EVALUATION OF THE GENETIC DIVERSITY AND POPULATION STRUCTURE OF POTENTIAL BOSNIAN MOUNTAIN HORSE BASED ON MICROSATELLITE MARKERS
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
R U K AV I N A E T A L . G E N E T I C S T R U C T U R E O F P O T E N T I A L B O S N I A N M O U N TA I N H O R S E 219
RESEARCH ARTICLE
EVALUATION OF THE GENETIC DIVERSITY AND
POPULATION STRUCTURE OF POTENTIAL BOSNIAN
MOUNTAIN HORSE BASED ON MICROSATELLITE
MARKERS
Dunja Rukavina1*, Belma Kalamujić Stroil2, Adaleta Durmić-Pašić2, Mirela Mačkić-
Đurović3, Senka Babić4, Atifa Ajanović5, Naris Pojskić2
1
Department of Biology, University of
Sarajevo-Veterinary Faculty, Sarajevo, ABSTRACT
Bosnia and Herzegovina
2
University of Sarajevo-Institute
for Genetic Engineering and
The focus of this study was to evaluate the genetic diversity and
Biotechnology, Sarajevo, Bosnia and population structure of potential Bosnian mountain horse and to
Herzegovina
determine the possible sources of undoubtedly autochthonous
3
University of Sarajevo-Faculty
of Medicine, Center for Genetics, germplasm using microsatellite markers. Genomic DNA was
Sarajevo, Bosnia and Herzegovina extracted from the whole blood collected from 61 potential
4
Department of Microbiology and
Infectious Diseases, University of Bosnian mountain horse specimens divided into three groups
Sarajevo-Veterinary Faculty, Sarajevo, according to the population of origin and genotyped by using
Bosnia and Herzegovina
5
Department of Chemistry, 17 specific microsatellite markers. Existence of higher genetic
Biochemistry and Physiology, variability was obtained in Group 1 and 3. Group 2 showed
University of Sarajevo-Veterinary
Faculty, Sarajevo, Bosnia and genetic specificity as well as high level of heterogeneity. An
Herzegovina increased inbreeding coefficient was detected in Group 1 and
*Corresponding author: 3 (0.1678 and 0.1813, respectively). Deviation from Hardy-
Prof. Dr. Dunja Rukavina Weinberg equilibrium was noticed in Group 1 and 3. Analysis of
Department of Biology, Veterinary
Faculty, University of Sarajevo, molecular variance showed that from all genetic variation, 5%
Sarajevo/Bosnia and Herzegovina was among populations, 14% among individuals and 81% within
Zmaja od Bosne 90, 71000 Sarajevo,
Bosnia and Herzegovina individuals. Results of the fixation index showed moderate level
Phone: +38733 729 100 of genetic differentiation among groups (5.1%). The results of
ORCID: 0000-0002-7523-2813
Email: dunja.rukavina@vfs.unsa.ba
pairwise differences showed clear genetic differentiation among
Group 2 and other observed groups. Analyses of admixture
Original Submission:
08 February 2021
model showed that second cluster consist almost all individuals
from Group 2, since other clusters consists individuals from
Revised Submission:
09 April 2021 two other observed groups. Factorial Component Analysis has
Accepted: confirmed results of admixture model. Group 2 has indications
15 April 2021 the most genetically characteristics of autochthonous Bosnian
How to cite this article mountain horses. Results of genetic diversity and F statistic data
Rukavina D, Kalamujuć Stroil B,
Durmić-Pašić A, Mačkić-Đurović M, indicate that in the gene pool of Group 1 and 3 is present a part
Babić S, Ajanović A, Pojskić N. 2021. of the gene pool of other breeds.
Evaluation of the genetic diversity
and population structure of potential Keywords: Autochthonous breed, genetic diversity, gene pole,
Bosnian mountain horse based on
microsatellite markers. Veterinaria, genetic structure, microsatellites
70(2), 219-27.
doi.org/10.51607/22331360.2021.70.2.219220 VETERINARIA VO L . 7 0 • I S S U E 2 • 2 0 2 1
INTRODUCTION assessment of genetic diversity of livestock (Semik
and Zabek, 2013). The designation and number of
The only autochthonous horse breed in Bosnia microsatellites that should be used in genotyping
and Herzegovina is Bosnian mountain horse depend on the variability of the breed under study,
(Bosnian pony, Bosnian and Herzegovinian as well as the characteristics of each locus and
mountain horse). In our country, this breed has is still a matter of discussion (Moshkelani et al.,
been selectively bred since 1900s, and today 2011).
is well adapted to ecological and geographical
In this study, 17 specific microsatellite markers,
conditions of the region it inhabits (Žiga and
currently recommended by International Society
Telalbašić, 2009). Bosnian mountain horse has
for Animal Genetics (ISAG) (ISAG, 2014), were
been created by crossing between the Asian wild
employed in order to evaluate the genetic diversity
horse (Equus caballus prsewalskii) and the tarpan
and population structure of potential Bosnian
(Equus caballus gmelini) for a long time. It is
mountain horses, and to determine the possible
believed that with the arrival of the Turks in the
sources of undoubtedly autochthonous germplasm.
area of our country during the Ottoman Empire,
oriental stock has been introduced to the breed
(Žiga and Telalbašić, 2009). Bosnian mountain MATERIAL AND METHODS
horse is a “warm-blooded” horse breed and, based
on its appearance, it belongs to the ponies (weight Genomic DNA was extracted from the
is 250-350 kg, height at the withers and a short whole blood collected from 61 specimens
range from 135 to 145 cm, straight back, sloping morphologically described as Bosnian mountain
quarters, wide and deep chest muscular neck, horse. The specimens were divided into three
with long sloping shoulders) (Dekic et al., 2014). groups according to the population of origin:
This breed is used for light draft, light farm work, Group 1 = 28 individuals (Bosnian mountain
pack and riding and is very surefooted on a terrain horses from Herzegovina region); Group 2 = 20
unsuitable for motor vehicles (Dekic et al., 2014). individuals (Bosnian mountain horses from the
It is important to evaluate the genetic diversity stud „Borike”) and Group 3 = 13 individuals
and population structure of potential Bosnian (Bosnian mountain horses from the other parts
mountain horse in order to develop conservation of Bosnia and Herzegovina). Blood samples (3
programs. This population has small contribution ml) were collected from v. jugularis using the
to the global diversity, but should be maintained sterile venipuncture needles and EDTA vacuum
because of other values, such as cultural value or containers.
utility.
Isolation of genomic DNA was done according to
The most commonly used class of genetic markers the modified protocol for the isolation of DNA from
for genetic diversity studies in livestock are the human blood by salting-out method (Miller et
microsatellites (Fornal et al., 2013; Semik and al., 1988). Modified protocol: 3 ml of blood; 10 ml
Zabek, 2013). Microsatellite loci constitute an of Lysis buffer; 4 ml of PBS; 4 ml of Kern-lysis
informative source concerning population history, buffer; 150 μl of 20% SDS; 100 μl of protease and
structure and genetic diversity. Polymorphism of 0,5 ml 6 M NaCl. The concentration of isolated
microsatellites still plays an important role in the DNA was determined by spectrophotometry, usingR U K AV I N A E T A L . G E N E T I C S T R U C T U R E O F P O T E N T I A L B O S N I A N M O U N TA I N H O R S E 221
UV mini - 1240 (Shimadzu) spectrophotometer. RESULTS
Improved StockMarks® Equine Genotyping Kit
(Applied Biosystems), designed for simultaneous All equine microsatellite markers reported in the
amplification of 17 horse microsatellite markers, study were amplified successfully. Number of
was used for the analysis of nuclear DNA detected alleles (AN), expected heterozygosity
polymorphism. PCR was performed according (HE), observed heterozygosity (HO), polymorphic
to the manufacturer’s protocol. PCR products information content (PIC), inbreeding coefficient
were analyzed on an ABI PrismTM 310 Genetic (F) and deviation from Hardy-Weinberg equilibrium
Analyzer. Sizing of the amplified fragments was (HWE) are given in Table 1.
performed using GeneMapper ID v3.2 software. In Group 1 the mean number of alleles was 8.5882
Number of different alleles (AN), polymorphic and varied from 5 (HTG6) to 13 (ASB17). The
information content (PIC) (Botstein et al., observed heterozygosity ranged from 0.4643
1980), observed heterozygosity (HO), expected (HTG7, LEX3) to 0.8929 (CA425) with mean of
heterozygosity (HE) (Nei, 1987), inbreeding 0.6757, while the expected heterozygosity ranged
coefficient (F) (Weir, 1996) and deviation from from 0.5134 (HTG6) to 0.8756 (ASB17) with mean
Hardy-Weinberg equilibrium (HWE) (Guo of 0.7949. Deviation from HWE was found in eight
and Thompson, 1992) were calculated using loci. The mean value of inbreeding coefficient was
POWERMARKER 3.25 (Liu and Muse, 2005). 0.1678 (Table 1). The mean number of alleles in
The same software was used for calculating Group 2 was 5.5880, varied from 3 (HTG7) to 10
genetic differentiation (FST) as well as pairwise (ASB17). The observed heterozygosity ranged from
FST (Weir, 1996). To determine genetic variation 0.1360 (HMS3) to 0.9090 (ASB2) with mean of
among populations, among and within individuals, 0.6310, while the expected heterozygosity ranged
AMOVA analysis was performed within from 0.4410 (HMS7) to 0.8530 (ASB17) with mean
PowerMarker v 3.25 (Liu and Muse, 2005). of 0.6900. Deviation from HWE was found in three
loci, while the mean value of inbreeding coefficient
STRUCTURE v.2.3.3 software was used for
was 0.0770 (Table 1). In Group 3 the mean number
population structure analysis (Pritchard et al.,
of alleles was 6.7647 and varied from 2 (HMS1) to
2000). Estimation of the most likely K (ΔK) value
12 (ASB17). The observed heterozygosity ranged
was performed according to Evanno et al. (2005)
from 0.1538 (HTG6) to 0.9231 (HMS3) with
using STRUCTURE HARVESTER software (Dent
mean of 0.6290, while the expected heterozygosity
and von Holdt, 2012). Burn-in period was 100,000
ranged from 0.1450 (HTG6) to 0.8698 (VHL20)
with 200,000 iterations. Concerning the genetic
with mean of 0.7334. In five loci was detected
divergence of analyzed populations, the Factorial
the deviation from HWE and the mean value of
Correspondence Analysis (FCA) was performed
inbreeding coefficient was 0.1813 (Table 1).
within the Genetix 4.05 software (Belkhir et al.,
1996–2004). Based on all observed parameters, the highest
microsatellite diversity in Group 1 showed ASB2
locus, in Group 2 HMS2 locus and in Group 3
locus VHL20. Locus HTG6 was the least diverse
in Groups 1 and 3, while HMS7 locus was the least
diverse in Group 2.222
Table 1. Number of detected alleles (AN), expected heterozygosity (HE), observed heterozygosity (HO), polymorphic information content (PIC),
inbreeding coefficient (F) and deviation from Hardy-Weinberg equilibrium (HWE) at 17 microsatellite loci in Bosnian mountain horses
Group 1* Group 2** Group 3***
Locus AN HE HO PIC F HWE AN HE HO PIC F HWE AN HE HO PIC F HWE
VETERINARIA
VHL20 9 0.82 0.643 0.801 0.233 0 4 0.706 0.545 0.653 0.227 0.13 10 0.8698 0.8462 0.8564 0.0671 0.06
HTG4 8 0.701 0.821 0.668 -0.15 0.86 5 0.728 0.864 0.678 -0.19 0.4 6 0.7189 0.8462 0.6866 -0.138 0.922
AHT4 9 0.856 0.786 0.839 0.1 0.03 6 0.752 0.818 0.71 -0.09 0.23 7 0.7811 0.7692 0.7511 0.0551 0.257
HMS7 8 0.802 0.607 0.776 0.26 0.14 5 0.441 0.455 0.409 -0.03 0.35 6 0.784 0.8462 0.7511 -0.039 0.1
HTG6 5 0.513 0.536 0.489 -0.03 0.34 4 0.604 0.591 0.527 0.022 0.48 3 0.145 0.1538 0.1399 -0.021 1
AHT5 8 0.772 0.607 0.74 0.231 0.02 6 0.698 0.727 0.65 -0.04 0.65 8 0.8225 0.6923 0.7999 0.197 0.374
HMS6 6 0.784 0.714 0.752 0.107 0.67 6 0.701 0.591 0.651 0.158 0.61 5 0.6893 0.3846 0.6425 0.4737 0.013
ASB23 10 0.824 0.63 0.802 0.254 0.02 5 0.595 0.545 0.554 0.083 0.19 7 0.7899 0.2308 0.7633 0.7273 0
VO L . 7 0 • I S S U E 2 • 2 0 2 1
ASB2 10 0.859 0.821 0.844 0.062 0.1 8 0.803 0.909 0.777 -0.13 0.49 7 0.8343 0.8462 0.8133 0.0258 0.202
HTG10 9 0.867 0.786 0.852 0.111 0.23 8 0.79 0.55 0.761 0.304 0.02 9 0.8402 0.5385 0.8212 0.3935 0.002
HTG7 7 0.816 0.464 0.79 0.446 0 3 0.615 0.639 0.536 -0.04 0.63 6 0.784 0.5385 0.7511 0.3488 0.003
HMS3 8 0.824 0.5 0.805 0.409 0 6 0.707 0.136 0.673 0.807 0 7 0.7959 0.9231 0.7675 -0.121 0.334
HMS2 9 0.853 0.75 0.837 0.139 0.29 7 0.825 0.864 0.801 -0.05 0.23 7 0.7959 0.6923 0.7703 0.1692 0.062
ASB17 13 0.876 0.714 0.864 0.202 0.03 10 0.853 0.591 0.837 0.308 0 12 0.8195 0.7692 0.8028 0.1011 0.816
LEX3 10 0.837 0.464 0.821 0.46 0 4 0.681 0.545 0.617 0.199 0.12 8 0.8284 0.3846 0.8062 0.5636 0
HMS1 7 0.722 0.75 0.692 -0.02 0.29 4 0.584 0.682 0.52 -0.17 0.37 2 0.497 0.6154 0.3735 -0.2 0.605
CA425 10 0.789 0.893 0.761 -0.11 0.19 4 0.64 0.682 0.58 -0.07 0.88 5 0.6716 0.6154 0.6293 0.1233 0.506
Mean 8.6 0.795 0.676 0.772 0.168 5.59 0.69 0.631 0.643 0.077 6.765 0.7334 0.629 0.7015 0.1813
*- Bosnian mountain horses from Herzegovina region; **- Bosnian mountain horses from the stud „Borike”; ***- Bosnian mountain horses from the other parts
of Bosnia and HerzegovinaR U K AV I N A E T A L . G E N E T I C S T R U C T U R E O F P O T E N T I A L B O S N I A N M O U N TA I N H O R S E 223
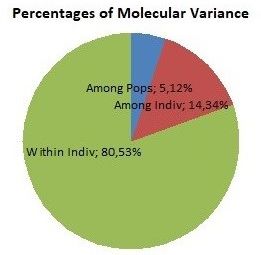
Analysis of Molecular Variance (AMOVA test) STRUCTURE HARVESTER software showed
showed that of the total genetic variation 5% was 2 clusters (K=2). MCMC analyses (admixture
among populations, 14% among individuals and model) showed that the second cluster included
81% within individuals (Figure 1). almost all individuals from Group 2, while other
clusters included individuals from the two other
observed groups (Figure 2). Factorial Component
Analysis (FCA) has confirmed MCMC results
(Figure 3).
1- Group 1; 2- Group 2; 3- Group 3
Figure 1 Results of Analysis of Molecular Figure 2 Results of MCMC analysis (admixture
Variance (AMOVA test) model)
The results of pairwise differences (pFST) for all
tested groups are presented in Table 2. Group 2
showed the differentiation compared to the Group
1 (0.318) and Group 3 (0.374). Differentiation
between Group 1 and 3 was 0.141.
Figure 3 Results of Factorial Component Analysis
Table 2 Results of pairwise differences (pFST) for (FCA)
all tested groups
GROUP GROUP GROUP
1 2 3
GROUP
-
1
GROUP
0.318 -
2
GROUP
0.141 0.374 -
3224 VETERINARIA VO L . 7 0 • I S S U E 2 • 2 0 2 1
DISCUSSION AND CONCLUSION locus ASB17 was also the most variable.
The average HE in Groups 1 and 3 indicated the
In this paper, we carried out evaluation of the genetic
existence of higher genetic variability in these
diversity and population structure of potential
populations. The greatest differences between
Bosnian mountain horses and determination of the
HO and HE in our study were observed for LEX3
possible sources of undoubtedly autochthonous
(Group 1 and 3) and HMS3 (Group 2) loci. The
germplasm using microsatellite markers.
same loci showed the highest deviation from HWE
Indicators of microsatellite diversity, reported in and inbreeding coefficient as well as substantial
the literature for other horse breeds, mostly ranged heterozygote deficit. According to Galov et al.
from 3.3 to 10.7 for number of alleles, from 0.45 to (2013), highly significant deviation from HWE
0.78 for HO and from 0.47 to 0.875 for HE (Canon combined with substantial heterozygote deficit
et al., 2000; Juras and Cothran, 2004; Solis et is likely to indicate a locus-specific genotyping
al., 2005; Behl et al., 2006; Plante et al., 2007; problem due to null alleles. The larger disproportion
Di Stasio et al., 2008; Shasavarani and Rahimi- between observed and expected heterozygosity
Mianji, 2010; Silva et al., 2012; Fornal et al., 2013; was found in Group 1 and 3. Berber et al. (2014)
Berber et al., 2014; Jiskrova et al., 2016; Jemmali reported that larger disproportion between
et al., 2017; Seyedebadi and Sofla, 2017). Results observed and expected heterozygosity could be
of microsatellite diversity for all investigated an indicator of conversely population subdivision
groups are consistent with the results published in reduction or within population, inbreeding.
previous researches.
Genetic markers with PIC values higher than
It was found that the number of detected alleles 0.5 in population genetic analysis are normally
and heterozygosity levels in Group 2 were within considered to be informative (Shasavarani and
the range for Arabian horse (Rukavina et al., 2015) Rahimi-Mianji, 2010). Mean PIC values in all
and Thoroughbred horse populations (Rukavina et investigated groups included in our study were
al., 2016), while the same observed parameters in above this level. Botstein et al. (1980) reported
Group 1 and 3 showed higher values than in “pure” that when the expected heterozygosity was above
breeds observed in the mentioned studies. 0.6 and PIC above 0.5, they s were considered the
Common and simple measure of genetic diversity markers with high diversity. Therefore, chosen
is the number of alleles per loci. In some cases, markers used in this study, are appropriate for the
number of alleles per loci may be more informative analysis of genetic diversity.
than genetic heterozygosity. The differences Deviation from HWE in Group 1 was found in
among breeds, populations and their mean number eight loci, in Group 2 in three and in Group 3 in
of alleles could depend on the analyzed number of five loci. Possible causes for disequilibrium in
alleles, number of samples as well as the population Group 1 and 3 were small population size and
structure. In our study, the lowest mean number of inbreeding. According to Putnova et al. (2019),
alleles was observed in Group 2. In all investigated the deviation could be explained by the effects of
groups, locus ASB17 showed the highest number inbreeding, random influences, individual stallions
of alleles. In Lithuanian (Juras and Cothran, 2004), and selective breeding. Statistically significant
Hucul (Fornal et al., 2013), Halla (Seo et al., 2016) deviation from HWE detected for only three loci
and Tunisian horse breeds (Jemmali et al., 2017) (HTG10, HMS3 and ASB17) in Group 2 supportsR U K AV I N A E T A L . G E N E T I C S T R U C T U R E O F P O T E N T I A L B O S N I A N M O U N TA I N H O R S E 225
the starting assumption that there is no inbreeding gene pool of Group 1 and 3 is present a part of the
among the animals in the observed group. gene pool of other breeds, and that these groups
probably cannot be treated as a complete purebred.
An increased inbreeding coefficient was detected
Clear differentiation of Group 2 compared to other
in Group 1 and 3 (0.1678 and 0.1813, respectively).
investigated groups is not a product of inbreeding
High level of inbreeding coefficient in these groups
or an increase in heterozygosity. Also, almost all
could be due to small sample size, or could indicate
of the analyzed loci in Group 2 were in Hardy-
shortage of heterozygote. Inbreeding coefficient
Weinberg equilibrium.
values for Group 2 (with mean of 0.0770) did
not differ significantly from zero, and indicate no If we consider that the level of inbreeding in Group
shortage of heterozygotes in investigated group. 2 was the lowest, and that almost all of the loci
are within Hardy-Weinberg equilibrium, almost no
The results of AMOVA test indicate a differentiation
admixture, as well as the lowest, but relatively high
among investigated groups of horses. Results of
level of genetic diversity compared to the observed
the Fixation index (FST) (5.1 %) (data not shown)
groups, we can conclude that the individuals from
showed a moderate level of genetic differentiation
Group 2 show the most probable characteristics
among groups, and indicated differentiation
of autochthonous Bosnian mountain horses. In
among tested groups. Value of FST, obtained in this
support of the above, significant data indicate a
study may indicate the presence of the gene flow
carefully bred Bosnian mountain horses from the
between investigated groups. Also, our results of
stud „Borike”. Most likely, it shares the highest
AMOVA test and FST were similar and demonstrate
percentage of genetic similarities with the original
differentiation among tested groups, which
gene pool of autochthonous Bosnian mountain
probably couldn’t be treated as one population.
horses. Other investigated groups did not show
Intermediate levels of gene flow may be due to the
these characteristics. Obtained results suggest that
absence of homozygosity, or the presence of gene
in the gene pool of Group 1 and 3 is present a part
flow between groups may be due to their common
of the gene pool of other breeds, and that these
origin. The values of the genetic differentiation
groups probably cannot be treated as a complete
of 10% are typical for differentiation between
purebred. The obtained results can be very useful
geographically isolated populations of horses as
to breeders as well as for the development of
well as within the different breeds (Zabek et al.,
conservation programs and management strategies
2005).
in order to protect and preserve our autochthonous
Results of pFST showed genetic differentiation breed.
between Group 2 and other observed groups.
Higher inter-group genetic differentiation, observed
among the Group 2 and other investigated groups, CONFLICT OF INTEREST
indicates that the Group 2 is differentiated in
relation to Group 1 and 3. The authors declare that there is no conflict of
It is noteworthy that Group 1 and 3 showed greater interest.
genetic variability than Group 2, probably because
of its wide genetic base. Our results of microsatellite
diversity and F statistic data indicate that in the226 VETERINARIA VO L . 7 0 • I S S U E 2 • 2 0 2 1
REFERENCES Jemmali B, Haddad MM, Barhoumi N, Tounsi S, Lasfer F,
Trabels A, et al. 2017. Genetic diversity in Tunisian horse
breeds. Arch Anim Breed, 60, 153-60.
Behl R, Behl J, Gupta N, Gupta SC, Ahlawat SPS, Ragnekar
M, Ahmed Z. 2006. Genetic characterization of Zanskari Jiskrova I., Vrtkova I, Prausova M. 2016. Genetic diversity of
breed of horse. J Genet, 85, 199-203. populations of Akhal-Teke horses from the Czech Republic,
Russia, Estonia and Switzerland. Acta Univ Agric Silvic
Belkhir K, Borsa P, Chikhi L, Raufaste N, Bonhomme F.
Mendelianae Brunn, 64, 461-6.
1996–2004. GENETIX 4.05, logiciel sous Windows TM
pour la ge´ne´tique des populations. Laboratoire Genome, Juras R, Cothran EG. 2004. Microsatellites in Lithuanian
Populations, Interactions, CNRS UMR 5000, Universite´ native horse breeds: usefulness for parentage testing.
de Montpellier II, Montpellier, France. URL: http://www. Biologija, 4, 6-9.
genetix.univmontp2.fr/genetix/intro.htm.
Liu K, Muse S. 2005. Powermarker: Integrated analysis
Berber N, Gaouar S, Leroy G, Kididi S, Tabet Aouel N, enviroment for genetic marker data. Bioinformatics, 21,
Saidi Mehtar N. 2014. Molecular characterization and 2128-29.
differentiation of five horse breeds raised in Algeria using
Miller SA, Dykes DD, Polesky HF. 1988. A simple salting-out
polymorphic microsatellite markers. J Anim Breed Genet,
procedure for extracting DNA from human nucleated cells.
131, 387-94.
Nucleic Acids Res, 16, 1215.
Botstein D, White RL, Skolnick M, Davis RW. 1980.
Moshkelani S, Rabiee S, Javaheri-Koupaei M. 2011.
Construction of a genetic linkage map in man using restriction
DNA fingerprinting of Iranian Arab horse using fourteen
fragment length polymorphisms. Am J Hum Genet, 32, 314-
microsatellites marker. Res J Biol Sci, 6, 402-5.
31.
Nei M. 1987. Molecular Evolutionary Genetics. Columbia
Canon J, Checa ML, Vega-Pla JL, Vallejo M, Dunner S. 2000.
University Press, New York.
The genetic structure of Spanish Celtic horse breeds inferred
from microsatellite data. Anim Genet, 31, 39-48. Plante Y, Vega-Pla JL, Lucas Z, Colling D, deMarch B,
Buchanan F. 2007. Genetic diversity in a feral horse
Dekic R., Ivanc A, Cetkovic D, Dolicanin Z, Obradovic S.
population from Sable Island, Canada. J Hered, 98, 594-602.
2014. Hematology of Bosnian pony. Bulgarian J Agricult Sci,
20(5), 1237-44. Pritchard J, Stephens M, Donnelly P. 2000. Inference of
population structure using multilocus genotype data. Genetics,
Dent EA, vonHoldt B M. 2012. STRUCTURE HARVESTER:
155, 945–59.
a website and program for visualizing STRUCTURE output
and implementing the Evanno method. In: Conservation Putnová L, Štohl R, Vrtková I. 2019. Using nuclear
Genetics Resources, 4(2), 359-61. microsatellite data to trace the gene flow and population
structure in Czech horses. Czech J Anim Sci, 64, 67-7.
Di Stasio L, Perrotta G, Blasi M, Lisa C. 2008. Genetic
characterization of the Bardigiano horse using microsatellite Rukavina D, Hasanbašić D, Durmić-Pašić A, Kalamujić
markers. Ital J Anim Sci, 7, 243-50. B, Zahirović A, Ramić J, et al. 2015. Genetic Diversity of
Arabian Horse from Stud “Borike” (Bosnia and Herzegovina)
Evanno G, Regnaut S, Goudet J. 2005. Detecting the number
Using Microsatellite Markers. J Vet Sci, 1, 21-5.
of clusters of individuals using the software STRUCTURE: a
simulation study. Mol Ecol, 14(8), 2611-20. Rukavina D, Hasanbašić D, Ramić J, Zahirović A, Ajanović A,
Beganović K, et al. 2016. Genetic diversity of Thoroughbred
Fornal A, Radko A, Piestrzynska-Kajtoch A. 2013. Genetic
horse population from Bosnia and Herzegovina based on 17
polymorphism of Hucul horse population based on 17
microsatellite markers. Jpn J Vet Res, 64, 215-20.
microsatellite loci. Acta Biochim. Pol, 60, 761-65.
Semik E, Ząbek T. 2013. Application of 7 STR markers for
Galov A, Byrne K, Gomerčić T, Duras M, Arbanasić H,
parentage testing and genetic distance study of Equidae. Ann
Sindičić M, et al. 2013.Genetic structure and admixture
Anim Sci, 13, 229-39.
between the Posavina and Croatian Coldblood in contrast to
Lipizzan horse from Croatia. Czech J Anim Sci, 58, 71-8. Seo JH, Kyung-Do Park KD, Hak-Kyo Lee HK, Hong-Sik
Kong HS. 2016. Genetic diversity of Halla horses using
Guo SW, Thompson EA. 1992. Performing the exact test of
microsatellite markers. J An Sci Technol, 58(40), 1-5.
Hardy-Weinberg proportion for multiple alleles. Biometrics,
48, 361-72. Seyedabadi HR, Savar Sofla S. 2017. Microsatellite Analysis
for Parentage Verification and Genetic Characterization of the
ISAG, 2014. ISAG 2014 Workshop Report. http://www.isag.
Turkmen Horse Population. Kafkas Univ Vet Fak Derg, 23,
us/committees.asp (accessed 2. 17. 2016)
467-71.R U K AV I N A E T A L . G E N E T I C S T R U C T U R E O F P O T E N T I A L B O S N I A N M O U N TA I N H O R S E 227
Shasavarani H, Rahimi-Mianji G. 2010. Analysis of genetic Weir BS. 1996. Genetic data analysis II, Sunderland, MA:
diversity and estimation of inbreeding coefficient within Sinauer Associates, Inc.
Caspian horse population using microsatellite markers. Afr J
Zabek T, Nogaj A, Radko A, Nogaj J, Slota E. 2005. Genetic
Biotechnol, 9, 293-9.
variation of Polish endangered Bilgoraj horses and two
Silva ACM, Paiva SR, Alboquerque MSM, Egito AA, Santos common horse breeds in microsatellite loci. J Appl Genet,
SA, Lima FC, et al. 2012. Genetic variability in local Brazilian 46(3), 299-305.
horse lines using microsatellite markers. Genet Mol Res, 11,
Žiga E, Telalbašić R. 2009. The Bosnian Mountain horse – a
881-90.
monograph. IP Šahimpašić, Sarajevo, 350.
Solis A, Jugo BM, Meriaux JC, Iriondo M, Mazon LI, Aguirre
AI, et al. 2005. Genetic diversity within and among four South
European native horse breeds based on microsatellite DNA
analysis: Implications for conservation. J Hered, 96(6), 670-8.
PROCJENA GENETIČKE RAZNOLIKOSTI I STRUKTURE POPULACIJE
POTENCIJALNOG BOSANSKOG BRDSKOG KONJA NA OSNOVI
MIKROSATELITSKIH MARKERA
SAŽETAK
Cilj našeg istraživanja jeste procijeniti genetičku raznolikost i strukturu populacije potencijalnog
Bosanskog brdskog konja te odrediti moguće izvore nesumnjive autohtone germplazme korištenjem
mikrosatelistkih markera. Genomska DNA je ekstrahovana iz pune krvi 61 potencijalnog Bosanskog
brdskog konja. Uzorci su podijeljeni u tri grupe prema porijeklu populacije i genotipizirani
korištenjem 17 specifičnih mikrosatelistkih markera. U grupama 1 i 3 je dokazano postojanje veće
genetičke raznolikosti. Grupa 2 je pokazala genetičku specifičnost, kao i visok stupanj heterogenosti.
U grupama 1 i 3 je dokazan povišen koeficijent inbridinga (0.1678 i 0.1813). Odstupanje od Hardy-
Weinbergovog ekvilibrija je uočeno u grupama 1 i 3. Analiza molekularne varijance je pokazala da
se od svih genetičkih varijacija, 5% odnosi na varijacije među populacijama, 14% među jedinkama i
81% unutar jedinki. Rezultati indeksa fiksacije su pokazali umjereni stepen genetičke diferencijacije
među grupama (5.1%). Rezultati razlika među parovima su pokazali jasnu genetičku diferencijaciju
između grupe 2 i ostalih opserviranih grupa. Analize modela mješovitog porijekla jedinki su
pokazale da drugi klaster čine skoro sve jedinke iz grupe 2, dok ostale klastere čine jedinke iz
druge dvije opservirane grupe. Faktorijalna analiza korespondencije je potvrdila rezultate modela
mješovitog porijekla. Zaključak je da grupa 2 posjeduje većinu genetskih karakteristika autohtonog
Bosanskog brdskog konja. Rezultati genetičke raznolikosti i F statistički podaci ukazuju da su u
genskom fondu grupa 1 i 3 prisutni dijelovi genskog fonda ostalih pasmina.
Ključne riječi: Autohtona pasmina, genetička raznolikost, genski fond, genetička struktura,
mikrosatelitiYou can also read

















































