Genital coupling and copulatory wounding in the Drosophila auraria species complex (Diptera: Drosophilidae)
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Biological Journal of the Linnean Society, 2022, 135, 195–207. With 7 figures.
Genital coupling and copulatory wounding in the
Drosophila auraria species complex (Diptera:
Drosophilidae)
MOE ONUMA1, , YOSHITAKA KAMIMURA2, and KYOICHI SAWAMURA3,*,
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
1
Doctoral Program in Biology, University of Tsukuba, 1-1-1 Tennodai, Tsukuba, Ibaraki, 305-8572,
Japan
2
Department of Biology, Keio University, 4-1-1 Hiyoshi, Kita, Yokohama, Kanagawa, 223-8521, Japan
3
Faculty of Life and Environmental Sciences, University of Tsukuba, 1-1-1 Tennodai, Tsukuba, Ibaraki,
305-8572, Japan
Received 9 July 2021; revised 27 August 2021; accepted for publication 29 August 2021
Animal genitalia have changed substantially and rapidly during evolution, and functionally interacting anatomical
structures complementarily match between the sexes. Several hypotheses have been proposed to explain how such
structure-matching evolved. A test of these hypotheses would require a detailed analysis of male and female genitalia
among closely related species and a comparison of the functional aspects of the interacting structures between the
sexes. Therefore, here we document genital coupling and copulatory wounds in the four species of the Drosophila
auraria complex. The position of the protrusion of the median gonocoxite of males relative to the female terminalia
differed among the species, which may reflect differences in protrusion morphology. Species-specific female structures
were discovered on the membrane between the genitalia and analia and on the vaginal wall. The former makes
contact with the protrusion, and the latter makes contact with appendages of the aedeagus. Copulatory wounds,
which are produced during copulation, were seen at three locations on females: depressions near the genital orifice,
the membrane between the genitalia and analia, and the vaginal wall. Some of the copulatory wounds were located
at sites that could potentially make contact with the species-specific structures that we identified. We speculate that
the female structures that differ between species of the D. auraria complex evolved in concert with the genitalia of
male conspecifics.
ADDITIONAL KEYWORDS: coevolution – Drosophila biauraria – Drosophila subauraria – Drosophila
triauraria – species-specificity.
INTRODUCTION The genitalia of male animals are generally among
the most rapidly evolving body structures, and their
‘… in many insects the male and female organs morphology can vary substantially even among closely
of copulation show closed adjustments to each related species (Eberhard, 1985; Hosken & Stockley,
other; those of the male having parts that fit 2004). Recent studies have suggested that female
precisely corresponding parts of the females. genitalia may also evolve rapidly – possibly more rapidly
These fittings vary from species to species, and a than male genitalia for certain species (Ah-King et al.,
change in the male finds a corresponding change 2014; Simmons & Fitzpatrick, 2019; Solan & Simmons,
in the female of the same species. I shall call 2019). According to the classic lock-and-key hypothesis
these lock and key adaptations―structures and (Dufour, 1844; Shapiro & Porter, 1989; Sota & Kubota,
functions complete at birth of organism.’ – T. 1998; Masly, 2012), male and female genitalia coevolve
H. Morgan (1910) to promote the success of conspecific mating and
prevent heterospecific mating, but this hypothesis has
rarely been supported empirically (Eberhard, 1985;
Hosken & Stockley, 2004). Instead, genital coevolution
*Corresponding author. E-mail: sawamura@biol.tsukuba.ac.jp may be a consequence of sexual conflict over mating, in
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207 195
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://
creativecommons.org/licenses/by/4.0/), which permits unrestricted reuse, distribution, and reproduction in any medium, provided
the original work is properly cited.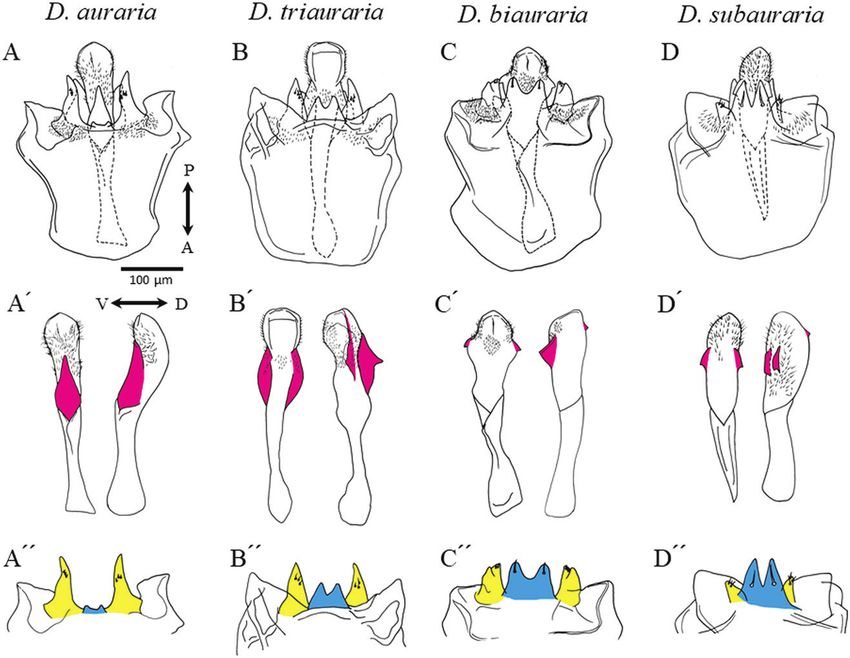
196 M. ONUMA ET AL.
which males voraciously pursue mating while females ablate part of the male genitalia using scissors or a laser
avoid mating, resulting in antagonistic coevolution (Polak & Rashed, 2010; Grieshop & Polak, 2012, 2014;
of genitalia (Arnqvist & Rowe, 1995, 2002; Rönn LeVasseur-Viens et al., 2015; Rhebergen et al., 2016).
et al., 2007; Kokko & Jennions, 2014). For species In the present work, we studied the four Drosophila
that are potentially polygamous, post-copulatory auraria complex species distributed in the Oriental
sexual selection can also promote genital coevolution region. They are under subgenus Sophophora, group
between the sexes. For example, females may select montium and subgroup montium (Sturtevant, 1939;
sperm from high-quality males for fertilization Hsu, 1949; Bock & Wheeler, 1972; Da Lage et al.,
during or after mating, and female behaviour and 2007; Yassin, 2018). Drosophila auraria as originally
genital morphology for this process might promote described by Peng (1937) was separated to ‘race A’, ‘race
the evolution of corresponding male traits – that is, B’ and ‘race C’ because they are sexually isolated from
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
cryptic female choice (Thornhill, 1983; Eberhard, each other (Moriwaki et al., 1952), and later described as
1985, 1996; Eberhard & Ramirez, 2004; Peretti & D. auraria, D. biauraria and D. triauraria, respectively
Aisenberg, 2015). Another example of post-copulatory (Bock & Wheeler, 1972). Drosophila subauraria was
sexual selection, namely sperm competition, is seen in described by Kimura (1983). Bock & Wheeler (1972) also
damselflies in which males use their genitalia to rake described D. quadraria, but this has been synonymized
sperm of previous males from the reproductive tract with D. triauraria because the morphology of the male
of females (Waage, 1979; Brennan & Prum, 2014). genitalia is essentially indistinguishable from that of
Data supporting sexual conflict and sexual selection D. triauraria (Watada et al., 2011).
are accumulating, and the aforementioned cases The phylogeny of the four species has not been well
constitute the major hypotheses for explaining rapid resolved (Gan et al., 2017, and references therein;
genital evolution (Eberhard, 1996; Hosken & Stockley, Chen et al., 2019; Conner et al., 2021) owing to their
2004; Arnqvist & Rowe, 2005; Brennan & Prum, 2014; close inter-relationships; it has been established
Kamimura, 2015, 2016). However, disentangling these that speciation took place less than 5 Mya (Yassin
causes of genital evolution is often difficult empirically, et al., 2016). Incomplete lineage sorting of ancient
as these forces are probably not mutually exclusive polymorphisms and/or interspecific introgression via
and the genital evolution could have a composite basis hybridization may have affected the gene tree. In fact,
with respect to mechanisms of divergence. a mitochondrial DNA-based phylogeny suggested that
The diversity of male genitalia has been well natural gene flow occurred between D. auraria and
established for Drosophila (e.g. Okada, 1954; Bock D. triauraria (Miyake & Watada, 2007). Interspecific
& Wheeler, 1972), but detailed analyses of female crossing within the D. auraria complex – especially
genitalia, including the relationship between genital between D. auraria and D. triauraria – is an easy
structures of both sexes, started only in recent decades. laboratory procedure that sometimes produces
For example, mating pairs of Drosophila were initially fertile hybrids (Kimura, 1987; Kim et al., 1989).
observed by scanning electron microscopy, and it The geographical distribution of the four D. auraria
seemed that female genitalia are less morphologically complex species overlaps, although each species has
diverse than male genitalia (Robertson, 1988; a distinct habitat preference (Kimura, 1987, and
Eberhard & Ramirez, 2004; Jagadeeshen & Singh, references therein). Sexual isolation among the four
2006). A method to clear cuticular structures was later species has been studied (Kurokawa, 1959, 1960,
introduced to assist the observation of genital coupling, 1963; Oguma et al., 1987), and it is known that the
and it is now possible to observe the positioning of inter-pulse interval of the male courtship song also
each part of the male and female genitalia during differs among the species and is recognized only by
copulation (Kamimura, 2010, 2016; Kamimura & conspecific females (Tomaru & Oguma, 1994a, b;
Mitsumoto, 2011, 2012a; Yassin & Orgogozo, 2013; Tomaru et al., 1998).
Muto et al., 2018). It is now generally accepted that The D. auraria species complex is a suitable model
female genitalia of Drosophila are also diverse, and for studies of genital coevolution because it is easy to
determining the function of each part of the male observe interspecific copulations and obtain artificial
and female genitalia is an ongoing pursuit. During introgressions from sibling species (M.O., unpublished
copulation, spines of Drosophila (especially, subgenus observations). Furthermore, this species complex may
Sophophora) male genitalia produce ‘copulatory wounds’ undergo natural, interspecific introgressions (Miyake
in the female genitalia and surrounding anatomy, and & Watada, 2007), and this may have led to intraspecific
these wounds are recognizable as melanized patches variation in the male genitalia (Kurokawa, 1967; the
(Kamimura, 2007, 2010, 2012, 2016; Kamimura & present report). To estimate causes of the interspecific
Mitsumoto, 2011, 2012a, b). Furthermore, to understand differences of genital morphology, which may underlie
the complementarity of the male and female genital the reproductive isolation, we describe and analyse
structures, surgical treatments have been conducted to coupling of the male and female genitalia during
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207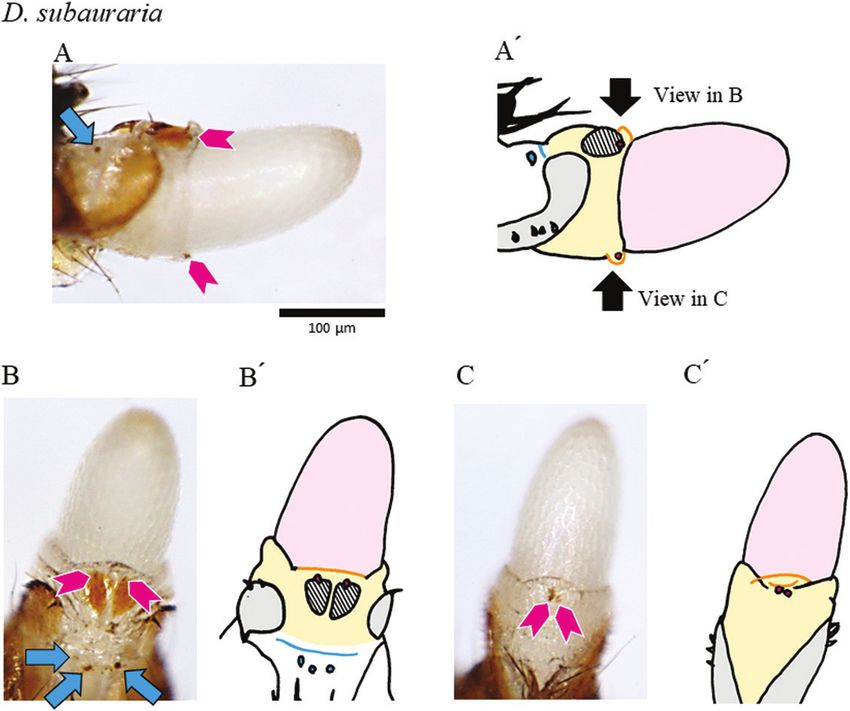
GENITAL COUPLING AND COPULATORY WOUNDING IN DROSOPHILA 197
copulation for the four D. auraria complex species. The Male dissection
significance of copulatory wounding found during the Wild-caught males and males from newly established
course of this study is also discussed. iso-female lines other than the aforementioned
standards were also dissected to assess morphological
variations. We collected flies in Ibaraki, Saitama,
MATERIALS AND METHODS Nagano, Shizuoka and Shiga Prefectures, Japan,
in 2014–2019 and established six D. auraria and 14
Flies D. triauraria iso-female lines. Males from five iso-
Each iso-female line was derived from a wild-caught female lines of D. triauraria from the Nansei (Ryukyu)
female in Japan, and the lines were used as standards Islands, Japan, were also dissected.
for the experiments: D. auraria YSM016 (Yoshimi, Male terminalia were dissected with a pair of
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
Saitama Pref.; established by Q. Araye in 2009), forceps in phosphate-buffered saline under an
D. triauraria T544 (Tsukuba, Ibaraki Pref.; established S8APO stereomicroscope (Leica Microsystems,
in 1980; EHIME-Fly stock number E-15301), Japan), removed from the carcass, and boiled in a
D. biauraria SG028 (Ueda, Nagano Pref.; established microtube containing 1 m potassium hydroxide at
by M.O. in 2019) and D. subauraria ONM29 (Ohnuma, 100 °C for 10 min. Phallic organs were cleaned and
Hokkaido; established in 1981; E-14901). Flies were their orientation adjusted in a drop of glycerol on a
reared at 25 ± 1 °C under a light/dark cycle of 14:10 h glass slide. They were observed under a CX23 light
with standard cornmeal medium for D. auraria and microscope (Olympus, Japan), and images were
D. triauraria or banana medium for D. biauraria and acquired using a single-lens reflex camera (EOS60D;
D. subauraria. Canon, Japan) connected to its dedicated adaptor
Figure 1. Phallic organs of the four species of the D. auraria complex. A, D. auraria. B, D. triauraria. C, D. biauraria.
D, D. subauraria. A–D, ventral view. P, posterior; A, anterior. A′–D′, aedeagus (left, ventral view; right, lateral view; V,
ventral; D, dorsal; appendages are indicated in magenta); A″–D″, a pair of pregonites (yellow) and protrusion of the median
gonocoxite (cyan).
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207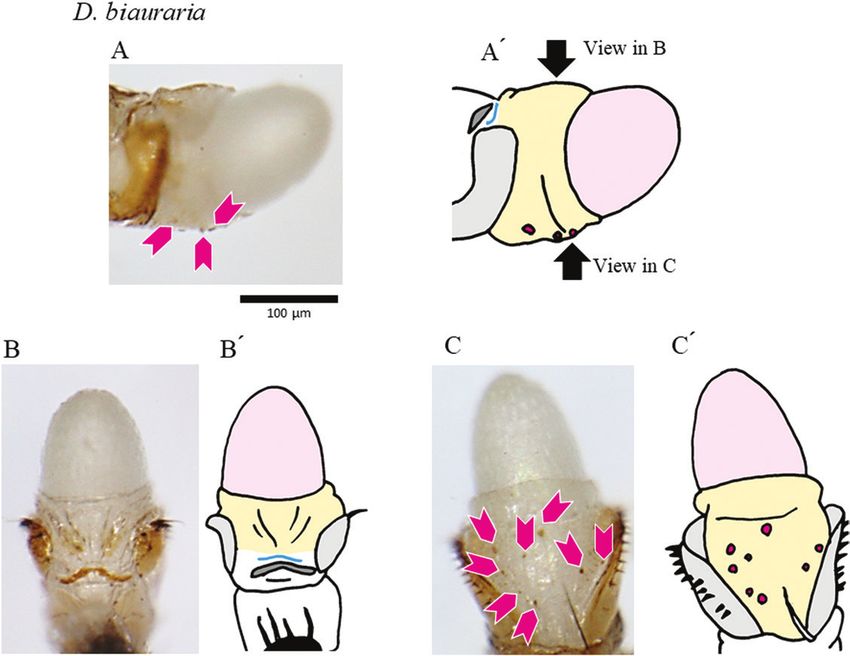
198 M. ONUMA ET AL.
(NY-1S; Canon). The terminology used for the phallic anaesthesia and kept separate based on sex for
organ structures follows that of Rice et al. (2019) 4–6 days, during which mating frequency is highest
applied to D. melanogaster; the homology of each for the D. auraria species complex (Oguma et al.,
structure examined in the present study is reliably 1987). For each species, 30 virgin females were
assigned in the D. auraria species complex. Briefly, the observed. After equal numbers of virgin females and
aedeagus is a tubular, sperm-transferring organ. The males were mixed in vials, they were left for 7 days.
hypandrium is the male ninth abdominal sternum that They were anaesthetized with diethyl ether and, for
extends beneath the aedeagus. The posterior part of each species, 30 females that had presumably mated
the hypandrium, termed gonocoxite, is characterized were observed to assess the presence of melanized
by its medial projection (median gonocoxite) which copulatory wounds.
bears hypandrial bristles. Pregonites are a pair of
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
lobes arising from and attached to the hypandrium,
anterior to the aedeagus, and are synonymous with Duration of copulation
hypandrial process and (anterior) paramere. A virgin female and male paired at 4–6 days after
emergence were put into a sample cup (Technicon
1.5 ml; Kartell, Italy) and covered with a piece of
Female dissection tissue paper (as an observation container), and
Female terminalia were dissected with a pair of forceps mating behaviour was observed. All observations were
in phosphate-buffered saline under a stereomicroscope conducted at 22–26 °C. Copulation duration (from
and then observed under a light microscope. Flies copulation initiation to separation) was measured for
were collected within 8 h after emergence without 20 pairs for each species.
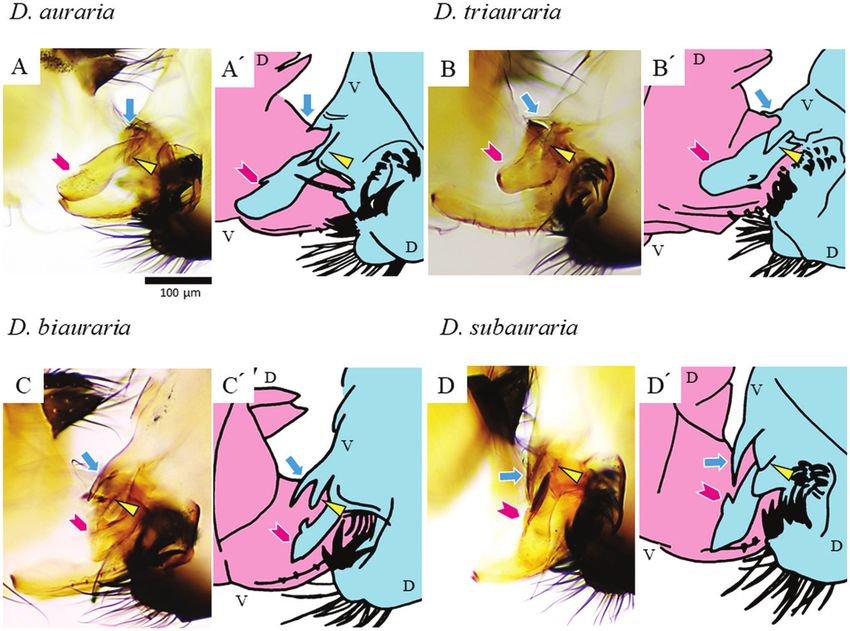
Figure 2. Positions of genital coupling of the four species of the D. auraria complex 4 min after initiation of coupling.
A, D. auraria. B, D. triauraria. C, D. biauraria. D, D. subauraria. The configuration of coupled genitalia is indicated in
the schematic drawing to the right of each photo (A′–D′): pink, female; light blue, male; V, ventral; D, dorsal. Arrowheads
(magenta), triangles (yellow) and arrows (cyan) indicate the aedeagus, pregonite and protrusion of the median gonocoxite,
respectively.
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207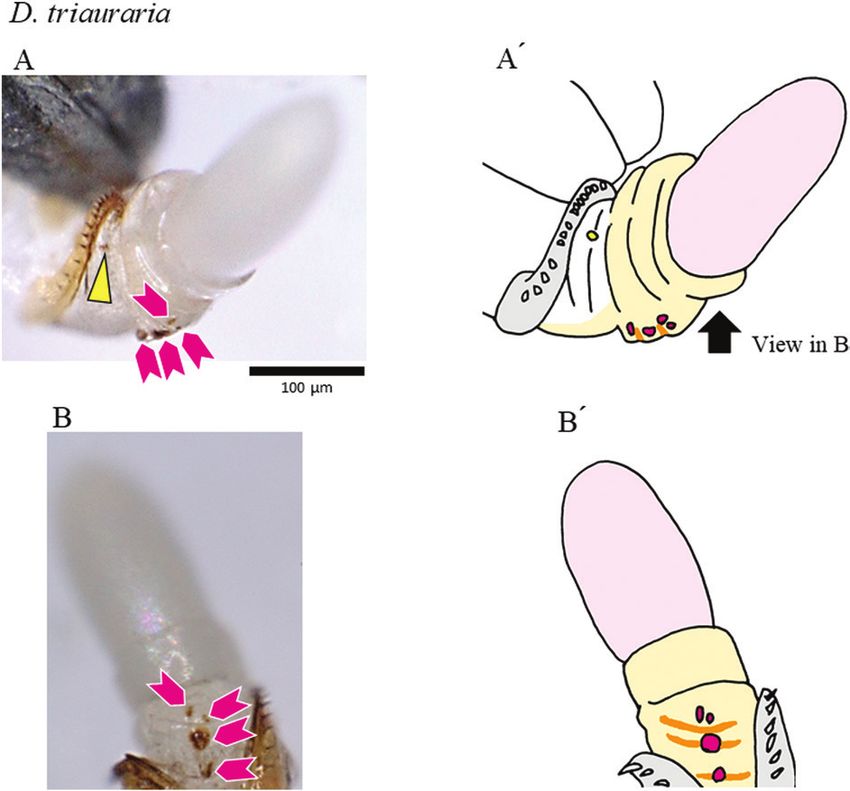
GENITAL COUPLING AND COPULATORY WOUNDING IN DROSOPHILA 199
Fixation of copula confirmed the results of previous studies (Okada, 1954;
Our method followed that of Kamimura & Mitsumoto Bock & Wheeler, 1972; Kimura 1983). The aedeagus
(2012a). Previous studies of Drosophila species have was found to possess associated structures (shown in
shown that stable coupling of male and female genitalia magenta in Fig. 1A′–D′) that were species-specific: a
and initiation of sperm transfer usually occur by the large, ventral projection in D. auraria, a dorsolateral
midpoint of the copulation timeframe (Jagadeeshan & expansion with a pair of claws in D. triauraria, and
Singh, 2006; Kamimura & Mitsumoto, 2011; Tanaka a pair of ventrolateral claws and a dorsal process in
et al., 2018). Copulation of the D. auraria species D. biauraria and D. subauraria. A pair of pregonites
complex lasts ~7−9 min (see Results), and therefore (shown in yellow in Fig. 1A″–D″) had several sensilla
we fixed copulating pairs at 1, 2, 3 or 4 min after for each of the four species and had bifid tips in
copulation initiation. For this, copulating pairs were D. biauraria, which were smaller in D. subauraria. The
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
flash-frozen in liquid nitrogen, fixed in 99.5% ethanol protrusion of the median gonocoxite of the hypandrium
at −18 °C for at least 14 days, and embedded in a 1% (shown in cyan in Fig. 1A″–D″) was small or absent in
agarose block. Each sample was dehydrated again in D. auraria but was larger and had a pair of spines in
99.5% ethanol and made transparent via absorption D. biauraria and D. subauraria.
with BABB solution (benzyl alcohol/benzyl benzoate, Intraspecific variations were also seen (Supporting
1:2) for 5 days. Genital coupling was observed under Information, Fig. S1 and Table S1). For D. auraria,
a light microscope (three mating pairs per time point). there was diversity with respect to the size and shape
of the protrusion of the median gonocoxite. Extra
spines were frequently seen on the pregonite or the
RESULTS protrusion of the median gonocoxite in individuals of
D. auraria and D. triauraria; those were similar to
Phallic organs variations previously described for local populations
As detailed below, our results concerning the where two or more species coexisted (Kurokawa,
interspecific differences in male genitalia (Fig. 1) 1967). As has been noted previously (M. T. Kimura and
Figure 3. Genital structures and copulatory wounds of a D. auraria female. The distal part of the female’s reproductive
tract was extruded by the pressure of an egg. A, lateral view. B, dorsal view (schematic drawings in A′ and B′). Black, analia;
light grey, oviscapt valves; dark grey, sclerotized plate between the genitalia and analia; pale orange, reproductive tract;
pink, egg. Magenta arrowheads denote spots that reflect wounds inside the reproductive tract, and the yellow triangle
denotes spots that reflect wounds in the depressions near the genital orifice.
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207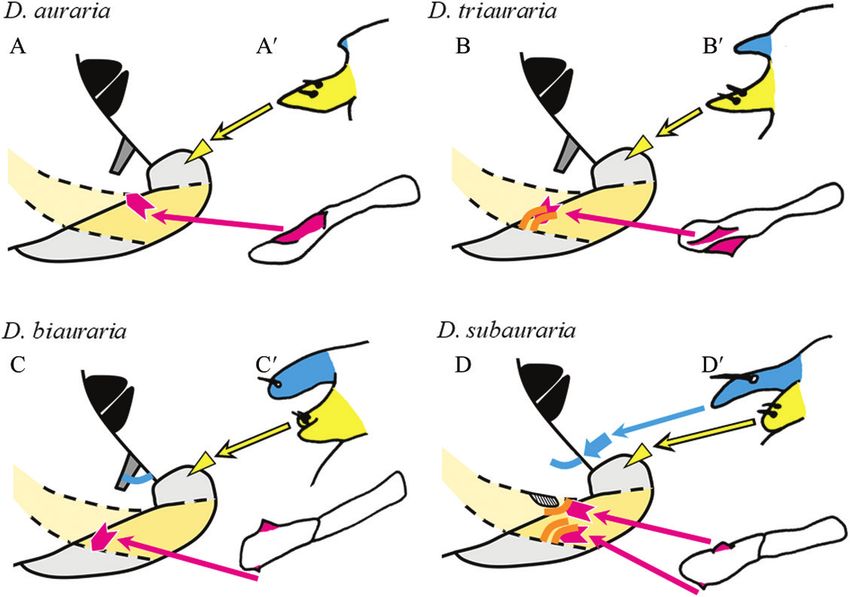
200 M. ONUMA ET AL.
M. Watada, personal communications), the aedeagus Female genitalia and copulatory wounds
morphology of D. triauraria varies with geographical Because the structures of the female terminalia have not
location – the strains from Nansei (Ryukyu) Islands been described in detail for the four Drosophila species
differ from the others. of the current study, we briefly describe them here
(Supporting Information, Figs S4, S5). A sclerotized plate
was present on the membrane between the genitalia
Genital coupling and analia in D. auraria, D. triauraria and D. biauraria
The duration of copulation (mean ± SD, N = 20) was (shown in dark grey in Figs 3–5), but not in D. subauraria
558 ± 149 s for D. auraria, 430 ± 216 s for D. triauraria, (Fig. 6). We noted a furrow on the membrane caudal
410 ± 154 s for D. biauraria and 455 ± 156 s for to the plate in D. biauraria and at a similar position
D. subauraria (Supporting Information, Fig. S2). These in D. subauraria (cyan lines in Figs 5, 6). A species-
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
results indicated that pairs should be fixed 1–4 min after specific fold structure(s) was seen in the vaginal wall,
copulation was initiated. For each of the four species, specifically on the ventral side in D. triauraria and on
the coupling of genitalia was established by 4 min after the dorsal and ventral sides in D. subauraria (orange
the initiation of copulation (Fig. S3). By 4 min, the lines in Figs 4, 6). For D. subauraria, there was also a
aedeagus was inserted into the female vagina (magenta nicked, round, sclerotized structure on the dorsal side of
arrowheads in Fig. 2), and the pair of pregonites touched the vaginal wall, close to one of the folds (hatched in Fig.
the female near her oviscapt valves (yellow triangles 6). As has been noted for other Drosophila species, a pair
in Fig. 2). The position of the protrusion of the median of depressions was present between the genital orifice
gonocoxite against the female differed somewhat and each oviscapt valve (Supporting Information, Fig.
among species: it rarely touched in D. auraria, barely S4C–E). The depressions are presumably functionally
touched alongside the female body in D. triauraria, and analogous to the blind invaginations (pockets) reported
deeply penetrated the female body in D. biauraria and for several other Drosophila species (Kamimura, 2007;
D. subauraria (cyan arrows in Fig. 2). Kamimura & Mitsumoto, 2012a).
Figure 4. Genital structures and copulatory wounds of a D. triauraria female. The distal part of the female’s reproductive
tract was extruded by the pressure of an egg. A, lateral view. B, ventral view (schematic drawings in A′ and B′). Light grey,
oviscapt valves; pale orange, reproductive tract; pink, egg; orange lines, the fold structure. Magenta arrowheads denote
spots that reflect wounds inside the reproductive tract, and the yellow triangle denotes spots that reflect wounds in the
depressions near the genital orifice. The sclerotized plate between the genitalia and analia is out of focus in these photos.
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207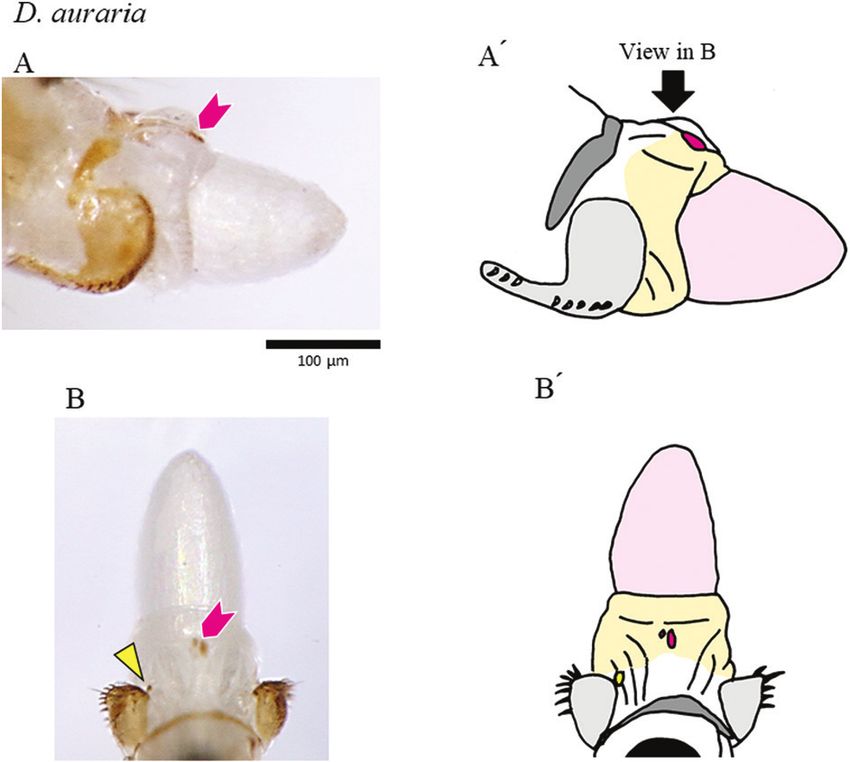
GENITAL COUPLING AND COPULATORY WOUNDING IN DROSOPHILA 201
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
Figure 5. Genital structures and copulatory wounds of a D. biauraria female. The distal part of the female’s reproductive
tract was extruded by the pressure of an egg. A, lateral view. B, dorsal view. C, ventral view (schematic drawings in A′–C′).
Light grey, oviscapt valves; dark grey, sclerotized plate between the genitalia and analia; pale orange, reproductive tract;
pink, egg; cyan, the furrow. Magenta arrowheads denote spots that reflect wounds inside the reproductive tract. Wounds in
the depressions near the genital orifice are out of focus in these photos.
Copulatory wounds, i.e. pigmented patches found on females were found in unique, species-specific
only in mated females, were observed in ~70% of locations, and Figure 7 presents possible scenarios to
mated females in each of the four species, and certain explain the origin of those unique wounds. First, the
species-specific wounds were observed at some copulatory wounds on the depressions near the genital
positions (Figs 3–6; Supporting Information, Fig. S6 orifice appear to have been caused by the male’s
and Table S2). Wounds on the depressions near the pregonites, which touched the female at that position.
genital orifice were common (Fig. S4D), whereas those Although copulatory wounds have been documented
on the membrane between the genitalia and analia in similar positions in other Drosophila species, those
(around the furrow) were specific to D. subauraria wounds were attributed to having been caused by
(Fig. 6). Wounds were also seen in the vaginal wall: different structures of the male genitalia (Kamimura,
on the dorsal side in D. auraria (Fig. 3), on the ventral 2007; Kamimura & Mitsumoto, 2012a). Second, the
side in D. triauraria (around the fold structure; Fig. 4), copulatory wounds on the vaginal wall appear to
on the ventral side in D. biauraria (Fig. 5), and on the have been caused by appendages of the aedeagus: (1)
dorsal and ventral sides (around the fold structures) in wounds on the dorsal side of the vagina by the ventral
D. subauraria (Fig. 6). projection of the male D. auraria, (2) wounds on the
ventral side by the dorsolateral claws of the male
D. triauraria, (3) wounds on the ventral side by the
DISCUSSION dorsal process of the male D. biauraria, and (4) wounds
on the dorsal side by the pair of ventrolateral claws and
Coevolution of genital structures between wounds on the ventral side by the dorsal process of the
sexes male D. subauraria. Third, the D. subauraria-specific
Our results highlight the diversity of genital structures wounds on the membrane between the genitalia and
– in both females and males – among the four species analia seemed to have been caused by the median
of the D. auraria complex. Certain copulatory wounds gonocoxite of the hypandrium, which penetrates the
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207202 M. ONUMA ET AL.
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
Figure 6. Genital structures and copulatory wounds of a D. subauraria female. The distal part of the female’s reproductive
tract was extruded by the pressure of an egg. A, lateral view. B, dorsal view. C, ventral view (schematic drawings in A′–C′).
Light grey, oviscapt valves; pale orange, reproductive tract; pink, egg; cyan, the furrow; hatched, the sclerite structure on
the reproductive tract; orange, the fold structures on the reproductive tract. Magenta arrowheads denote spots that reflect
wounds inside the reproductive tract, and cyan arrows denote spots that reflect wounds on the membrane between the
genitalia and analia. Wounds in the depressions near the genital orifice are out of focus in these photos.
female at that position. The functions of copulatory aedeagal appendages. Those are the fold structure(s)
wounding are much debated and may differ among on the ventral side in D. triauraria and on the dorsal
taxa (Lange et al., 2013; Reinhardt et al., 2014; Tatarnic and ventral sides in D. subauraria. The sclerotized
et al., 2014). Because the male organs responsible for structure on the dorsal side in D. subauraria may have
the wounds are located posterior to the phallotrema evolved to protect females from potential harm by the
in the female vagina, the wounds are unlikely to pair of aedeagal ventrolateral claws. This scenario is
function as entrances for sperm and/or seminal fluids similar to what has been reported for D. erecta and
to the female haemocoel (traumatic insemination or D. orena, which have that type of sclerotized structure
traumatic secretion transfer; sensu Lange et al., 2013). close to the position where the phallic spines or
Instead, copulatory wounding of these species may hooks touch the female (Yassin & Orgogozo, 2013;
retard remating of the females or may represent a side Kamimura, 2016). The furrow on the membrane
effect of mate-anchoring or stimulatory functions of between the genitalia and analia in D. biauraria and
male genitalia during mating. D. subauraria also may have evolved to accept the
Our results also allow speculation as to how genital gonocoxite.
morphology has coevolved between the sexes, i.e. based In summary, genital coupling and the positions
on species-specific postures of genital coupling and the of copulatory wounds differ among species of the
positions of copulatory wounds. Because the unique D. auraria complex. We speculate that structures on
positions of certain copulatory wounds coincided the vaginal wall and on the membrane between the
with species-specific structures on the vaginal wall, genitalia and analia have evolved to accept the species-
female structures may have evolved to accept the specific male structures. This may reduce the harm
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207GENITAL COUPLING AND COPULATORY WOUNDING IN DROSOPHILA 203
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
Figure 7. Possible causes of copulatory wounds. A, D. auraria. B, D. triauraria. C, D. biauraria. D, D. subauraria. Left,
lateral view of the female terminalia: black, the analia; dark grey, sclerotized plate between the genitalia and analia;
light grey, oviscapt valves; pale orange, vaginal wall; orange lines, the fold structure; cyan, the furrow. The hatched region
in D represents the sclerite structure on the reproductive tract. Magenta arrowheads denote wounds inside the vaginal
wall, yellow triangles denote wounds in the pockets near the genital orifice, and the cyan arrow denotes wounds on the
membrane between the genitalia and analia. Right (A′–D′), lateral view of the principal structures of the male genitalia.
Cyan, protrusion of the median gonocoxite; yellow, pregonites; magenta, appendages of the aedeagus.
caused by male genital structures without affecting evolutionary scenarios for genital coevolution in the
male fitness (tolerance trait). Alternatively, this may D. auraria species complex.
represent a resistance trait for efficient mate holding
by males. Such differences in genital structures may
prevent interspecific mating and/or lead to failure of Intraspecific variations
sperm transfer and thus may have played a role in Intraspecific variations were also seen. For example,
mechanical isolation, as has been suggested for other the protrusion of the median gonocoxite in D. auraria
species (Sota & Kubota, 1998; Kamimura & Mitsumoto, is diverse in terms of both size and shape, possibly
2012a; Muto et al., 2018). For example, D. subauraria because the protrusion does not play an important
males and females rarely mate with other sibling species role in copulation of D. auraria. In fact, the protrusion
(Kim et al., 1989), possibly because the sclerotized does not touch the female body in D. auraria, thus
plate on the membrane between the genitalia and leading to a relaxed constraint. A similar case has
analia is absent only in D. subauraria females (thus, been reported for D. suzukii, where pregonites do not
D. subauraria males have adapted to such females). touch the female in copula owing to the female’s large
Moreover, a cross between D. auraria females and oviscapt valves, and intraspecific variations are seen
D. triauraria males is less common than the reciprocal (Muto et al., 2018).
cross (Kim et al., 1989), possibly because D. auraria Extra spines were frequently seen on pregonites or
females do not have the fold structure on the vaginal the protrusion of the median gonocoxite in D. auraria
wall that accepts the appendage of the D. triauraria and D. triauraria. There are two possible explanations
aedeagus. Future examination of mounting success/ for this polymorphism: one is de novo mutations in the
failure, genital mismatch and sperm transfer rates species, and the other is interspecific introgression
in copulation trials with males and females from from, say, D. biauraria. Similar variations have been
different species or geographical populations could be reported in local populations of these species where
helpful to discriminate the relative importance of the two or more species coexist, and a hypothesis of
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207204 M. ONUMA ET AL.
such interspecific introgression has been proposed Arnqvist G, Rowe L. 2005. Sexual conflict. Princeton:
(Kurokawa, 1967). It should be stressed that the Princeton University Press.
polymorphism has lasted more than 50 years in Bock IR, Wheeler MR. 1972. The Drosophila melanogaster
natural populations (the present study). To understand species group. The University of Texas Publications 7213: 1–102.
the longevity of this polymorphism, it is necessary to Brennan PLR, Prum RO. 2014. Mechanisms and evidence
measure the fitness of variants. To test the possibility of genital coevolution: the roles of natural selection, male
of interspecific introgression, genital morphology choice, and sexual conflict. In: Rice WR, Gavrilets S, eds. The
genetics and biology of sexual conflict. New York: Cold Spring
must be compared between the natural variants and
Harbor Laboratory Press, 385–405.
the artificially produced hybrid descendants. Indeed,
Chen A, Chen C, Katoh T, Katoh TK, Watada M, Toda MJ,
future genomic analyses of polymorphic populations
Ritchie MG, Wen Sy. 2019. Evolution and diversity of the
may clarify this issue. A similar morphological
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
courtship repertoire in the Drosophila montium species group
variation is known – hypandrial bristles are usually (Diptera: Drosophilidae). Journal of Evolutionary Biology 32:
present in species of the D. melanogaster subgroup 1124–1140.
but are absent in D. santomea. This is a consequence Conner WR, Delaney EK, Bronski MJ, Ginsberg PS,
of a single nucleotide substitution at the enhancer Wheeler TB, Richardson KM, Peckenpaugh B, Kim KJ,
site of the scute gene (Nagy et al., 2018), and it would Watada M, Hoffman AA, Eisen MB, Kopp A, Cooper BS,
be interesting to know whether a similar genetic Turelli M. 2021. A phylogeny for the Drosophila montium
polymorphism has occurred in the D. auraria species species group: a model clade for comparative analyses.
complex. Molecular Phylogenetics and Evolution 158: 107061.
Da Lage JL, Kergoat GJ, Maczkowiak F, Silvain JF,
Cariou ML, Lachaise D. 2007. A phylogeny of Drosophilidae
using the Amyrel gene: questioning the Drosophila
ACKNOWLEDGEMENTS
melanogaster species group boundaries. Journal of Zoological
We are grateful to EHIME-Fly, KYORIN-Fly, and Systematics and Evolutionary Research 45: 47–63.
Drs Yukie Sato, Quenta Araye, Masayoshi Watada, Dufour L. 1844. Anatomie générale des diptères. Annales des
Masahito T. Kimura and Masanori J. Toda for providing Science Naturelles Zoologie et Biologie Animale 1: 244–264.
flies and useful information. Dr Shinichi Miyamura, Eberhard WG. 1985. Sexual selection and animal genitalia.
Dr Fumiaki Maruo and Ms Izumi Yamazaki permitted Cambridge: Harvard University Press.
use of the scanning electron micrograph presented Eberhard WG. 1996. Female control: sexual selection by
in the Supporting Information. Comments from cryptic female choice. Princeton: Princeton University Press.
Eberhard WG, Ramirez N. 2004. Functional morphology of the
anonymous reviewers improved the manuscript.
male genitalia of four species of Drosophila: failure to confirm
This study was partly supported by Grants-in-Aid
both lock and key and male–female conflict predictions. Annals
for Science Research from the Japan Society for the
of the Entomological Society of America 97: 1007–1017.
Promotion of Science (18K06383 to K.S. and 19H03276
Gan L, Li G, Li W, Zeng Q, Yang Y. 2017. Increase data
to Y.K.). The authors have no conflicts of interests to characters to construct the molecular phylogeny of the
declare. Drosophila auraria species complex. Open Journal of
Genetics 7: 40–49.
Grieshop K, Polak M. 2012. The precopulatory function of
DATA AVAILABILITY male genital spines in Drosophila ananassae [Doleschall]
(Diptera: Drosophilidae) revealed by laser surgery. Evolution
The data underlying the study are available in the main 66: 2637–2645.
figures and tables and in the Supporting Information. Grieshop K, Polak M. 2014. Evaluating the post-copulatory
sexual selection hypothesis for genital evolution reveals
evidence for pleiotropic harm exerted by the male genital
spines of Drosophila ananassae. Journal of Evolutionary
REFERENCES
Biology 27: 2676–2686.
Ah-King M, Barron AB, Herberstein ME. 2014. Genital Hosken DJ, Stockley P. 2004. Sexual selection and genital
evolution: why are females still understudied? PLoS Biology evolution. Trends in Ecology and Evolution 19: 87–93.
12: e1001851. Hsu TC. 1949. The external genital apparatus of male
Arnqvist G, Rowe L. 1995. Sexual conflict and arms races Drosophilidae in relation to systematics. The University of
between the sexes: morphological adaptation for control of Texas Publications 4920: 80–142.
mating in a female insect. Proceedings of the Royal Society Jagadeeshan S, Singh RS. 2006. A time-sequence functional
London B 261: 123–127. analysis of mating behaviour and genital coupling in
Arnqvist G, Rowe L. 2002. Correlated evolution of male Drosophila: role of cryptic female choice and male sex-drive
and female morphologies in water striders. Evolution 56: in the evolution of male genitalia. Journal of Evolutionary
936–947. Biology 19: 1058–1070.
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207GENITAL COUPLING AND COPULATORY WOUNDING IN DROSOPHILA 205
Kamimura Y. 2007. Twin intromittent organs of Drosophila LeVasseur-Viens H, Polak M, Moehring AJ. 2015. No
for traumatic insemination. Biology Letters 3: 401–404. evidence for external genital morphology affecting cryptic
Kamimura Y. 2010. Copulation anatomy of Drosophila female choice and reproductive isolation in Drosophila.
melanogaster (Diptera: Drosophilidae): wound-making Evolution 69: 1797–1807.
organs and their possible roles. Zoomorphology 129: Masly JP. 2012. 170 years of ‘lock-and-key’: genital morphology
163–174. and reproductive isolation. International Journal of
Kamimura Y. 2012. Correlated evolutionary changes in Evolutionary Biology 2012: 247352.
Drosophila female genitalia reduce the possible infection Miyake H, Watada M. 2007. Molecular phylogeny of the
risk caused by male copulatory wounding. Behavioral Drosophila auraria species complex and allied species of
Ecology and Sociobiology 66: 1107–1114. Japan based on nuclear and mitochondrial DNA sequences.
Kamimura Y. 2015. What is indirect cryptic female choice? Genes and Genetic Systems 82: 77–88.
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
Theoretical considerations and an example from a promiscuous Morgan TH. 1910. The American Society of Naturalists:
earwig. In: Peretti AV, Aisenberg A, eds. Cryptic female choice chance or purpose in the origin and evolution of adaptation.
in arthropods – patterns, mechanisms and prospects. Cham: Science 31: 201–210.
Springer International Publishing, 255–283. Moriwaki D, Okada T, Kurokawa H. 1952. Two types of
Kamimura Y. 2016. Significance of constraints on genital Drosophila auraria. Drosophila Information Service 26:
coevolution: why do female Drosophila appear to cooperate 112.
with male by accepting harmful matings? Evolution 70: Muto L, Kamimura Y, Tanaka KM, Takahashi A. 2018.
1674–1683. An innovative ovipositor for niche exploitation impacts
Kamimura Y, Mitsumoto H. 2011. Comparative copulation genital coevolution between sexes in a fruit-damaging
anatomy of the Drosophila melanogaster species complex Drosophila. Proceedings of the Royal Society London B
(Diptera: Drosophilidae). Entomological Science 14: 399–410. 285: 20181635.
Kamimura Y, Mitsumoto H. 2012a. Lock-and-key structural Nagy O, Nuez I, Savisaar R, Peluffo AE, Yassin A, Lang M,
isolation between sibling Drosophila species. Entomological Stern DL, Matute DR, David JR, Courtier-Orgogozo V.
Science 15: 197–201. 2018. Correlated evolution of two copulatory organs via a
Kamimura Y, Mitsumoto H. 2012b. Genital coupling and single cis-regulatory nucleotide change. Current Biology 28:
copulatory wounding in Drosophila teissieri (Diptera: 3450–3457.
D r o s o p h i l i d a e ) . C a n a d i a n Jo u r n a l o f Z o o l o g y 1 4 : Oguma Y, Kurokawa H, Akai SM, Tamaki H, Kajita J.
1437–1440. 1987. Interspecific differences in some courtship behavioral
Kim BK, Watanabe TK, Kitagawa O. 1989. Evolutionary properties among the four species belonging to the Drosophila
genetics of the Drosophila montium subgroup. I. Reproductive auraria complex. Zoological Science 4: 889–896.
isolations and the phylogeny. The Japanese Journal of Okada T. 1954. Comparative morphology of the Drosophilid
Genetics 64: 177–190. flies. I. Kontyu 22: 36–49.
Kimura MT. 1983. A new species of the Drosophila auraria Peng FT. 1937. On some species of Drosophila from China.
complex, D. subauraria, from northern Japan. Kontyu 51: Annotations Zoologicae Japonenses 16: 20–27.
593–595. Peretti AV, Aisenberg A. 2015. Cryptic female choice in
Kimura MT. 1987. Habitat differentiation and speciation arthropods – patterns, mechanisms and prospects. Cham:
in the Drosophila auraria species-complex (Diptera, Springer International Publishing.
Drosophilidae). Kontyu 55: 429–436. Polak M, Rashed A. 2010. Microscale laser surgery reveals
Kokko H, Jennions MD. 2014. The relationship between adaptive function of male intromittent genitalia. Proceedings
sexual selection and sexual conflict. In: Rice WR, Gavrilets S, of the Royal Society London B 277: 1371–1376.
eds. The genetics and biology of sexual conflict. New York: Reinhardt K, Anthes N, Lange R. 2014. Copulatory wounding
Cold Spring Harbor Laboratory Press, 23–35. and traumatic insemination. In: Rice WR, Gavrilets S, eds.
Kurokawa H. 1959. Experiments on sexual isolation between The genetics and biology of sexual conflict. New York: Cold
races A and B of Drorophila auraria. Annotationes Zoologicae Spring Harbor Laboratory Press, 115–139.
Japonenses 32: 220–224. Rhebergen FT, Coutrier-Orgogozo V, Dumont J,
Kurokawa H. 1960. Sexual isolation among the three races, Schilthuizen M, Lang M. 2016. Drosophila pachea
A, B, and C of Drosophila auraria. The Japanese Journal of asymmetric lobes are part of a grasping device and stabilize
Genetics 35: 161–166. one-sided mating. BMC Evolutionary Biology 16: 176.
Kurokawa, H. 1963. Population genetics on three races of Rice G, David JR, Kamimura Y, Masly JP. Mcgregor AP,
Drosophila auraria Peng. II. Sexual isolation among strains Nagy O, Noselli S, Nunes MDS, O’Grady P, Sánchez-
within a race C. The Japanese Journal of Genetics 38: 1–5. Herrero E, Siegal ML, Toda MJ, Rebeiz M, Courtier-
Kurokawa H. 1967. Variation in male genitalia of Drosophila Orgogozo V, Yassin A. 2019. A standardized nomenclature
auraria collected in nature. Annotationes Zoologicae and atlas of the male terminalia of Drosophila melanogaster.
Japonenses 40: 154–160. Fly 13: 51–64.
Lange R, Reinhardt K, Michaels NK, Anthes N. 2013. Robertson HM. 1988. Mating asymmetries and phylogeny in the
Functions, diversity, and evolution of traumatic mating. Drosophila melanogaster species complex. Pacific Science 42:
Biological Reviews 88: 585–601. 72–80.
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207206 M. ONUMA ET AL.
Rönn J, Katvala M, Arnqvist G. 2007. Coevolution between Tomaru M, Matsubayashi H, Oguma Y. 1998. Effects of
harmful male genitalia and female resistance in seed beetle. courtship song in interspecific crosses among the species of
Proceedings of the National Academy of Science of the United the Drosophila auraria complex (Diptera: Drosophilidae).
States of America 104: 10921–10925. Journal of Insect Behavior 11: 383–398.
S h a p i r o A M , Po r t e r A H . 1 9 8 9 . T h e l o c k - a n d - Tomaru M, Oguma Y. 1994a. Genetic basis and evolution
key hypothesis: evolutionary and biosystematics of species-specific courtship song in Drosophila auraria
interpretation of insect genitalia. Annual Review of complex. Genetical Research 63: 11–17.
Entomology 34: 231–245. Tomaru M, Oguma Y. 1994b. Differences in courtship song
Simmons LW, Fitzpatrik JL. 2019. Female genitalia can in the species of the Drosophila auraria complex. Animal
evolve more rapidly and divergently than male genitalia. Behaviour 47: 133–140.
Nature Communications 10: 1–8. Waage JK. 1979. Dual function of the damselfly penis: sperm
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
Sloan NS, Simmons LW. 2019. The evolution of female removal and transfer. Science 203: 916–918.
genitalia. Journal of Evolutionary Biology 32: 882–899. Watada M, Matsumoto M, Kondo M, Kimura MT. 2011.
Sota T, Kubota K. 1998. Genital lock-and-key as a selective Taxonomic study of the Drosophila auraria species complex
agent against hybridization. Evolution 52: 1507–1513. (Diptera: Drosophilidae) with description of a new species.
Sturtevant AH. 1939. On the subdivision of the genus Entomological Science 14: 392–398.
Drosophila. Proceedings of the National Academy of Science Yassin A. 2018. Phylogenetic biogeography and classification
of the United States of America 25: 137–141. of the Drosophila montium species group (Diptera:
Tanaka KM, Kamimura Y, Takahashi A. 2018. Mechanical Drosophilidae). Annales de la Société Entomologique de
incompatibility caused by modifications of multiple male France 54: 167–175.
genital structures using genomic introgression in Drosophila. Yassin A, Delaney EK, Reddiex AJ, Seher TD, Bastide H,
Evolution 72: 2406–2418. Appleton NC, Lack JB, David JR, Chenoweth SF,
Tatarnic NJ, Cassis G, Siva-Jothy MT. 2014. Traumatic Pool JE, Kopp A. 2016. The pdm3 locus is a hotspot for
insemination in terrestrial arthropods. Annual Review of recurrent evolution of female-limited color dimorphism in
Entomology 59: 245–261. Drosophila. Current Biology 26: 2412–2422.
Thornhill R. 1983. Cryptic female choice and its implications Yassin A, Orgogozo V. 2013. Coevolution between male and
in the Scorpionfly Harpobittacus nigriceps. The American female genitalia in the Drosophila melanogaster species
Naturalist 122: 765–788. subgroup. PLoS ONE 8: e57158.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the online version of this article at the publisher’s web-site:
Figure S1. Intraspecific variations of phallic organs. A–F, D. auraria. G–I, D. triauraria. A–D, representative
variations for protrusion of the median gonocoxite. A prominent protrusion (e.g. C and D) was seen in 3.4%
(16/471) of individuals. E and H, extra spines on the pregonite. F and I, extra spines on the protrusion of the
median gonocoxite. G, aedeagus of a male from Kume-jima Island (stock number E-15305). Left, ventral view;
right, lateral view; V, ventral; D, dorsal. Appendages are indicated in magenta. The aedeagus of D. triauraria
from Nansei (Ryukyu) Islands is round distally and has smaller, dorsolateral claws. Arrowheads indicate the
protrusion of the median gonocoxite or extra spines.
Figure S2. Copulation duration of the four species of the D. auraria complex. Error bars represent standard
deviation (N = 20).
Figure S3. Change of coupling posture over time. A, D. auraria. B, D. triauraria. C, D. biauraria. D, D. subauraria.
A–D, posture at 1 or 2 min (A′–D′, 3 or 4 min) after initiation of copulation. Note that the position of the protrusion
of the median gonocoxite (cyan arrow) differs between the two time points.
Figure S4. Anatomy of the female terminalia of D. auraria. A, lateral view. B, caudal view. C–E, the focus was on
the structures near the genital orifice (schematic drawings in A′–C′). Oviscapt valves were opened by pressing the
female’s abdomen. C, a virgin female. D, a mated female. E, scanning electron micrograph (courtesy of I. Yamazaki,
S. Miyamura and F. Maruo). Black, analia; dark grey, sclerotized plate between the genitalia and analia; light grey,
oviscapt valves; pale orange, vaginal wall. Yellow triangles denote wounds on the depressions near the genital
orifice.
Figure S5. Species-specific structures observed in virgin females of the four species of the D. auraria complex.
A, D. auraria. B, D. triauraria. C, D. biauraria. D, D. subauraria. The structures are indicated in the schematic
drawing to the right of each photo. A′–D′: light grey, oviscapt valves; orange, fold structures; cyan, furrow;
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207GENITAL COUPLING AND COPULATORY WOUNDING IN DROSOPHILA 207
dark grey, sclerotized plate between the genitalia and analia; hatched region in D′, sclerite structure on the
reproductive tract.
Figure S6. Copulatory wounds observed in females of the four species of the D. auraria complex. A, D. auraria. B,
D. triauraria. C, D. biauraria. D, D. subauraria. Species-specific structures and positions of wounds are indicated
in the schematic drawing to the right of each photo. A′–D′: light grey, oviscapt valves; orange, fold structures;
cyan, furrow; magenta, wounds inside the reproductive tract; yellow, wounds in the depressions near the genital
orifice; cyan, wounds on the membrane between the genitalia and analia; dark grey, sclerotized plate between the
genitalia and analia; hatched region in D′, sclerite structure on the reproductive tract.
Table S1. Intraspecific variation in the number of extra spines.
Table S2. Presence of copulatory wounds.
Downloaded from https://academic.oup.com/biolinnean/article/135/1/195/6381422 by guest on 04 February 2022
© 2021 The Linnean Society of London, Biological Journal of the Linnean Society, 2022, 135, 195–207You can also read

















































