist lated yeast mutants that overproduce isoamyl acetate, which is
at the transcriptional, protein, and metabolic levels. For exam- known as the key ingredient providing the flavor of sake, which
ple, microbial cells perceive and transduce some external stimuli is made from fermented rice (Takagi et al. 2015; Abe et al. 2019). By
via signal transduction pathways, resulting in the global remod- brewing with these mutants, several kinds of awamori, which is
eling of gene expression, which itself is governed by the cells’ a traditional distilled alcohol beverage in Okinawa Islands, have
transcriptional activators and repressors. been recently commercialized.
Stress applied to cells generally affects both transcription Yeast cells are subjected to a variety of environmental
and translation activities, which results in an inhibition of stresses during fermentation processes, and severe stress condi-
de novo protein synthesis. Protein quality control and protein tions can reduce their fermentation efficiency. The cellular tol-
homeostasis are essential for the cellular responses to stress, erance to various types of stress is thus desired for their use in
since fluctuations in environmental parameters can damage biotechnological applications. Freezing and other extreme tem-
proteins; examples of such damage are a destabilization of cel- peratures, high ethanol concentrations, high osmotic pressure,
lular structures, inhibition of the activities of enzymes, and and drying are some of the stresses to which yeast may be sub-
chemical gradient instability, any of which will eventually result jected during fermentation, and this stress can induce both the
in disruption of the cells. In response to harsh stress, energy- denaturation of proteins and the generation of reactive oxygen
producing and energy-consuming processes in cells are down- species (ROS) due to mitochondrial damage, inhibiting the yeast
regulated and enter a quiescent state. A dynamic shift in the cells’ growth and/or causing their death. In fact, we observed a
central metabolic pathways that convert nutrients into energy significant increase in intracellular ROS levels under a variety of
and biomass often accompanies this state. stress conditions that mimic fermentation processes, indicating
The budding yeast Saccharomyces cerevisiae is a single-cell that yeast cells are commonly exposed to oxidative stresses dur-
eukaryote that has been studied in basic science for decades. ing fermentation (Kitagaki and Takagi 2014).
Stress tolerance mechanisms in S. cerevisiae 1019
Downloaded from https://academic.oup.com/bbb/article/85/5/1017/6134096 by guest on 14 December 2021
Figure 1. Metabolic pathways of l-proline (Pro) and l-arginine (Arg) in S. cerevisiae. Protein names: Pro1, γ -glutamyl kinase; Pro2, γ -glutamyl phosphate reductase;
Pro3, 1 -pyrroline-5-carboxylate (P5C) reductase; Put1, Pro oxidase; Put2, P5C dehydrogenase; Arg2, N-acetyl-glutamate synthase; Arg6, N-acetyl-glutamate kinase;
Arg5, N-acetyl-glutamyl-5-phosphate reductase; Arg8, N-acetyl-ornithine aminotransferase; Arg7, N-acetyl-ornithine acetyltransferase; Arg11, Ornithine transporter;
Arg3, ornithine carbamoyltransferase; Arg1, arginosuccinate synthetase; Arg4, arginosuccinate lyase; Car1, arginase; Car2, ornithine aminotransferase; Mpr1, N-
acetyltransferase; and Tah18, flavoprotein (diflavin reductase family). Mainly, Pro and Arg are synthesized from l-glutamate. In response to high-temperature stress,
nitric oxide (NO) is produced from the increased Arg through the Mpr1- and Tah18-dependent manner. Activity of Pro1 or Arg2, Arg6, and Arg5 is subject to feedback
inhibition by Pro or Arg, respectively.
In the present review, I focus on my research group’s findings carboxylate (P5C) reductase (Pro3) (Figure 1). The activity of GK is
regarding several stress tolerance mechanisms in yeast and the subjected to feedback inhibition by Pro, indicating that GK is the
highly functional development of yeast. The physiological roles rate-limiting enzyme that controls the intracellular level of Pro
and metabolic regulations of the amino acids l-proline (Pro) and (Sekine et al. 2007). Pro is degraded generally in the mitochondria,
l-arginine (Arg) in stress tolerance are described, as is the mech- but the import mechanism responsible for this is not known; the
anism underlying the repair or degradation of stress-induced process involves mitochondrial Pro oxidase (Put1) converting Pro
abnormal proteins via the ubiquitin system. Stress-related into P5C, which is then processed into Glu by P5C dehydrogenase
transcription factors that can be manipulated for the improve- (Put2). S. cerevisiae cells can also synthesize Pro from Arg via l-
ment of stress tolerance are also presented. ornithine (Orn) catalyzed by Orn aminotransferase (Car2) in the
cytoplasm (Kaino et al. 2012).
We have reported the importance of the localization of Pro in
vacuoles. An S. cerevisiae mutant with intracellular Pro accumu-
The mechanisms underlying stress tolerance lation cultured in a minimal medium also accumulated Pro in its
via l-proline/l-arginine metabolism vacuoles (Matsuura and Takagi 2005). Deletion of the PEP3 gene,
The metabolic regulations of l-proline in yeast which encodes a vacuolar membrane protein that is required for
vacuolar biogenesis, conferred stress sensitivity to yeast cells,
The amino acid Pro serves as both a source of nitrogen and a indicating that a specific vacuolar function is involved in stress-
stress protectant in S. cerevisiae (Takagi 2008). It is used by this protective effects of Pro. Seven AVT genes (AVT1-7) in S. cerevisiae
yeast as a carbon source via the mitochondrial Pro metabolic encode vacuolar amino acid transporters that are members of
pathway and the subsequent tricarboxylic acid (TCA) cycle, as the amino acid vacuolar transport (AVT) family. In response to
our group recently demonstrated (Nishida, Watanabe and Takagi the application of exogenous Pro, these genes were significantly
2016a). The putative mitochondrial α-ketoglutarate-dependent upregulated. We added Pro to S. cerevisiae cells grown in a mini-
dioxygenase Fmp12 inhibits the growth of yeast cells by avoid- mal medium and then observed the effects of both the deletion
ing the ATP production step in the TCA cycle, as suggested by and the overexpression of the AVT genes on the subcellular dis-
the results of growth-phenotype analyses using gene disruption tribution of amino acids (Nishida et al. 2016b). The results indi-
and overexpression (Nishida, Watanabe and Takagi 2016a). The cated that Avt3 is the major protein involved in the export of Pro
metabolic regulation of Pro, including its biosynthesis, cellular from the vacuole, and that the AVT genes induced by exogenous
localization, transport, and degradation, is thus of great interest. Pro are involved in the bidirectional transport of Pro across the
S. cerevisiae cells synthesize Pro mainly from l-glutamate vacuolar membrane.
(Glu) via 3 cytoplasmic enzymes: γ -glutamyl kinase (GK) (Pro1); Pro is a predominant amino acid in grape must (the freshly
γ -glutamyl phosphate reductase (Pro2); and l-1 -pyrroline-5- crushed juice that contains the grapes’ skins, seeds, and stems),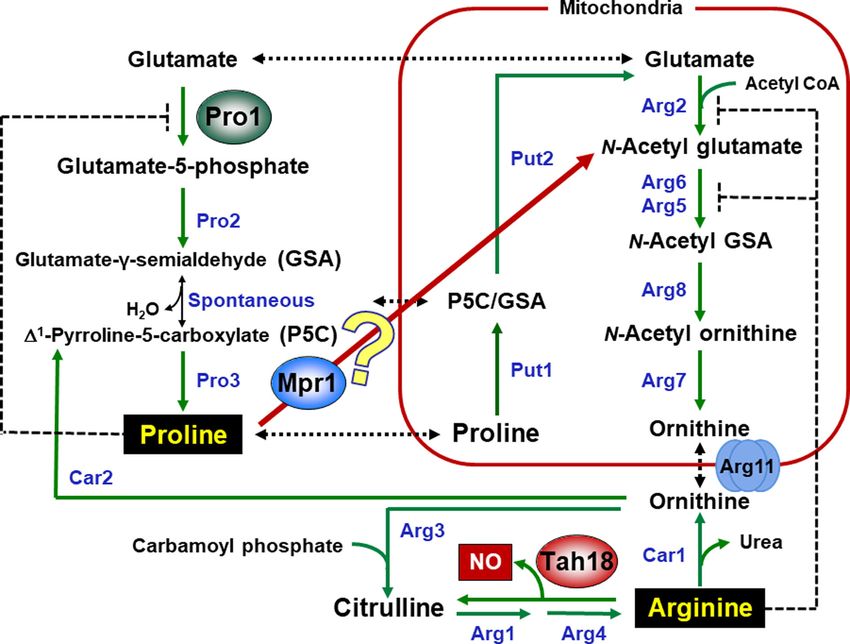
1020 Bioscience, Biotechnology, and Biochemistry, 2021, Vol. 85, No. 5
but Pro is not utilized effectively by S. cerevisiae in wine-making for cell protection against stress in yeast (Matsuura and Takagi
processes, resulting in a nitrogen deficiency during fermen- 2005; Takagi, Taguchi and Kaino 2016).
tation and the accumulation of Pro in the wine. We recently New physiological functions of Pro and GK were recently re-
revealed that Arg inhibits the utilization of Pro by specifically ported. We observed that intracellular Pro regulates the replica-
inducing the endocytosis of the high-affinity Pro transporter tive lifespan of S. cerevisiae (Mukai et al. 2019). The deletion
Put4 (Nishimura, Tanikawa and Takagi 2020b). We recently of PUT1 and the expression of PRO1I150T , which encodes the
revealed that Arg inhibits the utilization of Pro by specifically Ile150Thr variant of GK, resulted in the accumulation of Pro
inducing the endocytosis of Put4, a high-affinity Pro transporter. and extended the lifespan of yeast cells. Inversely, we found
A key regulator for Put4 endocytosis is an adaptor protein for that the disruption of the Pro biosynthetic genes PRO1, PRO2,
the ubiquitin ligase Rsp5, ie Art3 (Nishimura, Tanahashi and and CAR2 decreased the stationary Pro level and shortened
Takagi 2020a). We discovered that deletion of the ART3 gene the cellular lifespan. Pro metabolism may thus have a physi-
thoroughly canceled inhibition of Pro utilization by Arg. These ological role in maintaining the lifespan of yeast cells. In the
findings may contribute to the development of strains of wine nematode Caenorhabditis elegans, higher Pro levels increased
yeast that can efficiently assimilate the abundant Pro in grape longevity (Edwards et al. 2015). Impaired insulin/insulin-like
Downloaded from https://academic.oup.com/bbb/article/85/5/1017/6134096 by guest on 14 December 2021
must during the fermentation processes. growth factor (IGF-1) signaling was shown to extend the lifespan
of C. elegans by promoting mitochondrial Pro catabolism, and Pro
supplementation also extended the lifespan (Zarse et al. 2012).
The physiological functions of l-proline in yeast
Although these observations indicated that Pro directly affects
Several types of bacteria and plants accumulate Pro as an osmo- longevity in C. elegans, it should be noted that in yeast, changes
protectant (ie a compatible solute) in response to osmotic stress in amino acid metabolism caused by the intracellular Pro might
(Verbruggen and Hermans 2008). In contrast, yeast cells do not also be involved in longevity.
increase their level of intracellular Pro in response to various A novel function of GK that is independent of the biosyn-
stress conditions including osmotic stress; rather, they respond thesis of Pro was suggested by the finding that the deletion of
by synthesizing glycerol or trehalose (Kaino and Takagi 2008). PRO1 leads to high sensitivity to various stresses, but the dele-
The functions of Pro that have been observed in vitro include a tion of PRO2, which encodes γ -glutamyl phosphate reductase,
reduction of the melting temperature (Tm ) of DNA in response does not. We demonstrated that PRO1 genetically interacts with
to a destabilization of the double helix in salinity–stress tests, UBP3, which encodes ubiquitin-specific protease and is required
plus the stabilization of proteins and the cell membrane against for the selective autophagy of ribosomes (ribophagy) (Figure 2c)
elevated temperatures, freezing, and/or dehydrations (Takagi (Tatehashi, Watanabe and Takagi 2016). Interestingly, GK activity
2008). Pro also inhibits protein aggregation during its refolding or is indispensable for ribophagy, which is important for cell sur-
folding (Ignatova and Gierasch 2006), increases the solubility of vival during nitrogen starvation, while nonselective autophagy
sparingly soluble proteins, and scavenges ROS (Takagi 2008), par- is not.
ticularly hydroxyl radical (Smirnoff and Cumbes 1989). In plants
subjected to osmotic stress, the increase in Pro was shown to
reduce the levels of free radicals (Hong et al. 2000). In cultured The stress tolerance and fermentation ability of
tobacco cells, treatment with Pro conferred tolerance to salinity
industrial yeasts with l-proline accumulation
and cadmium stress (Islam et al. 2009). The mechanisms under-
lying these functions in vivo are not established. In S. cerevisiae, In the fermentation process used to produce Japanese sake,
the oxidation level in wild-type cells was clearly increased after yeast cells are exposed to high ethanol concentrations that block
ethanol exposure, suggesting the generation of ROS. Notably, in the cell growth, viability, and fermentation (Kunkee and Bisson
yeast cells at the stationary phase subjected to ethanol stress, 1993). An appropriate Pro level in yeast cells can protect against
the accumulation of Pro significantly reduced the level of ROS ethanol stress, and the development of ethanol-tolerant sake
and increased the cell survival rate (Takagi, Taguchi and Kaino yeasts could therefore reduce the time needed for fermentation.
2016). In addition, the taste of sake is derived from the amino acids
Pro and Arg are both known to exert significant that the yeast produces, and the creation of yeast strains with
cryoprotective activity that is nearly equal to those of glycerol an added accumulation of “sweet” Pro could thus expand the
and trehalose, which are major cryoprotectants for S. cerevisiae range of sake flavors.
(Takagi, Iwamoto and Nakamori 1997). It has been speculated A novel sake yeast strain was constructed by (i) replac-
that Pro inhibits ice crystal formation and dehydration by ing the wild-type PRO1 allele with the PRO1D154N allele en-
forming strong hydrogen bonds with intracellular free water, coding the Asp154Asn variant of GK and (ii) disrupting the
probably due to the extremely high water solubility of Pro. PUT1 gene encoding Pro oxidase, which is required for the
We previously isolated or constructed S. cerevisiae strains that utilization of Pro (Takagi et al. 2005). As expected, this strain
accumulated Pro, and showed that these strains enhanced accumulated Pro and was more tolerant to ethanol stress
tolerance to many types of stress treatments including ethanol, compared to the control strain. Moreover, the sake brewed with
hydrogen peroxide, desiccation, and freeze-thawing (Takagi, the Pro-accumulating strain contained 5 times more Pro than
Iwamoto and Nakamori 1997, 2000a, 2000b; Morita, Nakamori sake brewed with the control strain, with no effect on the fer-
and Takagi 2001; Morita, Nakamori and Takagi 2003; Terao, mentation profiles. The Pro-rich sake had a distinctive sweet and
Nakamori and Takagi 2003). Interestingly, the Asp154Asn and light flavor (unpublished results).
Ile150Thr variants of GK (the PRO1 gene product) were less Recombinant yeasts have been investigated for commercial
sensitive to feedback inhibition, leading to the oversynthesis use. Since many consumers have indicated objections to genet-
of Pro. The yeast cells that expressed these 2 variants could ically modified (GM) yeasts, the use of self-cloning (SC) yeast
therefore accumulate Pro, and exhibited higher tolerance to the that do not contain foreign genes or DNA sequences other than
freeze-thawing stress (Figure 2a and b) (Sekine et al. 2007). The yeast DNA may be more acceptable. We have constructed SC
appropriate level of intracellular Pro might thus be important diploid sake yeast strains that accumulate Pro (Takagi et al.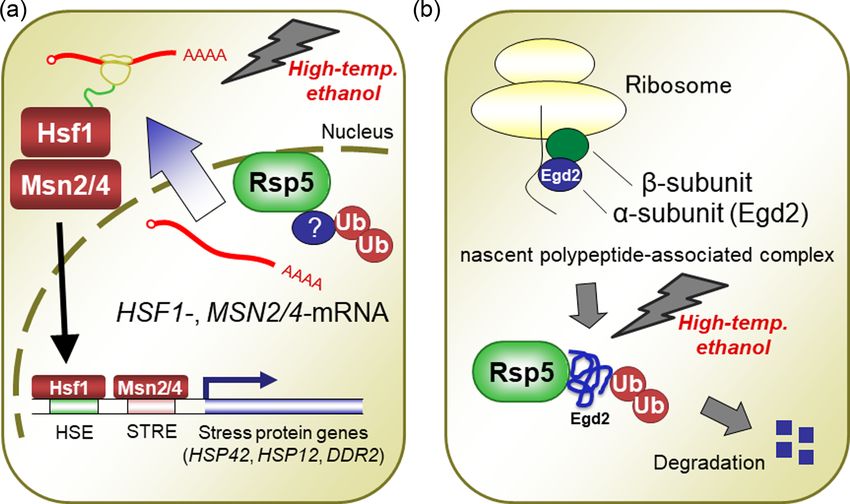
Stress tolerance mechanisms in S. cerevisiae 1021
Downloaded from https://academic.oup.com/bbb/article/85/5/1017/6134096 by guest on 14 December 2021
Figure 2. Physiological functions of Pro and GK in S. cerevisiae. (a) Effect of Pro on the GK activity. The activities of various GKs were measured using the hydroxamate
assay in the presence of the Pro concentrations indicated. The enzyme activities in the absence of Pro were defined as 100%. The values are the means of results from
3 independent experiments (Sekine et al. 2007). (b) Freezing stress tolerance of S. cerevisiae strains expressing GK. The cell viabilities of each strain were measured
after freezing at −20 °C for the times indicated. The number of colonies before freezing was defined as 100%. The values are the means of results from 3 independent
experiments (Sekine et al. 2007). (c) Proposed model for regulatory mechanism of ribophagy in yeast. First, nitrogen starvation induces cell death and ribophagy
mediated by the Ubp3–Bre5 complex and GK. GK activity is required not only for Pro biosynthesis, but also for ribophagy. Finally, ribophagy suppresses cell death under
nitrogen-starvation conditions (Tatehashi et al. 2016).
2007), and we observed that compared to the parent strain, the and/or respiratory chain (Ando et al. 2007; Landolfo et al. 2008;
Pro-accumulating strains produced greater amounts of Pro in Shima, Ando and Takagi 2008). Dried yeast is generally used in
sake (approx. 3 times more Pro) (Figure 3a) and approx. 30% baking processes because it can be stored for longer periods and
more total amino acids. The ethanol production rate of the Pro- is less costly to transport compared to compressed yeast. Sweet
accumulating strains was somewhat faster than that of the par- (high-sugar) dough is up to 40% sucrose per weight of flour. The
ent strain (Figure 3b). These results suggest that intracellular Pro widely available frozen-dough technology can supply oven-fresh
improves the fermentation rate, and that fermentation times of bakery products to consumers.
Pro-accumulating yeast strains might be shorter than those of Kaino et al. (2008) constructed SC diploid baker’s yeast strains
wild-type control strains. that accumulate Pro. They achieved this by replacing the wild-
We recently isolated yeast mutants that are resistant to the type PRO1 gene encoding GK with the PRO1D154N or PROI150T al-
Pro analogue, l-azetidine-2-carboxylate (AZC), derived from a lele and disrupting the PUT1 gene (Figure 3c, and d). When the
diploid sake yeast strain. Some of these mutants produced more dough was prefermented before freezing and then kept frozen,
Pro in the brewed sake, and one of the mutants carried a novel the gassing power of wild-type cells was greatly decreased com-
mutation in the PRO1 gene encoding the Gln79His variant of GK pared to the power observed before freezing. On the other hand,
(Murakami et al. 2020). This mutation resulted in extreme de- the cells that accumulated Pro showed ∼50% greater fermen-
sensitization to feedback inhibition by Pro, leading to an over- tation ability compared to the wild-type cells, indicating that
production of Pro. The sake brewed with this mutant contained baker’s yeast with Pro accumulation has promise for the frozen-
more Pro but less succinate than the sake brewed with the dough baking industry (Figure 3c). Compared to baker’s yeast
parent strain. A metabolome analysis indicated that the de- strains that accumulated only Pro or only trehalose, strains that
crease in succinate was due to a lower level of 2-oxoglutarate, simultaneously accumulated Pro and trehalose showed higher
which is converted into Glu. This approach could be a practical fermentation ability in frozen dough (Sasano et al. 2012a). An-
method for breeding yeast strains to increase the diversity of other study examined the results of inoculating baker’s yeast
sake flavors. cells into a liquid fermentation medium that contained a high
Yeast strains for baking need high fermentation ability and level of sucrose. Under this stress condition, the level of ROS
durability in response to various baking methods. Baker’s yeast increased (Sasano et al. 2012d). This was also observed with
cells are exposed to baking-associated stresses including high wine yeast (Landolfo et al. 2007). Pro-accumulating SC diploid
sucrose levels, freeze-thawing, and air-drying, all of which are baker’s yeast strains were constructed by Sasano et al. (2012d),
believed to induce oxidative stress to the yeast cells (Attfield and their findings demonstrated that Pro accumulation gives
1997; Shima and Takagi 2009). This oxidative stress may be yeast cells greater tolerance to high-sucrose stress. When they
attributable to the denaturation of proteins (eg antioxidant evaluated baker’s yeasts that are used in sweet dough, the Pro-
enzymes) and severe damage to the mitochondrial membrane accumulating strain showed an approx. 40% increase in gassing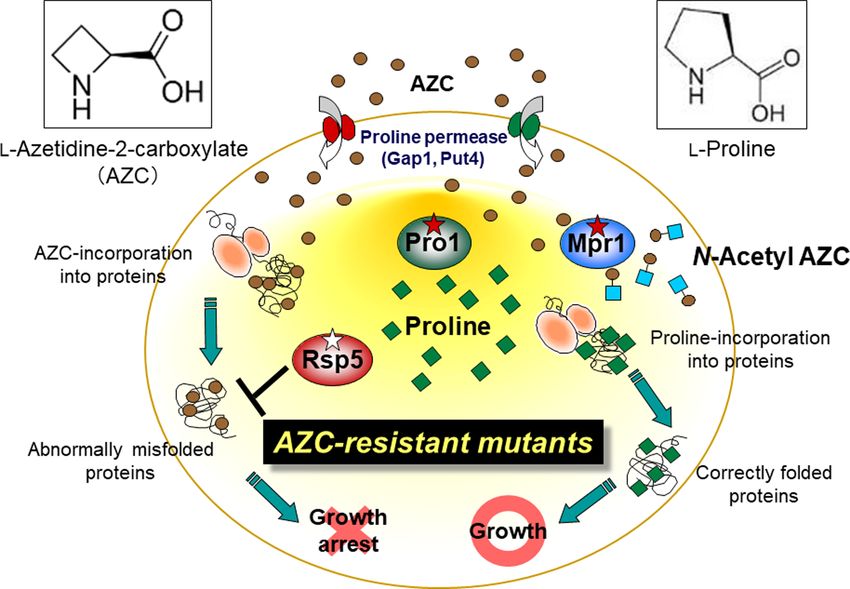
1022 Bioscience, Biotechnology, and Biochemistry, 2021, Vol. 85, No. 5
Downloaded from https://academic.oup.com/bbb/article/85/5/1017/6134096 by guest on 14 December 2021
Figure 3. Fermentation abilities of Pro-accumulating sake and baker’s yeast strains. (a, b) Laboratory-scale sake brewing was performed with a sake mash consisting
of 160 g of steamed rice, 40 g of koji rice, and 260 mL of water added in 3 steps. Each strain was grown in SD medium at 30 °C for 2 days under static conditions
and inoculated into the mash. Fermentation profiles were monitored by a loss of weight in conjunction with CO2 evolution. The sake mash was centrifuged and
the supernatant was obtained as sake. General components of the sake, including Pro (a) and ethanol (b), were analyzed by the standard method established by the
Japanese National Tax Administration Agency. The values are the means and standard deviation of results from 3 independent experiments (Takagi et al. 2007). (c) The
doughs were prefermented for 120 min at 30 °C and then frozen for 9 days at −20 °C. The frozen dough was thawed for 30 min at 30 °C, and the remaining CO2 gas
production was measured. The gassing power before freezing was defined as 100%. The values are the means and standard deviation of results from 3 independent
experiments (Kaino et al. 2008). (d) Fermentation ability in sweet dough (30% sucrose per weight of flour) was monitored by CO2 gas production. The total amounts of
CO2 production after 2 h were measured. The gassing power of wild-type strain (WT) was defined as 100%. The values are the means and standard deviation of results
from 3 independent experiments (Sasano et al. 2012d).
power compared to the wild-type strain (Figure 3d). Thus, Pro- tance to AZC in the yeast cells as well as desensitization to the
accumulating baker’s yeast strains are superior for the pro- Pro feedback inhibition of GK, leading to an intracellular accu-
duction of sweet bread. Dried yeast is exposed to air-drying mulation of Pro. Compared to the baker’s yeast cells that ex-
stress during the preparation process. Air-drying stress is ac- pressed the wild-type PRO1 gene, baker’s yeast cells expressing
tually a combination of 2 stresses: high temperature and de- the PRO1P247S or PRO1E415K gene demonstrated higher tolerance
hydration, which cause the accumulation of intracellular ROS to freezing stress. This approach could thus be effective for
(Franca, Panek and Eleutherio 2007). Air-drying also presents breeding Pro-accumulating baker’s yeast strains that have
disadvantages that reduce fermentation ability, including mito- higher tolerance to baking-associated stresses.
chondrial malfunction, vacuolar acidification, and protein mis- Figure 4 shows the positions of amino acid residues in GK
folding (Shima, Ando and Takagi 2008). Tolerance to air-drying where mutations occur in the Pro-accumulating strains.
stress is thus a necessary characteristic of the baker’s yeast used
in dried yeast preparations. We demonstrated that fermenta-
tion ability of baker’s yeast after air-drying was significantly en- The molecular functions of the novel
hanced by the accumulation of Pro in the cells (Sasano et al. N-acetyltransferase Mpr1 found in yeast
2010).
The methods used for breeding baker’s yeasts are generally When Arg was used as the sole source of nitrogen in yeast, the
more acceptable to consumers since SC yeasts do not need to fermentation rate was increased more than when ammonium
be treated the same as GM yeasts. Mutant yeast strains that are sulfate or Glu used as the sole source (Gutiérrez et al. 2012); the
resistant to the Pro analogue AZC were isolated from diploid addition of Arg also increased the fermentation rate (Thomas
baker’s yeast (Tsolmonbaatar et al. 2016), and under freezing or and Ingledew 1992). These findings suggest that the synthesis of
high-sucrose stress conditions some of these strains that accu- Arg promotes fermentation. In a study of the effects of amino
mulated Pro showed higher cell viability compared to the par- acids on freezing stress applied to yeast cells, Arg and other
ent wild-type strain. It also appeared that most of the mutant charged amino acids (Glu and Lys) provided higher freeze tol-
Pro-accumulating strains had enhanced fermentation ability in erance (Takagi, Iwamoto and Nakamori 1997). The mechanisms
frozen and sweet doughs. Two of those strains carried novel mu- underlying cryoprotective effect of Arg are not yet clear. Arg may
tations in the PRO1 gene encoding the Pro247Ser or Glu415Lys serve as an ion coating on the membrane components’ and pro-
variant of GK. Interestingly, these mutations produced resis- teins’ surfaces that prevents denaturation by the NH2 group in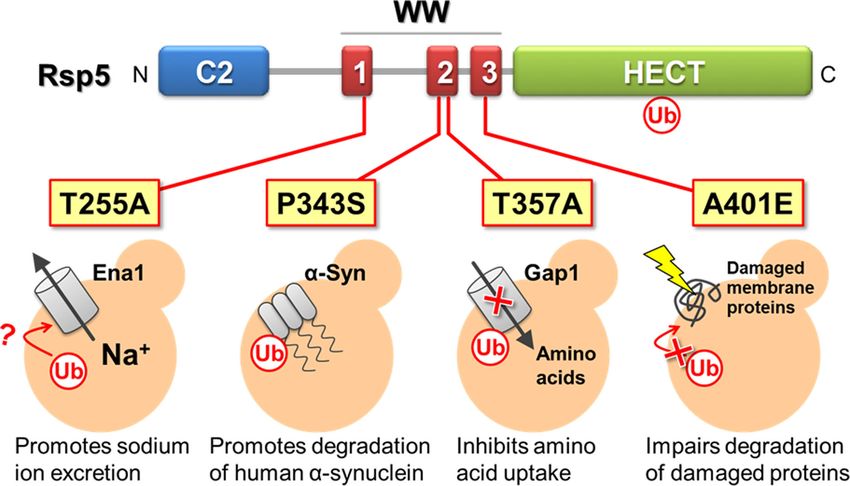
Stress tolerance mechanisms in S. cerevisiae 1023
Downloaded from https://academic.oup.com/bbb/article/85/5/1017/6134096 by guest on 14 December 2021
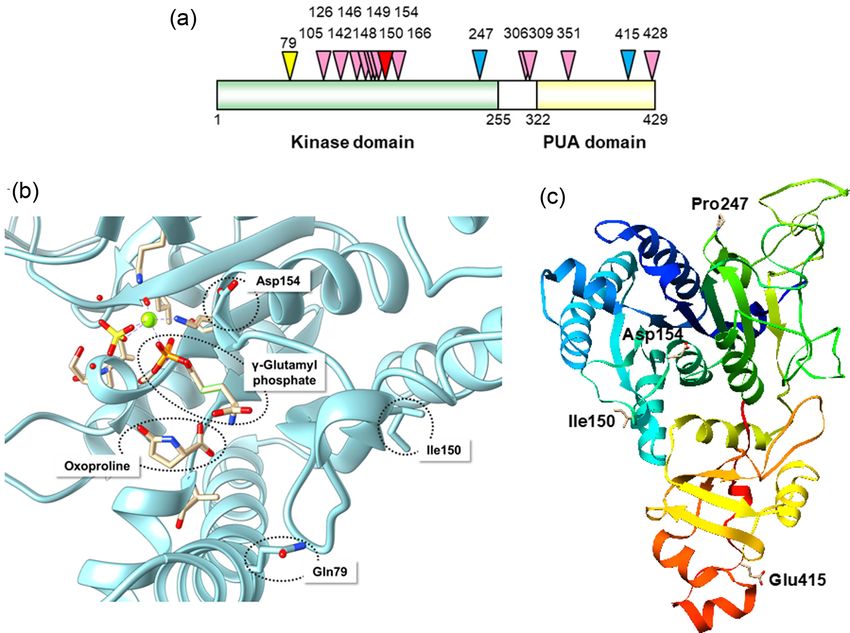
Figure 4. Features of GK variants. (a) Schematic representation of the S. cerevisiae GK. The positions of amino acids where mutations occur in the Pro-accumulating
mutants are indicated by arrowheads. The numbers are residue numbers. The kinase and PUA domains are green and yellow, respectively. (b) Homology model of S.
cerevisiae GK (Pro1, UniProtKB accession number P32264) was constructed by SWISS-MODEL using the structure of Escherichia coli GK bound with γ -glutamyl phosphate
and oxoproline (39% sequence identity, PDB ID: 2J5V) as the template. Asp154, Ile150, and Gln79 (Sekine et al. 2007; Murakami et al. 2020) are shown in a stick model.
(c) The structure of S. cerevisiae GK was predicted by homology modeling on I-TASSER based on the structure of E.coli GK (PDB ID: 2j5t) as the template. Asp154, Ile150,
Pro247, and Glu415 (Sekine et al. 2007; Tsolmonbaatar et al. 2016) are shown in a stick model. Figure of GK structure was drawn using UCSF chimera. Some residues or
the substrate CHOP are shown in a stick model or sphere model, respectively.
the molecules. The effect of intracellular charged amino acids (GSA) to provide N-acetyl-GSA, an intermediate of the Arg
on freeze tolerance in dough was examined by constructing biosynthesis pathway (Nishimura et al. 2010) (Figure 5b), thereby
homozygous diploid arginase-deficient mutants of commercial promoting the presence of N-acetyl-GSA in yeast. These find-
baker’s yeast (Shima et al. 2003). During the frozen-dough baking ings suggested that an Mpr1-mediated enhancement of Arg
process, one of the arginase mutants accumulated higher levels synthesis could enable the construction of new yeast strains
of Arg and/or Glu and showed increased leavening ability, sug- with better fermentation rates.
gesting that freeze tolerance is enhanced by the disruption of The results of our crystallographic analysis revealed that
the CAR1 gene. the overall structure of Mpr1 is based on folding that is typi-
During my investigations of Pro, I noticed the N- cal among the proteins in the Gcn5-related N-acetyltransferase
acetyltransferase Mpr1, which uses cyclic secondary amines superfamily (Figure 5c) (Nasuno et al. 2013). Mpr1 is folded into
(AZC and cis-4-hydroxy Pro) as substrates (Figure 5a). Among an α/β structure with eight-stranded mixed β-sheets and six α-
the many laboratory strains of Saccharomyces, the 1278b back- helices. The substrate binds to Asn135 and the backbone amide
ground strain has 2 copies of the MPR gene, MPR1 and MPR2 of Asn172 and Leu173, and the predicted acetyl-CoA (AcCoA)
(sigma 1278b gene for proline-analogue resistance). A single binding site is located near the backbone amide of Phe138 and
base substitution exists at position 254, leading to Gly and Glu the side chain of Asn178 (Figure 5d). An alanine substitution of
at position 85 in MPR1 and MPR2, respectively; nevertheless, the Asn178, which can interact with the sulfur of AcCoA, markedly
gene products (Mpr1 and Mpr2) play similar roles in resistance reduced the apparent kcat value. In addition, the replacement
to AZC (Takagi, Iwamoto and Nakamori 1997, 2000a, 2000b). of Asn135 led to a markedly increased apparent Km value.
The Mpr1 protein was revealed as a novel N-acetyltransferase Together these results indicated that Asn178 plays an important
that detoxifies AZC through its N-acetylation in the S. cerevisiae role in catalysis, and Asn135 has an important role in substrate
1278b strain (Shichiri et al. 2001). The genomes of various recognition (Nasuno et al. 2013). Structure-based molecular
yeasts and fungi have been shown to contain genes that are design enabled the creation of the 2 stable variants (ie
homologous to MPR1, and AZC acetyltransferase activity has Asn203Lys-Mpr1 and Asn203Arg-Mpr1) each of which exhib-
been observed in many types of yeast. Mpr1 homologues thus ited longer half-life activity compared to the wild-type Mpr1
appear to be widely conserved in yeasts and fungi (Kimura, (Nasuno et al. 2016). Polymerase chain reaction (PCR) random
Nakamori and Takagi 2002; Wada et al. 2008). Interestingly, our mutagenesis was used to isolate the Phe65Leu variant, which
research group has demonstrated the involvement of Mpr1 in led to a further stabilization of Mpr1 (Iinoya et al. 2009). In
a novel Arg biosynthesis pathway, and we observed that Mpr1 growth assays, the overexpression of stable Mpr1 variants in
may acetylate P5C or its tautomer l-glutamate-γ -semialdehyde yeast cells appeared to increase the synthesis of Arg. The con-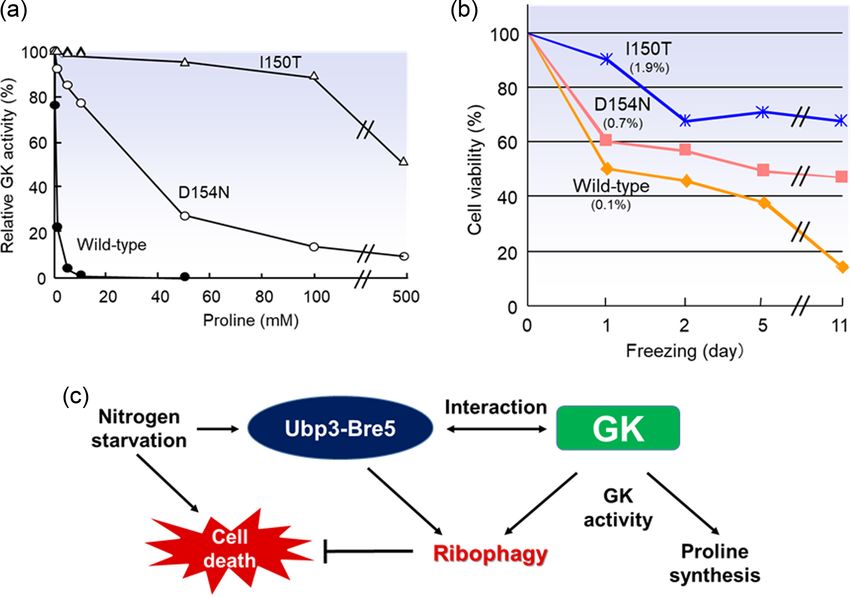
1024 Bioscience, Biotechnology, and Biochemistry, 2021, Vol. 85, No. 5
Downloaded from https://academic.oup.com/bbb/article/85/5/1017/6134096 by guest on 14 December 2021
Figure 5. Molecular profiles of Mpr1 found in S. cerevisiae. (a) Proposed scheme for the AZC acetyltransferase reaction by Mpr1. cis-4-Hydroxy Pro (CHOP) is also acetylated
in the same manner. (b) Growth curves of yeast strains during cultivation in SD medium at 30 °C. Mpr1 restored growth in arg2-deficient cells, even in the absence of
Arg, but could not compensate the arg8-disruption blocking the downstream pathway of N-acetyl glutamate (Nishimura et al. 2010). (c) The structure of Mpr1-CHOP
(light blue). It is superimposed with that of an aminoglycoside 6 -N-acetyltransferase from Enterococcus faecium (PDB ID code 1B87) (salmon pink). AcCoA in the structure
1B87 and CHOP in the structure of Mpr1-CHOP are shown as a sphere model (Nasuno et al. 2013). (d) Schematic illustration of CHOP binding site on Mpr1. CHOP is
shown by black color, and red letters indicate the carbon atom numbering of CHOP. The red and blue residues interact with the substrates through their side chain and
backbone, respectively. Black dotted lines show the possible van der Waals interaction and hydrogen bonds. CHOP binds to the side chain of Asn135 and the backbone
amide N-H group of Asn172 and Leu173 through its carboxyl group, and to a water molecule bound to Phe138 through its amine group. CHOP also forms van der Waals
contact with the phenolic side chain of Tyr75 through its Cγ atom (Nasuno et al. 2013).
struction of new yeast strains with both higher Arg synthetic exposing the strains to air-drying stress. The results revealed
ability and improved fermentation ability can make use of these that the MPR2 in baker’s yeast is involved in the tolerance to
findings. air-drying stress by reducing the intracellular ROS levels
Mpr1 was reported to protect yeast cells against various (Figure 6a) (Sasano et al. 2010). We also observed that compared
oxidative stress conditions by reducing their intracellular ROS to the wild-type Mpr1, the fermentation ability of bread dough
levels (Nomura and Takagi 2004; Du and Takagi 2005, 2007). after its exposure to air-drying stress was increased by the ex-
High temperatures cause oxidative stress by generating ROS in pression of the Lys63Arg and Phe65Leu variants with enhanced
the mitochondria (Moraitis and Curran 2007), prompting strong enzymatic functions (Figure 6a). The antioxidant enzyme Mpr1
transcriptions of MPR1 and PUT1 and leading to an increased thus has potential for the breeding of novel yeast strains that are
intracellular level of Arg (Nishimura et al. 2010). The increased tolerant to air-drying stress.
conversion of Pro into Arg endowed yeast cells with oxida- We recently investigated the effects of Asn203Lys (a sta-
tive stress tolerance, indicating that an unknown antioxidative ble variant of Mpr1) on the ethanol fermentation of a sake
mechanism is involved in the stress-induced synthesis of Arg yeast strain that lacks the MPR1 gene (Ohashi et al. 2019). The
requiring Mpr1 and Put1 (Nishimura et al. 2010). fermentation performance of this variant was improved com-
pared to the wild-type Mpr1 when the variant was expressed in
The stress tolerance and fermentation ability of the diploid Japanese sake strain (Figure 6b). In laboratory-scale
industrial yeasts with Mpr1 expression brewing, a sake strain that expresses the Asn203Lys variant pro-
duced more ethanol than the wild-type Mpr1. The Asn203Lys
Interestingly, among the many yeast strains used in industry, the variant also affected the contents of flavor compounds and or-
Japanese baker’s yeast strains have 1 copy of the MPR2 gene on ganic acids. This stable Mpr1 variant could therefore contributes
chromosome X (Sasano et al. 2010). For an examination of the to the construction of new industrial yeast strains with im-
role of MPR2 in baker’s yeast, we tested cell viability of diploid proved fermentation ability and diversity of taste and flavor.
industrial baker’s yeast strains and intracellular ROS level after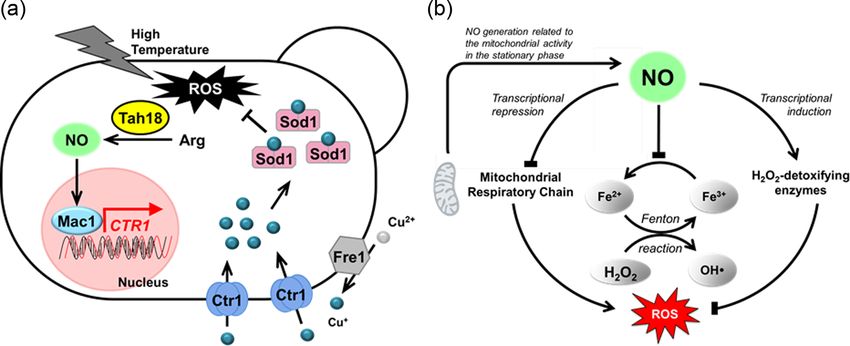
Stress tolerance mechanisms in S. cerevisiae 1025
Downloaded from https://academic.oup.com/bbb/article/85/5/1017/6134096 by guest on 14 December 2021
Figure 6. Effects of the wild-type and variant of Mpr1 on baker’s and sake yeast strains. (a) Fermentation ability of baker’s yeast strains after air-drying. Compressed
yeast was treated with air-drying stress for 4 h at 37 °C. The dough containing the stress-treated yeasts was fermented for 3 h, and the remaining CO2 gas production
was measured. The amount of CO2 production of the wild-type strain (WT) after air-drying stress treatment was defined as 100%. The values are the means and
standard deviation of results from 3 independent experiments (Sasano et al. 2010). (b) Total CO2 emission (L) of sake yeast strain harboring the empty vector (EV),
the wild-type Mpr1 (WT), and the Asn203Lys variant Mpr1 (N203K) in SC20-ura medium at 25 °C. The values are the means and standard deviation of results from 4
independent experiments. Statistically significant differences between N203K and WT or EV were determined by Student’s t-test (*P < .05) (Ohashi et al. 2019).
The physiological roles of nitric oxide in yeast Cytoprotective actions exerted by NO have been reported
in yeast cells subjected to a variety of environmental stress
The cellular signaling molecule nitric oxide (NO) is widely con- conditions, particularly oxidative stress. This cytoprotection is
served among organisms, including microorganisms and higher achieved by NO’s regulation of certain transcriptional factors,
eukaryotes. As a free radical gaseous molecule, NO participates including the S. cerevisiae Mac1 (Nasuno et al. 2014), the fis-
in many biological processes in mammals such as protec- sion yeast Schizosaccharomyces pombe Pap1 (Kang et al. 2011), Sty1
tion against pathogens and the regulation of blood pressure (Astuti, Watanabe and Takagi 2016), and Rst2 (Kato, Zhou and
(Astuti, Nasuno and Takagi 2018). NO has dual, concentration- Ma 2013), which modulate different mechanisms. For example,
dependent functions in both unicellular organisms and plants, the activation of Mac1 by NO that was produced under a high-
and in bacteria, a low nanomolar cellular level of NO contributes temperature-stress condition was shown to be important for
to the self-defense against oxidative stress (Gardner et al. 2000). the activation of the copper-dependent superoxide dismutase
The formation of the biofilm is regulated by NO, thus promoting Sod1, which is a crucial antioxidative enzyme (Nasuno et al. 2014)
the formation of the bacterial colony (Allain et al. 2011). NO (Figure 7a). In S. pombe, NO is localized in the mitochondria at the
also has a role in the synthesis of toxins, and at higher levels stationary phase, suggesting that 2 distinct types of NO signaling
NO can be toxic and induce cell death (Poole 2005). In yeasts, a exist. For mitochondria, pretreatment with an NO donor rescued
low NO level mediates resistance to oxidative and temperature the cell growth by repressing the generation of ROS under oxida-
stress (Nishimura, Kawahara and Takagi 2013; Nasuno et al. tive stress. The results of a DNA microarray analysis of S. pombe
2014; Liu et al 2015). At high micromolar levels, NO can enhance demonstrated that exogenous NO contributed to the yeast’s tol-
oxidative and nitrosative stress (Almeida et al. 2007; Tillmann, erance to H2 O2 (hydrogen peroxide) by (i) inhibiting the conver-
Gow and Brown 2011). A low level of NO is also important for sions of Fe3+ to Fe2+ , (ii) upregulating the H2 O2 -detoxifying en-
the resistance to oxidative stress in plants (Considine, Sandalio zymes, and (iii) downregulating the mitochondrial respiratory
and Foyer 2015). Based on its high diffusibility and reactivity, chain (MRC) genes. These findings indicated that NO plays a piv-
NO exerts opposing effects in cells, “acting as a double-edged otal role in the negative feedback system to regulate ROS levels
sword”; for example, many physiological aspects of cardiovascu- in S. pombe (Figure 7b) (Astuti, Watanabe and Takagi 2016).
lar protection involve NO by activating a cGMP-mediated signal In S. cerevisiae, NO has a pivotal role in the tolerance
transduction pathway, and NO also enhances the tolerance of of cells to high temperatures, heat shock, and hydrostatic
mammals and plants to oxidative stress by amplifying the cellu- pressure imbalance (Domitrovic et al. 2003). Our research
lar antioxidative activity (Petrovic et al. 2008; Martin et al. 2009). has demonstrated that an increased NO level conferred tol-
However, at higher NO levels or in the presence of superoxide, erance to high-temperature stress (Nishimura et al. 2013;
NO can have cytotoxic effects, potentially through the formation Nasuno et al. 2014). The activity of NOS is repressed by
of the oxidant peroxynitrite, which induces cell damage via the loss of Cka2, which further decreases the cell viabil-
lipid peroxidation and protein inactivation by oxidation and ity under H2 O2 -induced apoptosis or high-temperature stress
nitration, leading to cardiovascular dysfunction. NO can react (Liu et al. 2015). Calorie restriction in S. cerevisiae induced a
with other radical molecules, and this reaction generates more mitochondria-dependent synthesis of NO, leading to a metabolic
reactive secondary products, including reactive nitrogen species shift that improved the cellular metabolism and the cellular re-
(RNS). The RNS can react with proteins that have free sulfhydryl sponse to an age-related accumulation of oxidative stress (Li
groups at cysteine residues, and they react with tyrosine et al. 2011). Those findings indicated that NO may help combat
residues or with metals and heme to perform nitrosylation and the effects of aging and age-related diseases. In addition, a com-
nitration processes (Ridnour et al. 2004). Nitrosylation influences prehensive study using a yeast model revealed the pathology of
the activities of proteins, protein–protein interactions, and the Batten disease, a fatal disease of the nervous system that is the
localization of proteins (Benhar, Forrester and Stamler 2009).
1026 Bioscience, Biotechnology, and Biochemistry, 2021, Vol. 85, No. 5
Downloaded from https://academic.oup.com/bbb/article/85/5/1017/6134096 by guest on 14 December 2021
Figure 7. Proposed models for NO-mediated oxidative stress responses in yeast. (a) When S. cerevisiae cells are exposed to high-temperature stress, NO is pro-
duced from Arg by an NOS-like activity. NO-mediated Mac1 is activated by post-translational modification, then upregulates the expression of CTR1, which
encodes a copper transporter. Protein names: Fre1, cupric reductase; Ctr1, copper ion transporter; Sod1, Cu, Zn-superoxide dismutase; Ccs1, copper chaperone; Mac1,
transcription factor responsible for copper metabolism (Nasuno et al. 2014). (b) In S. pombe, NO is mainly generated by the activities of an unidentified NOS and the
MRC complex III during log phase growth and is detoxified by Yhb1 (NOD) and Fmd2 (GSNOR) in stationary phase. NO triggers at least 3 antioxidant mechanisms: (i)
inhibition of the conversion of Fe3+ into Fe2+ (ii) upregulation of the genes encoding H2 O2 -detoxifying enzymes, and (iii) downregulation of the MRC genes (Astuti et al.
2016).
result of reduced NO production due to the loss of Cln3, a pro- the induction of these 2 genes led to a significant increase in
tein that is important for the maintenance of the intracellular the level of Arg (ie the NOS substrate), especially in response to
Arg level and pH homeostasis (Osório et al. 2007). environmental stressors such as air-drying, high temperatures,
and freeze-thawing (Sasano et al. 2012c; Nishimura, Kawahara
and Takagi 2013). Despite the finding that TAH18 expression is
The synthetic regulations of nitric oxide in yeast required for NOS-like activity in S. cerevisiae, the details of Tah18-
associated NOS activity are not yet clear; this is because al-
Living organisms use 2 major mechanisms for NO production: though Tah18 is homologous to the reductase domain, the Tah18
(i) the Arg pathway catalyzed by NO synthases (NOSs) and (ii) protein does not contain an intact oxygenase domain of NOS.
the nitrite pathway, which provides a simple way to produce Tah18 transfers electrons from NADPH to the Fe-S clusters of
NO through the electron reduction of nitrate catalyzed by ni- the Dre2 protein via flavin molecules (FAD and FMD), leading to
trate reductase (NR) and nitrite reductase (NIR). The NO in mam- the biogenesis of Fe-S cluster proteins (Netz et al. 2010). Our bio-
malian cells is endogenously synthesized from Arg by the enzy- chemical analyses recently showed that in response to oxidative
matic reaction of NOSs. Arg is oxidized to form l-citrulline (Cit) stress Tah18 dissociates from Dre2, accompanied by an increase
and NO, and NADPH and oxygen are involved in the reaction. in the intracellular NO level. We also observed that under oxida-
Bacteria can also generate Arg-dependent NO through NOS sys- tive stress conditions the enhancement of the Tah18–Dre2 in-
tems. NOS-like proteins have been identified in many prokary- teraction suppressed the production of NO, indicating that the
otes based on their genome sequences (Chen and Rosazza 1995; interaction of Tah18 with Dre2 inhibits the Tah18-dependent
Adak, Aulak and Stuehr 2002; Hong et al. 2003; Kers et al. 2004; NOS-like activity (Yoshikawa et al. 2016). These findings suggest
Gusarov et al. 2008; Agapie et al. 2009). Bacterial NOSs (bNOSs) that (i) Tah18 may acts as a reductase domain to transfer elec-
are comprised of only an oxygenase domain, unlike mammalian trons from NADPH to an unidentified oxygenase protein, which
NOSs (Adak, Aulak and Stuehr 2002; Gusarov et al. 2008; Bird oxidizes Arg to Cit and NO, and (ii) Dre2 inhibits the Tah18-
et al. 2009). It is believed that evolutionarily, eukaryotic NOS orig- dependent NOS-like activity by removing electrons derived ex-
inated earlier than bNOS, and that bNOS evolved to use a dedi- clusively from NADPH. The Tah18-Dre2 complex might there-
cated reductase domain (Gusarov et al. 2008). However, the bNOS fore function as a molecular switch to control NO production in
of the soil-dwelling Gram-negative bacterium Sorangium cellulo- response to environmental oxidative conditions (Figure 8)
sum have both an oxygenase domain and a reductase domain, (Astuti, Watanabe and Takagi 2016; Astuti, Nasuno and Takagi
providing a new indication that the NOS present in both eukary- 2018).
otes and prokaryotes evolved from a common ancestor (Agapie Physiological functions and metabolism of NO in yeast
et al. 2009). remain to be clarified. A new method for obtaining more precise
The generation of NO via NOS-like activity in S. cerevisiae and measurements of the NO content in S. cerevisiae cells with
S. pombe has been observed in both in vitro and in vivo anal- the detection limit of 6 nm was recently developed in our
yses (Almeida et al. 2007; Kig and Temizkan 2009; Yoshikawa laboratory. The measurements are achieved by first treating
et al. 2016), but the corresponding NOS-encoding gene has not the cells with an NO-specific fluorescence probe, and the
been identified. Our laboratory revealed the generation of NO performing high-performance liquid chromatography with
in S. cerevisiae via an Arg-dependent mechanism that is medi- fluorescence detection (HPLC/FLD) (Nasuno et al. 2020). With
ated by the flavoprotein Tah18 (Nishimura, Kawahara and Takagi this method we successfully quantified the NO content inside
2013) (Figure 1). We observed that in laboratory and baker’s yeast yeast cells that were treated with an NO donor. Other results of
strains, this particular Tah18-dependent NO generation was as- this HPLC/FLD analysis indicated that the fluorescence that is
sociated with the activities of MPR1 and PUT1 in the Pro-Arg induced under some stress conditions (eg heat-shock, ethanol,
metabolic pathway. This association is based on the following: and vanillin treatment) was not derived from NO. The HPLC/FLDStress tolerance mechanisms in S. cerevisiae 1027
We recently identified a novel nitrosative stress tolerance
gene in S. cerevisiae, RIB1, which encodes GTP cyclohydrolase II
(GTPCH2) (Anam, Nasuno and Takagi 2020). The first step in ri-
boflavin biosynthesis is catalyzed by GTPCH2. The GTPCH2 en-
zymatic activity of Rib1 is essential for the nitrosative stress
tolerance that is RIB1-dependent, but riboflavin itself is not
required for this tolerance. We also observed that the reac-
tion mixture of recombinant purified Rib1 quenched NO or
its derivatives, whereas formate or pyrophosphate (byproducts
of the Rib1 reaction) did not. These findings suggested that
the reaction product of Rib1, ie 2,5-diamino-6-(5-phospho-d-
ribosylamino)-pyrimidin-4(3H)-one (DARP), scavenges NO or its
derivatives. Our analyses demonstrated that 2,4,5-triamino-1H-
pyrimidin-6-one, which is identical to a pyrimidine moiety of
Downloaded from https://academic.oup.com/bbb/article/85/5/1017/6134096 by guest on 14 December 2021
DARP, also scavenged NO or its derivatives, suggesting that
DARP reacts with the dinitrogen trioxide (N2 O3 ) generated via
its pyrimidine moiety. Since there has been no report that
DARP or another riboflavin metabolism intermediate scavenges
RNS or attenuates nitrosative stress, we identified RIB1/DARP
for the first time as a novel gene/metabolite that functions in
the induction of the nitrosative stress tolerance mechanism in
Figure 8. Dual effects of NO in S. cerevisiae. Like mammalian cells, Tah18- S. cerevisiae.
dependent NO production exhibits 2 opposed effects in yeast cells. For exam-
ple, appropriate NO level confers tolerance to high temperature to cells (Nasuno
et al. 2014). In contrast, under severe stress conditions, such as high levels of
Biotechnological applications of nitric oxide in yeast
H2 O2 , excess NO induces cell death (Yoshikawa et al. 2016).
NO signaling is known to be involved in the pathology of sev-
eral natural processes (eg aging) and degenerative diseases (eg
method measures the intracellular NO concentration with
neurodegenerative syndromes, cancer) (Khurana and Lindquist
higher accuracy than has been available in the past. We also
2010). Investigations of yeast may help reveal the underlying
developed an analytical method to identify the nitrogen source
mechanisms of these NO-related diseases, as a model of degen-
for NO generation; this method uses liquid chromatography
erative diseases, mitochondria-related diseases (Lasserre et al.
with tandem mass spectrometry and stable isotope labeling
2015), and aging phenomena (Li et al. 2011). The findings regard-
(Nasuno, Yoshikawa and Takagi 2021). We detected a 15 N-labeled
ing the involvement of NO in the stress response of industrially
NO-containing compound generated by a 15 N-labeled substrate
beneficial yeasts suggests that yeast cells could be modified by
nitrite both in vitro and in vivo by using this new method.
NO-targeted engineering at both the genetic and physiological
levels.
The detoxification systems of nitric oxide in yeast Modifications of the synthesis of NO designed to enhance the
fermentation ability of baker’s yeast have been described. For
Nitrosative stress is induced by the accumulation of NO deriva-
example, Sasano et al. (2012c) engineered a SC diploid baker’s
tives, especially RNS (Ridnour et al. 2004). Several RNS molecules,
yeast strain with enhanced Pro and NO synthesis that also
such as peroxynitrite, the higher oxides of nitrogen, S-
shows an increased intracellular NO level in response to air-
nitrosothiols (RSNO), and dinitrosyl iron complexes, are reported
drying stress; the strain better tolerates air-drying, oxidative,
to have roles in cellular systems (Heck 2001). The mechanisms
and freeze-thawing, which may be due to the strain’s reduced in-
of intracellular NO detoxification are important for ensuring that
tracellular ROS level. Compared to the wild-type strain, the new
NO can exert its cytoprotective actions rather than cytotoxic ef-
strain retained higher leavening activity in bread dough after air-
fects on cells. The maintenance of intracellular NO homeostasis
drying and freeze-thawing. These characteristics suggest that
involves S-nitrosoglutathione reductase (GSNOR) and NO dioxy-
(i) NO is synthesized in baker’s yeast in response to oxidative
genase (NOD) (Gardner 2005). S-nitrosoglutathione (GSNO) is of-
stresses, and (ii) increased NO is important in baking-associated
ten used as a marker of the intracellular level of RSNO, because
stress tolerance.
glutathione (GSH, the substrate for GSNO synthesis) is the most
abundant thiol-containing compound in eukaryotic cells. NOD
(or flavohemoglobin) mediates deoxygenation by using the re- The protein quality-control mechanism used
ductive power of NAD(P)H to covalently bond the 2 oxygen atoms by the ubiquitin system under stress
in O2 with NO, yielding NO3 − (Gardner 2005).
conditions
Our latest study of S. pombe cells demonstrated that
as NO-detoxification enzymes, the putative NO dioxygenase One of the fundamental cellular processes for regulating cellular
SPAC869.02c (named Yhb1) and the GSNO Fmd2 cooperatively activities at the post-translational level is protein ubiquitination,
reduced intracellular levels of NO (Astuti, Watanabe and Tak- achieved through protein modification by the small regulatory
agi 2016). Yhb1 is likely NOS-dependent, whereas Fmd2 is NOS- protein ubiquitin (Ub), a highly conserved protein that contains
independent and may be associated with a mitochondrial- 76 amino acids. Ubiquitination has roles in a variety of cellular
dependent production of NO. These two NO-detoxification processes such as regulation of the cell cycle and apoptosis, pro-
enzymes are conserved among eukaryotes from yeast to mam- tein degradation and trafficking, and an array of signal transduc-
mals, indicating that NO is likely to be involved in primary phys- tions (Mukhopadhyay and Riezman 2007). The ubiquitination
iological activities that are conserved throughout evolution. process occurs via a multistep cascade. As the first step, Ub is1028 Bioscience, Biotechnology, and Biochemistry, 2021, Vol. 85, No. 5
activated by the E1-activating enzyme in an ATP-dependent Repair and degradation mechanisms of stress-induced
manner. The formation of a thioester bond between the C- abnormal proteins mediated by Rsp5
terminal glycine residue of Ub and a cysteine in the active center
of E1 then takes place. The activated Ub is transferred to a cys- Cells have a quality-control mechanism for proteins that are
teine residue located at the active center of the E2-conjugating damaged by stress; that is, a repair and degradation mechanism.
enzyme. The next step is the prompt transfer of the active Ub to To investigate cellular functions under various stress conditions
the target protein with the aid of the E3 Ub ligase. As the process during the fermentation processes, our research group isolated a
continues, the ligation of Ub with the target protein occurs via an laboratory AZC-hypersensitive mutant yeast strain that is sen-
isopeptide bond formed between the C-terminal glycine residue sitive to various stresses (Hoshikawa et al. 2003). This mutant
of Ub and the lysine residues of the target protein (or another carries an allele of RSP5, leading to a single-amino acid change
Ub, if a poly-Ub chain is formed). Ub ligases can be categorized within the WW3 domain (Ala401Glu). The addition of NH4 + to
into 2 types: the RING (really interesting new genes)-finger type yeast cells growing on induced rapid ubiquitination, endocy-
(Jackson et al. 2000) and the HECT (homologous to E6-AP carboxyl tosis and the vacuolar degradation of the plasma membrane
terminus) type (Huibregtse et al. 1995). The many HECT-type Ub protein (the general amino acid permease Gap1). The Gap1 in
the rsp5A401E mutant remained stable and active on the plasma
Downloaded from https://academic.oup.com/bbb/article/85/5/1017/6134096 by guest on 14 December 2021
ligases in S. cerevisiae include Hul4, Hul5, Ufd4, Tom1, and Rsp5
(Wang, Yang and Huibregtse 1999). The sole essential HECT-type membrane (and probably without ubiquitination), leading to an
Ub ligase in the Nedd4 family found in S. cerevisiae is Rsp5. intracellular accumulation of AZC. Interestingly, the rsp5A401E
mutant also showed hypersensitivity to various stressors that
induce protein misfolding: ethanol, heat shock, high tempera-
ture in a rich medium, toxic amino acid analogs, and oxidative
The ubiquitin ligase Rsp5 treatments (Hoshikawa et al. 2003). Based on these findings, we
speculated that Rsp5 is involved in a selective degradation of
Rsp5 (Reverses Spt− phenotype protein 5) is originally isolated as
stress-induced abnormal proteins, and in a nitrogen-regulated
a mutated gene that suppresses SPT3 mutations that negatively
degradation of Gap1. In general, the accumulation of stress-
affect the gene expression (Winston 1993). Rsp5 is the orthologue
induced abnormal proteins poses a serious challenge for cells.
of the human Nedd4 Ub ligase in S. cerevisiae, and it is thus spec-
Two strategies to address such an accumulation can be consid-
ulated to play important roles in signal transduction (Dunn and
ered: degradation in the proteasome or vacuole, or refolding by
Hicke 2001), the quality control of plasma membrane proteins
molecular chaperones, such as stress proteins. It is of interest to
(Shiga et al. 2014), and intracellular trafficking (Jarmoszewicz
determine whether or how Rsp5 is involved in these processes.
et al. 2012). Rsp5 is composed of an N-terminal Ca2+ -dependent
We observed that when the rsp5A401E mutant was exposed
phospholipid membrane binding (C2) domain, 3 substrate recog-
to ethanol, a temperature up-shift, or sorbitol, significantly less
nition (WW) domains (commonly referred to 2 conserved tryp-
transcription of stress protein genes occurred compared to that
tophan residues in the domains), and the C-terminal catalytic
in the wild-type strain (Haitani, Shimoi and Takagi 2006). The
(HECT) domain (Rotin and Kumar 2009). The C2 domain of Rsp5,
amounts of the transcription factors Hsf1 and Msn4 in the
comprised of ∼130 amino acids, is normally regulated by the
rsp5A401E mutant were remarkably defective, suggesting that (i)
Ca2+ ions. This domain is important for the ability of Rsp5 to
expression of stress proteins are mediated by Rsp5 and (ii) Rsp5
electrostatically interact with either the endosomal membrane
primarily regulates the post-translational modification of Hsf1
or plasma membrane (Cho 2001). Each of the WW domains of
and Msn4. After rsp5A401E cells were exposed to ethanol and a
Rsp5 consists of ∼40 amino acid residues, as Sudol (1996) ini-
temperature upshift, the protein levels of Hsf1 and Msn2/4 were
tially described. The WW domains interact with substrates or
remarkably defective in the cells, although these proteins were
adaptor proteins by recognizing proline-rich sequences (PY mo-
localized mainly in the cell nucleus under these stress condi-
tifs; Pro-Pro-X-Tyr) within these proteins. The three WW do-
tions (Haitani and Takagi 2008). The mRNAs of HSF1 and MSN2/4
mains in Rsp5 are WW1, WW2, and WW3. The HECT domain
also accumulated in the nuclei of rsp5A401E cells, indicating that
is composed of ∼350 amino acid residues and accounts for the
Rsp5 may be required for the nuclear export of these mRNAs.
ubiquitination activity of Rsp5. The important residue in this
Rsp5 primarily regulates the expressions of these 2 transcrip-
catalytic domain is Cys777, which is the location for thioester
tion factors at the post-transcriptional level and is involved in
formation with ubiquitin. The mutation of Cys777 to Ala thus
the repair of stress-induced abnormal proteins (Figure 9a).
abolishes the overall ubiquitination activity of Rsp5 (Huibregtse
We conducted a comparative proteome analysis of the
et al. 1995). The mutation of Leu733 to Ser, known as rsp5-1 (a
rsp5A401E mutant under stress conditions to identify the pro-
temperature-sensitive allele) was reported to cause a defect in
tein substrates of Rsp5, and we observed that several proteins,
the Ub-thioester formation and decrease the stability of Rsp5 at
including the alpha subunit of nascent polypeptide-associated
restrictive temperatures (Wang, Yang and Huibregtse 1999).
complex (αNAC; Egd2) were accumulated in this mutant
Rsp5 must interact with the substrate through an interaction
(Hiraishi et al. 2009b). Interestingly, under the stress conditions
between its WW domains and the PY motifs located in the sub-
used, Egd2 was ubiquitinated in the wild-type cells but not in the
strate, but most of the substrates for Rsp5 do not contain PY mo-
rsp5A401E mutant cells. At elevated temperature, we also detected
tifs (Gupta et al. 2007). Thus, Rsp5 may interact with a substrate
the in vitro ubiquitination of Egd2 by Rsp5. The ubiquitination of
via non-PY motifs or with the assistance of adaptors that con-
Egd2 is independent of the RING-type Ub ligase Not4. Our study
tain a PY-motif, eg the arrestin-related trafficking adaptor (Art)
also demonstrated that under stress conditions, Egd2 was de-
proteins (Léon and Haguenauer-Tsapis 2009). Yeasts possess 14
graded mainly via the proteasome pathway, which strongly im-
Art proteins (Art1-10, Bul1-3, and Spo23), all of which contain
plies the involvement of Rsp5 in a selective ubiquitination and
the PY motifs recognized by Rsp5. This arrangement functions
degradation of stress-induced unstable proteins, such as Egd2
to direct nonfunctional or misfolded proteins out of the plasma
(Figure 9b).
membrane and prevent their aggregation, thereby maintaining
We also investigated the intracellular trafficking of Gap1
the plasma membrane’s integrity under proteotoxic stress, such
as a model substrate of cell membrane proteins under var-
as heat shock (Zhao et al. 2013).You can also read

















































