A Molecular Switch between Mammalian MLL Complexes Dictates Response to Menin-MLL Inhibition
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

RESEARCH ARTICLE
A Molecular Switch between Mammalian
MLL Complexes Dictates Response to
Menin–MLL Inhibition
Yadira M. Soto-Feliciano1, Francisco J. Sánchez-Rivera2, Florian Perner3,4,5, Douglas W. Barrows1,6,
Edward R. Kastenhuber2, Yu-Jui Ho2, Thomas Carroll6, Yijun Xiong3,4, Disha Anand3,5, Alexey A. Soshnev1,
Leah Gates1, Mary Clare Beytagh1, David Cheon1, Shengqing Gu7,8, X. Shirley Liu7,8, Andrei V. Krivtsov3,4,
Maximiliano Meneses9, Elisa de Stanchina9, Richard M. Stone10, Scott A. Armstrong3,4, Scott W. Lowe2, and
C. David Allis1
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
ABSTRACT Menin interacts with oncogenic MLL1-fusion proteins, and small molecules that
disrupt these associations are in clinical trials for leukemia treatment. By integrat-
ing chromatin-focused and genome-wide CRISPR screens with genetic, pharmacologic, and biochemical
approaches, we discovered a conserved molecular switch between the MLL1–Menin and MLL3/4–UTX
chromatin-modifying complexes that dictates response to Menin–MLL inhibitors. MLL1–Menin safe-
guards leukemia survival by impeding the binding of the MLL3/4–UTX complex at a subset of target
gene promoters. Disrupting the Menin–MLL1 interaction triggers UTX-dependent transcriptional acti-
vation of a tumor-suppressive program that dictates therapeutic responses in murine and human leuke-
mia. Therapeutic reactivation of this program using CDK4/6 inhibitors mitigates treatment resistance
in leukemia cells that are insensitive to Menin inhibitors. These findings shed light on novel functions of
evolutionarily conserved epigenetic mediators like MLL1–Menin and MLL3/4–UTX and are relevant to
understand and target molecular pathways determining therapeutic responses in ongoing clinical trials.
SIGNIFICANCE: Menin–MLL inhibitors silence a canonical HOX- and MEIS1-dependent oncogenic
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
gene expression program in leukemia. We discovered a parallel, noncanonical transcriptional program
involving tumor suppressor genes that are repressed in Menin–MLL inhibitor–resistant leukemia cells
but that can be reactivated upon combinatorial treatment with CDK4/6 inhibitors to augment therapy
responses.
INTRODUCTION lular carcinoma (19) and MLL1-rearranged (MLL1-r) leukemias
Menin is an evolutionarily conserved nuclear factor that (4, 20). Furthermore, over 1,000 germline and somatic MEN1
associates with chromatin to recruit (adapt) interacting pro- variants have been identified, some of which are linked to can-
teins (1). These include the Trithorax (Trx)-related MLL1 cer predisposition (21).
(KMT2A) and MLL2 (KMT2B) histone methyltransferase Given the pro-oncogenic role of Menin in acute leukemia
complexes (2, 3), MLL1 oncogenic fusion proteins (4), tran- and other malignancies, small-molecule inhibitors targeting
scription factors [e.g., c-MYC (5), JUND (6, 7), SMADs the Menin–MLL1 and Menin–MLL2 protein–protein interac-
(8, 9)], and other chromatin-bound proteins [e.g., LEDGF tions have shown great promise for intercepting and treat-
(10); reviewed in ref. 11]. ing different types of cancers (19, 22–27). Notably, three
Menin is a core subunit of the MLL1 (ref. 12) and MLL2 structurally different Menin–MLL inhibitors have recently
complexes (2) and is responsible for targeting these to chroma- entered clinical trials (NCT04065399, NCT04067336, and
tin (3). Menin is required for MLL1/MLL2-dependent H3K4 NCT04811560), and at least one has been granted fast-track
trimethylation of HOX genes and their stable long-term expres- designation by the FDA for the treatment of relapsed/refrac-
sion during development (2, 13). Menin has context-specific tory acute leukemias (25, 27, 28). Thus, an understanding of
functions in human diseases, acting as a tumor suppressor in the molecular mechanisms of action of these drugs would
neuroendocrine malignancies (14, 15) and in certain skin (16), facilitate the development of biomarkers to predict therapeu-
lung (17), and central nervous system (CNS) tumors (18) and tic response and resistance, and lead to the rational design of
as an oncogenic cofactor in other cancers, including hepatocel- more effective combination treatments.
1
Laboratory of Chromatin Biology and Epigenetics, The Rockefeller Corresponding Authors: C. David Allis, The Rockefeller University,
University, New York, New York. 2Cancer Biology and Genetics, Memo- Allis Lab, Box #78, 1230 York Avenue, New York, NY 10065. Phone:
rial Sloan Kettering Cancer Center, New York, New York. 3Department of 212-327-7839; E-mail: alliscd@rockefeller.edu; Scott W. Lowe,
Pediatric Oncology, Dana-Farber Cancer Institute, Boston, Massachusetts. Memorial Sloan Kettering Cancer Center, Sloan Kettering Institute,
4
Division of Hematology/Oncology, Boston Children’s Hospital and Harvard Cancer Biology and Genetics Program, New York, NY, 10065. Phone:
Medical School, Boston, Massachusetts. 5Internal Medicine C, Greifswald 646-888-3342; E-mail: lowes@mskcc.org; and Scott A. Armstrong,
University Medical Center, Greifswald, Germany. 6Bioinformatics Resource Harvard Medical School, Dana-Farber Cancer Institute, Department of
Center, The Rockefeller University, New York, New York. 7Department of Pediatric Oncology, Boston, MA, 02115. Phone: 617-632-2991; E-mail:
Biostatistics and Computational Biology, Dana-Farber Cancer Institute, Scott_Armstrong@dfci.harvard.edu
Harvard School of Public Health, Boston, Massachusetts. 8Center for Func- Cancer Discov 2023;13:146–69
tional Cancer Epigenetics, Dana-Farber Cancer Institute, Boston, Massa-
chusetts. 9Antitumor Assessment Core Facility, Memorial Sloan Kettering doi: 10.1158/2159-8290.CD-22-0416
Cancer Center, New York, New York. 10Leukemia Division, Department of This open access article is distributed under the Creative Commons Attribution
Medical Oncology, Dana-Farber Cancer Institute, Boston, Massachusetts. 4.0 International (CC BY 4.0) license.
Current address for Y.M. Soto-Feliciano and F.J. Sánchez-Rivera: David H. ©2022 The Authors; Published by the American Association for Cancer Research
Koch Institute for Integrative Cancer Research and Department of Biology,
Massachusetts Institute of Technology, Cambridge, Massachusetts.
Y.M. Soto-Feliciano, F.J. Sánchez-Rivera, and F. Perner are co–first authors.
JANUARY 2023 CANCER DISCOVERY | 147
RESEARCH ARTICLE Soto-Feliciano et al.
A Chromatin-focused C Ring1
Kdm2b Bcor Pcgf1
or genome-wide
sgRNA library 1 Phc1
Lorem ipsum
+ MI-503
Resistance gene rank
5,000 Mll4
T0 Tf Ncoa6
MLL3/4 Utx
MOI
Switch by MLL Complexes Dictates Menin Inhibitor Effects RESEARCH ARTICLE
E F
sgMen1-RFP Flow
MI-503
cytometry
BFP
Viability?
sgRNA-RFP 96 h
sgUtx-BFP
RFP
P = 0.0003 P = 0.0100
Day 16
P < 0.0001
1.5 10 5 49.4 0.26 4 P = 0.0139
P = 0.0001
4
10
sgControl-BFP
P = 0.0487
103
(relative to DMSO)
1.0 2
Relative cell proliferation
102
Cell viability
101
50.1 0.27
100
0.5 100 101 102 103 104 105 0
sgMen1-RFP
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
105 49.5 16.9
0.0 −2
sgCtrl sg1 sg2 sg3 sgUtx_2-BFP 104
sgUtx 103
DMSO 102
−4
MI-503 101 sgCtrl sg1 sg2 sg3
29.6 3.97 BFP
100
100 101 102 103 104 105 sgUtx-BFP
sgMen1-RFP sgMen1-RFP
Figure 1. (Continued) E, Viability assay from cells treated with vehicle (DMSO, black) or Menin–MLL inhibitor (MI-503, red) for 96 hours (mean ± SEM,
n = 3 infection replicates, P values calculated by Student t test). sgCtrl, control sgRNA targeting a nongenic region on chromosome 8. F, Relative cell
proliferation is shown as the proliferation of double-positive cells (sgMen1-RFP + sgUtx-BFP or sgMen1-RFP + sgCtrl-BFP) relative to single-positive
cells (sgMen1-RFP) 16 days after infection measured by flow cytometry (mean ± SEM, n = 3 infection replicates, P values calculated by Student t test).
Representative FACS plots are shown for sgControl- and sgUtx-targeted cells.
lysine 4 (H3K4) monomethyltransferases Mll3 (Kmt2c) and inhibition, pointing to a previously unknown functional
Mll4 (Kmt2d) were the most significantly enriched in the cross-talk between these chromatin-modifying complexes.
Menin–MLL inhibitor context (Fig. 1B, red circles). This To determine the effects of MLL3/4–UTX loss of function
result was unexpected given previously described canoni- in leukemia cell proliferation, we performed in vitro growth
cal functions for the mammalian MLL complexes (35). For competition assays (Supplementary Fig. S3A). We found that
example, the MLL1/2–Menin complex (disrupted by MI-503) Utx or Mll3 inactivation by CRISPR did not have a significant
is known to catalyze chromatin modifications associated impact on leukemia cell proliferation, but Mll4 inactiva-
with transcription at promoters, including dimethylation or tion decreased leukemia cell growth (ref. 43; Supplementary
trimethylation of H3K4 (H3K4me2/3; refs. 13, 36, 37). On Fig. S3B–S3D). Consistent with our genetic screening results,
the other hand, the MLL3/4–UTX complex has been shown Utx inactivation using three distinct sgRNAs significantly
to regulate enhancer states by serving as the major H3K4 increased the viability of MI-503–treated leukemia cells rela-
monomethyltransferase (H3K4me1; refs. 38–42). tive to their wild-type counterparts (Fig. 1E; Supplemen-
To assess whether these results were idiosyncratic of the cell tary Fig. S3E). In addition, Mll3- or Mll4-deficient leukemia
line, library, or inhibitor used, we performed a genome-wide cells treated with MI-503 exhibited a proliferative advantage
CRISPR screen in an independently derived MLL-AF9 mouse over wild-type cells under drug treatment (Supplementary
cell line using VTP-50469, a more potent (than MI-503), Fig. S3F and S3G). These orthogonal results establish the
selective, and orally bioavailable Menin–MLL inhibitor (25, MLL3/4–UTX complex as a central modulator of therapy
27, 28). sgRNAs targeting Utx, Mll3, and Mll4 were also response to Menin–MLL inhibition in acute leukemia.
among the most significantly enriched candidate genes iden- Because UTX was the most significantly enriched chromatin
tified in this genome-wide screen (Fig. 1C; Supplementary factor in our screen and this protein is shared by both MLL3
Fig. S2A–S2C; Supplementary Table S2), whereas shared sub- and MLL4 complexes (42, 44, 45), we focused on UTX disrup-
units between the two types of MLL complexes scored simi- tion to probe the molecular mechanisms linked to resistance
larly in both vehicle and Menin–MLL inhibitor conditions to Menin–MLL inhibition. We first tested whether genetic Utx
(Fig. 1B–D; Supplementary Fig. S2D). These results suggest inactivation could rescue the effects of Menin-specific abla-
that the core subunits of the MLL3/4–UTX complex modu- tion in MLL-AF9 leukemia cells. CRISPR-mediated deletion of
late the therapeutic response of leukemia cells to Menin–MLL Men1 led to robust inhibition of proliferation (Supplementary
JANUARY 2023 CANCER DISCOVERY | 149
RESEARCH ARTICLE Soto-Feliciano et al.
Fig. S4A), but codeletion of Utx suppressed this phenotype, (TSS) ± 2 kb], which was decreased when its canonical
such that Men1-deficient MLL-AF9 cells were able to prolifer- interactions with MLL1/2 and MLL-FPs were disrupted by
ate despite the presence of the drug (Fig. 1F; Supplementary MI-503 (Fig. 2A). Genome-wide enrichment of Menin was
Fig. S4B–S4F). These results establish a previously unknown also decreased, as evidenced by the reduction in the num-
epistatic relationship between Men1 and Utx in acute leukemia. ber of ChIP peaks after MI-503 treatment (Supplementary
Of note, preexisting mutations in MLL3, MLL4, or UTX, some Fig. S6A). We observed only a small number of UTX peaks
of which lead to truncated versions of the respective proteins, under basal conditions; however, MI-503 treatment led to
are found in about 8% of patients with acute myeloid leuke- a substantial increase in UTX chromatin association, with
mia (AML), myelodysplastic syndromes, or myeloproliferative predominant binding at promoters (Fig. 2B; Supplementary
neoplasia (Fig. 1F; Supplementary Fig. S4G). The experimental Fig. S6A). UTX binding sites detected after MI-503 treat-
setup used for our experiments strived to model the conse- ment included those observed under basal conditions and
quent loss of function of MLL3/4–UTX proteins in these cases. more than 13,000 additional sites (Supplementary Fig. S6B
To determine if the genetic interaction between Menin and S6C). Interestingly, these UTX binding sites were not
and UTX depends on the MLL-fusion protein (MLL-FP), we enriched for lineage-specific enhancers (Supplementary
ablated the MLL1–AF9 oncogenic transgene used to generate Fig. S6C). We also confirmed that the MI-503–dependent
this model using CRISPR-Cas9 (46–48). Importantly, survival enrichment of UTX on chromatin was not simply the result
of leukemia cells remains dependent on the presence of the of increased Utx expression or UTX protein stability (Fig. 2C
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
MLL–AF9 fusion protein even when UTX is lost, demonstrating and D). These data show that disruption of the Menin–
that MLL-FP–driven canonical gene expression is intact even MLL1 interaction leads to dynamic recruitment of UTX to
after loss of MLL3/4 function (Supplementary Fig. S5A–S5C). promoter regions (Fig. 2B), implying a previously unrec-
Codeletion of UTX and MEN1 in an NPM1c-mutant human ognized functional role for the MLL3/4–UTX complex in
leukemia cell line (49, 50) also bypassed the proliferation promoter regulation in leukemia.
defects associated with loss of MEN1 alone (Supplementary Intriguingly, promoter regions that became uniquely occupied
Fig. S5D and S5E). Therefore, Menin–MLL inhibitors act by UTX significantly overlapped with those where Menin was
through an evolutionarily conserved pathway that involves a lost, suggesting a dynamic molecular mechanism between
functional cross-talk between Menin and UTX and is down- these complexes at a subset of gene promoters (Fig. 2E and F;
stream or independent of MLL-FPs. Supplementary Fig. S6D). Correlation analysis also suggested
Menin is required for expression of canonical MLL-FP that reduction of Menin binding by MI-503 coincided with
target genes like Meis1, which is required for leukemia mainte- increased UTX chromatin occupancy at the same genomic
nance (25, 36, 37, 51, 52). Moreover, Meis1 overexpression has loci (Supplementary Fig. S6E). Comparing UTX chromatin-
been shown to partially rescue the leukemic stem cell tran- binding between MLL-AF9 cells and normal hematopoietic
scriptional program suppressed by Menin–MLL inhibition stem/progenitor cells revealed that UTX occupies a unique
(27). To determine if the genetic interaction between Menin set of binding sites in leukemia cells under basal conditions.
and UTX regulates the expression of these MLL-FP targets, we On the other hand, MI-503 treatment led to more permis-
first treated mouse MLL-AF9 leukemia cells with MI-503 and sive binding, including to the majority of sites bound in
confirmed that it leads to the robust downregulation of Meis1 non–MLL-r cells, supporting the concept that MLL1–Menin
(Supplementary Fig. S5F). However, to our surprise, cells complexes restrict UTX binding in MLL-FP–driven leukemia
genetically deficient for both Men1 and Utx showed a similar (Supplementary Fig. S6F). Treatment of mouse fibroblasts
reduction in Meis1 levels (Supplementary Fig. S5G) yet were (that are not dependent on Menin) with MI-503 did not force
able to proliferate (Fig. 1F; Supplementary Fig. S4E). When UTX mobilization or binding to promoters, indicating that
we knocked out Utx in mouse leukemia cells, we observed this molecular switch was specific to Menin-dependent cells
that Meis1 mRNA levels are not significantly different when (Supplementary Fig. S7A–S7E).
compared with the UtxWT counterparts (Supplementary The above results indicate that Menin inhibition displaces
Fig. S5H). We also observed significant downregulation of MLL1–Menin transcriptional regulatory complexes from pro-
Meis1 expression after treatment with a Menin–MLL inhibi- moters, enabling UTX to bind and potentially regulate target
tor. These data support a genetic epistasis model between gene expression (Fig. 2). In agreement, analysis of the chroma-
Menin and UTX in Menin-dependent mammalian cells and tin-binding profiles of their cognate H3K4 methyltransferases
suggest that factors beyond Meis1 can sustain the proliferative and their enzymatic histone modifications (Supplementary
capacity of MLL1-r and non–MLL1-r leukemia cells. Fig. S8A and S8B) revealed that Menin–MLL1 inhibition
led to a significant decrease in MLL1 chromatin enrich-
The MLL1–Menin Complex Restricts Chromatin ment and a concomitant decrease of its enzymatic prod-
Occupancy of MLL3/4–UTX at Target uct, H3K4me3 (Supplementary Fig. S8C–S8E). In contrast,
Gene Promoters Menin–MLL1 inhibition led to increased enrichment of the
Given previous work suggesting a primary role for Menin MLL3/4 enzymes at promoter regions co-occupied by UTX
at promoters (6, 53) and UTX at enhancers (42), we performed and a concomitant increase in H3K4me1 signal at these same
chromatin immunoprecipitation followed by sequenc- loci (Supplementary Fig. S8C, S8F, and S8G). Of note, these
ing (ChIP-seq) to examine the genome-wide binding pat- loci are distinct from those known to be bound and regulated
terns of their respective protein complexes (Supplementary by the MLL-FPs (ref. 30; Supplementary Fig. S9A–S9E). These
Table S3). As expected, Menin showed strong enrichment at data indicate that disruption of the MLL1–Menin interac-
promoter regions [here defined as transcription start sites tion induces targeting of the core enzymatic subunits of the
150 | CANCER DISCOVERY JANUARY 2023 AACRJournals.org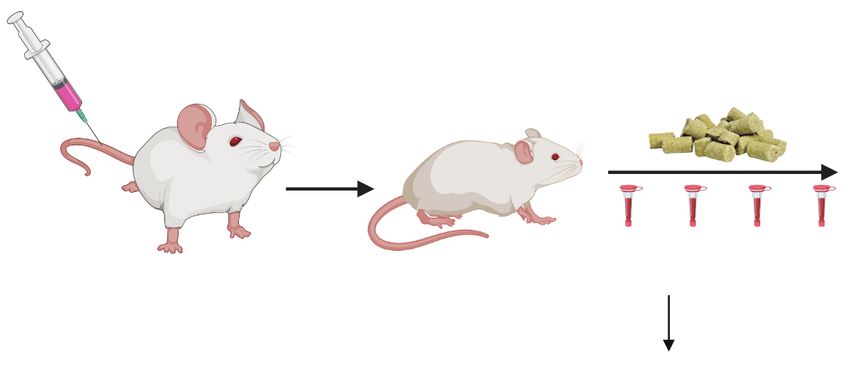
Switch by MLL Complexes Dictates Menin Inhibitor Effects RESEARCH ARTICLE
A E Menin ChIP-seq UTX ChIP-seq
Menin ChIP-seq
DMSO MI-503 DMSO MI-503
DMSO 1.0 1.0 1.5 1.5
UTX+
0.4 0.8 0.8
ChIP signal
MI-503
Menin occupancy (RPM)
1.0 1.0 UTX −
0.6 0.6
0.4 0.4 0.5
0.5
0.3
0.2 0.2
0.0 0.0 0.0 0.0
−2.0 TSS 2.0 −2.0 TSS 2.0 −2.0 TSS 2.0 −2.0 TSS 2.0
0.2
0.1
UTX+ promoters in MI-503
−2,000 −1,000 TSS 1,000 2,000
(8,856 peaks)
B UTX ChIP-Seq
DMSO
0.4
MI-503
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
UTX occupancy (RPM)
0.3
0.2
UTX− promoters in MI-503
(15,522 peaks)
0.1
−2,000 −1,000 TSS 1,000 2,000
C
Utx
1.5
P = 0.0018
Relative mRNA levels
S
−2 0
S
−2.0
S
−2.0
S
0
.0
.0
.0
.0
1.0
2.
2.
TS
TS
TS
TS
2
2
−2
F
[0−2.88]
DMSO
0.5
Menin
0.0 [0−2.88]
MI-503
S O
-5 03
DM MI
D SO 03 [0−8.10]
I-5
DMSO
DM M
Menin
UTX
[0−8.10]
MI-503
UTX
HSP90
RefSeq
Kifc5b Phf1 Cuta Syngap1 Zbtb9
Figure 2. MLL1–Menin complex restricts chromatin occupancy of MLL3/4–UTX at promoters of target genes. A, Metagene analysis showing the aver-
age chromatin occupancy of Menin at the TSS, and a region 2,000 bp downstream and upstream of the TSS. Signals corresponding to cells treated with a
Menin–MLL inhibitor (MI-503, solid) compared with cells treated with vehicle (DMSO, dotted) for 96 hours are shown. RPM, reads per million. B, Metagene
analysis showing the average chromatin occupancy of UTX at the TSS, and a region 2,000 bp downstream and upstream of the TSS. Signals corresponding
to cells treated with a Menin–MLL inhibitor (MI-503, solid) compared with cells treated with vehicle (DMSO, dotted) for 96 hours are shown. C, Relative
Utx mRNA levels determined by qPCR in mouse MLL-AF9 leukemia cells treated with a Menin–MLL inhibitor (MI-503, red) compared with vehicle (DMSO,
black) for 96 hours (mean ± SEM, n = 3 replicates, P value calculated by Student t test). D, Immunoblot analysis of Menin, UTX, and HSP90 proteins (load-
ing control) upon Menin–MLL inhibitor (MI-503) treatment of mouse MLL-AF9 leukemia cells for 96 hours. E, Heat maps displaying Menin (black) and
UTX (purple) ChIP-seq signals mapping to a 4-kb window around the TSS. Data are shown for DMSO and MI-503–treated cells for 96 hours. Metagene
plot shows the average ChIP-seq signal for Menin or UTX at promoters that are UTX+ (green) or UTX− (black) after MI-503 treatment. F, Genome browser
representation of ChIP-seq normalized reads for Menin (black) and UTX (purple) in mouse MLL-AF9 leukemia cells treated with either vehicle (DMSO) or a
Menin–MLL inhibitor (MI-503) for 96 hours.
JANUARY 2023 CANCER DISCOVERY | 151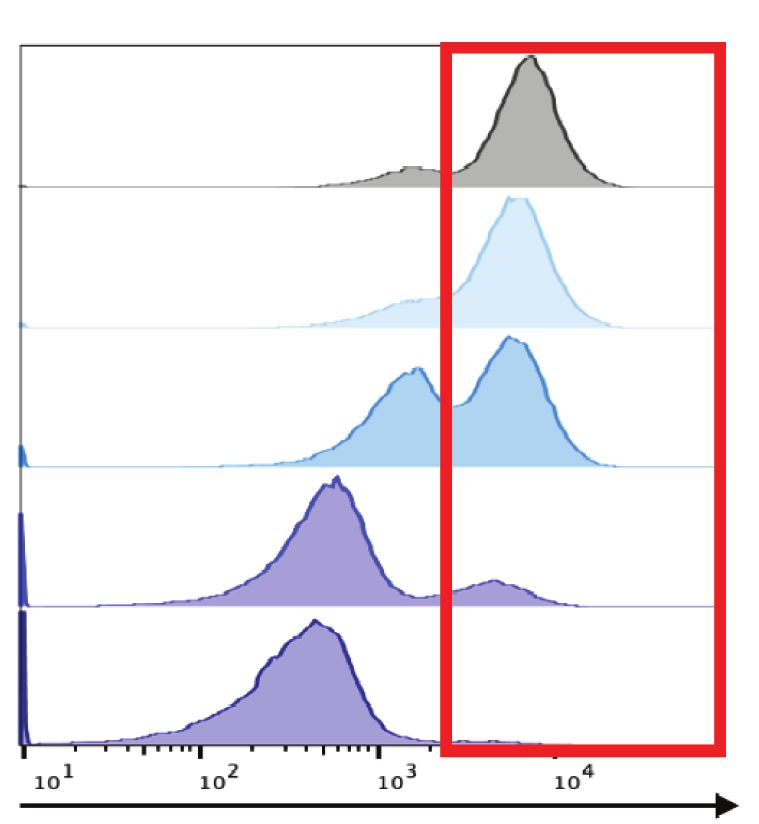
RESEARCH ARTICLE Soto-Feliciano et al.
MLL3/4–UTX complex to noncanonical sites that are nor- of NfyaKO cells with MI-503 led to a significant increase
mally bound by the MLL1–Menin complex. in UTX occupancy at genomic regions normally bound by
Promoter-associated H3K4me1 has been shown to facilitate Menin and NF-YA at steady state (Fig. 3E; Supplementary
transcriptional repression in other cellular settings (54), sug- Fig. S12C). Collectively, these results implicate NF-Y as one
gesting that deposition of H3K4me1 at gene promoters could of the sequence-specific transcription factors that marks
depend on context (55). To determine the functional implica- Menin–UTX target sites and potentially plays a functional
tions of the Menin–MLL inhibitor–induced colocalization of role in mediating the Menin–UTX switch at these loci.
UTX, MLL3/4, and H3K4me1 at target gene promoters, we lev-
eraged the H3 lysine-4-to-methionine (K4M) “oncohistone” Transcriptional Coregulation of Tumor-Suppressive
mutant as an orthogonal molecular tool to destabilize and Pathways by a Menin–UTX Molecular Switch
alter the function of the MLL3/4–UTX complex (ref. 56; To further probe the molecular phenotypes regulated by
Supplementary Fig. S10A). Expression of H3.1K4M in MLL- this epigenetic switch, we performed transcriptional profiling
AF9 leukemia cells led to a proliferative advantage only in the of MLL-AF9 leukemia cells treated with MI-503 and identi-
context of Menin–MLL inhibition (Supplementary Fig. S10B– fied pathways that are reciprocally regulated by Menin and
S10D), demonstrating that destabilization of the MLL3/4–UTX UTX. Although the activity of the MLL1–Menin complex
complex (Supplementary Fig. S10E) can phenocopy the is associated with actively transcribed developmental genes
intrinsic resistance of Utx-, Mll3-, or Mll4-deficient cells to (2, 13), MI-503 treatment resulted in both up- and downregu-
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
Menin–MLL inhibition (Fig. 1E; Supplementary Fig. S3E– lation of gene expression, with the majority of significantly
S3G). These results further support a model whereby the upregulated genes reciprocally bound by Menin and UTX
MLL3/4–UTX complex serves as a context-specific and cen- (Fig. 4A and B).
tral modulator of therapy response to Menin–MLL inhibition To gain further insights into the mechanism, we performed
in leukemia cells. ChIP-seq against acetylated H3K27 and H4K16 (H3K27ac
and H4K16ac; histone modifications associated with gene
NF-YA Contributes to Genomic Specificity of the activation; ref. 59) and found that the levels of these modi-
Menin–UTX Molecular Switch on Chromatin fications increased at sites where the MLL3/4–UTX complex
Because Menin lacks a defined DNA binding domain (57), is enriched upon Menin–MLL inhibition (Supplementary
we tested whether a sequence-specific DNA binding factor may Fig. S13A and S13B). These loci also showed increased bind-
regulate the switch between MLL1–Menin and MLL3/4–UTX ing of MOF, a histone H4K16 acetyltransferase (Supplemen-
occupancy at specific promoters. Motif analysis on the tary Fig. S13C; refs. 60–62). Thus, Menin–MLL inhibition
genomic regions bound by Menin and UTX in leukemia cells produces dynamic changes in gene expression due to loss of
and fibroblasts revealed that NF-Y sequence motifs were the MLL1–Menin-dependent repressive activity and a concomi-
most significantly and selectively enriched in MLL-AF9 tant increase in histone modifications associated with gene
leukemia cells (Fig. 3A; Supplementary Fig. S11A and activation (including H4K16ac and H3K27ac; refs. 63, 64).
S11B). In agreement, ChIP-seq analysis for NF-YA (the To determine if UTX was necessary for gene activation,
DNA binding and transactivation subunit of the NF-Y we generated UtxKO MLL-AF9 leukemia cells, treated these
complex; ref. 58) showed that it co-occupies sites recipro- with MI-503, and profiled them by RNA sequencing (RNA-
cally bound by Menin and UTX (Fig. 3B; Supplementary seq; Supplementary Fig. S14A and S14B). We found that
Fig. S11C). Furthermore, Menin–MLL inhibition did not genes bound by Menin and UTX that are induced by MI-503
affect NF-YA protein levels, but it decreased NF-YA binding failed to get activated in UtxKO cells, suggesting UTX func-
to chromatin (Fig. 3B and C; Supplementary Fig. S11D). tion is necessary for their transcriptional activation upon
To determine how changes in NF-YA genomic binding Menin displacement from chromatin (we refer to these genes
relate to changes in Menin, UTX, and MLL3/4 chromatin as “Menin–UTX targets”; Fig. 4C; Supplementary Table S4).
binding in the context of Menin–MLL inhibition, we per- Consistent with the idea that this mechanism is independent
formed correlation analysis of these ChIP-seq datasets. We of the MLL-FP, we found that MI-503-treated UtxKO cells still
found that genomic loci where NF-YA and Menin signals exhibited downregulation of canonical MLL–AF9 targets and
are depleted the most are also regions of high UTX and induction of myeloid differentiation programs (Supplemen-
MLL3/4 enrichment in cells treated with the Menin–MLL tary Fig. S15A–S15C), as has been observed in UtxWT cells (22,
inhibitor (Supplementary Fig. S11E). This pattern was 25, 27). Moreover, these cells were able to proliferate without
evident both genome-wide and more locally at promot- reexpression of Meis1 and Hoxa9—two critical Menin–MLL-FP
ers (TSSs). These results support a model whereby Menin targets (Fig. 1E; Supplementary Figs. S5G and S15D; refs. 25,
and NF-YA colocalize at certain genomic loci that become 27, 51, 52). These data suggest a new paradigm whereby the
available for binding by the MLL3/4–UTX complex in the effects of Menin–MLL inhibition on MLL-FP target genes
context of Menin–MLL inhibition. are independent of its effects on Menin–UTX transcriptional
To dissect the relationship between Menin, UTX, and targets, and that concomitant induction of tumor-suppres-
NF-YA, we generated NfyaKO MLL-AF9 leukemia cells and sive gene expression programs and repression of canonical
treated them with MI-503 (Supplementary Fig. S12A). MLL-FP targets are required for the antileukemic activity of
Disruption of Nfya significantly decreased the viability of Menin–MLL1 inhibitors.
MI-503–treated leukemia cells at two different time points, To gain insight into cellular and molecular pathways
suggesting a functional relationship between Nfya and Men1 regulated by the Menin–UTX switch, we performed gene
(Fig. 3D; Supplementary Fig. S12B). Moreover, treatment ontology analysis (65, 66) of Menin–UTX targets and found
152 | CANCER DISCOVERY JANUARY 2023 AACRJournals.org
Switch by MLL Complexes Dictates Menin Inhibitor Effects RESEARCH ARTICLE
A B
[0−2.88]
MI-503 DMSO MI-503 DMSO MI-503 DMSO
Motif Name P
Menin
[0−2.88]
GG
A
C
T
A
CCCAATCGGAG
T
G T A G T A G
C T G
C T G
C A TCT
A T
C
A
NFY 1e−130
G
C
GGGGGCGGAGC
A T
TA
CAA TC C T C T A C T A G
T
A C T A C T A
G
C TT C
A
T
A
G
Sp5 1e−63 [0−8.10]
C
AG CCGCCCCC
G CC
G
TATAAC T AG
T G T G T A C T A G T A G T A G A
A
TT G
T
G
A
Sp1 1e−50 UTX
[0−8.10]
A
T
G
C
GGGCTGGTGGC
C T A
C T A C T A G A C T
A
CT
AACGC
TT
A A
KLF5 1e−45
[0−39]
AGGGGGCGG
G T
TG
GC T AG KLF14 1e−41
TA
CCA
T CC
T C T A
A C T A G A C T A C
A C
ACT
TTA
NF-YA
[0−39]
C SO 03
I-5 Kifc5b Phf1 Cuta Syngap1 Zbtb9
DM M
UTX ChIP-seq
NF-YA E
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
NfyaWT + DMSO NfyaWT + MI-503 NfyaKO + DMSO NfyaKO + MI-503
ChIP signal 2
HSP90 1.5
1
0.5
D P = 0.0011 0
P = 0.0034
1.5
P = 0.0052
(relative to DMSO)
1.0
Cell viability
Overlap Menin (DMSO) and UTX (MI-503)
0.5
0.0
(8,856 peaks)
sgCtrl sg1 sg2 sg3
sgNfya
DMSO
MI-503
.0
.0
.0
.0
S
S
S
S
−2 0
−2.0
−2 0
0
2.
2.
2.
TS
TS
TS
TS
−2
2
Figure 3. NF-YA contributes to genomic specificity of the Menin–UTX molecular switch on chromatin. A, HOMER de novo motif analysis of overlapping
ChIP-seq peaks between Menin (in DMSO) and UTX (in MI-503) in mouse MLL-AF9 leukemia cells. B, Genome browser representation of ChIP-seq normal-
ized reads [average reads per kilobase per million mapped reads (RPKM)] for representative loci bound by Menin (black), UTX (purple), and NF-YA (red) in
cells treated with vehicle (DMSO) or a Menin–MLL inhibitor (MI-503) for 96 hours. C, Immunoblot analysis of NF-YA and HSP90 proteins (loading control)
upon Menin–MLL inhibitor (MI-503) treatment of mouse MLL-AF9 leukemia cells for 96 hours. D, Viability assay from cells treated with vehicle (DMSO,
black) or a Menin–MLL inhibitor (MI-503, red) for 96 hours (mean ± SEM, n = 3 infection replicates, P values calculated by Student t test). sgCtrl, control
sgRNA targeting a nongenic region on chromosome 8. E, Heat maps displaying UTX ChIP-seq signal mapping to a 4-kb window around the TSS in NfyaWT
(red) or NfyaKO (black) mouse MLL-AF9 leukemia cells treated with vehicle (DMSO) or a Menin–MLL inhibitor (MI-503) for 96 hours. Metaplot represents
the average UTX ChIP-seq signal at promoters.
JANUARY 2023 CANCER DISCOVERY | 153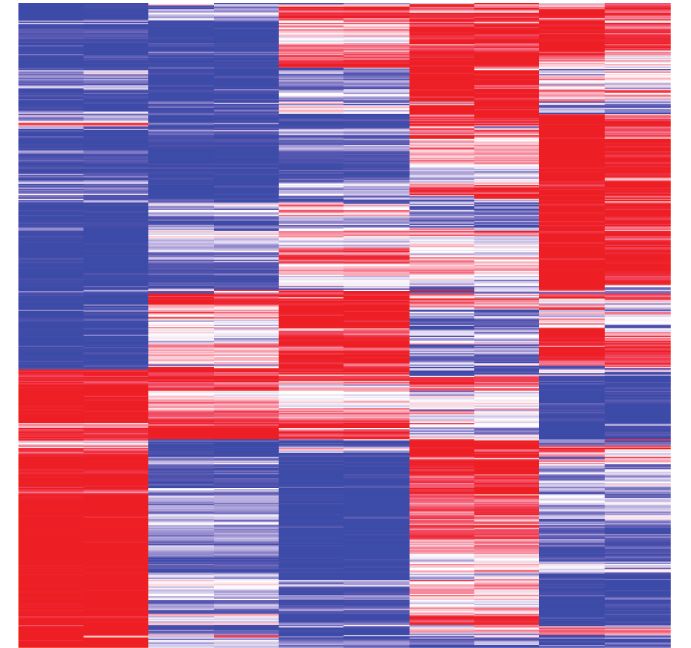
RESEARCH ARTICLE Soto-Feliciano et al.
A 921 genes DN 1,030 genes UP B
Il18rap Cxcr2
300 Il1r2
Mpeg 4,000
P < 0.0001
3,000
Number of genes
Aldh1a7 Muc13
−log10 (P value)
200
Gm1110 2,000
1,000
100 Pira6
Wt1
Pira1
0
Meis1 UTX Menin peaks
Piwil4 Flt3 Tgfbi UP in MI-503
0 Naip7
−4 0 4 8
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
Log2 (fold change MI-503 vs. DMSO)
C D Fridman senescence_UP
0.05
Utx WT Utx KO
−0.05 NES = −1.48
Enrichment score (ES)
P < 0.0001 FDR q value = 0.05
−0.15
−0.25
1 −0.35
−0.45
z-score
0
Utx WT + DMSO UtxWT + MI-503
−1 0.5
NES = 1.50
Enrichment score (ES)
0.4 FDR q value = 0.05
0.3
0.2
SO
03
SO
03
0.1
I-5
I-5
M
M
D
M
D
M
0.0
−0.1
Utx WT + MI-503 UtxKO + MI-503
Figure 4. Transcriptional coregulation of tumor-suppressive pathways by the Menin–UTX switch. A, Volcano plot of differentially expressed genes
in mouse MLL-AF9 leukemia cells treated with a Menin–MLL inhibitor (MI-503) or vehicle (DMSO) for 96 hours. Significantly (P < 0.05) downregulated
(DN) genes are shown on the left (n = 921 genes). Significantly (P < 0.05) upregulated (UP) genes are shown on the right (n = 1,030). B, Upset plot showing
significant overlap (red) between genes that undergo replacement of Menin by UTX at their promoters and MI-503–induced genes. P value for overlap is
shown. C, Box plot showing expression levels of genes that are induced by Menin–MLL inhibitor (MI-503) treatment and are bound by UTX at their pro-
moters in this condition and by Menin at steady state. Expression levels are shown for UtxWT and UtxKO leukemia cells. The midline in box plots represents
median. P value for MI-503 comparison is shown. D, GSEA showing that a Menin–UTX targets induced by a Menin–MLL inhibitor (MI-503) are significantly
enriched for genes regulating cellular senescence. FDR, false discovery rate; NES, normalized enrichment score. (continued on following page)
significant evidence for their association with transcriptional cells and observed a significant correlation between the pres-
programs related to proliferation, differentiation, and survival ence of UTX and the expression of senescence-associated genes
(Supplementary Fig. S16A; Supplementary Table S4). To fur- following treatment (Fig. 4D; Supplementary Fig. S16B–S16G;
ther evaluate the relevance of these gene ontology terms, we Supplementary Table S4; ref. 68). This correlation was also
performed gene set enrichment analysis (GSEA; ref. 67) on observed when we analyzed a curated list of genes involved
transcriptional data from MI-503-treated UtxWT and UtxKO in cell-cycle arrest and therapy-induced senescence (Fig. 4E;
154 | CANCER DISCOVERY JANUARY 2023 AACRJournals.orgSwitch by MLL Complexes Dictates Menin Inhibitor Effects RESEARCH ARTICLE
E Senescence-associated F
cell cycle and SASP
Utx WT Utx KO IL6
MI-503 − + − +
P = 0.0005
IL6 (pg/mL per million cells)
CSF1
TAP1 6,000
NPC1
TNFAIP3
UtxWT
ISG15
NFE2L2
TOP2A
Utx KO
MAP1LC3B
NFKB2
4,000 P = 0.0016
KLF6
CDK2
TAPBP P = 0.0700 P = 0.0806
PNRC1
NEURL1B
FOSL2
STAT1
2,000
CD82
FBXO5
L1CAM
TNFRSF9
EPS8
MX1
DNAJB4
0
IRF1
ASF1B
1.5 MI-503 − + − + − + − +
CTDSP1
RHOB
PLAU Day 4 Day 6
HMGB2
IFNAR1
1
RNF149
ATP2C1
TMPO
DLGAP5
KIF11
PSEN1
0.5 G IFNβ-1
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
B2M
z-score
IFIT3
IFNβ-1 (pg/mL per million cells)
CYP1B1
0 10 P = 0.0009
IGFBP7
CXCL11 UtxWT
HIF1A
HMMR 8 Utx KO
KIF20A
HMGN2 −0.5 P = 0.0051
CDKN1A
IFIT2
6 P = 0.3395
BIRC2 P = 0.0627
SAT1
OPTN −1 4
KDM6B
MUC1
COMMD4
RAB27B 2
BIRC5
UBE2C −1.5
STMN1
CCND1
0
XAF1
STXBP1
TK1 −2
CDK1
IL6
AURKB
MI-503 − + − + − + − +
PLK1
CCNA2
PRC1
Day 4 Day 6
FOXM1
IQGAP3
MKI67
STAT3
TNFAIP2
NCAPD2
KIF2C
DEPDC1B
CDKN3
NUSAP1
SPC25
CSTB
SGMS2
GM2A
CENPA
CDCA3
IL15
ACVRL1
GAS6
CCNB2
FAM83D
DNMT1
NUF2
SQSTM1
PVR
CCNE1
DEK
SMC4
IL1R1
CLU
GADD45A
ID1
ATF3
CXCL10
ITGA5
CDC20
IFNGR1
CCNB1
LMNB1
WEE1
NINJ1
NAMPT
PIR
BST2
RHNO1
RAD51AP1
CENPM
BUB1B
USP1
HMGB3
DUT
CCL2
CX3CL1
TYMS
SPC24
MYBL2
Figure 4. (Continued) E, Heat map showing relative gene
SERPINE1 expression levels of senescence-associated cell-cycle and
RCAN1
IL15RA
AURKA
senescence-associated secretory phenotype (SASP) genes in
CDK4
MCM3
mouse UtxWT and UtxKO MLL-AF9 leukemia cells treated with a
ORC6
RNASEH2A Menin–MLL inhibitor (MI-503) or vehicle (DMSO) for 96 hours.
IMPA2
DDR1 F and G, Secreted levels of IL6 and IFNβ-1 in conditioned media
NUCKS1
HMGA1 derived from mouse UtxWT (black) and UtxKO (red) MLL-AF9 leuke-
MCM6
CHEK1 mia cells treated with a Menin–MLL inhibitor (MI-503) or vehicle
(DMSO) for 4 or 6 days. Data are quantified as pg/mL of secreted
POLE2
ATP2A2
HELLS
EXO1 cytokine per million cells (mean ± SEM, n = 3 replicates, P values
EZH2
NASP calculated by Student t test).
JANUARY 2023 CANCER DISCOVERY | 155RESEARCH ARTICLE Soto-Feliciano et al.
ref. 69). These transcriptional changes are consistent with the UTX induced upon treatment with MI-503 (Fig. 2B and E)
fact that Menin–MLL inhibition induces a combination of did not affect global levels or distribution of H3K27me3
cell-cycle arrest, apoptosis, and differentiation (Supplemen- (Supplementary Fig. S20A and S20B), suggesting that this
tary Fig. S1E–S1G; ref. 22), as well as our observation that histone modification might not play a critical role in these
the senescence-associated H4K16ac modification increases phenotypes. To functionally test this possibility, we generated
with MI-503 treatment (Supplementary Fig. S13B and S13C; isogenic Ezh2KO;UtxWT and Ezh2KO;UtxKO MLL-AF9 leukemia
ref. 70). To gain additional insights into senescence-associated cells and confirmed the absence of H3K27me3 by immuno-
gene activation mechanisms, we performed ChIP-seq for RNA blotting (Supplementary Fig. S20C). Consistent with our
polymerase II and observed increased occupancy at the pro- model, Ezh2KO;UtxWT leukemia cells remained exquisitely sen-
moters of senescence-associated genes that are upregulated sitive to MI-503, whereas Ezh2KO;UtxKO cells were resistant to
by MI-503 treatment (Supplementary Fig. S16H). Thus, the Menin–MLL inhibition (Supplementary Fig. S20D). These
interplay between Menin and UTX in the context of Menin– results demonstrate that the catalytic domain of UTX is
MLL inhibition regulates the expression of genes involved in dispensable for UTX-dependent phenotypes and therapeutic
cell-cycle arrest and senescence. responses to Menin–MLL inhibitors.
The cellular senescence program is highly complex and
characterized in part by induction of permanent cell-cycle Combinatorial Targeting of Menin and CDK4/6
arrest and a senescence-associated secretory phenotype (SASP; Overcomes Resistance Associated with
MLL3/4–UTX Dysfunction
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
ref. 71). To determine if Menin–MLL inhibition induces SASP,
we measured cytokine and chemokine levels in conditioned Senescence can be regulated at both transcriptional and
media from MLL-AF9 leukemia cells treated with MI-503. posttranscriptional levels, and chromatin regulation has
Secretion of prototypical SASP cytokines such as IL6, IFNβ-1, been functionally implicated in modulating these programs
IL3, and IL15 was induced upon Menin–MLL inhibition in (70, 78, 79). We examined whether the Menin–UTX molecu-
a Utx-dependent manner, supporting a direct role for the lar switch directly regulates the expression of cell-cycle
Menin–UTX switch in regulating cellular senescence (Fig. 4F arrest and senescence–associated genes by direct binding
and G; Supplementary Fig. S17A–S17C). Thus, Menin–MLL to their promoters (80, 81). Consistent with this model,
inhibition engages a tumor-suppressive network that includes we found that Menin is bound to the promoters of the
therapy-induced cellular senescence and is regulated by the cyclin-dependent kinase (CDK) inhibitors Cdkn2c/Ink4c and
MLL3/4–UTX complex. Cdkn2d/Ink4d at basal conditions (82), and that enrich-
ment is decreased upon Menin–MLL inhibition, coinciding
The Enzymatic Domain of UTX Is Dispensable for with their increased expression (Fig. 6A; Supplementary
Tumor-Suppressive Functions in Response to Fig. S21A–S21C). Conversely, we found that UTX binds to
Menin–MLL Inhibition these promoters only in the context of Menin–MLL inhibi-
The UTX protein contains several functional domains, tion, leading to UTX-dependent upregulation of both CDK
including a JmjC histone demethylase domain that cata- inhibitors (Fig. 6A and B). Thus, the Menin–UTX molecular
lyzes the removal of the H3K27me3 modification (72–75). switch regulates the expression of these CDK inhibitors by
To determine the regions of UTX that are necessary for direct chromatin regulation.
treatment-associated UTX-dependent phenotypes, we per- Because the proteins encoded by these two genes are natu-
formed structure–function–rescue experiments in UtxKO-null ral inhibitors of the CDK4 and CDK6 kinases (82), we tested
leukemia cells using lentiviral constructs encoding dual whether pharmacologic inhibition of CDK4/6 could bypass
N-terminal HA- and C-terminal FLAG-tagged truncations of the intrinsic resistance of UtxKO cells to MI-503 while retain-
UTX (Fig. 5A; Supplementary Fig. S18A–S18C). Full-length ing the therapeutic effects of Menin–MLL inhibition on
UTX or a UTX truncation harboring the first 500 amino MLL-FP targets (Fig. 6C). Treatment of MLL-AF9 leukemia
acids (UTX1–500) and lacking the JmjC demethylase domain cells with MI-503 and the FDA-approved CDK4/6 inhibitor
(72–75) was sufficient to resensitize cells to MI-503 treat- palbociclib (83) showed that UtxKO cells were more resistant
ment whereas truncations proximal to the C-terminus were to either MI-503 or palbociclib alone relative to UtxWT cells,
unable to do so (Fig. 5B). Consistent with these cellular likely due to higher basal levels of Cdk6 transcripts (Fig. 6D;
phenotypes, full-length UTX and UTX1–500 were sufficient Supplementary Fig. S21D; refs. 80, 84). However, combined
to rescue UTX-dependent transcriptional phenotypes associ- inhibition of Menin–MLL and CDK4/6 resulted in a synergis-
ated with Menin–MLL inhibition (Fig. 5C), including induc- tic effect (CD = 0.4; ref. 85) on inhibiting cell proliferation to
tion of Menin–UTX target genes (Fig. 5D; Supplementary levels similar to those achieved by MI-503 treatment of UtxWT
Fig. S19). These results demonstrate that an N-terminal cells (Fig. 5D). These results demonstrate that targeting
truncation of UTX, lacking the JmjC demethylase domain pathways regulated by Menin and UTX can produce combi-
and a recently reported intrinsically disordered region (76), natorial therapeutic effects and suggest that the antileukemic
is both necessary and sufficient to drive treatment-associated effects of MI-503 (22) are primarily through reactivation of
tumor-suppressive responses. tumor suppressor pathways and not solely through damp-
To further confirm that UTX uses noncatalytic mecha- ening transcription of MLL-FP targets like Meis1 (refs. 22,
nisms to regulate gene expression in the context of Menin– 25, 27, 51; Supplementary Fig. S5G and S15D). Thus, the
MLL inhibition, we performed ChIP-seq for H3K27me3, a combination of palbociclib with Menin–MLL inhibitors may
modification catalyzed by PRC2 (77) and removed by UTX represent a novel and more effective therapeutic strategy for
(72–75). We found that the genomic redistribution of Menin-dependent cancers (23, 24, 26, 86).
156 | CANCER DISCOVERY JANUARY 2023 AACRJournals.orgSwitch by MLL Complexes Dictates Menin Inhibitor Effects RESEARCH ARTICLE
A C
PCA plot of gene expression
UTX
UTX UTX UTX
PC2: 8% variance
1−500 500−1,000 1,000−1,424 5
0
JmjC
−5
1
93
5
5
8
4
38
09
25
42
10
1,
1,
1,
TPR
−20 0 20
PC1: 82% variance MI-503
− +
B Empty UTX UTX UTX UTX Utx KO
vector full-length 1−500 500−1,000 1,000−1,424
2.0 Empty vector
P = 0.0141
UTX full-length
% cDNA-expressing cells
UTX rescue
1.5 UTX 1−500
P = 0.0003
(relative to d2)
P = 0.0051 P = 0.0368
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
P < 0.0001 P < 0.0001 P = 0.8866 P < 0.0001 P = 0.0005 P = 0.0003
UTX 500−1,000
UTX 1,000−1,424
1.0
0.5
0.0
d2 d6 d2 d6 d2 d6 d2 d6 d2 d6 d2 d6 d2 d6 d2 d6 d2 d6 d2 d6
MI-503 − + − + − + − + − +
D 4
P = 0.0020
P < 0.0001
Relative Cdkn2c expression
P = 0.0135
(normalized to DMSO)
3
2
P = 0.0395 P = 0.0601 P = 0.1598 P = 0.0788
1
0
MI-503 − + − + − + − + − + − + − +
Utx WT Utx KO Empty UTX UTX UTX UTX
vector full-length 1−500 500−1,000 1,000−1,424
Figure 5. Enzymatic domain of UTX is dispensable for its tumor-suppressive functions in response to Menin–MLL inhibition. A, Schematic of UTX
protein. Highlighted are the 8 tetratricopeptide repeats (TPR; 93–385 aa) and the histone demethylase (JmjC) domain (1,095–1,258aa). Three ∼500
amino-acid-long truncations are also represented. B, Growth competition assay in mouse UtxKO MLL-AF9 leukemia cells expressing different RFP-tagged
Utx cDNAs and treated with Menin–MLL inhibitor (MI-503) for 2 or 6 days. The graph shows the relative growth of leukemia cells infected with RFP-tagged
Utx cDNAs measured by flow cytometry (mean ± SEM, n = 3 infection replicates, P values calculated by Student t test). C, Principal component analysis
(PCA) of gene expression data from UtxKO MLL-AF9 leukemia cells expressing different RFP-tagged Utx cDNAs and treated with vehicle (DMSO) or
Menin–MLL inhibitor (MI-503) for 96 hours. D, Cdkn2c expression (mean normalized read counts) from different Utx truncations in mouse MLL-AF9 leuke-
mia cells treated with vehicle (DMSO) or a Menin–MLL inhibitor (MI-503) for 96 hours (mean ± SEM, n = 3 replicates, P values calculated by Student t test).
In Vivo Response to Menin–MLL Inhibitors Is in phase II clinical trials for treatment of patients with
Accompanied by Induction of MLL3/4–UTX- acute leukemia [SNDX-5613 (NCT04067336), KO-539
Dependent Tumor-Suppressive Programs (NCT04065399), and JNJ-75276617 (NCT04811560)]. Nota-
Small-molecule inhibitors of the Menin–MLL interaction bly, SNDX-5613 was recently granted fast-track designation
have shown significant promise in preclinical models of acute by the FDA for the treatment of relapsed/refractory acute
lymphoid and myeloid leukemias (25, 87) and are currently leukemias. To examine whether the Menin–UTX molecular
JANUARY 2023 CANCER DISCOVERY | 157RESEARCH ARTICLE Soto-Feliciano et al.
A 1 kb
[0–0.5] [0–2.3]
MI-503 DMSO MI-503 DMSO MI-503 DMSO MI-503 DMSO
Menin
[0–0.5] [0–2.3]
ChIP-seq
[0–0.9] [0–4.2]
Utx
[0–0.9] [0–4.2]
[0–1.5] [0–11]
Utx WT
[0–1.5] [0–11]
RNA-seq
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
[0–1.5] [0–11]
Utx KO
[0–1.5] [0–11]
Cdkn2c Kri1 Cdkn2d
B Cdkn2c Cdkn2d C
P = 0.8987 P = 0.0014
P < 0.0001
400 P < 0.0001 2,000 MI-503 Palbociclib
Normalized counts
Normalized counts
300 1,500
1
200 P = 0.0377 1,000 Menin UTX TSGs CDK4/6 Senescence
P < 0.0001 e.g., Cdkn2c/d
100 500 2
MLL-FP targets
0 0
M O
03
SO
03
M O
03
M O
03
S
S
S
I-5
I-5
I-5
I-5
M
M
M
M
D
D
M
D
D
Utx WT
Utx KO
Figure 6. Combinatorial targeting of Menin and CDK4/6 overcomes resistance associated with MLL3/4 dysfunction. A, Genome browser representa-
tion of ChIP-seq (top) and RNA-seq (bottom) normalized reads [average reads per kilobase per million mapped reads (RPKM)] for Cdkn2c and Cdkn2d
loci from mouse UtxWT or UtxKO MLL-AF9 leukemia cells treated with vehicle (DMSO) or a Menin–MLL inhibitor (MI-503) for 96 hours. B, Cdkn2c and
Cdkn2d expression (mean normalized read counts) from mouse UtxWT (black) and UtxKO (red) MLL-AF9 leukemia cells treated with vehicle (DMSO) or a
Menin–MLL inhibitor (MI-503) for 96 hours (mean ± SEM, n = 3 replicates, P values calculated by Student t test). C, Proposed model and rationale for
combination therapies based on Menin–MLL and CDK4/6 inhibitors. Our data support a model whereby (1) Menin restricts UTX-mediated transcriptional
activation of tumor suppressor genes (TSG), including Cdkn2c and Cdkn2d, which are natural inhibitors of the CDK4 and CDK6 kinases, which in turn
inhibit cell-cycle arrest and senescence. Our model predicts that CDK4/6 inhibition using palbociclib should boost the anticancer activity of Menin–MLL
inhibitors, which we show induces an MLL3/4–UTX tumor-suppressive axis, by more potently inhibiting these downstream kinases. On the other hand,
Menin is known to be required for the activation of MLL-FP targets like Meis1 and Cdk6 itself to sustain leukemia (2). Our model predicts that combina-
tion therapies based on Menin–MLL and CDK4/6 inhibitors should act synergistically to suppress leukemia proliferation by potently engaging two parallel
pathways that converge on regulation of cell-cycle progression. (continued on following page)
switch described above is operative in patients with leukemia 2) leukemia treated with SNDX-5613 (AUGMENT-101 clini-
treated with Menin–MLL inhibitors, we performed longitudi- cal trial; NCT04065399; Fig. 7A; Supplementary Fig. S22A).
nal RNA-seq analysis of primary AML cells derived from two Consistent with our hypothesis, SNDX-5613 led to con-
patients with NPM1c-mutated (patient 1) and MLL-r (patient comitant suppression of canonical MLL-FP target genes
158 | CANCER DISCOVERY JANUARY 2023 AACRJournals.orgSwitch by MLL Complexes Dictates Menin Inhibitor Effects RESEARCH ARTICLE
D 1.5
P < 0.0001 P < 0.0001 DMSO
P < 0.0001 P < 0.0001
MI-503
P < 0.0001 P = 0.0120
Palbociclib
(relative to DMSO)
1.0 MI-503 +
Cell viability
palbociclib
0.5
Figure 6. (Continued) D, Relative viability of
UtxWT and UtxKO MLL-AF9 leukemia cells treated with
either vehicle (DMSO), a Menin–MLL inhibitor (MI-
503), a CDK4/6 inhibitor (palbociclib), or a combina-
0.0 tion of both inhibitors for 6 days (mean ± SEM, n = 3
Utx WT Utx KO replicates, P values calculated by Student t test).
(e.g., MEIS1) and induction of Menin–UTX targets including skin (16), and CNS (18) malignancies and as an oncogenic
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
CDK inhibitors (e.g., CDKN2C; Fig. 7B–E; Supplementary cofactor in hepatocellular (19) and hematologic cancers
Fig. S22B and S22C). (4, 20). Menin can interact with similar cofactors in disparate
As an orthogonal approach, we analyzed the in vivo response settings, and the biological and molecular basis for these
and resistance of MLL-r acute leukemia patient-derived xeno- ostensibly paradoxical findings has remained unclear. For
grafts (PDX) to VTP-50469 (a close analogue of SNDX-5613; example, Menin functionally cooperates with MLL proteins
Fig. 7F; Supplementary Fig. S23A and S23B; ref. 25). Consist- to activate transcription of the Cdkn1b/p27Kip1 and Cdkn2d/
ent with our model, mice transplanted with an MLL3 wild-type Ink4d CDK inhibitors as a tumor-suppressive mechanism in
PDX showed a potent therapeutic response to VTP-50469 and neuroendocrine tumors and lung cancer (3, 17, 89, 90), yet
transcriptional induction of Menin–UTX targets (Fig. 7G and the same protein–protein interaction is critical for leukemia
H; Supplementary Fig. S24A). Conversely, an MLL3-mutant maintenance (4, 10).
PDX exhibited primary resistance to VTP-50469 and failed Our study sheds light on these paradoxical observations
to induce this gene expression program (Fig. 7I and J; Sup- by revealing a functional interaction between the mamma-
plementary Fig. S24A), linking the induction of Menin–UTX lian histone methyltransferase complexes MLL1–Menin and
targets to preclinical drug response. Thus, gene expression MLL3/4–UTX and, in doing so, challenging the paradigms
programs regulated by MLL1–Menin and MLL3/4–UTX com- that (i) these complexes are canonically restricted to cer-
plexes are operative in patients with AML and PDX models tain genomic compartments (some exceptions have been
treated with orally bioavailable Menin–MLL inhibitors that identified; refs. 91–93), and (ii) activation of gene promot-
are currently under clinical investigation. ers is strictly associated with H3K4me3 deposition, whereas
To test whether CDK4/6 inhibition can overcome the H3K4me1 enrichment at promoters is believed to be asso-
blunted induction of endogenous CDK inhibitors in the ciated with repression (Supplementary Fig. S25; refs. 54,
context of Menin inhibitor resistance, we treated mice har- 94). In leukemia cells harboring MLL fusions, Menin–MLL1
boring the MLL3-mutant AML PDX with VTP-50469 and represses the expression of a tumor-suppressive network that
palbociclib (Fig. 7K). Although single treatment with Menin involves Cdkn2c/Ink4c and Cdkn2d/Ink4d by binding to gene
or CDK4/6 inhibitors led to a minor decrease in leukemia promoters and depositing H3K4me3. Disruption of Menin–
burden in the bone marrow of recipient mice after 10 days MLL1 complexes using genetic or pharmacologic approaches
of treatment, combination treatment with VTP-50469 and triggers a molecular switch that leads to de novo recruit-
palbociclib induced significant leukemia regression (Fig. 7K ment of the MLL3/4–UTX complex to promoters of tumor-
and L). Moreover, RNA-seq from human leukemia cells iso- suppressive genes that were previously bound by Menin–
lated from these animals showed induction of cell-cycle MLL. This switch also leads to a UTX-dependent increase in
arrest and senescence–associated gene expression signa- activation-associated histone modifications and gene expres-
tures in mice treated with both VTP-50469 and palbociclib sion (70, 78). Importantly, these phenotypes are independent
(Fig. 7M; Supplementary Fig. S24B and S24C). Altogether, of UTX catalytic activity, as the first 500 amino acids of UTX,
these data provide preclinical evidence for the feasibility which lack the histone demethylase domain, are sufficient
and efficacy of combined Menin and CDK4/6 inhibition to to rescue UTX deficiency. This finding is particularly inter-
overcome the blunted induction of senescence-associated esting given a recent study suggesting that UTX requires a
programs in patient-derived leukemias resistant to Menin much bigger protein fragment to drive tumor-suppressive
inhibitor monotherapy. activity and transcriptional regulation via phase separation
mechanisms (76). Instead, our study demonstrates that UTX
does not require a core intrinsically disordered region to
DISCUSSION drive tumor-suppressive responses in the context of Menin–
The chromatin adapter protein Menin exhibits context- MLL inhibition. Thus, UTX uses noncatalytic mechanisms
specific functions in different tissues, acting as a tumor to regulate gene expression programs that affect response to
suppressor gene in neuroendocrine (14, 15), lung (17, 88), epigenetic therapies.
JANUARY 2023 CANCER DISCOVERY | 159RESEARCH ARTICLE Soto-Feliciano et al.
A B Patient 1: C
SDNX-5613 Patient 1: AML cells Menin–UTX targets Patient 1: CDKN1A
clinical trial: Days: 0 3 7 14 30 2 CDKN2B
0d
CDKN2C
Log2 fold change
3d
0 HOXA9
7d CDK6
14 d −2
PBX3
30 d
−4 MEIS1
101 102 103 104 0 5 10 15
CD117 (cKIT) Row max Row min Days of treatment
D E F
Patient 2: Patient 2:
Menin–UTX targets CDKN3
2 MLL3-WT
Days: 0 3 10 CDKN1B 4 weeks VTP-50469
MLL3-Mut
Log2 fold change
(0.1% in rodent diet)
Downloaded from http://aacrjournals.org/cancerdiscovery/article-pdf/13/1/146/3238184/146.pdf by guest on 30 May 2023
1 CDKN1A
Engraftment
HOXA7
0
CDK6
−1 Blood monitoring
MEF2C Transplant of AML PDX
−2 MEIS1 into NSG mice
0 2 4 6 8 10 Every 7 days:
Row max Row min Days of treatment harvest 3 mice/group
G MLL-r, MLL3-WT H MLL-r, MLL3-WT I MLL-r, MLL3-Mut J MLL-r, MLL3-Mut
Menin–UTX targets VTP-50469 Menin–UTX targets
VTP-50469
80 1.0 80 1.0 n.s
P < 0.0001
% hCD45+ cells in PB
% hCD45+ cells in PB
60 0.5 60 0.5
z-score
z-score
40 40
0.0 0.0
Vehicle Vehicle
20 20
VTP-50469 −0.5 VTP-50469 −0.5
0 0
0 10 20 30 40 50 −1.0 0 10 20 30 40 50 −1.0
Days after treatment Vehicle VTP-50469 Days after treatment Vehicle VTP-50469
Figure 7. In vivo response to Menin–MLL inhibition is accompanied by induction of MLL3/4–UTX-dependent tumor-suppressive programs. A, Longitu-
dinal flow cytometry analysis showing the fraction of CD45lo, cKIT+ leukemia cells in the peripheral blood of an NPM1c-mutant patient (patient 1) during
cycle 1 of Menin–MLL inhibitor (SDNX-5613) treatment as part of the AUGMENT-101 clinical trial (NCT04065399). B, Temporal gene expression changes
for Menin–UTX targets in FACS-sorted leukemia blast cells isolated from patient 1 as part of the AUGMENT-101 clinical trial (NCT04065399). Heat
map shows all Menin–UTX targets that are differentially expressed at day 14 versus day 0 of treatment cycle 1. C, Temporal expression levels of genes
involved in cell-cycle arrest and senescence (CDKN1A, CDKN2B, and CDKN2C) and Menin–UTX targets (HOXA9, CDK6, PBX3, and MEIS1) in leukemia blast
cells isolated from patient 1 treated with SDNX-5613. D, Temporal gene expression changes for Menin–UTX targets in FACS-sorted leukemia blast cells
isolated from patient 2 treated with SDNX-5613 as part of the AUGMENT-101 clinical trial (NCT04065399). Heat map shows all Menin–UTX targets that
are differentially expressed at day 10 versus day 0 of treatment cycle 1. E, Temporal expression levels of genes involved in cell-cycle arrest and senescence
(CDKN1A, CDKN1B, and CDKN3) and MLL-FP targets (HOXA7, CDK6, MEF2C, and MEIS1) in leukemia blast cells isolated from patient 2 treated with
SDNX-5613. F, Schematic of in vivo treatment experiments using genetically defined AML PDXs. NSG mice were transplanted with either MLL3 wild-type
(MLL3-WT) or MLL3-mutant (MLL3-Mut) AML PDXs and, upon disease engraftment, were randomized into Menin–MLL inhibitor (VTP-50469) or normal
chow for a duration of 4 weeks. Disease progression was monitored weekly by bleeding, and AML cells were sorted 7 days after initiation of treatment
using magnetic mouse cell depletion from the bone marrow of animals to perform RNA-seq. G, Disease progression as measured by the percentage of
human CD45+ cells in the peripheral blood (PB) of mice harboring MLL3-WT leukemia treated with vehicle (gray) or a Menin–MLL inhibitor (VTP-50469,
blue). H, Box plot denoting gene expression changes of Menin–UTX targets in AML cells isolated from mice harboring MLL3-WT leukemia treated with
vehicle (gray) or a Menin–MLL inhibitor (VTP-50469, blue). I, Disease progression as measured by the percentage of human CD45+ cells in the peripheral
blood of mice harboring MLL3-mutant leukemia treated with vehicle (gray) or a Menin–MLL inhibitor (VTP-50469, blue). J, Box plot denoting gene expres-
sion changes of Menin–UTX targets in AML cells isolated from mice harboring MLL3-mutant leukemia treated with vehicle (gray) or a Menin–MLL inhibitor
(VTP-50469, blue). (continued on following page)
Our findings have important translational implications, Menin–MLL1 complex maintains repression of a set of target
as they shed light on a novel noncanonical gene expres- genes under steady-state conditions, which only becomes
sion program regulated by Menin. We demonstrate that the activated upon Menin inhibitor–mediated displacement of
160 | CANCER DISCOVERY JANUARY 2023 AACRJournals.orgYou can also read

















































