Dental pulp stem cells as a therapy for congenital entero neuropathy
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
www.nature.com/scientificreports
OPEN Dental pulp stem cells as a therapy
for congenital entero‑neuropathy
Koichiro Yoshimaru1, Takayoshi Yamaza2*, Shunichi Kajioka3,7*, Soichiro Sonoda2,
Yusuke Yanagi1, Toshiharu Matsuura1, Junko Yoshizumi4, Yoshinao Oda5, Naoko Iwata6,
Chiho Takai6, Shinsuke Nakayama6* & Tomoaki Taguchi1
Hirschsprung’s disease is a congenital entero-neuropathy that causes chronic constipation and
intestinal obstruction. New treatments for entero-neuropathy are needed because current surgical
strategies have limitations5. Entero-neuropathy results from enteric nervous system dysfunction due
to incomplete colonization of the distal intestine by neural crest-derived cells. Impaired cooperation
between the enteric nervous system and intestinal pacemaker cells may also contribute to entero-
neuropathy. Stem cell therapy to repair these multiple defects represents a novel treatment
approach. Dental pulp stem cells derived from deciduous teeth (dDPSCs) are multipotent cranial
neural crest-derived cells, but it remains unknown whether dDPSCs have potential as a new therapy
for entero-neuropathy. Here we show that intravenous transplantation of dDPSCs into the Japanese
Fancy-1 mouse, an established model of hypoganglionosis and entero-neuropathy, improves large
intestinal structure and function and prolongs survival. Intravenously injected dDPSCs migrate to
affected regions of the intestine through interactions between stromal cell-derived factor-1α and
C-X-C chemokine receptor type-4. Transplanted dDPSCs differentiate into both pacemaker cells and
enteric neurons in the proximal colon to improve electrical and peristaltic activity, in addition to their
paracrine effects. Our findings indicate that transplanted dDPSCs can differentiate into different cell
types to correct entero-neuropathy-associated defects.
Hirschsprung’s disease (HSCR) and its allied disorders are congenital entero-neuropathies that occur with an
incidence of ~ 1 per 5000 livebirths in Japan1–3. HSCR is a potentially fatal disorder with clinical features that
include constipation, abdominal distension, megacolon and intestinal o bstruction1–3. The current management
of HSCR involves surgery to remove or bypass affected segments of the bowel4. Nevertheless, many children
suffer from life-long complications due to the dysfunction of ‘healthy’ bowel retained during s urgery5 and the
consequences of additional palliative surgery such as proctectomy and enterostomy. Therefore, novel therapies
are urgently needed to improve outcomes and quality of life in patients with HSCR.
HSCR was first reported in 1888 by Hirschsprung, who described two infants with local defects of enteric
neurons6. The pathogenesis of HSCR is incompletely understood but thought to involve the failure of enteric
neural crest-derived cells (NCCs) to complete their colonization of the distal intestine during foetal development.
Since the initial discovery that enteric neural circuits play an essential role in peristaltic movements of the gut
(the ‘law of the intestine’ proposed by Bayliss and Starling in 1899)7, it has become established that intestinal
motility is coordinated by an intrinsic nervous system. In the 1990s, the interstitial cells of Cajal (ICCs) were
identified as specialized cells that act as an intestinal pacemaker8. ICCs (especially those in the myenteric plexus)
are thought to contribute to the spatial organization of gut motility because of their network-like structure. Coop-
eration between the enteric nervous system (ENS) and ICC ‘pacemaker’ system is essential for the coordination
of complex intestinal m ovements9,10, and this cooperation between the ENS and ICCs is thought to be impaired
1
Department of Pediatric Surgery, Kyushu University Graduate School of Medical Sciences, Fukuoka,
Japan. 2Department of Molecular Cell Biology and Oral Anatomy, Kyushu University Graduate School of Dental
Science, 3‑1‑1 Maidashi, Higashi‑ku, Fukuoka 812‑8582, Japan. 3Department of Clinical Pharmacology, Kyushu
University Graduate School of Medicine, 3‑1‑1 Maidashi, Higashi‑ku, Fukuoka 812‑8582, Japan. 4Department of
Oral and Maxillofacial Surgery, Fukuoka Dental College, Fukuoka, Japan. 5Department of Anatomic Pathology,
Kyushu University Graduate School of Medical Sciences, Fukuoka, Japan. 6Department of Cell Physiology,
Nagoya University Graduate School of Medicine, 65 Tsurumai‑cho, Showa‑ku, Nagoya 466‑8550, Japan. 7Present
address: Department of Pharmaceutical Sciences, International University of Health and Welfare, 137‑1 Enokizu,
Okawa, Fukuoka 831‑8501, Japan. *email: yamazata@dent.kyushu-u.ac.jp; kajioka@uro.med.kyushu-u.ac.jp;
h44673a@nucc.cc.nagoya-u.ac.jp
Scientific Reports | (2022) 12:6990 | https://doi.org/10.1038/s41598-022-10077-3 1
Vol.:(0123456789)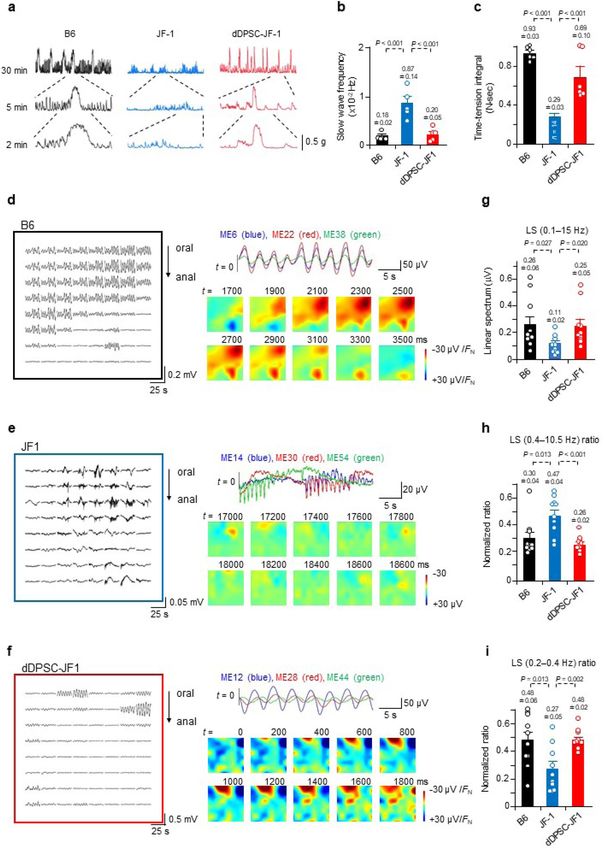
www.nature.com/scientificreports/
in HSCR and its allied disorders. The complex interaction between the ENS and ICCs may explain why familial
RET mutations, which are known to affect colonization of the intestine by NCCs, exhibit incomplete penetrance11.
Dental pulp stem cells derived from deciduous teeth (dDPSCs) are cranial neural crest-derived mesenchymal
stem cells (MSCs) that can differentiate to form structures such as the pulp-dentin complex due to their multiline-
age potential12–14. Our cultured dDPSCs are multipotent cells that express NCC markers and have low levels of
class II HLA, which is important for immune t olerance15. In view of their characteristics, we hypothesized that
dDPSCs may have potential for development into a novel cell-based therapy for entero-neuropathies. Here we
report the effects of intravenous transplantation of dDPSCs into the Japanese Fancy-1 (JF1) mouse, which is an
established animal model of entero-neuropathy. The JF1 mouse carries a homozygous mutation at the piebald
locus (Ednrbs) that encodes the endothelin-B receptor (ETBR), and the phenotype is characterized by a sparse
network of myenteric ganglia, especially in the proximal c olon16–19. We show that intravenously injected dDPSCs
migrate to affected regions of the intestine through interactions between stromal cell-derived factor-1α (SDF1α)
and C-X-C chemokine receptor type-4 (CXCR4). Furthermore, the transplanted dDPSCs differentiate into both
pacemaker cells and enteric neurons in the proximal colon to restore the slow rhythmic potentials observed
in wild-type mice. Our findings imply that congenital entero-neuropathies involve defects of the intestinal
pacemaker system and that dDPSCs have potential for development into a new treatment for HSCR in humans.
Results
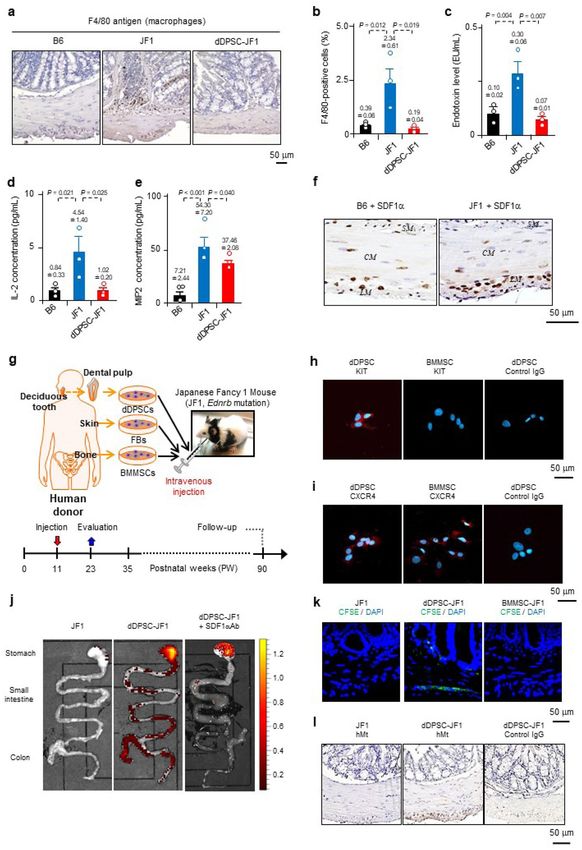
dDPSCs migrate to the intestine. JF1 mice had a shorter body length than age-matched wild-type B6
mice, and their colons exhibited faecal stasis and reduced mucosal thickness (Extended Data Fig. 1a–c). Proxi-
mal colon levels of endothelin receptor type B (Ednrb) mRNA and protein were lower in JF1 mice than in B6
mice (Extended Data Fig. 1d–f). Compared with B6 mouse colon, JF1 mouse colon had higher levels of mac-
rophage infiltration, intestinal endotoxin, interleukin-2 (IL-2) and macrophage inflammatory protein-2 (MIP2)
(Fig. 1a–e), higher faecal bacterial content, a higher proportion of Bacteroidetes (but not Firmicutes or other
bacteria) in the intestinal microbiota, and a lower Gini-Simpson index (Extended Data Fig. 2). Hence, JF1 mouse
colon exhibits intestinal inflammation and bacterial imbalance similar to that observed in Ednrb knock-out
mice20.
Stromal cell-derived factor-1α (SDF1α) is upregulated in inflamed tissue, and its receptor (CXCR4)21 is
expressed in MSCs such as dDPSCs and bone marrow MSCs (BMMSCs). SDF1α-positive cells were abundant
in the myenteric plexus of JF1 mouse proximal colon (Fig. 1f), raising the possibility that dDPSCs and BMMSCs
might target intestinal regions exhibiting hypoganglionosis-related local inflammatory changes. Therefore, our
subsequent experiments investigated the effects of intravenous injection of dDPSCs into JF1 mice at postnatal
week 11 (P11W), and in some experiments we compared the effects of transplanted dDPSCs with those of
intravenously injected BMMSCs or human skin fibroblasts (FBs) (Fig. 1g). Cultured dDPSCs at passage 3 (P3)
expressed KIT (CD117, a receptor tyrosine kinase) and CXCR4 (Fig. 1h,i), retained the characteristics of neural
crest cells, and had low immunogenicity (Extended Data Fig. 3).
We investigated the migration of transplanted dDPSCs (P11W) using cells labelled with 1,1’-dioctadecyl-
3,3,3’,3’-tetramethylindotricarbocyanine iodide (DiR). In vivo fluorescence imaging of dDPSC-transplanted JF1
mice (dDPSC-JF1 mice) revealed that DiR-labelled dDPSCs were widely distributed on day 1 but had migrated
to the intestine by day 7 (Extended Data Fig. 4a). Ex vivo fluorescence imaging 9 weeks after transplantation
revealed a high fluorescence intensity in the intestine (stomach, distal ileum and proximal colon), spleen and
liver (Fig. 1j and Extended Data Fig. 4b). Intraperitoneal administration of a neutralizing antibody to SDF1α
almost completely abolished the intestinal accumulation of DIR-labelled dDPSCs (Fig. 1j and Extended Data
Fig. 4a,b), suggesting that SDF1α-CXCR4 interactions play a role in dDPSC migration to the intestine. Little or
no fluorescence was detected in B6 mice injected with DiR-labelled dDPSCs (Extended Data Fig. 4c), implying
that dDPSCs migrate specifically to regions of hypoganglionosis. Carboxyfluorescein diacetate succinimidyl ester
(CFSE)-labelled dDPSCs were observed in the proximal colon of dDPSC-JF1 mice 3 days after transplantation,
whereas CFSE-labelled BMMSCs were not detected (Fig. 1k). Immunostaining for human mitochondria (hMt)
identified dDPSCs in the proximal colon 12 weeks after transplantation (Fig. 1l). The above results demonstrate
that SDF1α-CXCR4 interactions play a crucial role in the migration of dDPSCs to regions of hypoganglionosis
in JF1 mouse intestine.
dDPSCs regenerate neurons and ICCs. Morphological investigations (Fig. 2a,b) revealed that JF1
mouse proximal colon exhibited poorer development of the intestinal musculature and myenteric region (hae-
matoxylin–eosin staining), fewer myenteric ganglion cells (stained with the neuron-specific marker, HuC/HuD),
and weaker immunostaining for neurofilament M (NFM, a peripheral ganglion marker) and KIT (a pacemaker
interstitial cell marker). These observations suggest that congenital entero-neuropathy is accompanied by poor
development of the pacemaker system22,23. Notably, dDPSC-JF1 mouse colon exhibited increased numbers of
enteric neurons and pacemaker interstitial cells (Fig. 2a,b). Immunohistochemistry experiments in dDPSC-JF1
mice revealed that hMt was co-expressed in some HuC/HuD-positive and NFM-positive neurons in the myen-
teric plexus region (Fig. 2c,d) and in numerous KIT-positive cells (Fig. 2e), indicating that myenteric neurons
and pacemaker cells had developed from dDPSCs.
Since inadequate interactions between trophic factors may contribute to hypoganglionosis1–4, we assessed the
effects of dDPSCs on trophic factors. The concentrations of mouse nerve growth factor (NGF), glial cell-derived
neurotrophic factor (GDNF) and stem cell factor (SCF, a KIT ligand) were similar between B6 and JF1 mice
(Fig. 2f). dDPSC transplantation increased the protein levels of mouse NGF and GDNF and human GDNF, NGF
and SCF (Fig. 2f). These findings are consistent with dDPSC-mediated paracrine actions.
Scientific Reports | (2022) 12:6990 | https://doi.org/10.1038/s41598-022-10077-3 2
Vol:.(1234567890)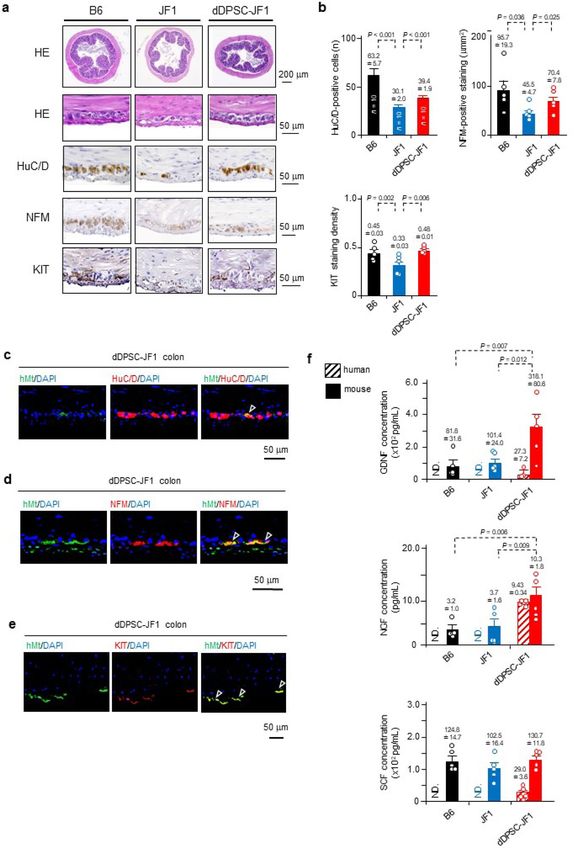
www.nature.com/scientificreports/
Figure 1. Intestinal migration of dDPSCs. (a) Macrophages in the proximal colon. (b) Macrophage percentage
in the proximal colon (mean ± SEM, n = 3). (c–e) Intestinal endotoxin, IL-2 and MIP2 levels (mean ± SEM,
n = 3). (f) Proximal colon SDF1α expression (× 200). CM: circular muscle; LM: longitudinal muscle; SM:
submucosal layer. (g) Experimental schedule. (h,i) Stem cell KIT and CXCR4 expression. j, Mouse intestine (ex
vivo fluorescence imaging; n = 1). (k) CFSE-labelled cells 3 days post-transplantation (proximal colon, × 200).
Representative of ≥ 3 experiments. (l) hMt-containing cells 12 weeks post-transplantation (proximal
colon, × 200). (a,f,h,i,k,l) Representative of ≥ 2 independent experiments.
Scientific Reports | (2022) 12:6990 | https://doi.org/10.1038/s41598-022-10077-3 3
Vol.:(0123456789)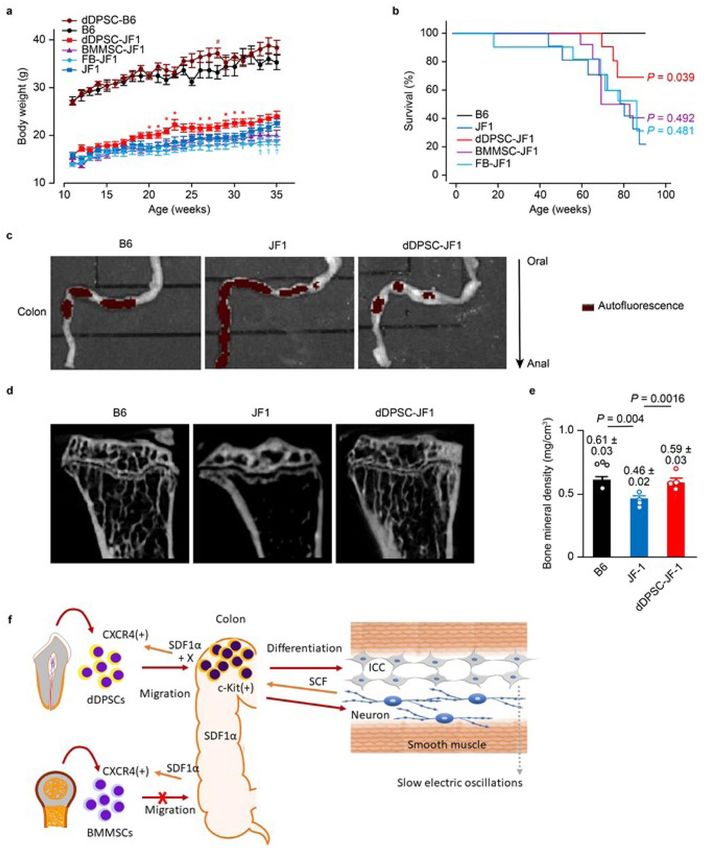
www.nature.com/scientificreports/
Figure 2. Differentiation of transplanted dDPSCs. (a) Proximal colon sections stained with haematoxylin–eosin
(upper: × 40; lower: × 400) or immunostained for HuC/D (neuron-specific marker; × 400), NFM (peripheral
ganglion marker; × 200) or KIT (pacemaker interstitial cell marker; × 200). (b) Number of HuC/D-positive cells
per tissue cross-section (mean ± SEM, n = 10), NFM-positive area in the myenteric plexus (mean ± SEM, n = 5),
and KIT-positive staining density in the proximal colon (mean ± SEM, n = 5). (c–e) Immunostaining for hMt,
HuC/D, NFM and KIT (nuclei counterstained with DAPI). Arrowheads: dDPSC-derived cells. (f) Human and
mouse NGF, GDNF and SCF concentrations (mean ± SEM, n = 5). ND, not detected. (a,c–e) Representative of ≥ 2
independent experiments.
Scientific Reports | (2022) 12:6990 | https://doi.org/10.1038/s41598-022-10077-3 4
Vol:.(1234567890)www.nature.com/scientificreports/
dDPSCs improve proximal colon function. We measured the mechanical and electrical activities of the
second part of the proximal colon (Extended Data Fig. 1b) to assess the functional effects of dDPSC transplan-
tation. Contractile responses to acetylcholine (Ach), endothelin-1 (ET-1) and electrical field stimulation (EFS)
were smaller in JF1 mice than in B6 mice but comparable between dDPSC-JF1 mice and B6 mice (Extended
Data Fig. 5a–d). The relaxation response to a nitric oxide (NO) donor was smaller in JF1 and dDPSC-JF1 mice
than in B6 mice (Extended Data Fig. 5e). Contractile responses to a high concentration (60 mM) of extracellular
K+ were similar among the three groups (Extended Data Fig. 5f), suggesting that the contractile ability of colonic
smooth muscle was preserved in JF1 mice. The above findings agree well with the data in Fig. 2 (recovery of
myenteric neurons after dDPSC transplantation).
We also evaluated the characteristics of spontaneous colonic contraction. The second part of the proximal
colon exhibited irregular, high-frequency, small-amplitude, transient contractions in JF1 mice, which contrasted
with the prolonged phasic contractions occurring at intervals of several minutes in B6 mice (Fig. 3a–c). dDPSC
transplantation into JF1 mice restored the magnitude and frequency of spontaneous contractions to levels com-
parable to those in B6 mice (Fig. 3a–c). dDPSC transplantation also restored spontaneous contraction magnitude
in the first part of the proximal colon (Extended Data Fig. 5g).
Spatiotemporally-coordinated electrical activity underlies the complex movements of the gastrointestinal
tract24. We used the dialysis membrane-reinforced microelectrode array (MEA) technique to monitor field poten-
tials in muscle sheets isolated from the second part of the proximal colon (Extended Data Fig. 6a–c). Samples
from B6 mice displayed basal electrical oscillations with a period of 3.78 ± 0.22 s (n = 10 recording regions, N = 6
animals) in most of the recording area, and occasionally a large potential occurred (Fig. 3d and Extended Data
Fig. 6d). Basal rhythmic oscillations rarely occurred in JF1 mouse colon (observed in 1/10 recording regions,
N = 3), whereas electrical complexes of rapid and slow potentials occurred frequently (Fig. 3e and Extended Data
Fig. 6e). The rapid potentials propagated in limited micro-regions (suggesting poor spatial synchronicity) with
a frequency (1–2 Hz) comparable to that of myoelectric complexes25–27. dDPSC transplantation into JF1 mice
restored the basal rhythmic oscillations, which occurred with an interval (4.12 ± 0.20 s, n = 10, N = 3) comparable
to that in B6 mice (Fig. 3f and Extended Data Fig. 6f). However, the basal oscillation phase varied within the
small recording area, and rapid potentials occurred locally in some samples, suggesting incomplete recovery
of spatially-coordinated motor function. Spectral analysis (0.1–15 Hz) revealed that the electrical potential
magnitude in JF1 mice was smaller than that in B6 mice but restored by dDPSC transplantation (Fig. 3g). The
low-frequency component was smaller in JF1 mice than in B6 mice but comparable between dDPSC-JF1 and B6
mice (Fig. 3h,i), indicating that colons from dDPSC-JF1 and B6 mice exhibited similar regular basal rhythmicity.
The effects of dDPSC transplantation on electrical characteristics are consistent with the restoration of myenteric
neurons and pacemaker interstitial cells (Fig. 2).
dDPSCs improve nutrition and survival. JF1 mice exhibited slower bodyweight gain (from P11W to
P35W) than B6 mice, and intravenous transplantation of dDPSCs (but not BMMSCs or FBs) caused a small but
significant enhancement of bodyweight gain (during P20W to P31W: Fig. 4a and the photo in Extended Data
Fig. 1a). Intraperitoneal injection of dDPSCs did not improve bodyweight gain in JF1 mice (data not shown).
Median survival time (up to P90W) of JF1 mice was shorter than that of B6 mice (77.8 weeks vs. 112.2 weeks,
n = 10, P = 0.004) and prolonged by the transplantation of dDPSCs (90.2 weeks, n = 10, P = 0.039 vs. JF1 mice)
but not BMMSCs (80.0 weeks, n = 10, P = 0.492 vs. JF1 mice) or FBs (73.8 weeks, n = 10, P = 0.481 vs. JF1 mice;
Fig. 4b). dDPSC transplantation into JF1 mice improved colonic transportation (ex vivo imaging of colonic
content after a 24-h fast; Fig. 4c), restored long bone (tibial) mineral density (micro-computed tomography;
Fig. 4d,e), enhanced hepatic glucose storage and improved the small intestinal mucosa (Extended Data Figs. 1c
and 7) in P23W. These results indicate that multipotent dDPSCs can partially correct entero-neuropathy in JF1
mice (Fig. 4f), probably due to both their multi-lineage differentiation and paracrine effects.
Finally, we investigated potential off-target effects of dDPSC transplantation. dDPSC-JF1 mice exhibited
no morphological alterations or T-cell accumulation in the lungs, liver or kidneys (Extended Data Fig. 8a) and
no changes in the serum levels of enzymes related to the function of these organs (Extended Data Fig. 8b) or
cytokines related to systemic inflammation (Extended Data Fig. 8c).
Discussion
Animal models of spinal cord transection and autoimmune encephalitis have been treated with dDPSCs28,29,
because these stem cells are known to differentiate into neural crest-derived cells, such as functional neurons
and glial cells under appropriate conditions30–32. The treatment of congenital entero-neuropathies is more chal-
lenging because the complex cytoarchitecture of the gastrointestinal tract develops from three germ layers. Our
study shows that dDPSCs have potential for development into a therapy for entero-neuropathies due to their
multipotent characteristics. Intravenous administration of dDPSCs improved the survival and nutritional sta-
tus (bodyweight and glucose uptake) of JF1 mice. We speculate that the latter effect was partly due to repair of
the mucosa (endoderm) by epithelial cells generated by dDPSCs. Furthermore, staining for hMt indicated that
dDPSCs differentiated into pacemaker interstitial cells (mesoderm) and enteric neurons (ectoderm). The ability
of dDPSCs to undergo multi-lineage differentiation is critical for their therapeutic potential because numerous
cells and mechanisms underlie coordinated gut m otility10 and because congenital entero-neuropathies likely
involve not only enteric neuron loss but also other impairments, as seen in JF1 mice. In addition, our data sug-
gest that paracrine mechanisms contribute to the therapeutic effects of dDPSC transplantation, as seen in the
treatment of the central nervous system28,29. In this study, although the body weight was temporarily improved
(P20W to P31W) and the survival ratio was significantly prolonged, it was unverified whether dDPSCs differ-
entiated into functional neurons and pacemaker cells, and how long they exist in recipients. In future study, to
Scientific Reports | (2022) 12:6990 | https://doi.org/10.1038/s41598-022-10077-3 5
Vol.:(0123456789)www.nature.com/scientificreports/
Figure 3. Effects of dDPSCs on colonic function. (a) Spontaneous contractions recorded from proximal
colon muscle strips. (b,c) Slow wave frequency (mean ± SEM, n = 5) and time-tension integral (area under the
curve) for the spontaneous contractions (mean ± SEM; B6, n = 6; JF1, n = 14; dDPSC-JF1, n = 7). (d–f) Electrical
potentials recorded from the proximal colon with an 8 × 8 MEA and the corresponding reconstructed potential
maps. (g–i) Characteristics of the electrical potentials recorded with the 8 × 8 MEA: potential magnitude,
proportion of high-frequency (0.4–10.5 Hz) potentials, and proportion of low-frequency (0.2–0.4 Hz) potentials
(mean ± SEM, n = 10). (a,d–f) Representative of ≥ 2 independent experiments.
Scientific Reports | (2022) 12:6990 | https://doi.org/10.1038/s41598-022-10077-3 6
Vol:.(1234567890)www.nature.com/scientificreports/
Figure 4. Therapeutic effects of dDPSCs. (a) Mouse bodyweight changes after the transplantation of stem cells
into JF1 (mean ± SEM, n = 10) and B6 (mean ± SEM, n = 5) mice. (b) Kaplan–Meier survival curves (n = 10).
Comparisons (vs. JF1 mice) were made using the log-rank test. (c) Autofluorescence of the colonic contents after
a 24-h fast. Representative of ≥ 2 independent experiments. (d) Micro-CT of the tibia. (e) Tibial mineral density
(mean ± SEM, n = 4). (f) Schematic diagram showing the possible mechanisms underlying dDPSC migration,
differentiation and beneficial effects on colonic motility in JF1 mice.
optimize the cell therapy of dDPSCs, these issues need to be addressed in relation with their paracrine effects
and booster injections.
According to our observation, dDPSCs may have advantages over BMMSCs, FBs and other stem cell types
in the treatment of refractory bowel disorders. Undifferentiated MSCs such as dDPSCs express high levels of
inflammation-activated adhesion proteins that enables them to avoid mechanical trapping in healthy capillaries
and settle in regions affected by inflammation33. The lack of intestinal accumulation of dDPSCs in B6 and anti-
SDF1α antibody-treated JF1 mice demonstrates the crucial role of SDF1α-CXCR4 interactions in the migration of
dDPSCs to affected bowel regions. However, CXCR4-expressing BMMSCs did not colonize the myenteric region
or improve the nutritional status of JF1 mice, indicating that mechanisms other than SDF1α-CXCR4 interactions
Scientific Reports | (2022) 12:6990 | https://doi.org/10.1038/s41598-022-10077-3 7
Vol.:(0123456789)www.nature.com/scientificreports/
contribute to the migration and colonization processes. For example, dDPSCs express KIT whereas BMMSCs
do not, and it has been reported that SCF (a KIT ligand released from enteric neurons and smooth muscle cells)
induces the differentiation of MSC-derived progenitor cells into I CCs34. It is likely that transplanted dDPSCs
differentiate into progenitors of enteric neurons and pacemaker cells, which act synergistically to correct bowel
dysmotility through interactions between SCF and KIT and the release of neurotransmitters and hormones that
promote the development of I CCs34.
Various cell types have been evaluated for possible use as a cell-based therapy for HSCR and its allied disor-
ders. Enteric NCC-derived progenitors from mice successfully develop into functional neurons when grafted
into the colon of wild-type recipient mice with the same background35. Moreover, pluripotent stem cells such as
embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs) can regenerate gut-like organoids that
exhibit neural responses and pacemaker potentials36,37. Recently, an ESC-derived NCC lineage was shown to
engraft successfully and reduce disease-related mortality in an animal model of HSCR (mice with an Ednrb muta-
tion)38. As with dDPSCs, the therapeutic effect of NCCs derived from pluripotent stem cells may be enhanced
by the simultaneous development of pacemaker cell lineages. However, the clinical application of ESC- and
iPSC-derived lineages is currently controversial due to possible carcinogenic and immunogenic e ffects39,40, which
highlights the low immunogenicity of dDPSCs as an important advantage (Extended Data Fig. 3). We observed
hMt-containing neurons and pacemaker cells in the colon of JF1 mice 12 weeks after transplantation as well as an
improvement in both mechanical and electrical function (Figs. 2 and 3). Moreover, the high expression of phos-
phatase and tensin homolog (PTEN) is thought to reduce the carcinogenicity of dental pulp stem c ells41. Although
autologous transplantation would be preferable for the clinical application of dDPSCs, the low immunogenicity of
these cells would likely allow allogeneic transplantation of dDPSCs obtained from deciduous or extracted teeth.
The colon exhibits characteristic patterns of electrical potentials. Slowly oscillating potentials (occurring at
intervals of 3–4 s) are prominent in the proximal colon of wild-type mice. KIT-positive pacemaker cells are the
otentials22,23,42. Spontaneous myoelectric complexes comprising rapid and slow components
likely origin of slow p
occur in the mid-to-distal colon, and this electrical activity is considered neurogenic in origin and the basis
of propulsion27. Activity resembling myoelectric complexes is observed in JF1 mouse proximal colon despite
the poor spatial synchronicity, suggesting that loss of spontaneous rhythmicity generated by network-forming
pacemaker cells is compensated for by the relatively preserved enteric neurons. We propose that both the ENS
and network-forming pacemaker system are impaired in entero-neuropathies. This hypothesis would account
for some of the observed discrepancies in colonic motility, such as bi-directional propagation of colonic migrat-
ing motor complexes26.
Heterozygous and homozygous sl mutations of Ednrb cause similar reductions in the myenteric ganglia of
mice, but the homozygous sl mutation is lethal due to a complete absence of m otility43, which would suggest a
role for factors other than ENS impairment. Mutations of Ret are known to be associated with defects in enteric
neurons, and homozygous mutations of Ret also yield HSCR-like mice, whereas the heterozygous mutation is
asymptomatic. Recent studies of the Ret-null mouse embryo suggest that Ret has much broader effects than
enteric neurogenesis44, as we suggest for the Ednrb mutation. We speculate that symptoms of bowel dysmotility
in animal models of congenital entero-neuropathies arise from the additional impairment of the pacemaker
system, because coordinated gastrointestinal movement is maintained by a multitude of cooperating systems.
This may also be true in humans, particularly given the discrepancy between bowel motility symptoms and
morphological defects in the ENS.
In conclusion, the pacemaker and intrinsic nervous systems of the proximal colon are impaired in JF1 mice
with Ednrb mutation. Therefore, appropriate cell-based therapy seems to require stem cells with multi-lineage
potential as well as their paracrine effects in this model of gut motility disorder. Notably, transplanted dDPSCs
migrate to affected regions of the colon and improve the electrical and mechanical activity, likely because they dif-
ferentiate into pacemaker interstitial cells and enteric neurons. Further investigations are required to optimise the
transplantation technique, for example by improving cell migration to affected intestinal regions and promoting
cell differentiation to repair defective tissues. Nevertheless, we consider dDPSC transplantation as a promising
cell-based therapy for congenital entero-neuropathies, especially for patients with pacemaker dysfunction in
apparently ‘healthy’ intestinal segments.
Methods
Ethics statement. All methods were performed in accordance with relevant guidelines and regulations
for humans and animals. All authors agreed that each is responsible for the accuracy and fairness of the related
research.
Healthy human deciduous teeth were collected as discarded biological/clinical samples from three healthy
donors (5–7 years old) who attended the Department of Pediatric Dentistry of Kyushu University Hospital.
The three samples were randomized to use in each experiment. The parents/guardians of the donors provided
written informed consent that stem cells harvested from the teeth could be used for research into regenerative
medicine for congenital diseases. Human peripheral blood mononuclear cells (PBMNCs) were collected from
peripheral venous blood obtained from donors (25–28 years old) unrelated to those who provided deciduous
teeth. Informed consent was obtained from all subjects for use of Human PBMNCs. Procedures for handling
human samples were approved by Kyushu University Institutional Review Board for Human Genome/Gene
Research (Protocol Number: 393-00).
All animal experiments were approved by the Institutional Animal Care and Use Committee of Kyushu Uni-
versity (Protocol Number: A21-044-1; A25-086–0) and Nagoya University (Permissions #28316, #29121, #30312,
#31199 and #20430). Stem cell transplantation caused no substantial biological alterations or welfare issues in
the animals (Fig. 4 and Extended Data Fig. 8). The study was carried out in compliance with ARRIVE guidelines.
Scientific Reports | (2022) 12:6990 | https://doi.org/10.1038/s41598-022-10077-3 8
Vol:.(1234567890)www.nature.com/scientificreports/
Animals. Male JF1 mice (National Institute of Genetics, Mishima, Japan) and male and female wild-type
C57BL/6J (B6) mice (Kyudo, Tosu, Japan; Japan SLC, Hamamatsu, Japan) were housed under controlled envi-
ronmental conditions (12-h light/12-h dark cycle) and given free access to sterile drinking water and a standard
MF chow diet (Oriental Yeast, Tokyo, Japan).
Stem cell isolation and culture. Isolation and culture of stem cells from dental pulp tissue were per-
formed using enzyme digestion and colony-forming unit fibroblast (CFU-F) methods14,15,45,46. Remnant dental
pulp tissues from extracted human deciduous teeth were digested with 0.3% collagenase type I (Worthington
Biochemicals, Lakewood, NJ, USA) and 0.4% dispase II (Sanko Junyaku Co., Ltd., Tokyo, Japan) for 60 min at
37 °C. The resultant cells were passed through a 70-µm cell strainer and seeded on T-75 culture flasks. Three
hours after cell seeding, the culture flasks were washed with sterilized Ca2+-free and M
g2+-free phosphate-buff-
ered saline (PBS). The remaining adherent cells were incubated in Minimum Essential Medium Eagle-Alpha
Modification (αMEM; Thermo Fisher Scientific, Waltham, MA, USA) containing 15% foetal bovine serum (FBS;
Equitech-Bio, Kerrville, TX, USA), 100 µM l-ascorbic acid 2-phosphate (Wako Pure Chemicals, Osaka, Japan),
2 mM l-glutamine (Nacalai Tesque, Kyoto, Japan) and premixed 100 U/mL penicillin/100 µg/mL streptomycin
(Nacalai Tesque) for 16 days. The resulting colonies were passaged to expand the number of dDPSCs. Cells at P3
were used for the experiments.
BMMSCs isolated from human whole bone marrow aspirates obtained from healthy adult volunteers (n = 3;
AllCells, Alameda, CA, USA) and human skin fibroblasts (FBs; PromoCell, Heidelberg, Germany) were prepared
reviously15.
as described p
Stem cell characterization. dDPSCs were characterized according to standard p rotocols47. The adherent
colony-forming capacity was evaluated using a CFU-F assay. Isolated cells (1 × 104) were seeded on T-75 culture
flasks and cultured in growth medium for 16 days. The flasks were treated with 4% paraformaldehyde and 0.1%
toluidine blue in PBS (pH 7.4) for 18 h. The number of fibroblast colonies containing > 50 cells was counted
under a microscope.
The multipotency of dDPSCs (P3) was verified according to previous s tudies12,15. Briefly, specific culture
conditions were used to induce dDPSCs to differentiate into osteoblasts, chondrocytes or adipocytes that were
stained with alizarin red-S, toluidine blue or oil red-O, respectively. Expressions of genes specific to osteoblasts
(runt-related transcription factor-2 [RUNX2] and osteocalcin [BGLAP]), chondrocytes (SRY-box-9 [SOX9] and
aggrecan [ACAN]) and adipocytes (peroxisome proliferator-activated receptor gamma-2 [PPARG] and lipopro-
tein lipase [LPL]) were analysed by quantitative reverse transcription polymerase chain reaction (RT-qPCR).
Expressions of genes specific for neural crest cells, including nerve growth factor receptor (NGFR), paired box-3
(PAX3), zinc finger protein SNAI-2 (SLUG), zinc finger protein SNAI-1 (SNAIL), SOX9 and endothelin recep-
tor type B (EDNRB), in dDPSCs (P3) were examined by RT-qPCR. Expressions of CXCR4 and KIT in dDPSCs
(P3) were assessed using immunofluorescence experiments (see Supplementary Table 1). We have previously
confirmed that cultured dDPSCs express stem cell markers (positive for CD146, CD73, CD105, and CD90, but
negative for hematopoietic markers of CD34, CD45, CD14, and CD11b) and low levels of class II HLA and T
cell costimulatory molecules46.
For mixed lymphocyte assays, human PBMNCs (1 × 106) were co-cultured with dDPSCs (1 × 105) and gamma-
irradiated human PBMNCs (1 × 105; 30 Gy irradiation with an MBR-1520R-3 irradiator; Hitachi, Tokyo, Japan)
or stimulated with phytohemagglutinin (5 μg/mL; Merck, Darmstadt, Germany) in RPMI-1640 medium (Sigma-
Aldrich) containing 10% heat-inactivated FBS (Equitech-Bio), 2 mM l-glutamine, 1 mM sodium pyruvate and
premixed 100 U/mL penicillin/100 µg/mL streptomycin (Nacalai Tesque). Five days after co-culture or stimula-
tion, cell viability was assayed using Cell Counting Kit-8 (Dojindo, Kumamoto, Japan) and a Multiscan GO plate
reader (Thermo Fisher Scientific).
Stem cell transplantation. dDPSCs, BMMSCs or FBs (104 cells/g bodyweight in 100 µL PBS) were
intravenously injected into JF1 mice at P11W, by reference to previous transplantation procedures in wild-type
mice15. Age-matched B6 and JF1 mice intravenously injected with PBS (100 µL) were used as controls. Some
JF1 mice were injected intraperitoneally with anti-SDF1α antibody (100 µg in 100 µL PBS; R&D Systems, Min-
neapolis, MN, USA) or PBS (100 µL) 24 h before transplantation.
Mice were divided into five experimental groups: B6 (n = 10), (PBS-infused) JF1 (n = 10), dDPSC-transplanted
JF1 (n = 10), BMMSC-transplanted JF1 (n = 10) and FB-transplanted JF1 (n = 10). Mouse bodyweight, water
intake, food intake and stool number were measured weekly from P11W to P35W. Some mice were selected
randomly for harvesting of organ tissues and peripheral blood at P23W (12 weeks after transplantation), and the
intestine was dissected into lengths of ~ 15 mm. Mice were sacrificed by cervical dislocation after deeply anesthe-
tized with inhaled isoflurane (Kyushu University), or were euthanized by cervical dislocation after respiratory
arrest with inhaled diethyl ether/carbon dioxide (Nagoya University).
Monitoring of transplanted cells. To reduce the background autofluorescence of standard chow, mice
were fed with alfalfa-free Ivid#2 feed (Oriental Yeast) for 10 days before in vivo and ex vivo imaging. dDP-
SCs were incubated with DiR (Perkin Elmer, Waltham, MA, USA) for 30 min at 37 °C and washed twice with
PBS. Fluorescent dDPSCs (105 cells/g bodyweight in 100 µL PBS) were intravenously transplanted into recipient
mice (P23W). PBS-infused mice were used as controls. Whole-body images and ex vivo images of organs were
acquired 1, 7 and 20 days after transplantation using an IVIS Lumina III system (Perkin Elmer). In the experi-
ment involving the injection of anti-SDF1α antibody into a JF1 mouse, ex vivo imaging was performed 18 days
Scientific Reports | (2022) 12:6990 | https://doi.org/10.1038/s41598-022-10077-3 9
Vol.:(0123456789)www.nature.com/scientificreports/
after dDPSC transplantation due to early death of the mouse. Colour images showing DiR fluorescence above a
threshold level were superimposed on greyscale images of the mouse body or resected organ.
Peristalsis. Mice were fed with standard chow, which exhibits autofluorescence. The entire gastrointestinal
tract was resected (P23W) after a 24-h fast. The autofluorescence of the intestinal contents was measured from
ex vivo images acquired with an IVIS Lumina III system (Perkin Elmer). A reduction in autofluorescence level
was interpreted as an increase in peristaltic movements.
Nutritional status. Improvements in nutritional status (hepatic glucose uptake and body weight) after
dDPSC transplantation were used as surrogate markers of mucosal repair in the small intestine. Tibial bones
were harvested from each experimental group at P23W, and the distal tibial epiphyses were imaged using a Sky-
scan 1076 micro-CT system (Skyscan, Kontich, Belgium). Bone mineral density (BMD) and bone parameters
were estimated using CT-Analyzer software (Skyscan). BMD values were calibrated using hydroxyapatite phan-
toms with BMD values of 0.25 and 0.75 g/cm3 (Skyscan).
In vivo hepatic glucose uptake was measured in mice (P23W) given free access to water containing 1%
2-deoxy-D-glucose (2DG; Merck)48. Liver tissue samples were collected after 7 days. Hepatic 2DG content (an
index of glucose storage) was assayed using a commercial kit (Cosmo Bio, Otaru, Japan) and plate reader (Mul-
tiscan GO; Thermo Fisher Scientific).
Histology and immunohistochemistry. Samples were fixed with 4% paraformaldehyde in PBS (pH 7.2)
at 4 °C overnight, dehydrated, cleaned and embedded in paraffin. For histology, sections were stained with hae-
matoxylin–eosin. For immunohistochemistry, sections were incubated first with 3% H 2O2 in ethanol for 30 min
to inhibit endogenous peroxidase and then with 10% bovine serum albumin in PBS for 60 min. Sections were
treated with 0.01 M citrate buffer (pH 6.0 or 9.0) in a microwave processor (MI-77, Azumaya, Tokyo, Japan) for
antigen retrieval. Sections were incubated with primary antibodies (Supplementary Table 2) at 4 °C overnight.
Visualisation was achieved using an EnVision + System (Agilent, Santa Clara, CA, USA). Sections were lightly
counterstained with haematoxylin. For double-immunofluorescence staining, sections were incubated with pri-
mary antibodies (Supplementary Table S2) and then treated with the Opal 3-plex kit (Perkin Elmer). Sections
were mounted onto slides using Vectashield Mounting Medium with DAPI (Vector Laboratories, Burlingame,
CA, USA). Negative controls utilized isotype-matched antibodies instead of primary antibodies. Images were
obtained using a BZ-X700 fluorescence microscope (Keyence, Tokyo, Japan). The number and area of ganglion
cells in myenteric ganglia were estimated using Hybrid Cell Count software (Keyence)49–51.
Quantitative assays. Blood samples were incubated overnight at 4 °C and centrifuged at 1,500 rpm for
10 min at 4 °C to obtain serum. Tissue samples (including liver, lung and kidney) were homogenized in T-PER
Tissue Protein Extraction Reagent (Thermo Fisher Scientific) containing a proteinase inhibitor cocktail (Nacalai
Tesque) using a Vibra-Cell ultrasonic processor (Sonic & Materials, Newtown, CT, USA). Total protein was
measured using a protein assay (Bio-Rad Laboratories, Hercules, CA, USA) and normalized to the tissue sample
weight.
Commercial kits were used to measure serum levels of creatinine (Creatinine Colorimetric Assay kit, Cayman
Chemical Company, Ann Arbor, MI, USA), alanine aminotransferase and aspartate aminotransferase (Transami-
nase CII-Test kit, Wako Pure Chemicals). Commercial enzyme-linked immunosorbent assay (ELISA) kits were
used to determine albumin, GDNF, IL-2, MIP-2, NGF, SCF and SDF1α protein levels in mouse tissues or serum
(see Supplementary Table 3). The colourimetric assay and ELISA results were analysed using a Multiscan GO
plate reader (Thermo Fisher Scientific).
Serum levels of IL-1α, IL-1β, IL-2, IL-5, IL-6, IL-12, IL-18, granulocyte-colony stimulating factor, tumour
necrosis factor-α, interferon-γ and macrophage inflammatory protein-1β (MIP1β) were quantified using a Bio-
Plex MAGPIX system and a Bio-Plex Pro multiplex cytokine assay system (Bio-Rad Laboratories).
RT‑qPCR. Samples were treated with TRIzol (Thermo Fisher Scientific). RNA extracts were digested with
DNase I (Promega, Fitchburg, WI, USA) and purified using the RNeasy Mini Kit (Qiagen, Venlo, Netherlands).
Total RNA samples were treated with a Revertra Ace qPCR kit (Toyobo, Osaka, Japan) to obtain cDNA. cDNA
samples were mixed with EagleTaq Universal Master Mix (Roche, Basel, Switzerland) and target-specific TaqMan
probes (Thermo Fisher Scientific; Supplementary Tables 4 and 5). RT-qPCR was performed using a Light Cycler
96 system (Roche) under the following conditions: 50 °C for 120 s; 95 °C for 600 s; 45 cycles of 95 °C for 15 s and
60 °C for 60 s. Glyceraldehyde 3-phosphate dehydrogenase mRNA (mouse samples) and 18S ribosomal RNA
(human samples) were used for data normalization.
Mouse intestinal microbiota. Stools were collected daily during P24W–P26W. Faecal bacterial DNA
content was analysed by the Primary Cell Division of Cosmo Bio (Sapporo, Japan; protocol #151228A1 and
#160125A1). Briefly, microbial DNA was extracted from the faecal sample using the Isospin Fecal DNA kit
(Nippon Gene, Tokyo, Japan). The V3V4 region of the 16S rRNA gene was amplified (Extended Data Fig. 2a,b:
Repertoire Genesis, Ibaraki, Japan) by PCR (forward: CCTACGGGNGGCWGCAG; reverse: GACTACHVGGG
TATCTAATCC) and sequenced using a MiSeq Deep sequencer and MiSeq Reagent Kit v3 (Illumina, San Diego,
CA, USA). The sequence data were pre-processed and analysed using Flora Genesis software (Extended Data
Fig. 2c: Repertoire Genesis, Ibaraki, Japan). The R1 and R2 read pairs were joined, and chimera sequences were
removed. Open-reference operational taxonomic unit (OTU) picking was performed using the 97% ID pre-
Scientific Reports | (2022) 12:6990 | https://doi.org/10.1038/s41598-022-10077-3 10
Vol:.(1234567890)www.nature.com/scientificreports/
filtered Greengenes database and Uclust. Representative sequences of each OTU were chosen, and taxonomy
assignment was performed using the Ribosomal Database Project (RDP) classifier and a threshold score ≥ 0.5.
OTUs were grouped if their annotation was the same regardless of RDP score. Bacterial floral communities were
characterised using the Gini-Simpson index (1/D)52,53.
Colonic mechanical activity. Mechanical responses to pharmacological agents and EFS were measured in
15-mm-long muscle strips isolated from the proximal colon. Macroscopically, the mouse proximal colon has two
bulging regions; here we refer to the oral one as the first part and the anal one as the second part. The second part
of the proximal colon was used in all experiments except those shown in Extended Data Fig. 5g. The modified
Krebs solution consisted of 121.6 mM NaCl, 4.7 mM KCl, 2.5 mM CaCl2, 1.2 mM MgCl2, 1.2 mM Na2KH2PO4,
15.4 mM NaHCO3 and 11.5 mM D-glucose gassed with 5% CO2 and 95% O2 (pH 7.4 at 37 °C). The strips were
mounted vertically in an organ bath maintained at 37 °C. Changes in isometric tension generated by the cir-
cular muscle were recorded using a force transducer (TB-612 T; Nihon Kohden, Tokyo, Japan) and amplifier
(AS1202; NEC, Tokyo, Japan) and analysed using LabChart and Scope software (ADInstruments, Bella Vista,
NSW, Australia). Enteric nerve stimulation was achieved using EFS (50 V, 0.5-ms step pulses repeated at 5 Hz
for 5 s) applied by an electrical stimulator (SEN-3301; Nihon Kohden). The following pharmacological agents
were used: 1 µM Ach (Wako Pure Chemicals), 10 nM ET-1 (Wako Pure Chemicals), 10 µM NO (Wako Pure
Chemicals), 60 mM KCl, 1 µM atropine sulphate (Wako Pure Chemicals; to block the response to Ach) and 1 µM
tetrodotoxin (Wako Pure Chemicals; to block the response to EFS). Tension development in response to EFS and
pharmacological agents was normalized to the tissue weight and response to 60 mM KCl.
Colonic electrical activity. The spatiotemporal characteristics of proximal colon electrical activity were
assessed using an 8 × 8 MEA recording system (Alpha MED Scientific, Ibaraki, Japan)54,55. Colonic smooth mus-
cle samples were prepared by gently removing the mucosa. Each sample was mounted on an 8 × 8 MEA (inter-
polar distance, 150 μm) with the longitudinal muscle layer facing downward using a piece of dialysis membrane
fixed with a slice anchor (SDH series, Harvard Apparatus Japan, Tokyo, Japan; Extended Data Fig. 6). The 2-mL
recording chamber was perfused (1–2 mL/min at 34 °C) with modified Krebs solution containing HEPES/Tris
pH-buffer instead of bicarbonate to prevent the development of microbubbles on the MEA. The sample was
perfused with solution for ~ 30 min before recording was started, and perfusion was discontinued during record-
ing to minimize electrical noise. A set of 8 × 8 field potentials were simultaneously recorded using a computer-
controlled, multi-channel AC amplifier (low-pass filtering at 10 kHz) and 14-bit A/D converters (sampling rate
of 20 kHz). The dynamic range of A/D conversion was usually ± 1 mV. High-pass (0.1 Hz) filtering was applied
to stabilize the baseline drift of the microelectrode potential.
A low-impedance MEA was employed to measure a wide range of electrical activity, including pacemaker
potentials occurring at ~ 4-s intervals and myoelectric complex-like potentials with rapid and slow c omponents54.
Each recording electrode was a square (~ 50 μm × 50 μm) made from platinum black nanoparticles, which
increased the surface area by ~ 200-fold to 0.5 m m2. The capacitance (CME) and resistance (RME) of each
microelectrode was 0.052 μF and 15 kΩ, respectively54,56, hence the impedance of the recording electrode at
0.1 Hz was estimated to be small enough [~ 31 MΩ = √ {1/(2π × 0.1 Hz × 0.052 μF)2 + (15 kΩ)2}] to follow oscil-
lating potentials when compared to the input impedance of the multi-channel amplifier (100 MΩ at 0.1 Hz). The
efficacy of electrical signal transmission (Tr) was estimated to be ~ 95% at 0.1 Hz [100 MΩ / √ {(100 MΩ)2 + (31
MΩ)2}]. Moreover, the low resistance (15 kΩ) of the electrodes was advantageous for reducing thermal noise
(NT), which was calculated to be ~ 1.6 μV at the low-pass frequency (10 kHz) of the AC amplifier.
To generate pseudocolour images, the field potential data were thinned 100–1000-fold in the time domain
and bandpass filtered at 0.25–30 Hz (Kaiseki Excel add-in software, Kyowa, Tokyo, Japan). At certain recording
times, the 8 × 8 field potentials were interpolated by a spline function with 50 points between each potential
using MATLAB (MathWorks, Natick, MA, USA)53. To reconstitute smooth potential images, the amplitude of
the field potential in each ME was compensated by the linear spectrum at 0.1–15 Hz54. The upper and lower
images corresponded to the oral and anal ends of the MEA recording region, respectively. Field potential videos
were produced at a frame rate of 200 Hz.
Statistical analysis. Data are expressed as the mean ± standard error of the mean (SEM) of at least three
determinations. Inter-group comparisons were made using two-tailed Student’s t-tests for distinct samples (two
groups), Mann–Whitney U-test or one-way repeated-measures analysis of variance (ANOVA) followed by the
Tukey post-hoc test (three or more groups). Kaplan–Meier analyses and log-rank tests were used for survival
analyses. P < 0.05 was considered significant. Analyses were performed using JMP 11 (SAS Institute, Cary, NC,
USA).
Data availability
The data that support the findings of this study are available from the corresponding author upon reasonable
request. Source data for figures are provided within this paper.
Received: 26 October 2021; Accepted: 22 March 2022
References
1. Burns, A. J. et al. White paper on guidelines concerning enteric nervous system stem cell therapy for enteric neuropathies. Dev.
Biol. 417, 229–251 (2016).
Scientific Reports | (2022) 12:6990 | https://doi.org/10.1038/s41598-022-10077-3 11
Vol.:(0123456789)www.nature.com/scientificreports/
2. Watanabe, Y., Kanamori, Y., Uchida, K. & Taguchi, T. Isolated hypoganglionosis: Results of a nationwide survey in Japan. Pediatr.
Surg. Int. 29, 1127–1130 (2013).
3. Taguchi, T. et al. The incidence and outcome of allied disorders of Hirschsprung’s disease in Japan: Results from a nationwide
survey. Asian J. Surg. 40, 29–34 (2017).
4. Heuckeroth, R. O. Hirschsprung disease: Integrating basic science and clinical medicine to improve outcomes. Nat. Rev. Gastro-
enterol. Hepatol. 15, 152–167 (2018).
5. Di Lorenzo, C., Solzi, G. F., Flores, A. F., Schwankovsky, L. & Hyman, P. E. Colonic motility after surgery for Hirschsprung’s disease.
Am. J. Gastroenterol. 95, 1759–1764 (2000).
6. Hirschsprung, H. Stuhlträgheit neugeborener in folge von dilatation und hypertrophie des colons. Jahrb. Kinderheilkd. Phys. Erzieh.
27, 1–7 (1888).
7. Furness, J. B. The enteric nervous system and neurogastroenterology. Nat. Rev. Gastroenterol. Hepatol. 9, 286–294 (2012).
8. Huizinga, J. D. et al. W/kit gene required for interstitial cells of Cajal and for intestinal pacemaker activity. Nature 373, 347–349
(1995).
9. Nakayama, S. et al. Pacemaker phase shift in the absence of neural activity in guinea-pig stomach: A microelectrode array study.
J. Physiol. 576, 727–738 (2006).
10. Huizinga, J. D. & Lammers, W. J. E. P. Gut peristalsis is governed by a multitude of cooperating mechanisms. Am. J. Physiol. 296,
G1–G8 (2009).
11. Attié, T. et al. Diversity of RET proto-oncogene mutations in familial and sporadic Hirschsprung disease. Hum. Mol. Genet. 4,
1381–1386 (1995).
12. Miura, M. et al. SHED: stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. U. S. A. 100, 5807–5812 (2003).
13. Chai, Y. et al. Fate of the mammalian cranial neural crest during tooth and mandibular morphogenesis. Development 127, 1671–
1679 (2000).
14. Xuan, K. et al. Deciduous autologous tooth stem cells regenerate dental pulp after implantation into injured teeth. Sci. Transl. Med.
10, eaaf3227 (2018).
15. Iwanaka, T. et al. A model study for the manufacture and validation of clinical-grade deciduous dental pulp stem cells for chronic
liver fibrosis treatment. Stem Cell Res. Ther. 11, 134 (2020).
16. Koide, T. et al. A new inbred strain JF1 established from Japanese fancy mouse carrying the classic piebald allele. Mamm. Genome
9, 15–19 (1998).
17. Yamada, T. et al. Reduced expression of the endothelin receptor type B gene in piebald mice caused by insertion of a retroposon-
like element in intron 1. J. Biol. Chem. 281, 10799–10807 (2006).
18. Dang, R., Sasaki, N., Torigoe, D. & Agui, T. Anatomic modifications in the enteric nervous system of JF1 mice with the classic
piebald mutation. J. Vet. Med. Sci. 74, 391–394 (2012).
19. Hosoda, K. et al. Targeted and natural (piebald-lethal) mutations of endothelin-B receptor gene produce megacolon associated
with spotted coat color in mice. Cell 79, 1267–1276 (1994).
20. Pierre, J. F. et al. Intestinal dysbiosis and bacterial enteroinvasion in a murine model of Hirschsprung’s disease. J. Pediatr. Surg. 49,
1242–1251 (2014).
21. Kitaori, T. et al. Stromal cell-derived factor 1/CXCR4 signaling is critical for the recruitment of mesenchymal stem cells to the
fracture site during skeletal repair in a mouse model. Arthritis Rheum. 60, 813–823 (2009).
22. Wedel, T. et al. Enteric nerves and interstitial cells of Cajal are altered in patients with slow-transit constipation and megacolon.
Gastroenterology 123, 1459–1467 (2002).
23. Burns, A. J. Disorders of interstitial cells of Cajal. J. Pediatr. Gastroenterol. Nutr. 45, S103–S106 (2007).
24. Szurszewski, J. H. Electrical basis for gastrointestinal motility. In Physiology of Gastrointestinal Tract 2nd edn (ed. Johnson, L. R.)
383–422 (Raven Press, 1987).
25. Wood, J. D. Electrical activity of the intestine of mice with hereditary megacolon and absence of enteric ganglion cells. Am. J. Dig.
Dis. 18, 477–488 (1973).
26. Sarna, S. K. Myoelectric correlates of colonic motor complexes and contractile activity. Am. J. Physiol. 250, G213–G220 (1986).
27. Spencer, N. J. & Bywater, R. A. Enteric nerve stimulation evokes a premature colonic migrating motor complex in mouse. Neuro-
gastroenterol. Motil. 14, 657–665 (2002).
28. Sakai, K. et al. Human dental pulp-derived stem cells promote locomotor recovery after complete transection of the rat spinal cord
by multiple neuro-regenerative mechanisms. J. Clin. Investig. 122, 80–90 (2012).
29. Shimojima, C. et al. Conditioned medium from the stem cells of human exfoliated deciduous teeth ameliorates experimental
autoimmune encephalomyelitis. J. Immunol. 196, 4164–4171 (2016).
30. Arthur, A. et al. Adult human dental pulp stem cells differentiate toward functionally active neurons under appropriate environ-
mental cues. Stem Cells 26, 1787–1795 (2008).
31. Király, M. et al. Simultaneous PKC and cAMP activation induces differentiation of human dental pulp stem cells into functionally
active neurons. Neurochem. Int. 55, 323–332 (2009).
32. Gazarian, K. G. & Ramírez-García, L. R. Human deciduous teeth stem cells (SHED) display neural crest signature characters. PLoS
ONE 12, e0170321 (2017).
33. Mabuchi, Y. et al. LNGFR(+)THY-1(+)VCAM-1(hi+) cells reveal functionally distinct subpopulations in mesenchymal stem cells.
Stem Cell Rep. 1, 152–165 (2013).
34. Huizinga, J. D. & White, E. J. Progenitor cells of interstitial cells of Cajal: on the road to tissue repair. Gastroenterology 134,
1252–1254 (2008).
35. Hotta, R. et al. Transplanted progenitors generate functional enteric neurons in the postnatal colon. J. Clin. Investig. 123, 1182–1191
(2013).
36. Workman, M. J. et al. Engineered human pluripotent-stem-cell-derived intestinal tissues with a functional enteric nervous system.
Nat. Med. 23, 49–59 (2017).
37. Takaki, M., Nakayama, S., Misawa, H., Nakagawa, T. & Kuniyasu, H. In vitro formation of enteric neural network structure in a
gut-like organ differentiated from mouse embryonic stem cells. Stem Cells 24, 1414–1422 (2006).
38. Fattahi, F. et al. Deriving human ENS lineages for cell therapy and drug discovery in Hirschsprung disease. Nature 531, 105–109
(2016).
39. Zhao, T., Zhang, Z. N., Rong, Z. & Xu, Y. Immunogenicity of induced pluripotent stem cells. Nature 474, 212–215 (2011).
40. Araki, R. et al. Negligible immunogenicity of terminally differentiated cells derived from induced pluripotent or embryonic stem
cells. Nature 494, 100–104 (2013).
41. Shen, W. C. et al. Methylation and PTEN activation in dental pulp mesenchymal stem cells promotes osteogenesis and reduces
oncogenesis. Nat. Commun. 10, 2226 (2019).
42. Lies, B. et al. Nitrergic signalling via interstitial cells of Cajal regulates motor activity in murine colon. J. Physiol. 593, 4589–4601
(2015).
43. Ro, S., Hwang, S. J., Muto, M., Jewett, W. K. & Spencer, N. J. Anatomic modifications in the enteric nervous system of piebald mice
and physiological consequences to colonic motor activity. Am. J. Physiol. 290, G710–G718 (2006).
44. Chatterjee, S., Nandakumar, P., Auer, D. R., Gabriel, S. B. & Chakravarti, A. Gene- and tissue-level interactions in normal gastro-
intestinal development and Hirschsprung disease. Proc. Natl. Acad. Sci. U. S. A. 116, 26697–26708 (2019).
Scientific Reports | (2022) 12:6990 | https://doi.org/10.1038/s41598-022-10077-3 12
Vol:.(1234567890)You can also read

















































