Woodpecker foraging activity in oak-dominated hill forests in Hungary
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Ornis Hungarica 2021. 29(1): 82–97.
DOI: 10.2478/orhu-2021-0006
Woodpecker foraging activity in oak-
dominated hill forests in Hungary
Mariann Komlós1, Zoltán Botta-Dukát2, Dániel Winkler1, Réka
Aszalós2 & Gábor Ónodi2,3*
Received: March 14, 2021 – Revised: May 23, 2021 – Accepted: May 25, 2021
Komlós, M., Botta-Dukát, Z., Winkler, D., Aszalós, R. & Ónodi, G. 2021. Woodpecker forag-
ing activity in oak-dominated hill forests in Hungary. – Ornis Hungarica 29(1): 82–97. DOI:
10.2478/orhu-2021-0006
Abstract We documented the foraging activities of woodpeckers on selected trees in an established conservation-
oriented management study in five oak-dominated forests in Hungary. We examined the tree species preference
of woodpeckers as a group and the impact of specific tree characteristics on the habitat use of woodpeckers.
We estimated the percentage of visible foraging signs on the trunks and upper limbs of selected trees through
the winter and early spring of 2019–2020. Based on the Jacobs’ index, woodpeckers preferred oak species for
foraging and most foraging signs were on limbs rather than trunks. Foraging signs on trunks were more frequent
on those of larger diameters and greater heights. It was also found that the lower the tree, the greater the effect
of its diameter on the occurrence of signs.
Keywords: woodpecker ecology, woodpecker foraging signs, conservation, nature conservation management
Összefoglalás Kutatásunkban hazai harkályfajok táplálkozási nyomait mértük fel egy természetvédelmi erdőke-
zelési kutatás alapállapot felméréseként tölgy dominálta gazdasági erdőkben. Vizsgáltuk a harkályok fafaj-prefe-
renciáját, valamint a fák egyes jellemzőinek hatását a harkályok élőhely-használatára. 2019–2020 tele és kora ta-
vasza között mértük fel a kijelölt fákon megfigyelhető táplálkozási nyomok százalékos előfordulását a fák törzsén
és a lombkoronában. Eredményeink alapján a harkályfajok leginkább a különböző tölgyfajokat preferálták, ezen
belül is legnagyobb mértékben a kocsánytalan tölgyet, emellett pedig nagyobb eséllyel találtunk táplálkozási nyo-
mokat az ágakon, mint a fák törzsén. A detektált táplálkozási nyomok a nagyobb törzsátmérőjű és magasabbak fá-
kon voltak gyakoribbak. Minél alacsonyabb a fa, annál nagyobb hatása van a mellmagassági átmérőnek a táplál-
kozási nyomok előfordulására, a felsőbb magassági kategóriákban az átmérő hatása mérsékeltebb.
Kulcsszavak: harkály ökológia, harkály táplálkozási nyomok, természetvédelem, természetvédelmi erdőkezelés
1
Institute of Wildlife Management and Wildlife Biology, University of Sopron, 9400 Sopron, Bajcsy-Zsilinszky
utca 4., Hungary
2
Institute of Ecology and Botany, Centre for Ecological Research, 2163 Vácrátót, Alkotmány út 2–4., Hungary
3
Department of Botany and Biodiversity Research, University of Vienna, 1030 Vienna, Rennweg 14, Austria
*corresponding author, e-mail: onodi.dendrocopos@ecolres.hu
Introduction
The ecological impact of woodpecker species in wooded ecosystems is important for numerous
reasons. They are considered umbrella species because through their presence they can
support many other species (Melletti & Penteriani 2003, Garmendia et al. 2006, Puverel et al.
2019). They provide nesting and roosting site for various cavity-dwelling species, including
invertebrates, mammals and birds such as tits (Paridae), flycatchers (Ficedula spp.), nuthatches
(Sitta spp.), treecreepers (Certhia spp.) and some owls (Strigidae) and ducks (Anatidae)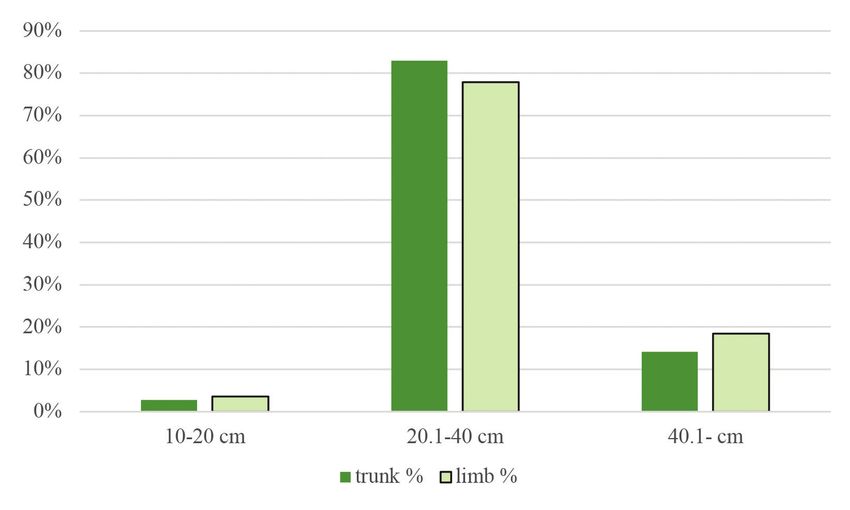
M. Komlós, Z. Botta-Dukát, D. Winkler, R. Aszalós & G. Ónodi 83 (Gorman 2004). Certain woodpeckers can also be regarded as keystone species (Mikusiński et al. 2001, Virkkala 2006), for example, Black Woodpecker (Dryocopus martius) in Europe (Gorman 2011, Puverel et al. 2019) and Red-cockaded Woodpecker (Dryobates borealis) in North America (Jusino et al. 2015). The most abundant European species, the Great Spotted Woodpecker (Dendrocopos major), is often categorized as a keystone species, especially in sub- optimal habitats, where there is invariably a lack of natural cavities (Pasinelli 2007). However, this species is a major nest predator of cavity-nesting songbirds and therefore, its keystone role is sometimes considered as controversial (Wesolowski 2007, Ónodi & Winkler 2016). Woodpeckers are particularly suitable as indicator organisms (Virkkala 2006, Drever et al. 2008). First, most of them are year-round resident and mostly easily detectable (Lõhmus et al. 2010). Second, they have large home ranges (e.g. Bocca et al. 2007, Charman et al. 2010, Campion et al. 2020), hence they can provide information on large areas. Third, this is the only bird family in the Western Palearctic that can forage for insects within the inner layers of wood, which are threatened with extinction in intensively managed regions so that they can be monitored relatively easily (Lõhmus et al. 2010). They are also highly susceptible to habitat change (Mikusiński et al. 2001, Thompson et al. 2003, Mikusiński 2006). Finally, they provide identifiable foraging signs that are often specific to the group (Gorman 2015). Many studies on the habitat and nesting requirements of woodpecker and cavity-nesting species in general have been published (Wesolowski & Tomialojc 1986, Hardersen 2004, Pasinelli 2007, Kosinski et al. 2017). There has been less focus on foraging habitat needs and on foraging signs. This is especially true for species-specific observations, especially in managed, commercial stands (Czeszczewik 2009, St-Amand et al. 2018, Aszalós et al. 2020). Woodpeckers that forage on bark may choose trees with different characteristics for foraging than they do for nesting, and thus, habitats appropriate for nesting may fail to include suitable foraging sites (Swallow et al. 1988, St-Amand et al. 2018). The motivation for studying artificially created snags usually relates to nesting (Brandeis et al. 2002, Kilgo & Vukovich 2014, Barry et al. 2018). Few studies examine this subject in the context of foraging needs (Aulén 1991, Farris & Zack 2005, Arnett et al. 2010, Aszalós et al. 2020). In this study, the following questions were posed. Which tree species are preferred for foraging by woodpeckers in the studied oak forests? Does the diameter or height of trees affect how they are used? Which attributes of the studied trees influence the presence of woodpecker foraging signs? What determines, and to what extent, the number of foraging signs observed on the studied trees? Materials and Methods Study sites This study was part of a detailed, conservation-oriented management project (LIFE 4 Oak Forests Project, LIFE16NAT/IT/000245), the purpose of which was to structurally enrich certain oak-dominated woodlands in Hungary through various management techniques. Within this framework, we designated ten 80×80 m (0.64 ha) square plots in 5 study sites,
84 ORNIS HUNGARICA 2021. 29(1)
in which artificial standing and downed deadwood and the opening of canopy gaps will be
created in the near future. The present study was a baseline research carried out prior to the
planned management. Therefore, all the trees surveyed would be selected for management
activities, and, in addition, control trees that would not be managed, were also selected. In the
forthcoming winter of 2021–2022, there will be a follow-up survey of both the managed and
control trees. Therefore, all trees selected for this study were living specimens at the time of
data collection. On 10 square plots in the 5 study sites, we measured a total of 1,471 individual
trees. In these study plots, we surveyed all trees with diameters of more than 10 cm at breast
height (DBH). Table 1 summarizes the data of the trees measured (species, height, diameter).
Table 1. Numbers, mean diameter, and height of the surveyed trees by species (QC – Quercus
cerris, RP – Robinia pseudoacacia, C – Crataegus sp., CB – Carpinus betulus, QR – Quercus
robur, QP – Quercus petraea, AC – Acer campestre, FE – Fraxinus excelsior, QPU – Quercus
pubescens, FO – Fraxinus ornus, QRU – Quercus rubra)
1. táblázat A vizsgált faegyedek száma, átlagos átmérője és magassága fafajok szerint bemutatva (QC
– Quercus cerris, RP – Robinia pseudoacacia, C – Crataegus sp., CB – Carpinus betulus, QR –
Quercus robur, QP – Quercus petraea, AC – Acer campestre, FE – Fraxinus excelsior, QPU –
Quercus pubescens, FO – Fraxinus ornus, QRU – Quercus rubra)
tree species QP QR QC QPU QRU CB FE FO AC RP C
number of individuals 616 71 750 7 1 11 1 1 11 1 1
mean DBH_cm 28.1 37.09 33.24 32.2 32.5 22.87 15.3 15.6 13.7 30.6 12.6
mean height_m 20.3 23.34 23.5 18.56 25.86 17.82 19.33 12.05 13.11 19 10.67
Figure 1. Location of the study sites
1. ábra Vizsgálati területek elhelyezkedése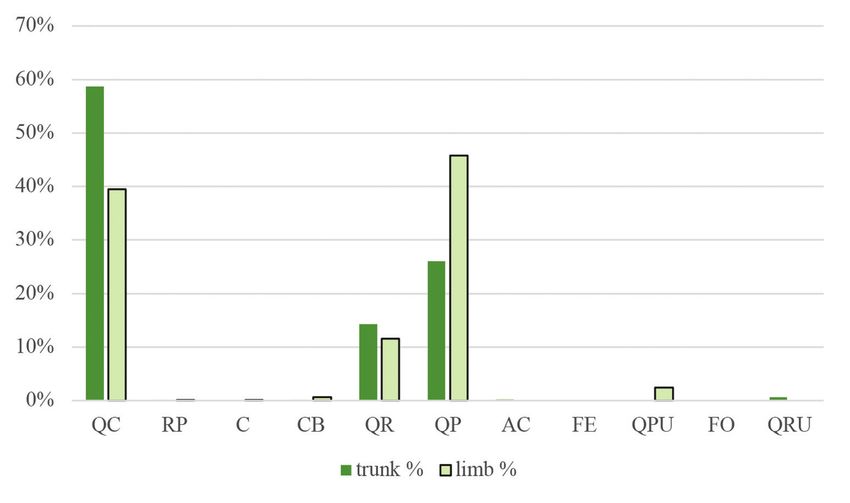
M. Komlós, Z. Botta-Dukát, D. Winkler, R. Aszalós & G. Ónodi 85 All study areas were within hill ranges in north-east Hungary (Figure 1). The first study site is located in the Duna-Ipoly National Park, specifically in the Börzsöny Hills near the settlement of Nagyoroszi (65°08’20”N-29°62’40”E, 250 m a.s.l, 10–15° slope, 80–99 years old stands). The studied forest was dominated by sessile oak (Quercus petraea) – Turkey oak (Quercus cerris) and hornbeam (Carpinus betulus). The second and third sites were situated in the Cserhát Hills, which are part of the East-Cserhát Landscape Protection Area. The second site, near the settlement of Buják (68°84’36”N-28°55’10”E, 300 m a.s.l, 10° slope, 80–99 years old stands), is a Turkey oak-pedunculate oak (Quercus robur) forest with a high scrub layer. The other site, in the vicinity of Garáb (69°48’84”N-29°32’60”E, 500 m a.s.l, 20–25° slope), is a 60–79-year-old deciduous woodland with sessile oak, hornbeam, beech (Fagus sylvatica) and Turkey oak. The final two study sites were located in the Bükk Hills (Bükk National Park) near the settlements of Cserépfalu (76°32’78”N-29°29’00”E, 250-350 a.s.l., 5–10° slope, 80–99-year-old stands) and Bükkzsérc (75°67’58”N-29°23’50”E, 350– 450 m a.s.l., 10–15° slope, 40–59 and 80–99-year-old stands). Both sites are predominantly covered by Turkey oak-sessile oak forests. Woodpecker foraging activity surveys In the course of previous point count surveys, we detected the presence of the Great (Dendrocopos major), Middle (Leiopicus medius), Lesser Spotted (Dryobates minor), Black (Dryocopus martius), Eurasian Green (Picus viridis) and Grey-headed Woodpeckers (Picus canus) within the study sites. In most cases, it was not possible to accurately distinguish between the foraging marks that the different species made, as most are not diagnostic (Gorman 2015). From the species present in the study areas, only the black woodpecker’s foraging marks are generally identifiable to species, owing to its large bill size. Therefore, we decided not to attempt to identify signs and marks to species level, but rather the wood layers that the birds accessed. The percentage of visible foraging signs on the trunks and limbs of the surveyed trees were estimated using binoculars and naked eyes through the winter and early spring of 2019–2020. The survey differentiated four depth categories for presumed woodpecker foraging signs on trees: marks on the bark, bark scaling, excavation marks in the sapwood, excavation marks in the heartwood. ‘Marks on the bark’ means that birds had searched for prey by only superficial pecking. ‘Bark scaling’ means that birds had peeled off sections of bark and foraged beneath it. We also identified deeper excavation marks: ones that penetrated only into the sapwood and others that entered deeper into the heartwood. Heartwood was recognized by its distinctive (usually reddish) colour. We also differentiated between marks on trunks (with an estimated diameter greater than 15 cm) and on limbs (with an estimated maximum diameter of 15 cm). Data analyses We calculated the frequency and distribution of woodpecker foraging signs according to tree species and categories of diameter at breast height (10–20; 20.1–40 and above 40 cm) and tree height (0.5–15; 15.1–25, and above 25.1 m). Jacobs’ preference index values (Jacobs
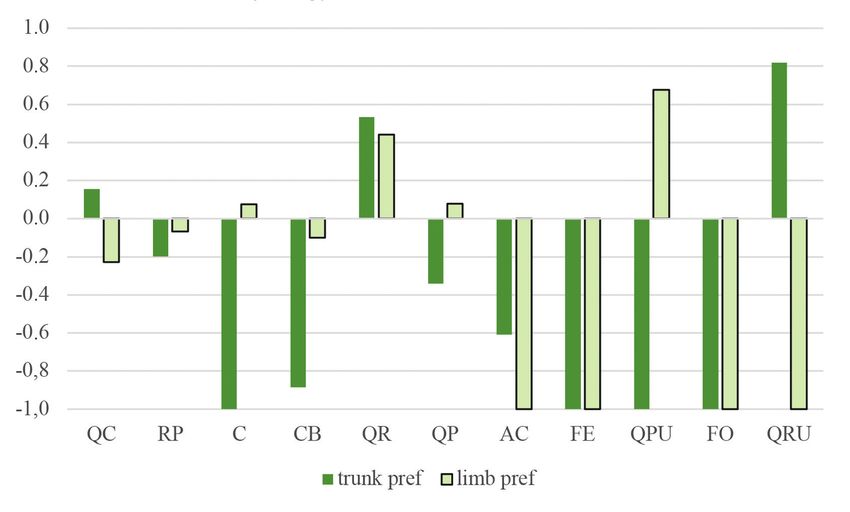
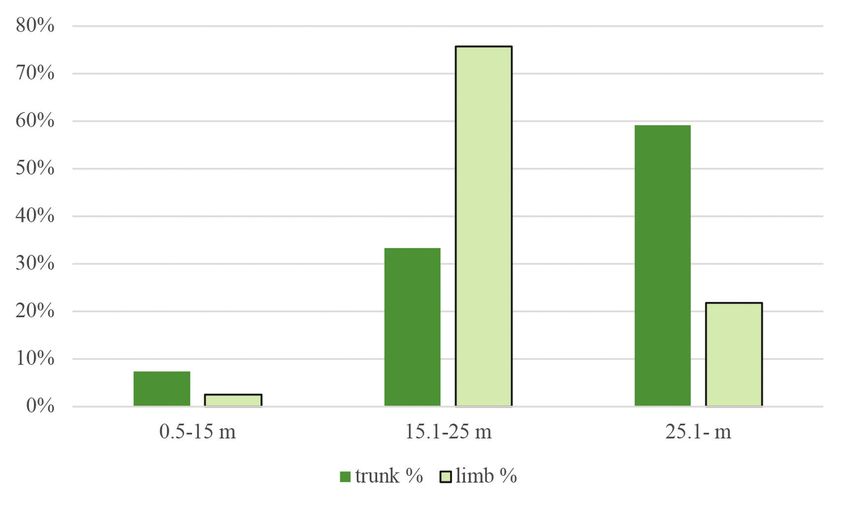
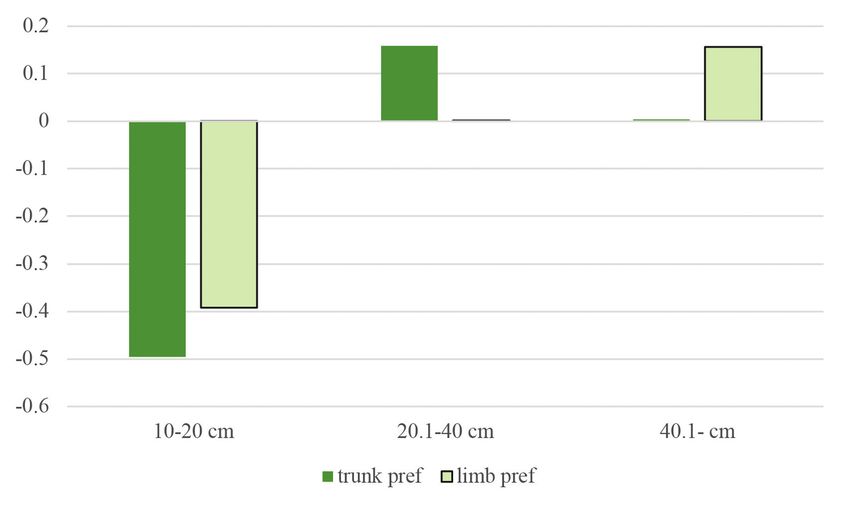
86 ORNIS HUNGARICA 2021. 29(1) 1974) were calculated using frequency data from a previous vegetation survey in the study sites. This index represents a –1 to +1 scale from avoidance to the examined variable’s preference, respectively. We used a mixed linear model with zero-inflated beta distribution and logit link. Since beta distribution is defined on interval [0, 1], the zero-inflation part models the probability of occurrence of foraging signs. The conditional part models the area of signs when they occur. Our models included tree species, DBH, and height as fixed effects and research plots as a random factor. Main effects were tested by Type II Wald’s chi-square test. Because of the limited data due to the limited availability of secondary tree species, only the pedunculate oak, sessile oak, and Turkey oak were included in the model. Analyses were done in the R statistical environment (version 4.0.2 R Development Core Team 2020) using the glmmTMB (version 1.0.2.1, Brooks et al. 2017), emmeans (version 1.5.4, Lenth 2021), multcomp (version 1.4-16, Hothorn et al. 2018) and effects (4.2-0, Fox 2003, Fox & Weisberg 2018) packages. Results Foraging preferences From the 1,471 individual trees examined, 798 (54.24%) had some type of foraging marks upon them. In the study areas, woodpeckers foraged mainly upon Turkey oaks and sessile oaks (Figure 2). The order of frequency of the tree species was the following: Turkey oak (51%), sessile oak (42%), pedunculate oak (5%). Other species had less than 1% of frequency: black locust (Robinia pseudoacacia), hawthorn (Crataegus sp.), hornbeam, common ash (Fraxinus excelsior), manna ash (Fraxinus ornus), red oak (Quercus rubra), downy oak (Quercus pubescens) and field maple (Acer campestre). The most preferred tree species in terms of both trunk and limb use was the pedunculate oak. When just trunk usage was examined, the most preferred species was the red oak, and when only limbs were considered, mainly downy oak was used. However, it must be mentioned that both red and downy oaks were hardly present in the study areas, thus, these data are biased to 1–2 individual trees (Figure 3). The Turkey oak can also be mentioned for trunk use and sessile oak and hawthorn for limb use as tree species utilized at a moderate level. As reflected in the negative Jacobs’ index values, woodpeckers avoided common ash, manna ash, hornbeam, and field maple. The percentage of coverage of foraging signs was the highest on trees falling in the middle DBH category. At the same time, on trees with a DBH under 20 cm, woodpecker foraging marks were scarcely present (Figure 4), as is also reflected in the preference index (Figure 5). In the case of tree height (Figure 6), the 15.01–25 m category was the most frequent, followed by the category of >25 m. The least utilized trees belonged to the lowest height category (below 15 m). Regarding both trunks and limbs, woodpeckers preferred the highest trees and avoided the medium-height category (Figure 7).
M. Komlós, Z. Botta-Dukát, D. Winkler, R. Aszalós & G. Ónodi 87
Figure 2. Percentage distribution of woodpecker foraging signs on the trunks and limbs on each tree
species. (For abbreviations of tree species see the legend of Table 1)
2. ábra A felmért harkály táplálkozási nyomok százalékos megoszlása a törzseken és az ágakon
fafajok szerint. (A fafajok rövidítését lásd az 1. táblázat jelmagyarázatánál)
Figure 3. Jacobs’ preference index of woodpecker foraging signs on trunks and limbs on each tree
species. (For abbreviations of tree species see the legend of Table 1)
3. ábra Jacobs preferencia értékek a törzseken és az ágakon fafajok szerint. (A fafajok rövidítését
lásd az 1. táblázat jelmagyarázatánál)88 ORNIS HUNGARICA 2021. 29(1)
Figure 4. Percentage distribution of woodpecker foraging signs on trunks and limbs according to
breast height categories
4. ábra A felmért harkály táplálkozási nyomok százalékos megoszlása a törzseken és az ágakon a
megadott átmérőkategóriák szerint
Figure 5. Jacobs’ preference index of woodpecker foraging signs on trunks and limbs according to
diameter at breast height categories
5. ábra Jacobs preferencia értékek a törzseken és az ágakon a megadott átmérőkategórák szerintM. Komlós, Z. Botta-Dukát, D. Winkler, R. Aszalós & G. Ónodi 89
Figure 6. Percentage distribution of woodpecker foraging signs on trunks and limbs according to
height categories
6. ábra A felmért harkály táplálkozási nyomok százalékos megoszlása a törzseken és az ágakon a
megadott magassági kategóriák szerint
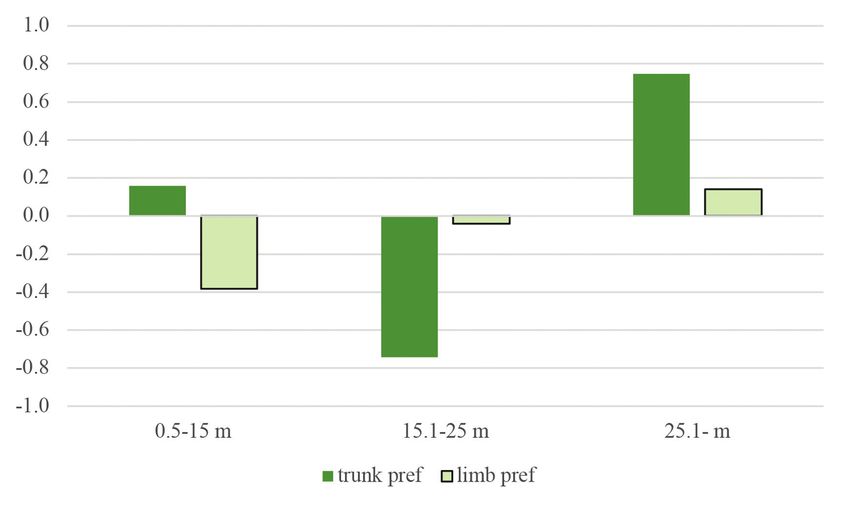
Figure 7. Jacobs’ preference index of woodpecker foraging signs on trunks and limbs according to
defined height categories
7. ábra Jacobs preferencia értékek a törzseken és az ágakon a megadott magassági kategóriákban90 ORNIS HUNGARICA 2021. 29(1)
Foraging sign distributions (glmmTMB)
Regarding trunks, tree species significantly affected the occurrence of foraging signs. At the
same time, interaction effects of the variables DBH and height also proved to be significant.
On limbs, the effects of both tree species, DBH, and height were significant (pChisq)
component=zi
tree species 12.08 2 0.0024
DBH 1.89 1 0.1696
Height 3.50 1 0.0615
trunk
species:DBH 0.85 2 0.6536
species:Height 1.57 2 0.4565
DBH:Height 11.13 1 0.0009
tree species 28.844 3M. Komlós, Z. Botta-Dukát, D. Winkler, R. Aszalós & G. Ónodi 91
Table 4. Estimated probability of the presence of foraging signs and their relative amount when pres-
ent on the different tree species. (For abbreviations of tree species see the legend of Table 1)
4. táblázat A táplálkozási nyomok előfordulásának és relatív kiterjedésének becsült valószínűségi érté-
ke, amennyiben előfordul a vizsgált nyom, különböző fafajokra vizsgálva. (A fafajok rövidí-
tését lásd az 1. táblázat jelmagyarázatánál)
Trunk Limb Limb (relative area of sign,
Species
(presence of sign, %) (presence of sign, %) when present, %)
QC 9.31 (2.18–32.1)b 37.3 (31.5–43.5)a 6.94 (6.18–7.78)a
QR 4.23 (0.85–18.6)a 55.8 (40.8–69.8)b 12.11 (9.28–15.65)b
QP 16.92 (4.25–48.3)c 55.1 (48.4–61.6)b 7.90 (7.12–8.76)a
When a foraging sign occurred on a trunk, its area was not related to any of the studied
predictors (tree species, DBH, height), while on limbs, only the tree species used had a
significant effect on the area of signs (Table 3). On trunks, the presence of foraging signs
was most frequent on sessile oak, while on limbs, both sessile oak and pedunculate oak
shared almost the same probability of presence. Regarding the relative area of signs, their
extent, when present, was the highest on the limbs of pedunculate oak (Table 4).
From the four different height categories, the following conclusions were drawn. The
lower the tree, the greater the effect of diameter on the occurrence of signs. In greater height
categories, the impact of diameter was moderate (Figure 8). The results also showed that
signs were more frequent on trunks with greater diameters and heights (Figure 9–10).
Figure 8. Interaction of the variables DBH and height with regards to the presence of foraging data
on trunks
8. ábra A mellmagassági átmérő és a magasság interakciója a törzseken előforduló táplálkozási
nyomok tekintetében92 ORNIS HUNGARICA 2021. 29(1)
Figure 9. Linear correspondence (logit distribution) of DBH and the probability of presence
of foraging signs on limbs
9. ábra A mellmagassági átmérő és a táplálkozási nyomok előfordulásának lineáris
korreszpondencia vizsgálata a vizsgált ágak vonatkozásában
Figure 10. Linear correspondence (logit distribution) of height and the probability of
presence of foraging signs on limbs
10. ábra A magasság és a táplálkozási nyomok előfordulásának lineáris korreszpondencia
vizsgálata a vizsgált ágak vonatkozásábanM. Komlós, Z. Botta-Dukát, D. Winkler, R. Aszalós & G. Ónodi 93 Discussion In the present study, we assessed managed, commercial forests, which are generally considered to be of little importance for nature conservation and biodiversity. Neither the structure nor the species composition is as rich in such forests as it is in natural ones (Tomiałojć et al. 1984, Kraigher et al. 2003, Bobiec et al. 2005, Christensen et al. 2005). Data from Central European managed deciduous forests are scarce (Mazgajski 1998, Kosinski 2006, Kosinski & Kempa 2007), compared to the number of similar research projects that have been carried out in so-called primeval forests such as Białowieża in Poland (Wesołowski et al. 2010, Walankievicz et al. 2011, Czeszczewik et al. 2015). Possibilities for comparison were, and are, therefore limited. Nevertheless, it is worth comparing our results with those that are available from unmanaged forests. Similar to the observations by Kosinski (2006) and Pasinelli (2007), our results confirm that in mixed forests in Europe, woodpeckers mainly prefer oak species. This is due to the less diverse nature of the studied managed forests, primarily composed of oak species with a lower proportion of secondary tree species. However, woodpeckers, especially Great and Middle Spotted Woodpeckers, forage mainly on oak trees that have a fissured bark rich in arthropod fauna, providing an important food source even in forests where oaks are less dominant (Török 1990, Pasinelli & Hegelbach 1997, Kruszyk 2003, Kosinski 2006). Our results showed a relatively high preference for red oak. Red oak is an adventive species in Hungary, and thus, few insect ‘pest’ species (bark and wood-boring insects) use it (Rédei et al. 2011, Keserű et al. 2017), thus this phenomenon is noteworthy. In this study, woodpeckers utilized hornbeam only moderately. Studies in Białowieża showed that woodpeckers prefer to nest and forage on hornbeam, while other researchers stated they avoid this species (Hardersen 2004). The reason for this deviation could be that in primeval forests, the hornbeam trees used are older, more decayed, sizable, and hence often have substantial amounts of deadwood (Kosinski & Kempa 2007). According to Roberge et al. (2008), the basal area/ha is one of the determining factors for foraging Middle Spotted and Great Spotted Woodpeckers, since they prefer older and wider trees. In the framework of this baseline survey, we studied foraging on living trees, a subject which is generally uncommon in woodpecker-related research. Nonetheless, some previous studies have indicated that Great Spotted Woodpeckers are often associated with living trees (Török 1990, Farris & Zack 2005, Pasinelli 2007, Ónodi & Csörgő 2014, Ónodi & Winkler 2016, Kosiński et al. 2017). Many papers discuss the relationship between snags and woodpeckers and the importance of deadwood to them (Angelstam et al. 2003, Farris & Zack 2005, Löhmus et al. 2010, Kosinski et al. 2017). Deadwood is usually scarce in managed forests, so our results might highlight some of the traits on trees that woodpeckers prefer if no deadwood material is available. We found stronger correlations for the canopy than for trunks, which may have been related to dead limbs being available in higher amounts in the canopy through the decaying processes initiated by wood-rotting fungi via mechanical damage e.g., windbreak (Gibbons & Lindenmayer 2002). Dead branches are crucial for, among other species, the Lesser Spotted Woodpecker (Roberge et al. 2008, Charman et al. 2010).
94 ORNIS HUNGARICA 2021. 29(1)
Nowadays, the vast majority of forests worldwide are managed in some way. To have
diverse and stable ecosystems, it is often necessary to improve their natural characteristics
through nature conservation management. To achieve this aim, it is important to diversify
forests both structurally and compositionally. Conservationists can use various methods to
create deadwood, new microhabitats and open canopy gaps, which can all help increase the
diversity and abundance of a wide range of taxa: plants, fungi, saproxylic beetles, birds and
mammals. Currently, such management activities are mostly employed in North America
(Swallow et al. 1988, Hallett et al. 2001, Arnett et al. 2010, Kilgo & Vukovich 2014, Weiss
et al. 2017, Sandström et al. 2019), although they are on the increase in Europe (Aulén 1991,
Aszalós et al. 2020).
Acknowledgements
We would like to express our gratitude to Péter Molnár for his support. We would like to
thank Gerard Gorman for his practical help with woodpecker signs in the field. Our research
was funded by the LIFE 4 Oak Forests project (LIFE16NAT/IT/000245), the ÚNKP-19-
3-I-33 New National Excellence Program of the Ministry for Innovation and Technology
and the Austrian Agency for International Cooperation in Education and Research (OeAD-
GmbH), Mobility Programmes, Bilateral and Multilateral Cooperation (MPC) (Scholarship
of the Scholarship Foundation of the Republic of Austria, Postdocs ICM-2020-00204).
References
Angelstam, P. K., Bütler, R., Lazdinis, M., Mikusiński, G. & Roberge, J. M. 2003. Habitat thresholds for focal
species at multiple scales and forest biodiversity conservation. – Dead wood as an example. – Annales
Zoologici Fennici 40(6): 473–482.
Arnett, E. B., Kroll, A. J. & Duke, S. D. 2010. Avian foraging and nesting use of created snags in intensively-
managed forests of western Oregon, USA. – Forest Ecology and Management 260(10): 1773–1779. DOI:
10.1016/j.foreco.2010.08.021
Aszalós, R., Szigeti, V., Harmos, K., Csernák, S., Frank, T. & Ónodi, G. 2020. Foraging activity of
woodpeckers on various forms of artificially created deadwood. – Acta Ornithologica 55(1): 63–76. DOI:
10.3161/00016454AO2020.55.1.007
Aulén, G. 1991. Increasing insect abundance by killing deciduous trees: A method of improving the food
situation for endangered woodpeckers. – Holarctic Ecology 14(1): 68–80. DOI: 10.1111/j.1600-
0587.1991.tb00635.x
Barry, A. M., Hagar, J. C. & Rivers, J. W. 2018. Use of created snags by cavity-nesting birds across 25 years.
– Journal of Wildlife Management 82(7): 1376–1384. DOI: 10.1002/jwmg.21489
Bobiec, A., Gutowski, J. M., Laudenslayer, W. F., Pawlaczyk, P. & Zub, K. 2005. The afterlife of a tree. –
WWF Poland, Warsawa-Hajnóvka
Bocca, M., Carisio, L. & Rolando, A. 2007. Habitat use, home ranges and census techniques in the Black
Woodpecker Dryocopus martius in the Alps. – Ardea 95: 17–29. DOI: 10.5253/078.095.0103
Brandeis, T. J., Newton, M., Filip, G. M. & Cole, E. C. 2002. Cavity-nester habitat development in artificially
made Douglas-Fir Snags. – Journal of Wildlife Management 66(3): 625–633. DOI: 10.2307/3803129
Brooks, M. E., Kristensen, K, van Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., Skaug, H.
J., Maechler, M. & Bolker, B. M. 2017. glmmTMB balances speed and flexibility among packages
for zero-inflated generalized linear mixed modeling. – The R Journal 9(2): 378–400. DOI: 10.3929/
ethz-b-000240890M. Komlós, Z. Botta-Dukát, D. Winkler, R. Aszalós & G. Ónodi 95
Campion, D., Pardo, I., Elósegui, M. & Villanua, D. 2020. Gps Telemetry and Home Range of the White-Backed
Woodpecker Dendrocopos leucotos: Results of the First Experience. – Acta Ornithologica 55(1): 77–87.
DOI: 10.3161/00016454AO2020.55.1.008
Charman, E. C., Smith, K. W., Gruar, D. J., Dodd, S. & Grice, P. V. 2010. Characteristics of woods used recently
and historically by Lesser Spotted Woodpeckers Dendrocopos minor in England. – Ibis 152(3): 543–555.
DOI: 10.1111/j.1474-919X.2010.01020.x
Christensen, M., Hanh, K., Mountford, E. P., Ódor, P., Standovár, T., Rozenbergar, D., Diaci, J., Wijdeven, S.,
Meyer, P., Winter, S. & Vrska, T. 2005. Dead wood in European beech (Fagus sylvatica) forest reserves. –
Forest Ecology and Management 210: 267–282. DOI: 10.1016/j.foreco.2005.02.032
Czeszczewik, D. 2009. Foraging behaviour of White-backed Woodpeckers Dendrocopos leucotos in a primeval
forest (Białowiea National Park, NE Poland): Dependence on habitat resources and season. – Acta
Ornithologica 44(2): 109–118. DOI: 10.3161/000164509X482687
Czeszczewik, D., Zub, K., Stanski, T., Sahel, M., Kapusta, A. & Walankiewicz, W. 2015. Effects of forest
management on bird assemblages in the Bialowieza Forest, Poland. – iForest 8: 377–385. DOI: 10.3832/
ifor1212-007
Drever, M. C., Aitken, K. E. H., Norris, A. R. & Martin, K. 2008. Woodpeckers as reliable indicators of
bird richness, forest health and harvest. – Biological Conservation 141: 624–634. DOI: 10.1016/j.
biocon.2007.12.004
Farris, K. L. & Zack, S. 2005. Woodpecker-snag interactions: an overview of current knowledge in ponderosa
pine systems. – USDA Forest Service General Technical Report 198: 183–195.
Fox, J. & Weisberg, S. 2018. Visualizing fit and lack of fit in complex regression models with predictor effect
plots and partial residuals. – Journal of Statistical Software 87(9): 1–27. https://www.jstatsoft.org/article/
view/v087i09.
Fox, J. 2003. Effect Displays in R for Generalised Linear Models. – Journal of Statistical Software 8(15): 1–27.
https://www.jstatsoft.org/article/view/v008i15.
Garmendia, A., Cárcamo, S. & Schwendtner, O. 2006. Forest management considerations for conservation of
Black Woodpecker Dryocopus martius and White-backed Woodpecker Dendrocopos leucotos populations
in Quinto Real (Spanish Western Pyrenees). – Biodiversity and Conservation 15(4): 1399–1415. DOI:
10.1007/s10531-005-5410-0
Gibbons, P. & Lindenmayer, D. 2002. Tree hollows and wildlife conservation in Australia. – CSIRO Publishing
Gorman, G. 2004. Woodpeckers of Europe. A study of the European Picidae. – Bruce Coleman
Gorman, G. 2011. The Black Woodpecker. A monograph on Dryocopus martius. – Lynx Edicions
Gorman, G. 2015. Foraging signs and cavities of some European woodpeckers (Picidae): Identifying the clues
that lead to establishing the presence of species. – Denisia 36: 87–97.
Grubb, T. C. Jr. 1975. Weather dependent foraging behaviour of some birds wintering in a deciduous woodland.
– Condor 77: 175–182.
Hallett, J. G., Lopez, T., O’Connell, M. A. & Borysewicz, M. A. 2001. Decay dynamics and avian use of
artificially created snags. – Northwest Science 75(4): 378–386.
Hardersen, S. 2004. Habitat usage of Woodpeckers and Nuthatch. – Ricerche Naturalistiche a Bosco Della
Fontana – Quaderni Conservazione Habitat 3: 49–59. (in Italian with English Summary)
Hothorn, T., Bretz, F. & Westfall, P. 2008. Simultaneous inference in general parametric models. – Biometrical
Journal 50(3): 346–363. DOI: 10.1002/bimj.200810425
Jacobs, J. 1974. Quantitative measurement of food selection. – Oecologia 14: 413–417. DOI: 10.1007/
BF00384581
Jusino, M. A., Lindner, D. L., Banik, M. T. & Walters, J. R. 2015. Heart rot hotel: fungal communities in Red-
cockaded Woodpecker excavations. – Fungal Ecology 14: 33–43. DOI: 10.1016/j.funeco.2014.11.002
Keserű, Zs., Csiha, I., Kovács, Cs., Rásó, J. & Rédei, K. 2017. Vörös tölgyesek természetes felújítása és
erdőnevelése: Esettanulmányok [Natural regeneration of red oak (Quercus rubra) stands: case studies]. –
Erdészettudományi Közlemények 7(2): 115–125. DOI: 10.17164/EK.2017.008 (in Hungarian)
Kilgo, J. C. & Vukovich, M. A. 2014. Can snag creation benefit a primary cavity nester: Response to an experimental
pulse in snag abundance. – Biological Conservation 171: 21–28. DOI: 10.1016/j.biocon.2014.01.003
Kosiński, Z. 2006. Factors affecting the occurrence of Middle Spotted and Great Spotted Woodpeckers in
deciduous forests – a case study from Poland. – Annales Zoologici Fennici 43(2): 198–210.
Kosiński, Z. & Kempa, M. 2007. Density, distribution and nest-sites of woodpeckers Picidae, in a managed
forest of Western Poland. – Polish Journal of Ecology 55(3): 519–533.96 ORNIS HUNGARICA 2021. 29(1)
Kosiński, Z., Pluta, M., Ulanowska, A., Walczak, Ł., Winiecki, A. & Zarębski, M. 2017. Do increases in the
availability of standing dead trees affect the abundance, nest-site use, and niche partitioning of Great Spotted
and Middle Spotted Woodpeckers in riverine forests? – Biodiversity and Conservation 27(1): 123–145. DOI:
10.1007/s10531-017-1425-6
Kraigher, H., Jurc, D., Kalan, P., Kutnar, L., Levanic, T., Rupel, M. & Smolej, I. 2002. Beech coarse woody
debris characteristics in two virgin forest reserves in southern Slovenia. – Zbornik Gozdarstva in Lesarstva
69: 91–134.
Kruszyk, R. 2003. Population density and foraging habits of the Middle Spotted Woodpecker Dendrocopos medius
and Great Spotted Woodpecker D. major in the Odra valley woods near Wroclaw. – Notatki Ornitologiczne
44: 75–88.
Lenth, R. V. 2021. emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.5.4. –
https://CRAN.R-project.org/package=emmeans
Lõhmus, A., Kinks, R. & Soon, M. 2010. The importance of dead-wood supply for woodpeckers in Estonia. –
Baltic Forestry 16(1): 76–86.
Mazgajski, T. D. 1998. Nest-site characteristic of Great Spotted Woodpecker Dendrocopos major in central
Poland. – Polish Journal of Ecology 46(1): 33–41.
Melletti, M. & Penteriani, V. 2003. Nesting and feeding tree selection in the endangered White-backed
Woodpecker, Dendrocopos leucotos lilfordi. – Wilson Bulletin 115(3): 299–306. DOI: 10.1676/03-022
Mikusinski, G. 2006. Woodpeckers: distribution, conservation, and research in a global perspective. – Annales
Zoologici Fennici 43: 86–95.
Mikusiński, G., Gromadzki, M. & Chylarecki, P. 2001. Woodpeckers as indicators of forest bird diversity. –
Conservation Biology 15(1): 208–217. DOI: 10.1046/j.1523-1739.2001.99236.x
Ónodi, G. & Csörgő, T. 2014. Habitat preference of Great-spotted Woodpecker (Dendrocopos major linnaeus,
1758) and Lesser-spotted Woodpecker (Dendrocopos minor Linnaeus, 1758) in the presence of invasive plant
species – Preliminary study. – Ornis Hungarica 22(2): 50–64. DOI: 10.2478/orhu-2014-0018
Ónodi, G. & Winkler, D. 2016. Nest site characteristics of the Great-spotted Woodpecker in a bottomland riparian
forest in the presence of invasive tree species. – Ornis Hungarica 24(1): 81–95. DOI: 10.1515/orhu-2016-0005
Pasinelli, G. & Hegelbach, J. 1997. Characteristics of trees preferred by foraging Middle Spotted Woodpecker
Dendrocopos medius in Northern Switzerland. – Ardea 85: 203–209.
Pasinelli, G. 2007. Nest site selection in Middle and Great Spotted Woodpeckers Dendrocopos medius &
Dendrocopos major: implications for forest management and conservation. – Biodiversity Conservation 16:
1283–1298. DOI: 10.1007/s10531-007-9162-x
Puverel, C., Abourachid, A., Böhmer, C., Leban, J. M., Svoboda, M. & Paillet, Y. 2019. This is my spot: What
are the characteristics of the trees excavated by the Black Woodpecker? A case study in two managed French
forests. – Forest Ecology and Management 453: 117621. DOI: 10.1016/j.foreco.2019.117621
R Core Team 2018. R: A Language and environment for statistical computing. – https://www.r-project.org/
Rédei, K., Csiha, I. & Keserű, Zs. 2011. Vöröstölgyesek nevelése [Silvicultural treatment of red oak stands]. –
Erdészeti Lapok 146(11): 333–334. (in Hungarian)
Roberge, J. M., Angelstam, P. & Villard, M. A. 2008. Specialised woodpeckers and naturalness in hemiboreal
forests – Deriving quantitative targets for conservation planning. – Biological Conservation 141(4): 997–
1012. DOI: 10.1016/j.biocon.2008.01.010
Sandström, J., Bernes, C., Junninen, K., Lõhmus, A., Macdonald, E., Müller, J. & Jonsson, B. G. 2019. Impacts of
dead wood manipulation on the biodiversity of temperate and boreal forests. A systematic review. – Journal
of Applied Ecology 56(7): 1770–1781. DOI: 10.1111/1365-2664.13395
St-Amand, J., Tremblay, J. A. & Martin, K. 2018. Foraging ecology of the Williamson’s Sapsucker: Implications
for forest management. – Condor 120(3): 680–702. DOI: 10.1650/CONDOR-17-238.1
Swallow, S. K., Howard, R. A. & Gutierrez, R. J. 1988. Snag preferences of woodpeckers in a northeastern
hardwood foraging. – Wilson Bulletin 100(2): 236–246.
Thompson, I. D., Baker, J. A. & Ter-Mikaelian, M. 2003. A review of the long-term effects of post-harvest
silviculture on vertebrate wildlife, and predictive models, with an emphasis on boreal forests in Ontario,
Canada. – Forest Ecology and Management 177: 441–469. DOI: 10.1016/S0378-1127(02)00453-X
Tomiałojć, L., Wesolowski, T. & Walankiewicz, W. 1984. Breeding bird community of a primaeval temperate
forest (Bialowieza National Park, Poland). – Acta Ornithologica 20(8): 241–308.
Török, J. 1990. Resource partitioning among three woodpecker species Dendrocopos spp. during the breeding
season. – Holarctic Ecology 13: 257–264. DOI: 10.1111/j.1600-0587.1990.tb00617.xM. Komlós, Z. Botta-Dukát, D. Winkler, R. Aszalós & G. Ónodi 97
Virkkala, R. 2006. Why study woodpeckers? The significance of woodpeckers in forest ecosystems. – Annales
Zoologici Fennici 43(2): 82–85.
Walankiewicz, W., Czeszczewik, D., Tumiel, T. & Stański, T. 2011. Woodpecker abundance in the Białowieża
Forest – a comparison between deciduous, strictly protected and managed stands. – Ornis Polonica 52:
161–168.
Weiss, S. A., Corace, R. G., Toman, E. L., Herms, D. A. & Goebel, P. C. 2018. Wildlife implications across snag
treatment types in jack pine stands of Upper Michigan. – Forest Ecology and Management 409: 407–416.
DOI: 10.1016/j.foreco.2017.10.013
Wesolowski, T. 2007. Lessons from long-term hole-nester studies in a primeval temperate forest. – Journal of
Ornithology 148: 395–405. DOI: 10.1007/s10336-007-0198-1
Wesołowski, T., Mitrus, C., Czeszczewik, D. & Rowiński, P. 2010. Breeding bird dynamics in a primeval
temperate forest over thirty-five years: Variation and stability in the changing world. – Acta Ornithologica
45(2): 209–232. DOI: 10.3161/000164510x551354
Wesolowski, T. & Tomialojc, L. 1986. The breeding ecology of woodpeckers in a temperate primaeval forest –
preliminary data. – Acta Ornithologica 22(1): 1–21.You can also read

















































