Characteristics of Fetal Extrachromosomal Circular DNA in Maternal Plasma: Methylation Status and Clearance
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Clinical Chemistry 00:0 Molecular Diagnostics and Genetics
1–9 (2021)
Characteristics of Fetal Extrachromosomal Circular
DNA in Maternal Plasma: Methylation Status and
Clearance
Downloaded from https://academic.oup.com/clinchem/advance-article/doi/10.1093/clinchem/hvaa326/6146145 by guest on 24 March 2021
Sarah T.K. Sin,a,b,† Lu Ji,a,b,† Jiaen Deng,a,b,† Peiyong Jiang,a,b Suk Hang Cheng,a,b Macy M.S. Heung,a,b
Caitlyn S.L. Lau,c Tak Y. Leung,c K.C. Allen Chan,a,b Rossa W.K. Chiu,a,b and Y.M. Dennis Loa,b,*
BACKGROUND: Although the characterization of cell-free eccDNA was found to be rapidly cleared from the ma-
extrachromosomal circular DNA (eccDNA) has gained ternal blood after delivery, similar to fetal linear DNA.
much research interest, the methylation status of these
molecules is yet to be elucidated. We set out to compare
the methylation densities of plasma eccDNA of mater-
Introduction
nal and fetal origins, and between small and large mole-
cules. The clearance of fetal eccDNA from maternal cir-
The identification of extrachromosomal circular DNA
culation was also investigated.
(eccDNA) in murine and human blood circulation has
METHODS: We developed a sequencing protocol for
prompted much research interest in the characterization
eccDNA methylation analysis using tagmentation and of this topologic form of DNA molecules (1–3) and
enzymatic conversion approaches. A restriction enzyme- their potential in diagnostics (4). Emerging evidence has
based approach was applied to verify the tagmentation shown that cell-free eccDNA molecules possess several
results. The efficiency of cell-free fetal eccDNA clear- biological properties that are distinct from their linear
ance was investigated by fetal eccDNA fraction evalua- counterparts. For instance, plasma eccDNA molecules
tions at various postpartum time points. displayed bimodal size profiles with 2 major peak clus-
ters, exhibiting 202- and 338-bp summits and sharp 10-
RESULTS: The methylation densities of fetal eccDNA bp periodicities within both clusters (3), whereas the
(median: 56.3%; range: 40.5–67.6%) were lower than majority of plasma linear DNA molecules were found
the maternal eccDNA (median: 66.7%; range: 56.5– shorter than 200 bp with their 10-bp periodicities
75.7%) (P ¼ 0.02, paired t-test). In addition, eccDNA mainly present in small molecules (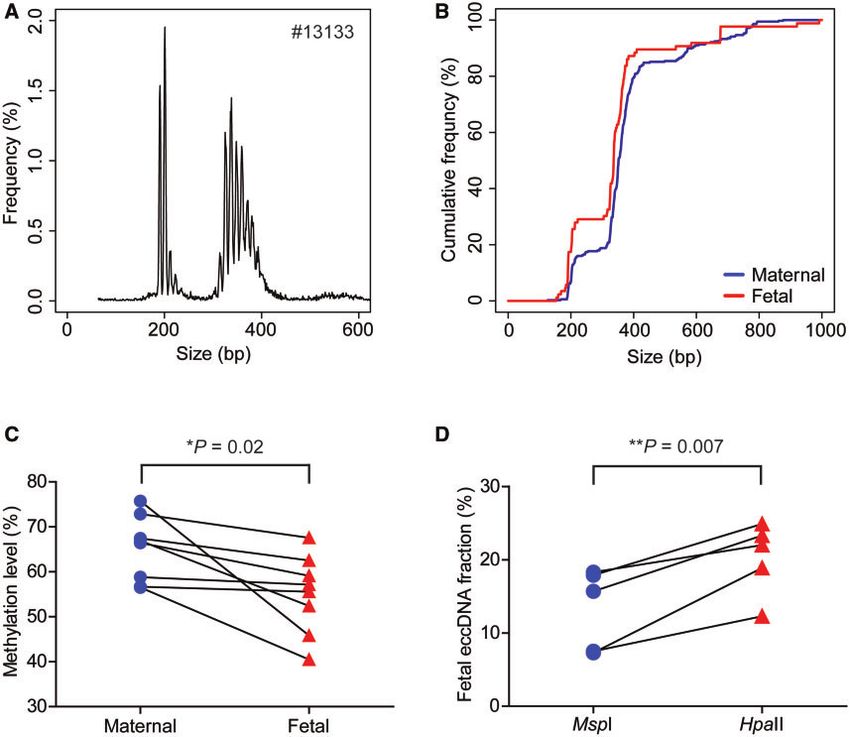
blood samples of pregnant women with relatively were performed at 37 C for 2 hours, stopped at 70 C
smaller size distributions than the maternal population for 10 min with the addition of 2 lL of EZ-Tn5 10
(3). However, the methylation patterns of plasma Reaction Buffer (Lucigen), and column-purified.
eccDNA and their maternal–fetal differences remain As illustrated in Fig. 1, the tagmentation reaction
unknown. In this study, we developed an approach would leave 9-nt gaps and single-stranded jagged ends
utilizing tagmentation and enzymatic cytosine to uracil on the DNA molecules. To fill these gaps and ends, the
Downloaded from https://academic.oup.com/clinchem/advance-article/doi/10.1093/clinchem/hvaa326/6146145 by guest on 24 March 2021
conversion technologies followed by massively parallel Klenow fragment (New England Biolabs) was used for
sequencing in an attempt to explore the methylation gap/end repair. The reaction mixtures (34 lL purified
status of plasma eccDNA. DNA, 5 lL 10 NEBuffer 2, 2.5 lL 10 mmol/L
The distinct size distributions and topological status dATP, 2.5 lL 10 mmol/L dTTP, 2.5 lL 10 mmol/L 5-
between cell-free linear and eccDNA also prompted us to mdCTP, 2.5 lL 10 mmol/L dGTP, 1 lL Klenow frag-
study whether these two forms of DNA molecules are of ment) were incubated at 37 C for 30 min and purified
differential biostability in the blood stream. Previous stud- using columns.
ies demonstrated that fetal linear DNA was cleared rapidly
from the maternal blood stream after delivery with half- ENZYMATIC CYTOSINE TO URACIL CONVERSION AND LIBRARY
lives as short as 16 min (9, 10). In the present study, we CONSTRUCTION
used maternal blood samples at various postnatal time To minimize DNA damage and maximize DNA mapp-
points to determine the efficiency of fetal eccDNA clear- ability, we performed unmethylated cytosine to uracil
ance in maternal plasma. conversion using the NEBNext Enzymatic Methyl-seq
Conversion Module (New England Biolabs) on the 5-
Materials and Methods mC-Tn5 reaction products according to the manufac-
turer’s instructions. Detailed procedures of library prep-
CASE RECRUITMENT, BLOOD SAMPLE COLLECTION, AND arations are described in the Supplemental Materials
PROCESSING and Methods.
This study was approved by the Joint Chinese
University of Hong Kong-Hospital Authority New ECCDNA IDENTIFICATION AND METHYLATION ANALYSIS
Territories East Cluster Clinical Research Ethics Experimental procedures and bioinformatics principles
Committee. Pregnant women attending the antenatal of eccDNA identification using the restriction enzyme
clinic at the Department of Obstetrics and approach, and the annotation of eccDNA to different
Gynaecology, Prince of Wales Hospital, Hong Kong types of genomic regions were performed as previously
SAR, China, as well as nonpregnant female participants, described (3).
were recruited with written informed consent. Blood For the analysis of sequencing data, a modified ver-
samples from pregnant women were collected before sion of the previously described bioinformatics pipeline
delivery [for methylation analysis using the MspI/HpaII (3) was applied. More detailed principles of eccDNA
restriction enzyme and 5-methyl cytosine Tn5 trans- identification are described in the Supplemental
posase (5-mC-Tn5) approaches; third trimester] Materials and Methods. For eccDNA methylation
and at various time points after delivery (for half-life analysis, the sequenced thymines at the genomic CpG
determination by tagmentation approach; 0, 30, 60 sites were assigned as unmethylated CpG sites, while the
and 120 min postpartum). Blood samples were proc- sequenced cytosines at the genomic CpG sites were
essed and plasma DNA was extracted as previously assigned as methylated CpG sites. The methylation den-
described (3). sities of eccDNA were defined as the proportion of
methylated sequenced CpG sites among the total se-
TAGMENTATION AND GAP/END REPAIR quenced CpG sites. The methylation densities of
The construction of 5-mC-Tn5 transposomes was mod- eccDNA at CH sites were measured in a similar
ified from Spektor et al. (11) and is described in more manner.
detail in the Materials and Methods in the online Data
Supplement. For tagmentation experiments using DETERMINATION OF FETAL ECCDNA HALF-LIVES
5-mC-Tn5, 50 ng of plasma DNA was digested with Identification of total, maternal, and fetal eccDNA from
5 units of exonuclease V (New England Biolabs) at maternal plasma was achieved by exonuclease V diges-
37 C for 5 min. The digestion products were then tion and tagmentation approaches as detailed previously
cleaned-up by column purification. The tagmentation (3). The cell-free fetal linear and eccDNA fractions for
reactions were prepared by mixing 17 lL of purified each postpartum time point were determined. The natu-
DNA, 2 lL of EZ-Tn5 10 Reaction Buffer (Lucigen) ral logarithm values of fetal DNA fractions for each time
and 1 lL of 5-mC-Tn5 transposomes. The reactions point were fit with linear regression lines against sample
2 Clinical Chemistry 00:0 (2021)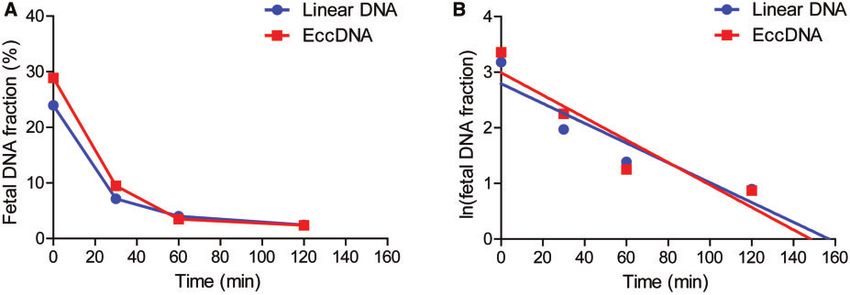
Features of Circulating Fetal eccDNA
collection time points and the slope values were
obtained. Fetal linear and eccDNA half-lives for each
case were calculated using the following formula:
Inð2Þ
half life ¼
Slope
Downloaded from https://academic.oup.com/clinchem/advance-article/doi/10.1093/clinchem/hvaa326/6146145 by guest on 24 March 2021
ILLUMINA SEQUENCING AND STATISTICAL ANALYSIS
DNA libraries for eccDNA methylation and fetal
eccDNA clearance analyses were sequenced on an
Illumina HiSeq 1500 platform as 2 250 bp paired-end
reads. Paired t-tests were performed to compare 2
groups of values of the same sample sets. A P value
lower than 0.05 was considered statistically significant.
Results
EXPERIMENTAL WORKFLOW
The principles and workflow of eccDNA methylation
analysis are illustrated in Fig. 1. The circular forms of
DNA from plasma samples were enriched by digesting
the linear portions with exonuclease V incubation,
which was then followed by tagmentation using 5-mC-
Tn5 transposomes. Such a tagmentation step was a one-
step process that simultaneously opened the DNA
circles and ligated sequencing adaptors: the 2 adaptors
from the 5-mC-Tn5 (highlighted in blue and red) were
added to the two ends of the same eccDNA fragment
(Fig. 1). The gap filling and end repair steps turned the
tagmented DNA fragments into blunt-ended molecules
for sequencing. The adapter-ligated DNA molecules
Fig. 1. Workflow of plasma eccDNA identification and were then subjected to enzymatic conversion by which
methylation analysis. Cell-free DNA comprising of linear the unmethylated cytosines were converted to uracils
and circular forms of molecules were extracted from the that were amplified as thymines, while the methylated
plasma samples of pregnant participants. Exonuclease V cytosines remained unchanged, thus allowing for deter-
treatment would digest most of the linear DNA and signifi-
mination of methylation patterns of each molecule. The
cantly enrich eccDNA from the plasma DNA samples. The
eccDNA molecules were identified from fragments that
enriched eccDNA was then tagmented by 5-mC-Tn5 trans-
were either with read 1 or read 2 unmappable and those
posomes: each 5-mC-Tn5 transposome contained a Tn5
transposase dimer and two outward-oriented sequences
mapped with eccDNA-specific outward alignment ori-
(all C bases were methylated Cs) of the Tn5 transposon entations as described previously (3, 12). The CpG sites
(adaptors) (13). The tagmentation reaction by the 5-mC-Tn5 residing in the eccDNA fragments were used for deter-
transposomes would open the DNA circles and ligate two mining the methylation densities of these molecules.
adaptors (indicated as red and blue bars) to both ends of
the same DNA fragment. This procedure would also gener- METHYLATION ANALYSIS OF CELL-FREE ECCDNA IN
ate 9-nt gaps and jagged ends on the linearized eccDNA MATERNAL PLASMA
fragments, which would be filled by the end/gap repair To compare the methylation densities between maternal
step using Klenow fragment. The completed fragments and fetal eccDNA, we analyzed 8 cases from third tri-
would then undergo enzymatic conversion of unmethy- mester pregnancies using the experimental design illus-
lated cytosine to uracil, leaving the methylated cytosine
trated in Fig. 1. Among these 8 cases, the median
unchanged. The converted DNA molecules were amplified
eccDNA count was 7874 (range: 4340–22 790). Fig. 2,
by PCR and sequenced on an Illumina HiSeq 1500 plat-
A shows the total eccDNA size profile of a representative
form. The sequencing data were processed using our bioin-
formatics pipelines for eccDNA identification and
pregnancy case (#13133). The size profiles of eccDNA
methylation level determination. of all 8 cases are shown in Supplemental Fig. 1. Two
predominant peak clusters with their summits at 202
Clinical Chemistry 00:0 (2021) 3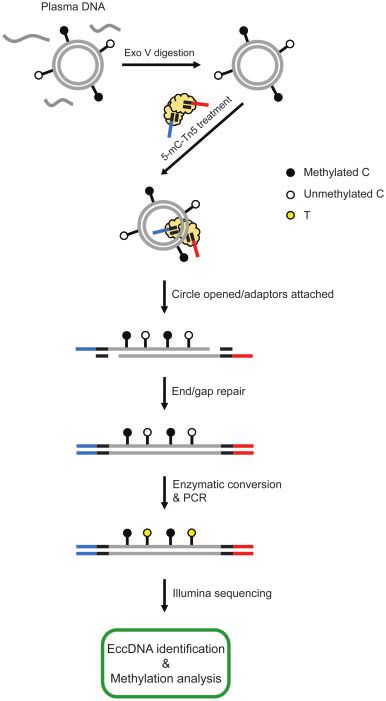
Downloaded from https://academic.oup.com/clinchem/advance-article/doi/10.1093/clinchem/hvaa326/6146145 by guest on 24 March 2021
Fig. 2. EccDNA size profiling and methylation analysis. (A), Size profiling of total eccDNA of representative case #13133; (B), cu-
mulative frequency plot of maternal (blue) and fetal (red) eccDNA in plasma (pooled data from 8 cases). Sizes were capped at
1000 bp for both groups of molecules. (C), Methylation densities of maternal and fetal eccDNA in the plasma of third trimester
pregnancy cases (8 cases in total). P ¼ 0.02, paired t-test; (D), fetal eccDNA fractions deduced from MspI- and HpaII-treated
plasma DNA samples of third trimester pregnancy cases (5 cases in total). P ¼ 0.007, paired t-test.
and 338 bp, respectively, and sharp small peaks at 10-bp the CpG methylation densities ranged from 56.1 to
intervals were observed. SNP-based approach was ap- 67.3% with a median of 58.5%. Using the 8 cases we
plied to mine the maternal- and fetal-derived eccDNA examined, the fetal eccDNA methylation densities (me-
from the sequencing data as detailed previously (3). The dian: 56.3%; range: 40.5–67.6%) were lower than those
median fetal eccDNA fraction of these 8 cases was of the maternal eccDNA (median: 66.7%; range: 56.5–
15.1% (range: 11.4–41.4%). Figure 2, B plots the cu- 75.7%) (P ¼ 0.02, paired t-test) (Fig. 2, C). However,
mulative frequencies of maternal- and fetal-specific for 2 of those 8 cases, the maternal–fetal differences in
eccDNA from pooled data of the 8 cases. The fetal- eccDNA methylation levels were modest: 56.7% (ma-
derived eccDNA molecules were shown to be enriched ternal) versus 55.6% (fetal) for case 13142, and 58.8%
for shorter DNA molecules when compared to their ma- (maternal) versus 57.1% (fetal) for case #13151. Our
ternal counterparts. data indicated that fetal eccDNA was generally hypome-
The CpG methylation densities were then deter- thylated in comparison with the maternal molecules.
mined for the total, fetal, and maternal eccDNA mole- It has previously been reported that eccDNA was
cules for each case. For the total eccDNA populations, relatively enriched in genic regions (3, 14). To study
4 Clinical Chemistry 00:0 (2021)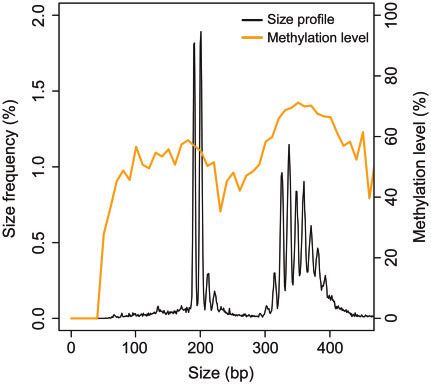
Features of Circulating Fetal eccDNA
whether eccDNA methylation correlates with their ge- molecules to 5 types of genomic element: 50 UTR, 30
nomic origins, we compared the CpG methylation levels UTR, exon, intron, and Alu. Again, we observed similar
of eccDNA between genic and intergenic origins levels of CpG methylation between the linear and
(P ¼ 0.064, paired t-test), as well as between origins of eccDNA populations in these genomic regions (Fig. 3,
high and low gene density regions (P ¼ 0.21; paired t- B).
test) (Supplemental Fig. 2). No statistically significant
Downloaded from https://academic.oup.com/clinchem/advance-article/doi/10.1093/clinchem/hvaa326/6146145 by guest on 24 March 2021
difference was detected for either of these comparisons. ASSOCIATIONS BETWEEN ECCDNA SIZE AND METHYLATION
In addition to the methylation status of eccDNA at DENSITIES
CpG dinucleotides, we also examined the non-CG Figure 4 plots the correlation between eccDNA size and
methylation levels of eccDNA. In particular, we com- methylation densities. There was a periodicity of around
pared the CHH and CHG methylation levels between 200 bp in the eccDNA methylation level, which was
maternal and fetal eccDNA (H: A, T, or C). For the 8 reminiscent of previous findings in plasma linear DNA
cases analyzed, no statistically significant difference was (15). In addition to the 200 bp periodicity, we observed
observed between maternal and fetal molecules in either a narrower periodicity in multiples of 10-bp.
CHH (P ¼ 0.71; paired t-test) or CHG (P ¼ 0.68; We also compared the methylation densities of
paired t-test) methylation levels (Supplemental Fig. 3). eccDNA populations from 2 size bands, namely the 2
Further studies of eccDNA with large-scale samples predominant peak clusters of 180–230 bp (peak cluster
might deepen our understanding of their methylation 1) and 300–450 bp (peak cluster 2) (highlighted in yel-
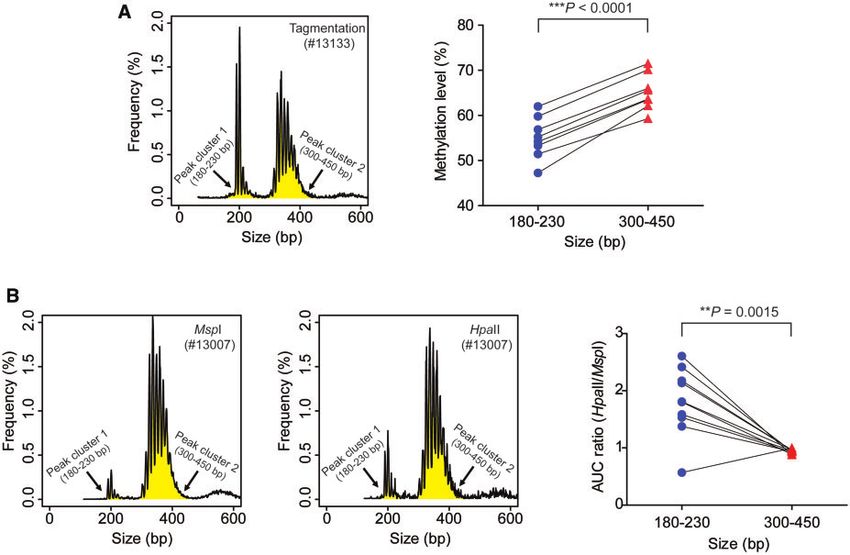
properties across different genomic regions. low for representative case #13133; left panel of Fig. 5,
We further attempted to use the methylation sensi- A). For all the 8 cases, the eccDNA of smaller sizes
tive and insensitive restriction enzymes to independently (peak cluster 1) exhibited lower methylation densities
verify our observation that the fetal eccDNA was hypo- than that of larger sizes (peak cluster 2) (P < 0.0001,
methylated compared to the maternal eccDNA. These paired t-test) (right panel of Fig. 5, A). To further verify
data were obtained from our previously published data- this finding, we applied the MspI/HpaII approach to
set generated by the restriction enzyme approach (3). compare the methylation densities of eccDNA from
Two restriction enzymes, namely MspI (methylation in- peak clusters 1 and 2 in both pregnant (5 cases) and
sensitive) and HpaII (blocked by CpG methylation on nonpregnant participants (5 cases). For eccDNA size
the CCGG recognition site), were chosen for cutting profiles obtained from MspI or HpaII treatment, we cal-
open the DNA circles. The MspI enzyme could poten- culated the areas under the curve (AUC) for both of the
tially open all eccDNA circles harboring the enzyme peak clusters (highlighted in yellow for representative
cutting site regardless of methylation states, while the case #13007 in the left and middle panels of Fig. 5, B).
HpaII enzyme would preferentially cut and thus detect We then calculated the ratios of AUCHpaII: AUCMspI for
the hypomethylated molecules. We compared the fetal the 2 peak clusters. If this AUC ratio is higher in peak
eccDNA fractions obtained from plasma DNA sub- cluster 1 than that in peak cluster 2, it means HpaII
jected to MspI and HpaII cutting. We found that the fe- preferentially cuts eccDNA within peak cluster 1.
tal fractions of eccDNA detected by HpaII treatment According to the data shown in the right panel of Fig. 5,
(median: 22.0%; range: 12.3–24.9%) were significantly B, the AUCHpaII: AUCMspI ratios were significantly
higher than those detected by MspI (median: 15.7%; higher in peak cluster 1 than those in peak cluster 2
range: 7.3–18.3%) (P ¼ 0.007, paired t-test) (Fig. 2, (P ¼ 0.0015, paired t-test). Therefore, the MspI/HpaII
D). This result suggested that HpaII enzyme preferen- approach further confirmed that the smaller eccDNA
tially acted on the fetal eccDNA molecules, supporting molecules (180–230 bp) were more susceptible to HpaII
the aforementioned conclusion that fetal eccDNA was cleavage and thus relatively hypomethylated compared
relatively hypomethylated compared to the maternal with the larger molecules (300–450 bp).
eccDNA population.
CLEARANCE KINETICS OF CIRCULATING FETAL ECCDNA
SIMILAR METHYLATION DENSITIES BETWEEN LINEAR AND To assess the efficiency of fetal eccDNA clearance
ECCDNA IN PLASMA in vivo, we evaluated the fetal eccDNA portions in the
We also sought to answer whether there was any differ- maternal plasma collected at different time points after
ence in terms of CpG methylation between linear and delivery and compared them with linear DNA.
eccDNA molecules in plasma. We did not observe sta- Collections of maternal blood samples were timed start-
tistically significant differences in methylation levels be- ing from the removal of the placenta from the pregnant
tween linear and eccDNA of either maternal (P ¼ 0.46, women and collected at 0, 30, 60, and 120 min postde-
paired t-test) or fetal (P ¼ 0.49, paired t-test) origin livery. Serial time point collections were performed on 3
(Fig. 3, A). We then annotated the linear and eccDNA pregnant individuals. Of note, sample collections at
Clinical Chemistry 00:0 (2021) 5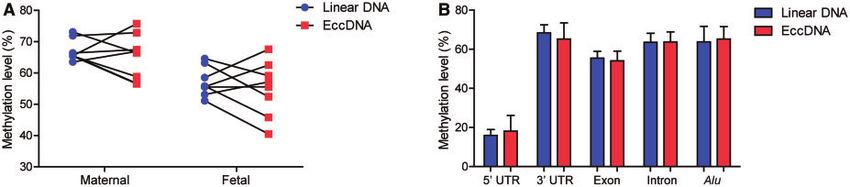
Downloaded from https://academic.oup.com/clinchem/advance-article/doi/10.1093/clinchem/hvaa326/6146145 by guest on 24 March 2021
Fig. 3. Methylation densities of cell-free linear and eccDNA. (A), Methylation densities of linear and eccDNA in the plasma of
third trimester pregnancy cases (8 cases in total). No statistically significant differences in methylation levels between linear and
eccDNA were found for either maternal (P ¼ 0.46, paired t-test) or fetal (P ¼ 0.49, paired t-test) origins; (B), methylation densi-
ties of linear and eccDNA in 5 types of genomic elements (8 cases in total). No statistically significant differences in methylation
levels were found between linear and eccDNA in any of these genomic regions.
which is in line with previous findings (9, 10). The per-
centages of fetal-specific molecules among the total
eccDNA populations also declined rapidly after delivery.
We further fit linear regression lines between the
natural logarithm values of fetal portions (linear and
eccDNA) and collection time points, as shown in
Fig. 6, B of case #13155 and online Supplemental Fig.
4, B of cases #13118 and #13139. The slope values of
the linear regression lines were used to determine the
half-lives of fetal linear and eccDNA in maternal
plasma. For the 3 cases, the fetal linear DNA half-lives
were 22.1, 29.5, and 39.0 min with a mean value of
30.2 min; the fetal eccDNA half-lives of the same cases
were 25.4, 29.3, and 34.4 min, respectively, with a
mean value of 29.7 min. Therefore, fetal linear and
eccDNA had similar half-lives in maternal blood, both
of which were rapidly cleared after delivery.
Fig. 4. Size-methylation correlations of eccDNA in plasma. Discussion
The CpG methylation densities (orange curve) were coplot-
ted with eccDNA size frequencies (black curve) (pooled data
This study investigated the methylation status of the
from 8 cases).
maternal and fetal eccDNA molecules. The tagmenta-
tion approach has been found to be both efficient and
sensitive in eccDNA detection in previous studies (3,
16). Therefore, we leveraged the tagmentation-based
certain time points in 2 cases were missed due to blood technologies for capturing eccDNA molecules and
clotting at the blood collection needles. EccDNA in the obtaining their epigenetic information. Improved DNA
maternal plasma DNA was enriched by the removal of quality could enhance DNA mapping, which would
linear DNA using exonuclease V digestion, followed by benefit high-confidence eccDNA identification that
sequencing using the tagmentation method as previ- requires the precise pinpointing of their junctional sites.
ously described (3). Figure 6, A shows the fetal linear In this regard, we utilized the enzymatic cytosine to ura-
and eccDNA portions at different postpartum time cil conversion in place of traditional bisulfite conversion
points of representative case #13155. Such plots of fetal to minimize DNA degradation and increase DNA
linear and eccDNA portions for the remaining 2 cases mappability (17). Through this method integrating tag-
(#13118 and #13139) are shown in Supplemental Fig. mentation and enzymatic conversion approaches fol-
4, A. For linear plasma DNA, the fetal DNA portions lowed by sequencing, we were able to identify
were rapidly reduced for all the 3 cases after delivery, thousands to tens of thousands of eccDNA molecules
6 Clinical Chemistry 00:0 (2021)Features of Circulating Fetal eccDNA
Downloaded from https://academic.oup.com/clinchem/advance-article/doi/10.1093/clinchem/hvaa326/6146145 by guest on 24 March 2021
Fig. 5. Positive correlations between eccDNA size and methylation densities. (A), Tagmentation-based analysis. The left panel
highlights the two peak clusters (180–230 bp, peak cluster 1; 300–450 bp, peak cluster 2) in yellow for representative case
#13133. The right panel shows a comparison of the eccDNA methylation densities between the two peak clusters for the 8 cases
of third trimester pregnancy. P < 0.0001, paired t-test. (B), Restriction enzyme-based analysis. Plasma eccDNA from both non-
pregnant (5 cases) and pregnant participants (5 cases) was analyzed using MspI and HpaII enzymes. The left and middle panels
highlight the two peak clusters (180–230 bp, peak cluster1; 300–450 bp, peak cluster 2) in yellow color for representative preg-
nancy case #13007 treated with MspI and HpaII, respectively. The AUC values of the two predominant peak clusters of eccDNA
in each case were calculated and plotted as ratios (AUCHpaII: AUCMspI), as shown in the right panel. P ¼ 0.0015, paired t-test.
for each pregnancy plasma sample. We observed the bi- This result hinted that the fetal and maternal eccDNA
modal size patterns (2 peak clusters displaying 202 and would largely preserve the DNA methylation status of
338 bp summits) and the sharp 10-bp periodicity of their genomes of origin. Interestingly, we observed that
eccDNA. These typical size patterns of eccDNA were the eccDNA population of smaller sizes (peak cluster 1
reported in our previous study using tagmentation- of 180–230 bp) tended to be less methylated than the
based nonbisulfite sequencing and restriction enzyme larger molecules (peak cluster 2 of 300–450 bp). Two
digestion followed by sequencing (3). Thus, the se- possible mechanisms might underlie such differences in
quencing protocol combining tagmentation and enzy- methylation levels. The first possible mechanism con-
matic conversion technologies we developed is a reliable cerns eccDNA generation: eccDNA with smaller sizes
method for eccDNA identification. might be preferentially derived from the hypomethy-
By reading into the epigenetic information of lated regions. Previous reports elaborated the close cor-
eccDNA molecules, we found that the overall methyla- relation between nucleosome assembly and methylation
tion level of fetal eccDNA was lower than that of the densities: CpG methylation facilitates (H3-H4)2 tetra-
maternal eccDNA. The lower methylation level present mer incorporation and DNA wrapping. The higher the
in the fetal eccDNA molecules was reminiscent of the methylation level, the higher the nucleosome occupancy
fact that the fetal linear DNA was featured with lower (18). Such correlations might help explain our observa-
methylation levels compared to the maternal DNA (7). tion that the di-nucleosomal eccDNA (the 300–450 bp
Clinical Chemistry 00:0 (2021) 7Downloaded from https://academic.oup.com/clinchem/advance-article/doi/10.1093/clinchem/hvaa326/6146145 by guest on 24 March 2021
Fig. 6. Fetal eccDNA clearance from maternal plasma. (A), Absolute values of fetal linear and eccDNA fractions at various postna-
tal time points of representative case #13155; (B), linear regression fitting between the natural logarithm values of fetal linear
or eccDNA fractions and the postnatal time points of representative case #13155. R2linear DNA ¼ 0.86; Slopelinear DNA ¼ 0.018;
R2eccDNA ¼ 0.859; SlopeeccDNA ¼ 0.02.
peak cluster) exhibited higher methylation levels than in the clearance kinetics of these topologic forms of cir-
the mono-nucleosomal molecules (the 180–230 bp peak culating DNA might suggest that there are clearance
cluster). Second, eccDNA molecules of lower methyla- mechanisms that are shared by both types of DNA.
tion densities might be more susceptible to degradation Cell-free DNA is an important tool for early disease
when exposed to nucleases present in the blood circula- diagnosis. Preeclampsia, which is an important cause for
tion. Previous studies illustrated the correlation between maternal and fetal mortality, has been proposed to be
size and methylation level of plasma linear DNA (7, associated with epigenetic alterations of genes such as
19): larger DNA fragments were of comparatively higher TIMP3 (22). Since eccDNA molecules exhibited larger
methylation levels than the short molecules. Also, it has size distributions compared to their linear counterparts
been reported that DNASE1 activity could be inhibited (3), they might be able to capture higher proportions of
by CpG methylation (20). Therefore, it is reasonable to CpG sites of target genes in diagnostics. Thus, the
speculate that eccDNA of higher methylation levels tagmentation-based sequencing technology for eccDNA
might be more resistant to degradation by certain nucle- methylation analysis might provide the possibility
ases and thus might be better preserved. Further studies for methylation biomarker development of early diagno-
are required to elucidate the relationships between the sis of diseases related to dysregulation of DNA
methylation configuration of different genomic regions methylation.
and eccDNA production. For instance, one could apply
5-azacytidine (a DNA demethylation agent) to an Supplemental Material
in vitro cell culture system to explore whether eccDNA
generation would be enhanced or attenuated. On the Supplemental material is available at Clinical Chemistry
other hand, with deep sequencing coverages, it would online.
be interesting to study eccDNA occurrence in the
imprinted regions, which might imply whether the
methylation patterns would affect eccDNA formation. Nonstandard Abbreviations: eccDNA, extrachromosomal circular
DNA; cfDNA, cell-free DNA.
Interestingly, we found that the overall clearance ki-
netics of cell-free eccDNA generally followed the clear-
ance patterns pertaining to the linear DNA in plasma, as Human Genes: DNASE1, Deoxyribonuclease 1; TIMP3, Tissue
revealed by the results from various postpartum time Inhibitor of Metalloproteinases 3.
points. Two routes of cell-free DNA clearance had been Author Contributions: All authors confirmed they have contributed to
proposed: nuclease digestion and phagocytic elimination the intellectual content of this paper and have met the following 4 require-
(21). The relative roles of these mechanisms to the clear- ments: (a) significant contributions to the conception and design, acquisi-
tion of data, or analysis and interpretation of data; (b) drafting or revising
ance of eccDNA and linear DNA molecules would re- the article for intellectual content; (c) final approval of the published arti-
quire further investigation. Nonetheless, the similarities cle; and (d) agreement to be accountable for all aspects of the article thus
8 Clinical Chemistry 00:0 (2021)Features of Circulating Fetal eccDNA
ensuring that questions related to the accuracy or integrity of any part of Honoraria: None declared.
the article are appropriately investigated and resolved. Research Funding: Hong Kong Research Grants Council Theme-
Based Research Scheme (T12-403/15N). Y.M.D. Lo, Li Ka Shing
L. Ji, statistical analysis; J. Deng, statistical analysis; P. Jiang, statistical
Foundation Endowed Professorship, funding to institution from Grail
analysis; M.M.S. Heung, administrative support; T.Y. Leung, provi- Collaborative Research Agreement.
sion of study material or patients; K.C.A. Chan, statistical analysis.
Expert Testimony: None declared.
Patents: S.T.K. Sin, patent application (16/829,771) related to
Downloaded from https://academic.oup.com/clinchem/advance-article/doi/10.1093/clinchem/hvaa326/6146145 by guest on 24 March 2021
Authors’ Disclosures or Potential Conflicts of Interest: Upon man-
uscript submission, all authors completed the author disclosure form. eccDNA; L. Ji, patent application (16/829,771) related to eccDNA; J.
Disclosures and/or potential conflicts of interest. Deng, patent application (16/829,771) related to eccDNA; P. Jiang,
patent application (16/829,771) related to eccDNA; K.C.A. Chan,
Employment or Leadership: K.C.A. Chan, Take2, DRA; R.W.K. 16/829,771; R.W.K. Chiu, US patent application 16/829,771;
Chiu, Clinical Chemistry, AACC; Y.M.D. Lo, Clinical Chemistry, Y.M.D. Lo, numerous patents and patent applications using circulat-
AACC; DRA Limited, Take2 Holdings Limited; P. Jiang, KingMed ing nucleic acids.
Future.
Consultant or Advisory Role: K.C.A. Chan, Grail; R.W.K. Chiu, Role of Sponsor: The funding organizations played no role in the de-
Illumina, Grail; Y.M.D. Lo, Grail, Decheng Capital. sign of study, review and interpretation of data, preparation of manu-
Stock Ownership: K.C.A. Chan, Illumina, Grail, Take2, DRA; script, or final approval of manuscript. The funding organizations
R.W.K. Chiu, Grail, Illumina, DRA, Take2; Y.M.D. Lo, Grail, DRA played no role in the choice of enrolled patients.
Limited, Take2 Holdings Limited; P. Jiang, Grail.
References
1. Kumar P, Dillon LW, Shibata Y, Jazaeri AA, Jones DR, and its cellular contributors uncovers placenta hypome- 16. Kumar P, Kiran S, Saha S, Su Z, Paulsen T, Chatrath A, et
Dutta A. Normal and cancerous tissues release extrachro- thylated domains. Genome Biol 2015;16:78. al. ATAC-seq identifies thousands of extrachromosomal
mosomal circular DNA (eccDNA) into the circulation. Mol 9. Lo YMD, Zhang J, Leung TN, Lau TK, Chang AMZ, Hjelm circular DNA in cancer and cell lines. Sci Adv 2020;6:
Cancer Res 2017;15:1197–205. NM. Rapid clearance of fetal DNA from maternal plasma. eaba2489.
2. Zhu J, Zhang F, Du M, Zhang P, Fu S, Wang L. Molecular Am J Hum Genet 1999;64:218–24. 17. Hoppers A, Williams L, Ponnaluri VKC, Sexton B, Saleh L,
characterization of cell-free eccDNAs in human plasma. 10. Yu SCY, Lee SWY, Jiang P, Leung TY, Chan KCA, Chiu Campbell M, et al. Enzymatic methyl-seq: next genera-
Sci Rep 2017;7:10968. RWK, et al. High-resolution profiling of fetal DNA clear- tion methylomes. J Biomol Tech 2020;31:S15.
3. Sin STK, Jiang P, Deng J, Ji L, Cheng SH, Dutta A, et al. ance from maternal plasma by massively parallel se- 18. Lee JY, Lee J, Yue H, Lee TH. Dynamics of nucleosome
Identification and characterization of extrachromosomal quencing. Clin Chem 2013;59:1228–37. assembly and effects of DNA methylation. J Biol Chem
circular DNA in maternal plasma. Proc Natl Acad Sci USA 11. Spektor R, Tippens ND, Mimoso CA, Soloway PD. Methyl- 2015;290:4291–303.
2020;117:1658–65. ATAC-seq measures DNA methylation at accessible chro- 19. Jiang P, Tong YK, Sun K, Cheng SH, Leung TY, Chan
4. Zhu J, Chen S, Zhang F, Wang L. Cell-free eccDNAs: a matin. Genome Res 2019;29:969–77. KCA, et al. Gestational age assessment by methylation
new type of nucleic acid component for liquid biopsy? 12. Møller HD, Parsons L, Jørgensen TS, Botstein D, and size profiling of maternal plasma DNA: a feasibility
Mol Diagn Ther 2018;22:515–22. Regenberg B. Extrachromosomal circular DNA is com- study. Clin Chem 2017;63:606–8.
5. Lo YMD, Chan KCA, Sun H, Chen EZ, Jiang P, Lun FMF, mon in yeast. Proc Natl Acad Sci USA 2015;112: 20. Kochanek S, Renz D, Doerfler W. Differences in the acces-
et al. Maternal plasma DNA sequencing reveals the E3114–22. sibility of methylated and unmethylated DNA to DNase I.
genome-wide genetic and mutational profile of the fe- 13. Sato S, Arimura Y, Kujirai T, Harada A, Maehara K, Nucl Acids Res 1993;21:5843–5.
tus. Sci Transl Med 2010;2:61ra91. Nogami J, et al. Biochemical analysis of nucleosome tar- 21. Kustanovich A, Schwartz R, Peretz T, Grinshpun A. Life
6. Chan KCA, Zhang J, Hui ABY, Wong N, Lau TK, Leung TN, geting by Tn5 transposase. Open Biol 2019;9:190116. and death of circulating cell-free DNA. Cancer Biol Ther
et al. Size distributions of maternal and fetal DNA in ma- 14. Shibata Y, Kumar P, Layer R, Willcox S, Gagan JR, Griffith 2019;20:1057–67.
ternal plasma. Clin Chem 2004;50:88–92. JD, Dutta A. Extrachromosomal microDNAs and chromo- 22. Yuen RK, Pe~naherrera MS, von Dadelszen P, McFadden
7. Lun FMF, Chiu RWK, Sun K, Leung TY, Jiang P, Chan somal microdeletions in normal tissues. Science 2012; DE, Robinson WP. DNA methylation profiling of human
KCA, et al. Noninvasive prenatal methylomic analysis by 336:82–6. placentas reveals promoter hypomethylation of multiple
genomewide bisulfite sequencing of maternal plasma 15. Kelly TK, Liu Y, Lay FD, Liang G, Berman BP, Jones PA. genes in early-onset preeclampsia. Eur J Hum Genet
DNA. Clin Chem 2013;59:1583–94. Genome-wide mapping of nucleosome positioning and 2010;18:1006–12.
8. Jensen TJ, Kim SK, Zhu Z, Chin C, Gebhard C, Lu T, et al. DNA methylation within individual DNA molecules.
Whole genome bisulfite sequencing of cell-free DNA Genome Res 2012;22:2497–506.
Clinical Chemistry 00:0 (2021) 9You can also read

















































