Filippa Bertilsson - Using the eminent toolkit of Wolbachia to study Culex pipiens populations and their relations in Europe
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Using the eminent toolkit of Wolbachia
to study Culex pipiens populations and
their relations in Europe
Filippa Bertilsson
Degree project C in Biology 15hp, Bachelor of Science, 2022
Examensarbete C I biologi 15hp, Kandidatexamen 2022
The National Veterinary Institute, SVA, Uppsala, Sweden
2022-05-26
Supervisor: Tobias Lilja PhD
Abstract
Culex pipiens, in the family Culicidae, has emerged as one of the biggest vectors for West
Nile virus. It has two bioforms, pipiens and molestus, which differ from each other regarding
habitat, diapause, and prey. Pipiens prefers to bite birds, and molestus prefers to bite humans.
There is to some extent hybridization between the two, which creates a bridge-vector between
birds and humans. One way to study the relationships and spreading of the mosquitos is using
the intracellular bacteria Wolbachia pipientis which is present in at least 99% of al Culex
mosquitoes. The bacteria have two fast evolving genes, pk1 and ank2 which are suitable for
this. Not only are the bacteria suitable for genetics, but it is also manipulating the
reproductive system of the mosquitoes through Cytoplasmic Incompatibility, which changes
structures of populations and allows for the bacteria to spread fast and efficient. We wanted to
investigate levels of Wolbachia in different populations, as well as if the two bioforms prefer
a prey, together with mapping the relationships between populations using the two genes. We
found that Wolbachia was present in all tested mosquitoes, with higher levels of it in the
abdomen than in the thorax. We also found that the theory of a preferred prey was false within
the tested populations, since both bioforms preferred birds. Lastly, we could identify five
different strains of Wolbachia pipientis concentrated to different locations. This study has
shown that Wolbachia is present in all tested mosquitoes and is a useful tool to determine
relationships within and between populations. This is important to be able to gain
understanding of the spread of West Nile virus and other vector borne diseases spread by
Culex pipiens mosquitoes.
List of abbreviations
WNV: West Nile Virus
ANK: Ankyrin
pk1: pyruvate kinase 1
ank2: ankyrin 2
Wsp: Wolbachia surface protein gene
FtsZ: Filamenting temperature-sensitive mutant Z gene
wPip: Wolbachia pipientis
CI: Cytoplasmic incompatibility
IIT: Incompatible Insect Technique
2
Table of Contents
1. INTRODUCTION .......................................................................................................................................... 4
1.1 THE CULEX MOSQUITO ........................................................................................................................................ 4
1.1.1 Culex pipiens pipiens and Culex pipiens molestus ................................................................................. 4
1.1.2 Hybridization between pipiens and molestus ....................................................................................... 5
1.1.3 Cx. pipiens as vector .............................................................................................................................. 5
1.2 WOLBACHIA ..................................................................................................................................................... 5
1.2.1 Variants of Wolbachia .......................................................................................................................... 6
1.2.2 Cytoplasmic incompatibility and Wolbachia ......................................................................................... 6
1.2.3 Applications for Wolbachia ................................................................................................................... 7
1.3 THE EXPERIMENTS .............................................................................................................................................. 7
2. METHODS ................................................................................................................................................... 8
2.1 DISSECTION OF MOSQUITOES ............................................................................................................................... 8
2.2 EXTRACTION OF DNA ......................................................................................................................................... 8
2.3 REAL-TIME PCR – DETECTION OF WOLBACHIA ........................................................................................................ 8
2.4 PCR FOR SEQUENCING ........................................................................................................................................ 8
2.5 GEL ELECTROPHORESIS ........................................................................................................................................ 9
2.6 SEQUENCING PREPARATION ................................................................................................................................. 9
2.7 REAL-TIME PCR – DETERMINE SPECIES ................................................................................................................... 9
2.8 STATISTIC ANALYSIS .......................................................................................................................................... 10
2.9 TREE CONSTRUCTION ........................................................................................................................................ 10
3. RESULTS .................................................................................................................................................... 11
3.1 EUROPEAN MOSQUITOES ................................................................................................................................... 11
3.1.1 qPCR results ........................................................................................................................................ 11
3.1.2 PCR results .......................................................................................................................................... 12
3.2 RESULTS FOR GOTHENBURG MOSQUITOES ............................................................................................................ 12
3.2.1 qPCR results – species ......................................................................................................................... 12
3.2.2 qPCR results – Wolbachia ................................................................................................................... 13
3.2.3 PCR results .......................................................................................................................................... 13
3.3 PHYLOGENETIC RESULTS .................................................................................................................................... 14
4. DISCUSSION .............................................................................................................................................. 16
4.1 DIFFERENT LEVELS OF WOLBACHIA IN ABDOMEN AND THORAX .................................................................................. 16
4.2 NO PREFERRED PREY OF PIPIENS NOR MOLESTUS – HYBRIDIZATION IMPORTANT ............................................................ 16
4.3 FIVE DIFFERENT WPIP-STRAINS IDENTIFIED ............................................................................................................ 16
5. CONCLUSIONS .......................................................................................................................................... 17
6. ACKNOWLEDGEMENTS ............................................................................................................................. 17
7. REFERENCES.............................................................................................................................................. 18
7.1 LITTERATURE, ARTICLES AND WEBPAGES ............................................................................................................... 18
7.2 SOFTWARE ..................................................................................................................................................... 20
APPENDIX ..................................................................................................................................................... 21
3
1. Introduction
Insects have important roles in nature and its ecosystems, such as pollinators, decomposers,
and providers (National Geographic, 2020), but they are at the same time the largest group of
vectors spreading some of the worst diseases in the world, such as Malaria, Zika virus fever,
and Dengue (World Health Organization, 2020). On Earth, there are more than 3,000 species
of mosquitoes (National Geographic, 2010) and they are seen as the deadliest animal in the
world (Harvard University, 2014) due to their vector abilities. In the Culex genus, the Culex
pipiens complex mosquitoes are vectors for West Nile fever, together with Japanese
encephalitis and Lymphatic filariasis. (World Health organization, 2020). Culex pipiens
complex has over the last two decades emerged as one of the major vectors of diseases over
the northern hemisphere, mainly due to it during this period spreading the West Nile Virus
(WNV) (Haba & McBride, 2022). Despite it being an important vector, Culex pipiens
complex are hosts of the bacterial endosymbiont Wolbachia, which is known to affect the
mosquito’s biology (Sicard et al. 2019).
1.1 The Culex mosquito
The Culex genus is a part of the tribe Culicini and the family Culicidae. The family Culicidae
consists of 3,593 known species, and the subfamily Culicinae is divided into 110 genera
consisting of 11 tribes, where Culicini is one of them (Mosquito Taxonomic Inventory, 2008).
Culex mosquitoes are found all over the world, except the small exception of the most
northern areas of temperate zones (Mike Service, 2008) (see figure 1). In northern regions,
Culex pipiens (Cx. pipiens) is the most common one, while Culex quinquefasciatus (Cx.
quinquefasciatus), which is closely related to Cx. pipiens, is more common in subtropic and
tropic regions (Britannica, 2020).
Figure 1: An illustration over Culex pipiens and Culex quinquefasciatus’ distribution ().
1.1.1 Culex pipiens pipiens and Culex pipiens molestus
The Cx. pipiens mosquito, also called the northern house mosquito, is associated with
transmitting three out of ten arboviruses that exist in Europe, one of them being the WNV
(Brugman et al. 2018). There are two bioforms of Cx. pipiens: Cx. pipiens pipiens and Cx.
pipiens molestus. One main difference between these variants is the prey: while the pipiens
bioform choose to bite birds, the molestus bioform prefer to bite humans (Haba & McBride,
4
2022). The two biotypes also diverge between each other in their physiology and behavior,
such that pipiens need blood to be able to develop the first batch of eggs, while molestus do
not. There is also a significant difference between the two biotypes regarding where they mate
and live. Molestus prefers to live underground with colder temperatures, meaning they do not
need large areas to mate. On the contrast, pipiens lives above ground and need wide, open
areas to mate (Shaikevich et al. 2016). Another difference in the behavior of the two biotypes
is noticeable during the colder part of the year. It is shown that molestus do not require
diapaus during the winter, while pipiens do. This is due to them originating from the
Mediterranean area and Nort Africa and thereby not accustomed to winter (Haba & McBride,
2022). One reason for it to not be able to enter diapause seems to be its short photoperiod, a
result of the sewers et cetera where they live not experiencing natural variations in light
(Epstein et al. 2021). The only morphological difference between pipiens and molestus is the
structure of the males’ genitals. In females, there is no difference (Shaikevich et al. 2016).
1.1.2 Hybridization between pipiens and molestus
The two bioforms, pipiens and molestus, can mate with each other, and thereby create a
pipiens-x-molestus hybrid. Both biotypes have been found in both underground and
aboveground environments in Europe where they were not genetically isolated, being
evidence for hybridization (Amraoui et al. 2012). However, hybridization between the
bioforms is not common in the northern regions of Europe due to the two habitats of the
bioforms being more separated (Vogels et al. 2015). At the same time, there is a more
complicated situation in more southern regions of Europe. There, molestus still occupy
underground environments, but above ground, the mosquitoes behaved more like hybrids.
Around the Mediterranean region there are varying structure withing populations, and not just
between place and habitat. Here, the bioforms are found in the same traps, meaning the live in
the same areas above ground (Haba & McBride, 2022).
1.1.3 Cx. pipiens as vector
The mosquitoes get infected with a pathogen when they feed on birds carrying the disease. It
is then transmitted to humans via the salivary glands of the mosquito when it stings (World
Health Organization, 2017). Understanding Cx. pipiens and the hybridization between the two
forms is important. This is due to the phenomenon having influence on spreading of the
WNV. Since the hybrid feeds on both birds and humans, they can act as bridge-vectors
between the two parts (Gomes et al. 2013), but Cx. pipiens overall plays an important role in
virus transmission. WNV is for example endemic in Italy, where the transmission rates vary
between 37% and 47%. At the same time, it is 33% in the Netherlands, suggesting that WNV
can emerge further north in Europe. In recent years WNV has been found in Germany
(European Center for disease prevention and control, 2021).
1.2 Wolbachia
Up to the 1990s, Wolbachia were seen to be a part of another rare bacteria genus, but thanks
to new biological and molecular instruments there was found out that Wolbachia were
distributed among many insects and other arthropods, e.g., spiders and scorpions. Not only
were they found in arthropods, but they also exist in filarial nematodes (Werren et al. 2008)
and strictly in the subfamilies Onchocercinae and Dirofilariinae (Taylor et al. 2005).
Wolbachia pipientis (wPip) is the type species of the genus, the name given due to it first
being described in Cx. pipiens (Werren et al. 2008). The genus is a part of the
Anaplasmataceae family in the order Rickettsiales, being a-proteobacterial (Taylor et al.
2005). There is believed to be Wolbachia present in 66% of all insect species known (da Silva
et al. 2021).
5
1.2.1 Variants of Wolbachia
There are eight “supergroups”, A-H (see figure 2), in the genus Wolbachia which is based on
Wsp, FtsZ, and 16S gene sequences. The supergroups A and B are the ones that cause most
infections in insects, while C and D infect the filarial nematodes. The other groups are more
specialized. wPip is a part of group B (see figure 1). Wolbachia have been found in many
medically important species, such as the main vectors of malaria and dengue. For Cx. pipiens,
more than 99% of all individuals are infected (Hughes et al. 2012).
Figure 2: Phylogeny over Wolbachia (Gerth et al. 2014).
The two genes ank2 and pk1 are used due to them being fast evolving markers when
establishing Wolbachia strains, which both encode for ankyrin (ANK) motifs (Dumas et al.
2013). Wolbachia that infect arthropods tend to have a high number of genes that code for
different proteins containing the ANK domain, higher than those infecting nematodes,
meaning they might play a critical role when creating a relationship between the host and
bacteria (Duron et al. 2007).
1.2.2 Cytoplasmic incompatibility and Wolbachia
Cytoplasmic incompatibility (CI) is a form of reproductive manipulation. It takes place when
a male, infected with e.g., Wolbachia, mates with an uninfected female. Females that are
infected can mate with males that are either infected or uninfected, meaning they have an
advantage fitness-wise over uninfected females, since uninfected females only can mate with
uninfected males. It is thanks to the fitness advantage in infected females that Wolbachia can
spread quick through populations, such as Cx. pipiens. Yet there is not only one way that
Wolbachia can affect mosquitoes’, and other arthropods’, reproduction, but regardless of the
method, the female’s offsprings made by infected females increase in numbers, compared to
number of female offsprings made by uninfected females, which decrease. The offspring
inherit the bacteria maternally which allows it to spread quickly through populations (Hughes
& Rasgon, 2012).
6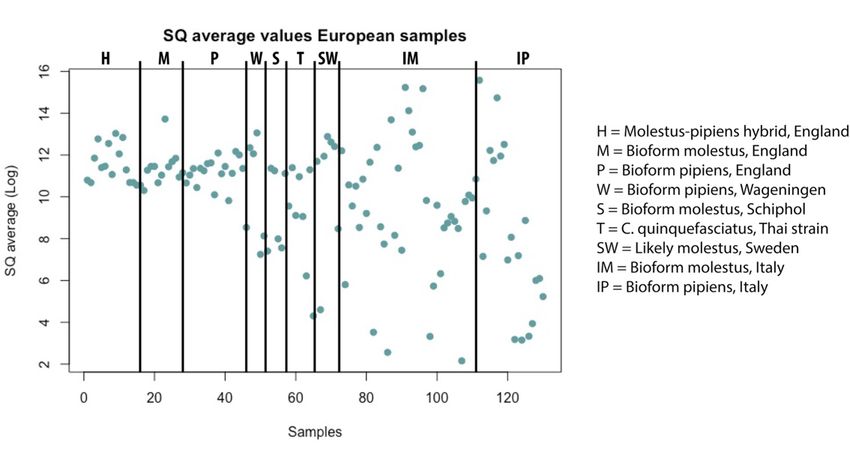
1.2.3 Applications for Wolbachia
The bacteria have two abilities in its host: pathogen protection or pathogen interference.
Pathogen protection leads to a fitness benefit for the insect through interference with the
development of the pathogen. For pathogen interference, the bacteria alter the insect’s ability
to get infected by vertebrate pathogens, but also its ability to spread it. The mechanisms
behind it are not understood, but one hypothesis suggests that an increased basal immunity in
the host is the reason behind pathogen interference. Another hypothesis suggests that
pathogen interference appears due to metabolic competition between Wolbachia and the
pathogen, meaning the bacteria steals nutrients from the pathogen, creating a non-pathogen-
friendly environment (Hughes & Rasgon, 2012).
Wolbachia can manipulate its host in more than one way, both interfering with the pathogen
and its reproductive systems, which could be used in our advantage. It could be used to
control arthropod-borne diseases, such as WNV. One way is using the incompatible insect
technique (IIT), which is a technique where bacteria-mediated CI suppresses populations of
insects. Infected males are introduced to a population which mate with un-infected females,
resulting in no progenies (Hughes & Rasgon, 2012). Although there is a problem with
reducing population size, since it is relying on a bite from a mosquito carrying the pathogen,
and there is yet little evidence on what method is effective. Another path to take is to modify
the vector, making it resistant to transmitting the disease. These methods would theoretically
work in populations such as the mosquito Aedes aegypti since it is not a natural host for the
bacteria, meaning it can be introduced to populations in a controlled way (Flores & O’Neill,
2018). As for Cx. pipiens, 99% of all individuals carry Wolbachia (Hughes & Rasgon, 2012),
meaning you would have to discover a strain of the bacteria that would not only compete with
the naturally occurring strain, but also reduce transmission efficiency (Ong, 2021).
1.3 The experiments
This project took place in two parts. The first part contained mosquitoes from different places:
England, Italy, and different strains grown in Alnarp, Sweden. The aim with these mosquitoes
was to see if there were higher levels of Wolbachia in the abdomen than in the thorax, and to
map the different strains of Wolbachia to see if they correlate with different populations of
mosquitoes. The second part contained mosquitoes collected in traps in Gothenburg. The traps
were chemically designed to smell like the mosquitos’ different preys: humans or birds. Here,
one aim was to see if the two biotypes of Cx. pipiens (molestus and pipiens) have a preferred
prey as it is hypothesised. The other aim was the same as with the European mosquitoes: to
map the Wolbachia strains (here, no comparison of Wolbachia levels between thorax and
abdomen was made). From here, the two parts of the project will be referred to as European
mosquitoes and Gothenburg mosquitoes.
Aims
1. Measure and compare levels of Wolbachia in abdomen and in thorax (European
mosquitoes)
2. Determine if the biotypes have a preferred prey, human or bird (Gothenburg
mosquitoes)
3. Map the different Wolbachia strains in Culex mosquitos (European and Gothenburg
mosquitoes)
7
2. Methods
2.1 Dissection of mosquitoes
The first step in the process of the European mosquitos was to separate the abdomen and
thorax. The dissection was carried out using two tweezers. Each mosquito came in a tube with
ethanol. The ethanol, together with the mosquito, was poured out on a plate, and the ethanol
discarded. It was then left to dry shortly before the dissection. A stereo microscope was used
to get good vision. The mosquito was then cut into two parts: abdomen, and thorax together
with head and legs. They were then stored in the freezer at -20 °C overnight.
2.2 Extraction of DNA
The following step was DNA extraction. The kit used was QIAGEN’s QIAamp DNA Mini
Kit (250). After the samples were taken out of the freezer, 180 µl ATL lysis buffer was added
to each tube, together with 20 µl Proteinase K. All samples were then mushed in the solutions
using a plastic pestle. This was carried out until the mosquito’s part was separated into
smaller bits. They were then incubated at 56°C for approximately 1 hour. After incubation, 20
µl AL buffer was added, and the tubes were then vortexed for approximately 10 seconds.
They were then incubated again at 70°C for 10 minutes, and then spun down quickly to
remove the condensation from the lids. After the second incubation, 20 µl 99% Ethanol was
added, and the tubes were shortly spun down. All samples were then transferred to new
column tubes, that came with the kit, and then got centrifuged at 8000 rpm for 1 minute. The
flowthrough was discarded. 500 µl AW1 buffer (washing buffer) was added, and tubes were
centrifuged at 8000 rpm for 1 minute. The flowthrough was discarded. Then 500 µl AW2
buffer (washing buffer) was added before they were centrifuged at 14000 rpm for 1 minute.
The flowthrough was discarded. Without additions, the tubes were then centrifuged further, at
14000 rpm for 2 minutes, to get all liquids out of the filter. The collection tubes were thrown
away and the columns were put in new Eppendorf tubes. 200 µl AE buffer, elution buffer,
was added and the tubes were incubated at room temperature for 1 minute before they were
centrifuged at 8000 rpm for 1 minute. The columns were thrown away and the Eppendorf
tubes with DNA was placed in the freezer at -20 °C.
2.3 Real-time PCR – detection of Wolbachia
To determine the level of bacteria in the mosquitos and see if there was a difference between
abdomen and thorax, a qPCR was performed. A master mix was prepared containing primers:
Wol_wsp_OSM_323(5’-TAGCGATTGAAGATATGC), and Wol_wsp_OSM_324(5’-
CTAGCTTCTGAAGGATTG) with the working stock 10 µM. The probe used was
Wol_wsp_probe_OSM_324(5’/56-FAM/CACCAACAC/ZEN/CAACACCAA) with working
stock 10 µM. Together with these, Perfecta Toughmix 2x (QuantaBio), and nuclease free
water were added. For 1 reaction: 10 µl Perfecta Toughmix 2X, 1.2 µl forward primer, 1.2 µl
reverse primer, 0.4 µl probe, 5.2 µl nuclease free water. One reaction without template was 18
µl, and 2 µl DNA was added to each well. A 96 well plate was used, containing samples and 3
controls with given concentrations (10-4, 10-5, 10-6). All samples including controls were run
as either duplicates or triplicates. The program was: 95°C 3 min, 95°C 5 sec, 55°C 20 sec,
72°C 30 sec, 45x.
2.4 PCR for sequencing
Only abdomen samples were used for sequencing. Only samples giving a higher SQ average
than 1000 were used in further experiments. The gene amplified for pk1 used the following
primers: Wol_pk1_Wpa_0256_f(5’-CCA-CTACATTGCGCTATAGA) and
8
Wol_pk1_Wpa_0256_r(5’-ACA-GTAGAACTACACTCCTCCA) in 10 µM working stock.
For 1 reaction: 2.5 µl Thermo Scientific PCR Buffer 10X, 1 µl Thermo Scientific MgCl2 25
mM, 0.5 µl dNTPs 10 mM, 1 µl forward primer, 1 µl reverse primer, 0.1 µl AmpliTaq GoldTM
DNA Polymerase with Buffer I (Thermo Fisher scientific), 17.9 µl nuclease free water.
Lastly, 1 µl of DNA was added, giving a final volume of 25 µl. The PCR program was 95°C
10 min, 95°C 15 sec, 52°C 30 sec, 72°C 1.5 min x35, 72°C 5 min, 4°C until further use.
The same procedure was done for the ank2 gene with the following primers:
Wol_ank2_Wpa_0652_f(CTTCTTCTGTGAGTGTACGT),
Wol_ank2_Wpa_0652_r(TCCATATCGATCTACTGCGT) (Duron et al. 2007).
2.5 Gel electrophoresis
To check the PCR results, a gel electrophoresis was performed. For the gel, a 1% agarose gel
was prepared with 1X TAE buffer, 1 g agarose, and 1.5 µl GelRed (Biotium), for 100 ml
buffer. For the chamber, 1X TAE buffer was used. The PCR products were prepared for
loading using 5 µl PCR product and 1.8 µl loading buffer (1x: 0.24 g bromophenol blue, 3 ml
glycerol). The ladder used was Thermo Scientific™ GeneRuler 100 bp DNA Ladder, Catalog
number: SM0241, and the gel was run at 120 V, 260 mA for about 40 minutes.
2.6 Sequencing preparation
A 96-well plate was used. 6 µl PCR product were used for each well, together with 1.5 µl
ExoSAP (0.5 µl Exonuclease, 1 µl Fast Alkaline Phosphatase). The plate was then incubated
in a PCR machine at 37°C for 15 minutes and 80°C for 15 minutes. Lastly, the primers for
either pk1 (Wol_pk1_Wpa_0256_f(5’-CCA-CTACATTGCGCTATAGA) and
Wol_pk1_Wpa_0256_r(5’-ACA-GTAGAACTACACTCCTCCA)), or ank2
(Wol_ank2_Wpa_0652_f(CTTCTTCTGTGAGTGTACGT),
Wol_ank2_Wpa_0652_r(TCCATATCGATCTACTGCGT), were added, 2.5 µl 10 µM in
each well. The sequencing method used was Sanger sequencing by Macrogen Europe.
2.7 Real-time PCR – determine species
For the Gothenburg mosquitoes, the first step was to determine which species it was since it
was unknown. After species determination, they went to the same procedures as the European
Mosquitoes (steps 2.2 to 2.6). Two master mixes were prepared, one to determine Cx. pipiens
pipiens from Cx. pipiens molestus (MM_1) using the CG11 loci, and one to determine Cx.
pipiens from Cx. torrentium (MM_2) as control, using the Ace2 loci. The primers were
ordered according to Vogels et al. 2015.
MM_1, 1 reaction: 8.5 µl nuclease free water, 12.5 µl QuantaBio PerfeCTa ToughMix 2X,
0.6 µl CQ11 forward primer: 5′-GCGGCCAAATATTGAGACTTTC 10µM, 0.6 µl CQ11
reverse primer: 5′-ACTCGTCCTCAAACATCCAGACATA 10 µM, 0.4 µl Probe
Cpp_pip_P1: ′-VIC-CACACAAAYCTTCACCGAA-MGB 10 µM, 0.2 Probe Cpp_pip_P2:
5′-VIC-ACACAAACCTTCATCGAA-MGB 10 µM, 0.2 Cpp_mol_P: 5′-FAM-
TGAACCCTCCAGTAAGGTA-MGB 10 µM and lastly 2 µl DNA template.
MM_2, 1 reaction: 9.1 µl nuclease free water, 12.5 µl QuantaBio PerfeCTa ToughMix, 0.6
Primer Cx_tor_F: 5′-CTTATTAGTATGACACAGGACGACAGAAA 10 µM, 0.6 Primer
Cx_tor_R: 5′-GCATAAACGCCTACGCAACTACTAA 10 µM, 0.2 µl Probe Cx_tor_P: 5′-
FAM-ATGATGCCTGTGCTACCA-MGB 10 µM and lastly 2 µl DNA template.
9
Each sample, together with 9 positive controls (3x Cx. pipiens pipiens, 3x Cx. pipiens
molestus and 3x Cx. torrentium) was run with each primer on the following program: 95°C 3
min, 95°C 13 sec, and 62°C 1 min, step 2 and 3 repeated 45x.
2.8 Statistic analysis
For the Gothenburg mosquitoes, a Fisher’s exact test was performed to see if each bioform
had a preferred prey. The test was done in RStudio with the following code:
fisher.test(data$Scent, data$Bioform), where Scent in the datafile was the trap, either Bird or
Human, and Bioform where either pipiens or molestus. The hypothesises were:
- H0: The two bioforms chose a preferred prey, molestus prefer humans and pipiens
prefer birds.
- H1: The two bioforms do not have a preferred prey, there is no difference between
molestus and pipiens.
With p=0.05.
2.9 Tree construction
The forward and reverse sequences for each locus and specimen were aligned to a consensus
in the software Assseq. Sequences from all specimens were aligned using Clustal W
implemented in MEGA 11(Tamura, Stecher, and Kumar, 2021). Phylogenetic trees were
calculated using the Neighbor-Joining method (Saitou & Nei 1987). The percentage of
replicate trees in which the associated taxa clustered together in the bootstrap test (500
replicates) are shown next to the branches (Felsenstein 1985). The tree is drawn on scale and
the evolutionary distances were computed using the Tamura-Nei method (Tamura & Nei
1993) and are in the units of the number of base substitutions per site. One tree for each gene,
pk1 and ank2 were make, together with one tree with concatenated sequences for the samples
that were positive for both loci.
103. Results
3.1 European mosquitoes
3.1.1 qPCR results
The aim with the European mosquitoes was to find Wolbachia and map the bacteria and by
finding their relations, see whether colonies interact with each other or not. This was done
with qPCR (2.3), to see if the bacteria were present, and sequencing to determine the genera.
The qPCR gave an average starting quantity value (SQ), telling where the sample pass the
threshold. The SQ vary a lot between different populations (see figure 3), where the English
populations had both more similar and higher values than the rest. All samples with an SQ
above 1000 were used further.
Figure 3: Plot over the qPCR results for each sample. The y-axis was transformed to logarithmic scale for visual
purposes. Each block represents a bioform from a specific location.
The second result from the same qPCR was the difference in level of Wolbachia between the
abdomen and thorax. Since the bacteria affects the reproductive tract of the mosquitoes, it was
Figure 4: Boxplot showing the levels of Wolbachia found in abdomen and thorax based on population. The
figure consists of 10 different populations of Cx. pipiens pipiens, Cx. pipiens molestus and Cx. quinquefasciatus.
“Ab.” represents abdomen, “Th.” represents thorax.
11believed there would be higher levels in the abdomen than in the thorax, this was for 7 out of
10 populations true (see figure 4). The two populations where this was not true were
Brookwood molestus and Caldbeck pipiens, together with Brookwood pipiens where it is a
marginal difference. Generally, the Mali population and the Swedish populations had
considerably higher levels in both abdomen and thorax than the rest of the populations,
although there is more variation within the sample size. Cx. quinquefasciatus from Mali had
significantly higher levels than the same species from Thailand.
3.1.2 PCR results
A PCR (2.4) was performed on the abdomen samples passing an SQ value of 1000 to see in
which samples there was a clear band of the pk1 and ank2 genes. The samples with a clear
band and an expected band size of 1334-1349 bp for pk1 and 313-511 bp for ank2 (figure 5)
were sequenced.
Figure 5: Results of the gel electrophoresis done on amplified samples. To the left, pk1 and to the right, ank2.
3.2 Results for Gothenburg mosquitoes
3.2.1 qPCR results – species
The second part was performed on mosquitoes collected in traps, designed to smell like birds,
humans, or nothing, in Gothenburg. Here, the purpose was to see if the two biotypes Cx.
pipiens pipiens and Cx. pipiens molestus chose a preferred prey. To determine the species, a
qPCR (2.7) was performed. All except 14 could be identified, indicating that these 14 are not
Culex mosquitoes. The Gothenburg mosquitoes were also tested for Wolbachia.
Once the species and biotypes were known, a Fisher’s exact test was performed only on the
traps that smelled like humans and bird. The control traps with no smelled where not used in
the test. The test gave a p-value of 0.6448, the H0 could be rejected and H1 accepted,
meaning there was no correlation between biotype and prey.
123.2.2 qPCR results – Wolbachia
The same experiment was performed on the Gothenburg mosquitoes, using qPCR to detect
bacteria. Generally, there were much lower levels of Wolbachia in Gothenburg than in the
European mosquitoes (see figure 6). The 48 with the highest levels were chosen for
sequencing.
Figure 6: Combined qPCR results European and Gothenburg mosquitoes. The values are on logarithmic scale
due to visual purposes. There is a big difference between the values, and generally much lower levels than the
European samples.
3.2.3 PCR results
Even though most of the Gothenburg mosquitoes did not have a higher SQ than 1000, 48
samples were chosen for PCR. The PCR went well with ank2, but not as good with pk1 (see
figure 7). The ones with band on both gels, together with the ones having the clearest bands
on ank2 were sequenced.
Figure 7: PCR results of Gothenburg mosquitoes. Left: ank2, right: pk1. The samples with bands on both gels
together with the samples with clearest bands on ank2 were sent for sequencing.
133.3 Phylogenetic results
The results gave three phylogenetic trees, one for pk1 (see figure 8), one for ank2 (see figure
9), and one concatenated with both genes (see appendix figure 1). Both European and
Gothenburg mosquitoes are included together with reference genes and some old samples.
Using pk1, five wPip-strains could be identified. wPip-I is found in clade one, Cx. pipiens
molestus from Sweden. wPip-II is found in group 1 consisting of bioforms pipiens and
molestus from England, as well as in clade 2 with Cx. pipiens pipiens from Sweden. wPip-III
is found in clade 3, with Cx. quinquefasciatus and Cx. pipiens from Italy, Netherlands,
Thailand, and Mali. wPip-IV is found in group 2 containing molestus from Norway. Lastly,
wPip-V is found in clade 4 with Cx. pipiens pipiens from Italy.
Figure 8: The relationships of Wolbachia based on the pk1 gene. Five wPip-strains where found and all could
be linked into clades and groups.
14ank2 gave a different result (see figure 9). Here, there were less difference between
populations within a country. Like figure 8, the Norwegian mosquitoes form a clade by their
own. The mosquitoes from Italy, Netherlands, Thailand, and Mali form a clade with ank2 as
well. A noticeable difference is that all populations from Sweden and the Netherlands form
one clade with both molestus and pipiens, suggesting there being no, or less difference in
ank2 than in pk1. Another difference is that molestus and pipiens from England is in the same
clade, suggesting no difference in the gene there either.
Figure 9: The relationships of Wolbachia based on the ank2 gene. Four clades could be found using this gene.
154. Discussion
4.1 Different levels of Wolbachia in abdomen and thorax
Understanding the spread of Wolbachia in vector mosquitoes is important and vital when
wanting to understand vector borne diseases. In this study, we could observe several
interesting finds. The first of them being the difference in level of Wolbachia between the
abdomen and thorax. Wolbachia was present in all European mosquitoes, with mostly
relatively high levels. When we compared abdomen and thorax, the hypothesis of abdomen
having higher level was shown to be true, at least in most cases. This is due to Wolbachia
foremost interfering with the reproductive organs of the mosquitoes, located in the abdomen
(Hughes & Rasgon, 2012). In this study we used qPCR after separating abdomen and thorax,
but another method, which would give visualization of exact location of the bacteria, would
be to use Fluorescent in situ hybridization (FISH), as a method for further studies. This has
been proven to be a reliable method in the mosquito Anopheles Gambiae, although with a
different strain of Wolbachia (Hughes & Rasgon, 2012).
4.2 No preferred prey of pipiens nor molestus – hybridization important
The second observation we did was with the Gothenburg mosquitoes, regarding what prey the
two bioforms of Cx. pipiens prefer. It was believed that pipiens, the bioform living above
ground in open, large areas, would prefer to bite birds, and that its opposite, molestus, which
lives underground in narrow spaces, would prefer to bite humans (Haba & McBride, 2022). In
Gothenburg, this could not be seen. We found no correlation between bioform and prey at all.
However, the sample size was relatively small, and the ration between pipiens and molestus
was not perfect, with 12 molestus and 75 pipiens. Regardless the ratio, only two out of 12
molestus chose humans over birds, suggesting that the prey might not matter as much as
thought. A study made in Portugal also found that molestus and pipiens were both mainly
ornithophilic in the studied region, meaning molestus not preferring its expected prey (Gomes
et al. 2013). There are many differences between Gothenburg in northern Europe and
Portugal regarding temperature, humidity et cetera. It is shown that both biotypes occupy the
same above ground regions in southern Europe (Haba & McBride, 2022) and that there is
more hybridization there than in northern Europe where there are larger differences in
climates and habitats (Vogels et al. 2015). However, Europe, especially Scandinavia, is
getting warmer faster than the global average (European Environment Agency, 2021), which
possibly could be the reason of the bioforms behaving more similar. Not only the behavior
changes, but hybridization increases since they occupy the same habitats (Haba & McBride,
2022). Studying the hybridization is vital due to them being bridge-vectors between humans
and birds (Gomes et al. 2013), meaning we could possibly see an increase in WNV further up
in Europe (Brugman et al. 2018) and an introduction of the WNV to Sweden (Hesson et al.
2016).
4.3 Five different wPip-strains identified
Using pk1, we could identify five, I-V, different strains linked to different geographical
locations. The same has been done in previous studies, where they used the same genes pk1,
ank2) due to them being fast evolving markers, identifying five different groups (Dumas et al.
2013) (Nten et al. 2011), meaning these two genes to be fitting for identification. One distinct
observation in our study is that pipiens and molestus from Sweden have different strains of
Wolbachia, where molestus got wPip-I and pipiens got wPip-II, which it shares with both
biotypes from England. It is suggested that wPip evolved into five different groups in Europe
where it probably initially spread due to Cx. pipiens populations there have been showing to
having the highest diversity of all examined locations (Dumas et al. 2013). Being able to
16determine populations using bacteria is important, since it can tell us about the relationships
between populations. For example, pk1 relationships suggests that wPip-1 in Swedish
molestus derived from molestus in Brookwood, England. By continuously monitoring the
movement of Cx. pipiens using Wolbachia, the bacteria existing in at least 99% of the
individuals (Hughes & Rasgon, 2012), we could gain knowledge in the mosquitos’
movements, and thereby get better understanding in pathogen distribution by vectors. There
was less difference between populations based on ank2, it is a shorter gene allowing for less
mutations. Although the Norwegian mosquitoes are separated in both phylogenies, suggesting
they are more incompatible with others, containing the wPip-IV strain, which also have been
found in Turkey and Italy (Duron et al. 2005). However, the relationship between them and
how Norway got wPip-IV while there is wPip-1 and wPip-II in Sweden remains unsolved.
5. Conclusions
We identified that there are higher levels of Wolbachia in abdomen than in thorax in Cx.
pipiens. Our results prove that mosquitoes in Gothenburg are mostly ornithophilic regardless
biotype. Lastly, we could identify five different strains of Wolbachia connected to different
locations, wPip-I-V, which seems to be the five different strains there is. This shows that
Wolbachia is an important bacterium with a suitable toolkit for understanding vector
mosquitoes. Further studies should examine whether there is a trend with molestus preferring
birds in different locations over northern Europe. Finally, investigating relationships between
populations should continue regarding the spread of WNV and other vector borne diseases.
6. Acknowledgements
First, I want to say thank you to Tobias Lilja, my supervisor at SVA who had time for me and
help me with my project. I am very grateful for getting this opportunity and thankful for
everything I have learned during these weeks, and for everything Tobias has taught me. I
would also like to thank Fredrik Sundström at Uppsala university for answering to all my
questions about statistics and coding, it was very helpful. Lastly, thanks to Anders Lindström
at SVA for letting me use his picture of Culex pipiens form my front page.
177. References
7.1 Litterature, articles and webpages
Amraoui F, Tijane M, Sarih M, Failloux A-B. 2012. Molecular evidence of Culex pipiens form
molestus and hybrids pipiens/molestus in Morocco, North Africa. Parasites & Vectors 5:
83.
Brugman VA, Hernández-Triana LM, Medlock JM, Fooks AR, Carpenter S, Johnson N. 2018.
The Role of Culex pipiens L. (Diptera: Culicidae) in Virus Transmission in Europe.
International Journal of Environmental Research and Public Health 15: E389.
Byrne K, Nichols RA. 1999. Culex pipiens in London Underground tunnels: differentiation
between surface and subterranean populations. Heredity 82: 7–15.
Culicidae Classification | Mosquito Taxonomic Inventory. online: https://mosquito-taxonomic-
inventory.myspecies.info/simpletaxonomy/term/6045. Accessed May 26, 2022.
Dumas E, Atyame CM, Milesi P, Fonseca DM, Shaikevich EV, Unal S, Makoundou P, Weill M,
Duron O. 2013. Population structure of Wolbachia and cytoplasmic introgression in a
complex of mosquito species. BMC Evolutionary Biology 13: 181.
Duron O, Boureux A, Echaubard P, Berthomieu A, Berticat C, Fort P, Weill M. 2007. Variability
and Expression of Ankyrin Domain Genes in Wolbachia Variants Infecting the Mosquito
Culex pipiens. Journal of Bacteriology 189: 4442–4448.
Duron O, Bernard C, Unal S, Lagnel J, Bourtzis K, Raymond M, Weill M. 2005. Transposable
element polymorphism of Wolbachia in the mosquito Culex pipiens: evidence of genetic
diversity, super-infection and recombination.
Epstein NR, Saez K, Polat A, Davis SR, Aardema ML. 2021. The urban-adapted underground
mosquito Culex pipiens form molestus maintains exogenously influenceable circadian
rhythms. Journal of Experimental Biology 224: jeb242231.
European Center for disease prevention and control. Weekly updates: 2021 West Nile virus
transmission season. online: https://www.ecdc.europa.eu/en/west-nile-fever/surveillance-and-
disease-data/disease-data-ecdc. Accessed May 25, 2022.
European Environment Agency. Global and European temperatures. online:
https://www.eea.europa.eu/ims/global-and-european-temperatures. Accessed May 22, 2022.
Felsenstein J. 1985. Confidence Limits on Phylogenies: An Approach Using the Bootstrap.
Evolution 39: 783–791.
Flores HA, O’Neill SL. 2018. Controlling vector-borne diseases by releasing modified
mosquitoes. Nature Reviews Microbiology 16: 508–518.
Gerth M, Gansauge M-T, Weigert A, Bleidorn C. 2014. Phylogenomic analyses uncover origin
and spread of the Wolbachia pandemic. Nature Communications 5: 5117.
18Gomes B, Sousa CA, Vicente JL, Pinho L, Calderón I, Arez E, Almeida AP, Donnelly MJ, Pinto
J. 2013. Feeding patterns of molestus and pipiens forms of Culex pipiens (Diptera:
Culicidae) in a region of high hybridization. Parasites & Vectors 6: 93.
Haba Y, McBride L. 2022. Origin and status of Culex pipiens mosquito ecotypes. Current
Biology 32: R237–R246.
Hesson JC, Schäfer M, Lundström JO. 2016. First report on human-biting Culex pipiens in
Sweden. Parasites & Vectors 9: 632.
Hughes GL, Rasgon JL. 2012. Chapter 9 - Wolbachia Infections in Arthropod Hosts. In: Vega
FE, Kaya HK (ed.). Insect Pathology (Second Edition), pp. 351–366. Academic Press, San
Diego.
Inácio da Silva LM, Dezordi FZ, Paiva MHS, Wallau GL. 2021. Systematic Review of
Wolbachia Symbiont Detection in Mosquitoes: An Entangled Topic about Methodological
Power and True Symbiosis. Pathogens (Basel, Switzerland) 10: 39.
Mike Service. Medical Entomology for Students. Cambridge University. 2008. pp. 53–54. ISBN
978-0-521-70928-6.
Mosquitoes | National Geographic. online April 11, 2010:
https://www.nationalgeographic.com/animals/invertebrates/facts/mosquitoes. Accessed
April 4, 2022.
mosquito | Description, Life Cycle, & Facts | Britannica. online:
https://www.britannica.com/animal/mosquito-insect. Accessed May 4, 2022.
Nten C, Delsuc F, Pasteur N, Weill M, Duron O. 2011. Diversification of Wolbachia
Endosymbiont in the Culex pipiens Mosquito. Molecular biology and evolution 28: 2761–
72.
Ong S. 2021. Wolbachia goes to work in the war on mosquitoes. Nature 598: S32–S34.
Saitou N, Nei M. 1987. The neighbor-joining method: a new method for reconstructing
phylogenetic trees. Molecular Biology and Evolution 4: 406–425.
Shaikevich EV, Vinogradova EB, Bouattour A, Gouveia de Almeida AP. 2016. Genetic diversity
of Culex pipiens mosquitoes in distinct populations from Europe: contribution of Cx.
quinquefasciatus in Mediterranean populations. Parasites & Vectors 9: 47.
Sicard M, Bonneau M, Weill M. 2019. Wolbachia prevalence, diversity, and ability to induce
cytoplasmic incompatibility in mosquitoes. Current Opinion in Insect Science 34: 12–20.
Tamura K, Nei M. 1993. Estimation of the number of nucleotide substituions in the control
region of mitochondrial DNA in humans and chimpanzees. Molecular biology and evolution
10: 512–26.
19Taylor MJ, Bandi C, Hoerauf A. 2005. Wolbachia.Bacterial Endosymbionts of Filarial
Nematodes. In: Baker JR, Muller R, Rollinson D (ed.). Advances in Parasitology, pp. 245–
284. Academic Press,
The Mosquito Hunters: Perspectives from Vector Biologists in the Harvard-MIT Community.
online December 31, 2014: https://sitn.hms.harvard.edu/flash/special-edition-on-infectious-
disease/2014/the-mosquito-hunters-perspectives-from-vector-biologists-in-the-harvard-mit-
community/. Accessed April 4, 2022.
Vector-borne diseases. online: https://www.who.int/news-room/fact-sheets/detail/vector-borne-
diseases. Accessed April 4, 2022.
Vogels CBF, van de Peppel LJJ, van Vliet AJH, Westenberg M, Ibañez-Justicia A, Stroo A,
Buijs JA, Visser TM, Koenraadt CJM. 2015. Winter Activity and Aboveground
Hybridization Between the Two Biotypes of the West Nile Virus Vector Culex pipiens.
Vector-Borne and Zoonotic Diseases 15: 619–626.
West Nile virus. online: https://www.who.int/news-room/fact-sheets/detail/west-nile-virus.
Accessed April 21, 2022.
Werren JH, Baldo L, Clark ME. 2008. Wolbachia: master manipulators of invertebrate biology.
Nature Reviews Microbiology 6: 741–751.
5 vital roles insects play in our ecosystem. online April 23, 2020:
https://www.nationalgeographic.com/magazine/article/insects-play-these-five-vital-roles-
in-our-ecosystem-feature. Accessed April 4, 2022.
7.2 Software
Assseq Mac, GitHub 2019, https://github.com/andersla/Assseq/tree/master/assseq-mac
Glen Stecher, Koichiro Tamura, and Sudhir Kumar (2020) Molecular Evolutionary Genetics
Analysis (MEGA) for macOS. Molecular Biology and
Evolution (https://doi.org/10.1093/molbev/msz312)
(Publication PDF available at https://www.megasoftware.net/citations)
RStudio Team (2020). RStudio: Integrated Development for R. RStudio, PBC, Boston, MA
URL http://www.rstudio.com/.
20Appendix
Appendix figure 1: A phylogeny of ank2 and pk1 combined.
21You can also read

















































