Early ionic events in activation of the mammalian egg
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Reviews of Reproduction (1998) 3, 96–103
Early ionic events in activation of the mammalian egg
Dalit Ben-Yosef1 and Ruth Shalgi2
1Sara Racine IVF Unit, LIS Maternity Hospital, Tel Aviv Sourasky Medical Center, Tel Aviv 64239 and
2Department of Embryology and Teratology, Sackler School of Medicine, Tel Aviv University, Ramat Aviv,
Tel Aviv 69978, Israel
At fertilization of the mammalian egg, the spermatozoon initially binds to and then fuses
with the egg plasma membrane. This critical event activates specific biochemical pathways
within the egg. Activation of the egg induces resumption of meiosis and the start of rapid
embryonic mitotic divisions on the one hand, and cortical granule exocytosis leading to modi-
fication of the zona pellucida and a block to polyspermy on the other. It has been shown in
different systems that changes in intracellular ion concentrations can serve as second mess-
engers of signal transduction mechanisms. The use of specific fluorescence probes, combined
with the image analysis technique, facilitates the measurement of their dynamics in real time
in the living cell and, thereby, assessment of their role in activation of the mammalian egg.
This review focuses on the dynamics of intracellular Ca2+ and pH and their role in transducing
the sperm signal to downstream cell cycle regulators.
The mammalian oocyte enters meiosis during embryonic life. Phillips, 1980; Myles et al., 1994; reviewed by Yanagimachi
The meiotic division proceeds to the first prophase, and is then 1994). The interaction between spermatazoon and egg mem-
arrested at the diplotene stage, when the germinal vesicle (GV), branes initiates a cascade of biochemical events that trigger the
a characteristic nuclear structure, can be observed. Resumption egg to proliferate and develop into a new organism.
of meiosis, also known as oocyte maturation, occurs in re-
sponse to the preovulatory surge of LH, and is followed by
Mechanisms by which a spermatazoon activates an egg
ovulation (Tsafriri and Dekel, 1994). The mature oocyte, com-
monly referred to as the egg, is again arrested at the second
The sperm factor theory
metaphase (MII), and considered a relatively quiet cell, with a
pre-set developmental programme. At fertilization, the sper- This hypothesis proposes that the cellular messenger is gen-
matozoon overcomes this second cell cycle arrest, and initiates erated as a result of fusion between the spermatazoon and egg
the sequence of biochemical events that leads to the rapid membranes and delivery of an activating component from the
embryonic mitotic divisions. These events, triggered within sperm cytosol to the ooplasm. The hypothesis was introduced
the egg at fertilization, are collectively referred to as ‘egg for mammalian spermatozoa by Swann (1990), after micro-
activation’. Cortical granule exocytosis (CGE) is an early event injection of soluble sperm extracts that were capable of inducing
characteristic of egg activation. Completion of CGE leads to early events of egg activation (that is, an increase in intracellu-
modification of the zona pellucida and the block to polyspermy lar Ca2+ concentration ([Ca2+]i) followed by oscillations). The
(Ducibella, 1991, Kline and Stewart-Savage, 1994; T. Raz and cytosolic sperm factor triggers Ca2+ oscillations in several mam-
R. Shalgi, unpublished). Activation by the spermatozoon also malian species, and is not species specific (Homa and Swann,
leads to later events including resumption of meiosis, pronuclei 1994; Wu et al., 1997). A widespread efficacy of the factor was
formation, initiation of DNA synthesis and cleavage (Xu et al., demonstrated by its ability also to induce Ca2+ oscillations in
1994). This review focuses on the dynamics of early ionic neurones and hepatocytes (Berrie et al., 1996). A soluble sperm
changes leading to mammalian egg activation. protein correlated with Ca2+ oscillation-inducing activity in
eggs has been identified, and termed oscillin. Oscillin is an
oligomer with a subunit of 33 kDa that exhibits a specific intra-
How does a spermatazoon activate an egg?
cellular localization at the equatorial segment of the sperm
Fertilization in eutherian mammals takes place in the ampul- head, an appropriate region for egg activation during fusion
lary region of the oviduct. The first interaction between the (Parrington et al., 1996).
spermatozoon and egg is at the zona pellucida, an extracellular The hypothesis requires fusion between the spermatozoon
coat that surrounds all mammalian eggs. The acrosome-reacted and egg before any increase in cellular second messengers
spermatozoon penetrates the zona pellucida, passes the peri- occurs. Using two independent fluorescence methods and
vitelline space and interacts with the plasma membrane of the confocal microscopy, Lawrence et al. (1997) demonstrated that
egg (reviewed by Schultz and Kopf, 1995). This initial inter- sperm–egg fusion is the prelude to the initial Ca2+ increase
action involves specific recognition between components on at fertilization. They estimated the time interval between
the postacrosomal region over the equatorial segment of the sperm–egg fusion and the onset of the [Ca2+]i oscillations
acrosome-reacted sperm head, and the oolemma (Shalgi and as 1–3 min. The clinical intracytoplasmic sperm injection
© 1998 Journals of Reproduction and Fertility
1359-6004/98 $12.50
Downloaded from Bioscientifica.com at 10/14/2023 01:08:56AM
via free access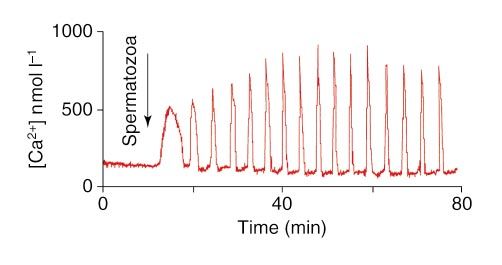
Early ionic events in mammalian egg activation 97
procedure (ICSI) further supports the concept of the spermato-
zoon containing an activating molecule in its cytosol. The ICSI
procedure results in full egg activation, as well as normal
embryonic development, although no contact between sper-
matozoon and egg membranes occurs.
The receptor theory
This hypothesis proposes that recognition between sper-
matozoon and egg receptors, analogous to hormone action in
somatic cells, is sufficient to induce activation. The model
suggests that the egg receptor for the spermatozoon transduces
a signal that mediates a cascade of subsequent cellular events.
It is suggested that the spermatozoon may use a G-protein- Fig. 1. Intracellular Ca2+ oscillations in fertilized rat eggs. A repre-
coupled receptor, or a tyrosine (Tyr) kinase receptor. Although sentative tracing of repetitive Ca2+ transients in rat eggs insemin-
the receptor itself has not yet been identified, studies using ated at the time indicated by the arrow. Eggs were isolated and
specific activators or inhibitors support either of the two loaded with Fura 2. Changes in intracellular Ca2+ were recorded in
receptor-mediated models. It has been demonstrated that com- individual fertilized eggs from insemination until 70 min after
plete activation of the mouse egg in the absence of spermatozoa insemination. An example of an in vitro fertilized zona-free egg
can be achieved by stimulation of an exogenous heterotrimeric that responded to sperm attachment by repetitive Ca2+ transients
and monomeric G-protein-coupled receptor (Williams et al., is shown.
1992; Moore et al., 1993, 1994). Strong support for the G-protein
model has been received from inhibition studies. Microinjection
of the G-protein antagonist, GDPβS, into hamster or mouse
arrested mammalian eggs is approximately 100 nmol l–1, a
eggs blocked several aspects of sperm-induced egg activation
characteristic value within many different inactive somatic
(Miyazaki, 1988; Moore et al., 1994). Some aspects of egg acti-
cells. The initial Ca2+ transient starts within 1–3 min of sperm
vation can occur through the Tyr kinase receptor pathway
fusion. This displays variability in its amplitude and duration,
(starfish: Shilling et al., 1994; Xenopus: Yim et al., 1994; sea
although it is usually lower and lasts longer than the subse-
urchin: Abassi and Foltz, 1994), yet the Tyr kinase receptor in
quent transients (duration 3–6 min, with an amplitude of
mammals has not been examined.
approximately 3–4 times the basal level). This first transient
is followed by a series of narrower Ca2+ transients (duration
Second messengers that mediate egg activation 0.5–1.5 min) of high amplitude (up to 6–8 times the basal
level). Rodent eggs, as well as those of humans, exhibit high
In the last decade, progress has been made in the development frequency oscillations (regular peak-to-peak intervals ranging
of fluorescence probes specific for different ions. These probes, between 2 and 4 min), whereas in bovine and porcine eggs,
combined with the image analysis technique, provide a 15–30 min elapse between two transients. Thus, it appears that
valuable tool with which to follow changes in intracellular the Ca2+ response during fertilization is characterized by a
ion concentrations in living cells, and to assess their role in major difference between the first transient and those that
mammalian egg activation. Some of these ions were tested follow, as well as a major difference between the kinetics of
initially in lower species and were shown to transduce the intracellular Ca2+ increase and its subsequent decrease in a
sperm signal to downstream cell cycle regulators. single transient. The first Ca2+ transient is, as a rule much wider
(fourfold) than the subsequent transient in a series in the same
egg. When analysed by video rate kinetics (25 frames s–1), the
Calcium
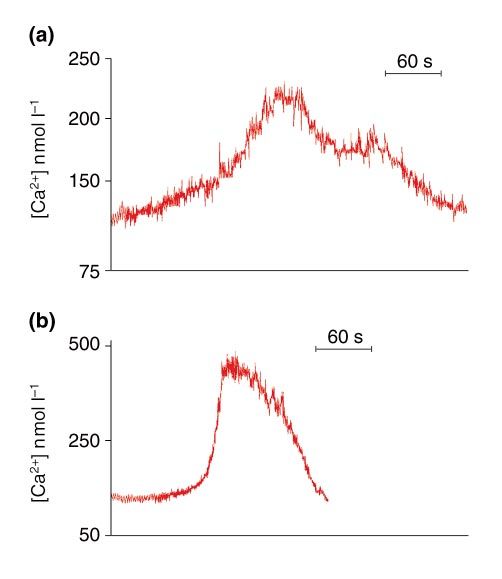
first transient is often biphasic (Fig. 2a). When the duration of
The primary signal transduction event observed at fertilization the first transient was compared with the duration of the
in all mammalian eggs studied, including those of humans, subsequent transients, it was suggestive of a fusion of two
is an increase in [Ca2+]i followed by oscillations (mouse: consecutive transients forming the first transient. This implies
Cuthbertson and Cobbold, 1985; hamster: Miyazaki, 1991; pig: that the mechanisms of intracellular Ca2+ sequestration and/
Sun et al., 1992; cow: Fissore et al., 1992; rat: Ben-Yosef et al., or extrusion are not yet coordinated during the first event.
1993; rabbit: Fissore and Robl, 1993; human: Taylor et al., 1993). Whatever the correct interpretation may be, it is clear from the
A representative pattern is shown (Fig. 1). Dynamic measure- phenomenology that the first transient differs markedly from
ments of [Ca2+]i revealed a marked change accompanying the subsequent transients. This pattern may be a result of
fertilization of the mammalian egg. This first fertilization the initial rise in [Ca2+]i itself, or other parallel cellular events
Ca2+ transient takes the form of a wave originating at the initiated by fertilization.
point of sperm entry (Miyazaki, 1988). A similar Ca2+ wave The shape of the subsequent transients is similar in practic-
was described in Xenopus and sea urchin eggs, in which fer- ally all eggs that responded (Figs 1 and 3). The increase in
tilization produces only a single transient. However, in mam- [Ca2+]i is very slow, and is followed by a rapid increase once
mals, the first distinctive transient is followed by a series a certain threshold Ca2+ concentration is attained (Fig. 2b).
of Ca2+ oscillations of high amplitude and short duration that This corresponds well to the biphasic sensitivity of IP3-
persist for several hours. The basal [Ca2+]i in ovulated MII mediated Ca2+-release described by Bezprozvanny et al. (1991).
Downloaded from Bioscientifica.com at 10/14/2023 01:08:56AM
via free access98 D. Ben-Yosef and R. Shalgi
Fig. 3. Ca2+ oscillations in fertilized human eggs. Aged human
oocytes (48 h after pickup) that failed to be fertilized after in-
semination in vitro were loaded with Fura 2. Changes in cellular
Ca2+ were recorded and a representative pattern of two fertilized
eggs is shown (O. Rufas, D. Ben-Yosef, B. Fisch and R. Shalgi,
unpublished).
transients in in vivo fertilized eggs has been shown to reflect
the physiological events that occur during fertilization in mam-
mals (Ben-Yosef et al., 1993).
It has been shown by studying the role of early changes in
egg Ca2+ during fertilization that a single Ca2+ transient is suf-
ficient to cause exocytosis of most of the cortical granules, lead-
Fig. 2. Video rate kinetics of the first distinctive Ca2+ transient,
ing to an effective block to polyspermy as well as inducing
compared with the subsequent oscillations. Rat eggs loaded with
Fura 2 were inseminated and changes in cellular Ca2+ were re- resumption of the second meiotic division (Jaffe, 1985; Tombes
corded (25 frames s–1). (a) Video rate kinetics of a representative et al., 1992; Vincent et al., 1992, Kline and Stewart-Savage, 1994).
pattern of the first distinctive Ca2+ transient. The transient is often Moreover, blocking the fertilization-induced [Ca2+]i increase
biphasic with an amplitude of about 3–4 times the basal con- inhibits egg activation (Kline and Kline, 1992). However, micro-
centration and a duration of 3–6 min. (b) Video rate kinetics of a injecting mouse eggs with physiological concentrations of in-
representative Ca2+ transient from a series of oscillations. These ositol 1,4,5-tris-phosphate (IP3), resulted in CGE but not in
transients last 0.5–1.5 min each, and increase to 6–8-fold of the basal emission of the second polar body or pronuclei formation
concentration. The regular peak-to-peak intervals range between (Kurasawa et al., 1989). Fine-tuning of the single Ca2+ transient
2 and 4 min. The transient starts with a very slow increase in
demonstrates that low Ca2+ concentrations allow CGE, while
[Ca2+]i, followed by a rapid increase once a certain threshold
higher concentrations are required for cell cycle resumption
[Ca2+]i is attained. However, the decrease of [Ca2+]i to the basal
concentration is biphasic with the slow phase followed by a rapid (Raz et al., 1998). Therefore, it is conceivable that cell cycle
phase. progression is dependent upon the temporal and spatial as-
pects of the Ca2+ transient. Most activators of Ca2+ release
induce a larger Ca2+ increase than the initial response at fertil-
ization, thus leading to complete activation of the egg. It is
The increasing concentrations of [Ca2+]i promote faster release, possible that the first physiological Ca2+ transient is not ade-
forming a feed-forward loop. Moreover, the decrease of quate for complete egg activation, and additional transients are
[Ca2+]i to basal concentrations is, in many eggs, clearly bi- required. In two studies, Ozil and coworkers simulated the
phasic (slow phase followed by a rapid phase), suggesting normal sperm-induced Ca2+ transients by repeated electric
two distinct mechanisms of intracellular Ca2+ removal acting field pulses, evoking Ca2+ increases at a frequency similar to
in tandem. that seen at fertilization (Ozil, 1990; Vitullo and Ozil, 1992).
The repetitive Ca2+ transients continue for several hours They suggested that the oscillations may somehow be coupled
after sperm entry (Cuthbertson and Cobbold, 1985). As fertil- to events occurring during the first cell cycle, and come into
ization progresses, the amplitude and frequency of the Ca2+ play when a prolonged signal is required after a single stimu-
transients decreases and the duration increases (Ben-Yosef lus, as in fertilization.
et al., 1993; Fissore and Robl, 1993) until an absolute cessation A deeper understanding of the mechanism associated with
during entry into interphase, at the time when pronuclei are physiological egg activation in mammals is important es-
forming (Jones et al., 1995). pecially in the field of human IVF. Therefore, it is of interest
Continuous measurements of intracellular Ca2+ changes to verify whether fertilization-associated events observed in
at fertilization have been performed systematically on zona- rodents resemble those characterized by activation of human
free eggs, which are practical for this purpose although sus- eggs. The first evidence of the existence of Ca2+ oscillations in
ceptible to polyspermy. The phenomenon of cyclical Ca2+ human eggs was provided by Taylor et al. (1993) who found
Downloaded from Bioscientifica.com at 10/14/2023 01:08:56AM
via free accessEarly ionic events in mammalian egg activation 99
that insemination of zona-free, as well as zona-enclosed,
human oocytes resulted in cyclical Ca2+ transients (every
(a) 5 min
10–35 min) of short duration (2 min) and high amplitude.
Rufas et al. (1994) demonstrated that aged human oocytes (48 h
490 Fluorescence
old) that failed to fertilize after insemination in vitro could fuse
with and decondense human spermatozoa. Using this model,
intensity
insemination of aged human oocytes was shown to result in
cyclical transients with regular peak-to-peak intervals of
1.5 min and fairly short duration (1.2 min; Fig. 3). Collectively,
these findings demonstrate that aged human oocytes may
serve as a model for the study of early events in human egg
activation.
The mechanism by which the fertilizing spermatozoon
triggers the intracellular Ca2+ increase is not completely de-
(b)
340:380 Fluorescence
termined. The most recent evidence points to sperm-initiated
hydrolysis of the egg phosphatidylinositol 4,5-biphosphate,
intensity ratio
generating IP3, which is known to release Ca2+ from intra-
cellular stores. G-protein-coupled receptors undergo a con-
formational change, which activates its coupled enzyme
phospholipase C (PLCβ) to induce IP3 production. Tyrosine
kinase receptors undergo ligand-mediated dimerization,
allowing the kinase domains to phosphorylate each other on
specific Tyrs. These phosphotyrosines (P-Tyrs) provide docking
sites for cytosolic PLCγ1 (a cytosolic Tyr kinase), which are
thereby translocated to the plasma membrane to catalyse IP3
and diacylglycerol (DAG) production. Dupont et al. (1996) have
demonstrated that mouse oocytes have mRNA encoding Fig. 4. pHi and [Ca2+]i of the same egg during fertilization. Rat
PLCβ1, PLCβ3 and PLCγ. However, when protein expression eggs were double loaded with Fura-2-acetoxymethyl (AM) ester
and 2’,7’-biscarboxyethyl-5(6) carboxyfluorescein (BCECF)-AM.
was studied, only PLCγ could be immunodetected. A specific
Simultaneous recording at four wavelengths (340, 380, 440, 490 nm),
PLC inhibitor (U73122) exerted an inhibitory effect on oocytes for [Ca2+]i and pHi in the same egg, was performed. (a) 490 fluor-
activated by spermatozoa, as well as on oocytes activated escence intensity of two representative fertilized eggs (double-
by acetylcholine, which stimulates Ca2+ spiking through PLC loaded). The emission in 490 nm, the pH-sensitive wavelength is
activation. Collectively, the above data suggest that the unchanged. Therefore, it can be concluded that no change in intra-
activation of PLC to generate IP3 plays a critical role in cellular pH accompanies fertilization. (b) 340:380 fluorescence in-
fertilization. tensity ratio of the same two fertilized eggs as in (a). The exact time
The site of Ca2+ release and sequestration is generally ac- of the first [Ca2+]i transient reflects sperm attachment to the
cepted as the endoplasmic reticulum. Mehlmann et al. (1995) oolemma and confirms fertilization of the specific monitored egg.
demonstrated that competence to undergo normal activation at
fertilization is associated with a marked reorganization of the
egg endoplasmic reticulum, and the development of the IP3- and Robl, 1994). Therefore, a single-pool model for Ca2+ oscil-
induced Ca2+ release system. Nevertheless, two types of mech- lations, assuming a continuous influx of Ca2+ from outside and
anism can mediate Ca2+ release from intracellular stores: IP3- Ca2+-sensitized IICR, was proposed to operate at fertilization of
induced Ca2+ release (IICR) mediated by the IP3 receptor, and mammalian eggs. Although ryanodine receptors are present
Ca2+-induced Ca2+ release (CICR) mediated either by the ryano- and functional in mouse and bovine eggs, release of Ca2+ medi-
dine receptor or by IP3 receptor. Studies on eggs of different ated by this receptor is not essential for sperm-induced egg
classes (sea urchin compared with Xenopus and mammals) activation (Ayabe et al., 1995; Yu et al., 1995). Preincubating
have claimed that different mechanism(s) operate at fertil- eggs with ryanodine, or its addition to oscillating oocytes,
ization. By microinjection of a functional blocking monoclonal indicates that the ryanodine receptor may be involved in
antibody to the IP3 receptor of the Ca2+ release channel, the modulation of oscillations, rather than in their initiation
Miyazaki et al. (1992) demonstrated that IICR from intracellular (Wu et al., 1997). Swann and Lai (1997) postulated that the
stores operates at fertilization in the hamster egg, and that it ‘sperm protein’ oscillin may affect the intracellular Ca2+ stores
is essential for the initiation, propagation, and oscillation of directly by altering the intracellular Ca2+ channel. Once modi-
the sperm-induced Ca2+ transients. When the same antibody fied, the Ca2+ release channels would display an increased sen-
was injected into the mouse egg before insemination, it pre- sitivity to CICR, which may involve IP3 or ryanodine receptor
vented CGE and pronuclear formation, as well as some other or both.
activation-related events (Xu et al., 1994). Likewise, in rabbit Either the sperm factor, or a sperm-receptor signal trans-
eggs, IICR has been shown to participate in the generation of duction mechanism, could lead to IP3 production and initiation
fertilization-associated intracellular Ca2+ increases, and it has of intracellular Ca2+ oscillations. At present, there is no direct
been suggested that the time to reach threshold concentrations evidence to support either of the sperm-mediated models, and
of IP3 dictates the periodicity of the oscillations (Fissore a combination of both theories appears more conceivable. It is
Downloaded from Bioscientifica.com at 10/14/2023 01:08:56AM
via free access100 D. Ben-Yosef and R. Shalgi
Ca2+
channel
G PIP2
R
protein PLC
rane PTK
memb Sperm protein
a DAG
sm
g pla oscillin
Eg Ca2+
Ca2+ PKC
IP3
H+ H+
Alkaline Protein
environment H+ phosphorylation
H+ ER, Ca2+ store
H+ ?
Ca2+
Ca2+ Cortical
Ca2+-dependent enzymes reaction
?
Block to
polyspermy
PTK/PTP
Protein tyrosine
phosphorylation
Protein tyrosine
? dephosphorylation
Cell cycle activation
Fig. 5. Suggested mechanism of sperm-induced egg activation. Binding of a spermatozoon to a G-protein-coupled receptor (G-protein) or,
alternatively, to a tyrosine-phosphorylated (PTK) receptor (R) induces an increase in intracellular Ca2+ concentration ([Ca2+]i). A soluble
sperm protein (oscillin) can also be responsible for this Ca2+ response. The source of Ca2+ for the transients originates from the endoplasmic
reticulum (ER). Moreover, the continuous oscillations also require an influx of Ca2+ from outside to refill the previously emptied stores. The
[Ca2+]i increase induces resumption of meiosis and activation of the cell cycle in one direction, and cortical granule exocytosis leading to
the block to polyspermy, in another direction. The increase in [Ca2+]i activates Ca2+-dependent enzymes, which can facilitate activation of
non-receptor protein tyrosine kinases (PTK) or inactivation of protein tyrosine phosphatases (PTPs). Phosphorylation of specific substrate
proteins further transduces the sperm signal to downstream cell cycle regulators. The naturally high pHi within the ovulated egg provides a
suitable environment in which the enzymes activated by the Ca2+ transient can function. Cortical reaction can be triggered by the initial Ca2+
increase, or by a Ca2+-dependent protein kinase C (PKC). DAG, diacylglycerol; IP3, inositol 1,4,5-tris-phosphate; PIP2, phosphatidylinositol
4,5-biphosphate; PLC, phospholipase C.
also possible that the sperm receptor is composed of multiple
pH
molecules that activate more than one signalling pathway (re-
viewed by Whitaker and Swann, 1993; Schultz and Kopf, 1995; Although the role of Ca2+
as an intracellular regulator in mam-
Kline, 1996). malian egg activation has been studied extensively, very little is
Downloaded from Bioscientifica.com at 10/14/2023 01:08:56AM
via free accessEarly ionic events in mammalian egg activation 101
known about intracellular pH (pHi) changes or function during 1981; Sasaki et al., 1992). This could be explained by the fact
this stage of mammalian egg development. that, in Xenopus, the eggs develop in fresh water where the
In different systems, growth factors may act, at least in part, [Na+] is usually much lower than the intracellular [Na+] of the
by increasing pHi. Events participating in cell division are egg. The Na+/H+ exchanger is the only pH regulation mechan-
also controlled by changes in pHi. In studies of the involvement ism known to act in the absence of bicarbonate. Nevertheless,
of pHi in mediating egg activation, it was found that, at least when studying pHi regulation in mammals it is essential to
in several lower species, alkalinization of 0.2–0.4 units ac- work under physiological HCO3– conditions, where additional
companies fertilization (sea urchin eggs, Urechis caupo and acid transporter pH-regulating mechanisms can be identified.
Xenopus; reviewed by Freeman and Ridgway, 1993). In these The limited information available regarding pH regulation
species, it was suggested initially that alkalinization is involved mechanisms in mammalian eggs suggests that mouse eggs and
in regulating later events of egg activation, such as the mi- embryos have only a bicarbonate–chloride exchanger and are
gration of the sperm or egg pronuclei, or the initiation of pro- capable of recovery from an alkaline load only in the presence
tein and DNA synthesis (reviewed by Whitaker and Steinhardt, of HCO3– (Baltz et al., 1990; House, 1994). Our findings of the
1985; Epel, 1990). However, Rees et al. (1995) showed that pHi is ability of rat eggs to maintain their pHi only in the presence of
only one of many signals involved in the activation of protein bicarbonate is in accordance with these studies.
synthesis at fertilization. On the basis of the aforementioned data, we suggest the
The milieu of the egg at the time of fertilization differs following sequence of events leading to mammalian egg
between species that undergo external fertilization (such as sea activation (Fig. 5). Binding of a spermatozoon to a G-protein-
urchins) and mammals, which are fertilized internally. coupled receptor, or alternatively to a Tyr-phosphorylated
Therefore, an experimental system was designed in which the receptor, induces an increase in [Ca2+]i. A soluble sperm pro-
pH of the ampullar fluid of the oviduct, the millieu in which tein that is delivered to the egg cytoplasm upon fusion can also
the spermatozoon fertilizes the egg, could be measured using be responsible for this Ca2+ response. The source of Ca2+ for the
bicarbonate-buffered medium under controlled CO2 conditions initial transient, as well as for the subsequent oscillations,
(Ben-Yosef et al., 1996). The pH of the rat ampullar fluid was originates from the endoplasmic reticulum. Moreover, the
determined as being alkaline (pH 8.0–8.2). In contrast with the continuous oscillations also require an influx of Ca2+ from out-
ampullar fluid, the pH of the follicular fluid is similar to that of side to refill the previously emptied stores, and are produced
serum (pH 7.2–7.3; reviewed by Fisch et al., 1990). In accordance by Ca2+-sensitized IICR. Calcium-induced Ca2+ release medi-
with these results, Maas et al. (1977) reported that monkey ated by the ryanodine receptor may be involved in the modu-
ampullar fluid is more alkaline after ovulation (pH 7.7) than lation of oscillations. The intracellular Ca2+ increase induces
during the follicular phase (pH ~ 7.2). Under physiological con- resumption of meiosis and activation of the cell cycle in one
ditions (pHout = 7.5), the pHi of rat eggs at MII is relatively high direction, and cortical granule exocytosis, which leads to block
(pH = 7.75). However, under the same conditions, the pHi of to polyspermy, in another direction (reviewed by Ducibella,
the GV oocyte is 0.2 pH units lower. These data correlate well 1996). The increase in [Ca2+]i activates Ca2+-dependent en-
with the values obtained for the ampullary fluid and with the zymes (Watanabe et al., 1989; Lorca et al., 1994; Malcov et al.,
assumption that the high concentrations of bicarbonate and K+ 1997) which, through an unknown cascade of biochemical
in the oviductal fluid would tend to increase the pHi of the egg events, facilitate activation of non-receptor protein tyrosine
(Baltz et al., 1990). kinases (PTKs) or, alternatively, inactivation of protein tyro-
The pHi of the rat egg does not change during fertilization, sine phosphatases (PTPs; Ben-Yosef et al., 1998; Talmor et al.,
and fertilization-induced Ca2+ changes do not affect pHi in rat 1998). Phosphorylation of specific substrate proteins further
eggs (Ben-Yosef et al., 1996). Similar results have been observed transduces the sperm signal to downstream cell cycle regu-
in mouse eggs (Kline and Zagray, 1995; Phillips and Baltz, lators. The naturally high pHi within the ovulated egg pro-
1996). In accordance with the assumption of Freeman and vides a suitable environment in which the enzymes activated
Ridgway, (1993), it is expected that, in species such as rats in by the Ca2+ transient at fertilization can function, and no
which the pHi of the egg is relatively high, no further alkalin- further increase in pHi is necessary to initiate changes in egg
ization upon fertilization is required to facilitate subsequent metabolism during fertilization. The cortical reaction can be
activation events. A representative pattern of the intracellular triggered by the initial Ca2+ increase, or by a Ca2+-dependent
pH and Ca2+ in two fertilized eggs is shown (Fig. 4a,b). PKC.
Several studies have attempted to explore the mechanism
of the increase in pHi in those species in which it is known The authors thank Yoram Oron for his thoughtful suggestions and
to accompany fertilization. Initially, it was reported that the the Desktop Publishing Unit in Tel-Aviv University for technical assist-
pHi increase in sea urchin eggs is caused by stimulation of an ance in graphics.
amiloride sensitive Na+–H+ exchanger, which was found to be
Ca2+-calmodulin-dependent (Johnson and Epel, 1976; Shen
and Sui, 1989; Kuhtreiber et al., 1993). However, some of the References
fertilization-triggered pHi increase may also be attributed to Key references are indicated by asterisks.
CO2 release from the egg (Gillies et al., 1981). In Spisula Abassi YA and Foltz KR (1994) Tyrosine phosphorylation of the egg receptor
oocytes, the Na+-dependent acid release is regulated by protein for sperm at fertilization Developmental Biology 164 430–443
Ayabe T, Kopf GS and Schultz RM (1995) Regulation of mouse egg acti-
kinase C (PKC; Dube, 1988). Xenopus oocytes also express the vation: presence of ryanodine receptors and effects of microinjected
Na+/H+ exchanger, yet alkalinization at fertilization in this ryanodine and cyclic ADP ribose in uninseminated and inseminated eggs
species is independent of external Na+ (Webb and Nuccitelli, Development 121 2233–2244
Downloaded from Bioscientifica.com at 10/14/2023 01:08:56AM
via free access102 D. Ben-Yosef and R. Shalgi
Baltz JM, Biggers JD and Lechene C (1990) Apparent absence of Na+/H+ Kline D and Zagray JA (1995) Absence of an intracellular pH change follow-
antiport activity in the two-cell mouse embryo Developmental Biology 138 ing fertilisation of the mouse egg Zygote 3 305–311
421–429 Kuhtreiber WM, Gillot I, Sardet C and Jaffe LF (1993) Net calcium and acid
Ben-Yosef D, Oron Y and Shalgi R (1993) Prolonged, repetitive calcium release at fertilization in eggs of sea urchins and ascidians Cell Calcium 14
transients in rat oocytes fertilized in vitro and in vivo. FEBS Letters 331 73–86
239–242 Kurasawa S, Schults RM and Kopf GS (1989) Egg-induced modifications of
*Ben-Yosef D, Oron Y and Shalgi R (1996) Intracellular pH in rat eggs is not the zona pellcida of mouse eggs: effects of microinjected inositol 1,4,5-
affected by fertilization and the resulting calcium oscillations Biology of trisphosphate Developmental Biology 133 295–304
Reproduction 55 461–468 Lawrence Y, Whitaker M and Swann K (1997) Sperm–egg fusion is the
Ben-Yosef D, Talmor A, Shwartz L, Granot Y and Shalgi R (1998) Tyrosyl- prelude to the initial Ca2+ increase at fertilization in the mouse
phosphorylated proteins are involved in regulation of meiosis in the rat Development 124 233–241
egg Molecular Reproduction and Development 49 176–185 Lorca T, Abrieu A, Means A and Doree M (1994) Ca2+ is involved through
Berrie CP, Cuthbertson KS, Parrington J, Lai FA and Swann K (1996) A type II calmodulin-dependent protein kinase in cyclin degradation and
cytosolic sperm factor triggers calcium oscillations in rat hepatocytes exit from metaphase Biochemical and Biophysical Acta 1223 325–332
Biochemical Journal 313 369–372 Maas DH, Storey BT and Mastroianni L, Jr (1977) Hydrogen ion and carbon
Bezprozvanny I, Watras J and Ehrlich BE (1991) Bell-shaped calcium- dioxide content of the oviductal fluid of the rhesus monkey Fertility and
response curves of Ins(1,4,5)P3- and calcium-gated channels from endo- Sterility 28 981–985
plasmic reticulum of cerebellum Nature 27 751–754 Malcov M, Ben-Yosef D, Glaser T and Shalgi R (1997) Changes in calpain
Cuthbertson KSR and Cobbold PH (1985) Phorbol ester and sperm activate during meiosis in the rat oocyte Molecular Reproduction and Development 48
mouse oocytes by inducing sustained oscillations in cell Ca2+ Nature 316 119–126
541–542 Mehlmann LM, Terasaki M, Jaffe LA, and Kline D (1995) Reorganization of
Dube F (1988) The relationships between early ionic events, the pattern the endoplasmic reticulum during meiotic maturation of the mouse
of protein synthesis, and oocyte activation in the surf clam, Spisula oocyte Developmental Biology 170 607–615
solidissima. Developmental Biology 126 233–241 Miyazaki S (1988) Inositol 1,4,5-trisphosphate-induced calcium release and
Ducibella T (1991) Mammalian egg cortical granules and the cortical reaction. guanine nucleotide-binding protein-mediated periodic calcium rises in
In Elements of Mammalian Fertilization pp 205–231 Ed. PM Wassarman. golden hamster eggs Journal of Cell Biology 106 345–353
CRC Press, Boston Miyazaki S (1991) Repetitive calcium transients in hamster oocytes Cell
Ducibella T (1996) The cortical reaction and development of activation com- Calcium 12 205–216
petence in mammalian oocytes Human Reproduction Update 2 29–49 *Miyazaki S, Yuzaki M, Nakada K, Shirakawa H, Nakanishi S, Nakada S
Dupont G, Mcguinness OM, Johnson MH, Berridge MJ and Borgese F and Mikoshiba K (1992) Block of Ca2+ and Ca2+ oscillation by antibody to
(1996) Phospholipase C in mouse oocytes: characterization of β and γ the inositol 1,4,5-trisphosphate receptor in fertilized hamster eggs Science
isoforms and their possible involvement in sperm-induced Ca2+ spiking 257 251–255
Biochemical Journal 316 583–591 Moore GD, Kopf GS and Schultz RM (1993) Complete mouse egg activation
Epel D (1990) The initiation of development at fertilization Cell Differentiation in the absence of sperm by stimulation of an exogenous G protein-
and Development 29 1–12 coupled receptor Developmental Biology 159 669–678
Fisch B Goldberg I Ovadia J and Tadir Y (1990) Physico-chemical properties Moore GD, Ayabe T, Visconti PE, Schultz RM and Kopf GS (1994) Role of
of follicular fluid and their relation to IVF outcome Journal of In vitro heterotrimeric and monomeric G proteins in sperm-induced activation
Fertilization and Embryo Transfer 7 67–73 of mouse eggs Development 120 3313–3323
Fissore RA and Robl JM (1993) Sperm, inositol trisphosphate, and thimerosal- Myles DG, Kimmel LH, Blobel CP, White JM and Primakoff P (1994)
induced intracellular Ca2+ elevations in rabbit eggs Developmental Biology Identification of a binding site in the disintegrin domain of fertilin re-
159 122–130 quired for sperm–egg fusion Proceedings of the National Academy of Sciences
Fissore RA and Robl JM (1994) Mechanism of calcium oscillations in fer- USA 91 4195–4198
tilized rabbit eggs Developmental Biology 166 634–642 Ozil JP (1990) The parthenogenetic development of rabbit oocytes after
Fissore RA, Dobirnsky JR, Balise JJ, Duby RT and Robl JM (1992) Patterns repetitive pulsatile electrical stimulation Development 109 117–127
of intracellular Ca2+ concentrations in fertilized bovine eggs Biology of Parrington J, Swann K, Shevchenko VI, Sesay AK and Anthony Lai F (1996)
Reproduction 47 960–969 Calcium oscillations in mammalian eggs triggered by a soluble sperm
*Freeman G and Ridgway EB (1993) The role of intracellular calcium and pH protein Nature 379 364–368
during fertilization and egg activation in the Hydrozoan Phialidium. Phillips KP and Baltz J (1996) An intracellular pH change does not accompany
Developmental Biology 156 176–190 egg activation in the mouse Molecular Reproduction and Development 45 52–60
Gillies RJ, Rosenberg MP and Deamer DW (1981) Carbon dioxide efflux Raz T, Ben-Yosef D and Shalgi R (1998) Segregation of the pathway leading
accompanies release of fertilization acid from sea urchin eggs Journal of to cortical reaction from that of cell cycle activation in the rat egg Biology
Cell Physiology 108 115–122 of Reproduction 58 94–102
Homa ST and Swann K (1994) A cytosolic sperm factor trcalcium oscillations Rees BB, Patton C, Grainger JL and Epel D (1995) Protein synthesis increases
and membrane hyperpolarization in human oocytes Human Reproduction 9 after fertilization of sea urchin eggs in the absence of an increase in intra-
2356–2361 cellular pH Developmental Biology 169 683–698
House CR (1994) Confocal ratio-imaging of intracellular pH in unfertilized Rufas O, Fish B, Phillips DM and Shalgi R (1994) Human gamete fusion
mouse oocytes Zygote 2 37–45 test: an experimental model for assessing the fusion potential of sperm
Jaffe LF (1985) The role of calcium explosions, waves, and pulses in activat- from nonfertilizing normospermic patients Fertility and Sterility 62 786–792
ing eggs. In Biology of Fertilization pp 127–165 Eds CB Metz and A Monroy. Sasaki S, Ishibashi K, Nagai T and Marumo F (1992) Regulation mechan-
Academic Press, Orlando, FL isms of intracellular pH of Xenopus laevis oocyte Biochemical and
Johnson JD and Epel D (1976) Intracellular pH and activation of sea urchin Biophysical Acta 1137 45–51
eggs after fertilization Nature 262 661–664 *Schultz RM and Kopf GS (1995) Molecular basis of mammalian egg acti-
Jones KT, Carroll J, Merriman JA, Whittingham DG and Kono T (1995) vation Current Topics in Developmental Biology 30 21–61
Repetitive sperm-induced Ca2+ transients in mouse oocytes are cell cycle Shalgi R and Phillips DM (1980) Mechanism of sperm entry in cyclic ham-
dependent Development 121 3259–3266 sters Journal of Ultrastructure Research 71 154–161
Kline D (1996) Activation of the mouse egg Theriogenology 45 81–90 Shen SS and Sui AL (1989) K+ activity and regulation of intracellular pH in the
Kline D and Kline JT (1992) Repetitive calcium transients and the role of sea urchin egg during fertilization Experimental Cell Research 183 343–352
calcium in exocytosis and cell cycle activation in the mouse egg Shilling FM, Carroll DJ, Muslin AJ, Escobedo JA, Williams LT and Jaffe LA
Developmental Biology 149 80–89 (1994) Evidence for both tyrosine kinase and G-protein-coupled pathways
Kline D and Stewart-Savage JS (1994) The timing of cortical granule fusion, leading to starfish egg activation Developmental Biology 162 590–599
content dispersal, and endocytosis during fertilization of the hamster egg: Sun FZ, Hoyland X, Huang W and Moor RM (1992) A comparison of intra-
an electrophysiological and histochemical study Developmental Biology 162 cellular changes in porcine eggs after fertilization and electroactivation
277–287 Development 115 947–956
Downloaded from Bioscientifica.com at 10/14/2023 01:08:56AM
via free accessEarly ionic events in mammalian egg activation 103
Swann K (1990) A cytosolic sperm factor stimulates repetitive calcium in- Webb DJ and Nuccitelli R (1981) Direct measurement of intracellular pH
creases and mimics fertilization in hamster eggs Development 110 changes in Xenopus eggs at fertilization and cleavage Journal of Cell Biology
1295–1302 91 562–567
*Swann K and Lai FA (1997) A novel signalling mechanism for generating Whitaker M and Steinhardt RA (1985) Ionic signaling in the sea urchin egg
Ca2+ oscillations at fertilization in mammals BioEssays 19 371–378 at fertilization. In Biology of Fertilization pp 168–222 Eds CB Metz and A
Talmor A, Kinsey WH and Shalgi R (1998) Expression and immuno- Monroy. Academic Press, Orlando, FL
localization of p59c-fyn tyrosine kinase in rat eggs Developmental Biology Whitaker M and Swann K (1993) Lighting the fuse at fertilization
194 38–46 Development 117 1–12
Taylor CT, Lawrence YM, Kingsland CR, Biljan MM and Cuthbertson KSR Williams CJ, Schultz RM and Kopf GS (1992) Role of G proteins in mouse
(1993) Oscillations in intracellular free calcium induced by spermatozoa in egg activation: stimulatory effects of acetylcholine on the ZP2 to ZP2f con-
human oocytes at fertilization Human Reproduction 8 2174–2179 version and pronuclear formation in eggs expressing a functional m1
Tombes RM, Simerly C, Borisy GG and Schatten G (1992) Meiosis, egg acti- muscarinic receptor Developmental Biology 151 288–296
vation, and nuclear envelope breakdown are differentially reliant on Ca2+, Wu H, He CL and Fissore RA (1997) Injection of a porcine sperm factor trig-
whereas germinal vesicle breakdown is Ca2+ independent in the mouse gers calcium oscillations in mouse oocytes and bovine eggs Molecular
oocytes Journal of Cell Biology 117 799–811 Reproduction and Development 46 176–189
Tsafriri A and Dekel N (1994) Molecular mechanisms in ovulation. In Molecular Xu Z, Kopf GS and Schultz RM (1994) Involvement of inositol 1,4,5-
Biology of the Female Reproductive System pp 207–258 Ed. JK Findlay. trisphosphate-mediated Ca2+ release in early and late events of mouse
Academic Press, New York egg activation Development 120 1851–1859
Vincent C, Cheek TR and Johnson MH (1992) Cell cycle progression of Yanagimachi R (1994) Mammalian fertilization. In The Physiology of
parthenogenetically activated mouse eggs to ionophore is dependent on Reproduction 2nd edn pp 189–317 Eds E Knobil and JD Neill. Raven Press,
the level of internal calcium Journal of Cell Science 103 389–396 New York
Vitullo AD and Ozil J (1992) Repetitive calcium stimuli drive meiotic re- Yim DL, Opresko LK, Wiley HS and Nuccitelli R (1994) Highly polarized
sumption and pronuclear development during mouse oocyte activation EGF receptor tyrosine kinase activity initiates egg activation in Xenopus.
Developmental Biology 151 128–136 Developmental Biology 162 41–55
Watanabe N, Vande Woude GF, Ikawa Y and Sagata N (1989) Specific pro- Yu C, White KL, Reed WA and Bunch TD (1995) The existence of inositol
teolysis of the c-mos proto-oncogene product by calpain on fertilization of 1,4,5-trisphosphate and ryanodine receptors in mature bovine oocytes
Xenopus eggs Nature 342 505–511 Development 121 2645–2654
Downloaded from Bioscientifica.com at 10/14/2023 01:08:56AM
via free accessYou can also read

















































