Warmer and drier ecosystems select for smaller bacterial genomes in global soils
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
2770596x, 0, Downloaded from https://onlinelibrary.wiley.com/doi/10.1002/imt2.70 by Readcube (Labtiva Inc.), Wiley Online Library on [12/01/2023]. See the Terms and Conditions (https://onlinelibrary.wiley.com/terms-and-conditions) on Wiley Online Library for rules of use; OA articles are governed by the applicable Creative Commons License
Received: 4 September 2022 | Revised: 1 November 2022 | Accepted: 28 November 2022
DOI: 10.1002/imt2.70
SHORT COMMUNICATION
Warmer and drier ecosystems select for smaller bacterial
genomes in global soils
Hongwei Liu1 | Haiyang Zhang1,2 | Jeff Powell1 |
Manuel Delgado‐Baquerizo3,4 | Juntao Wang1,5 | Brajesh Singh1,5
1
Hawkesbury Institute for the
Environment, Western Sydney Abstract
University, Penrith, New South Wales, Bacterial genome size reflects bacterial evolutionary processes and metabolic
Australia
2
lifestyles, with implications for microbial community assembly and ecosystem
College of Life Sciences, Hebei
University, Baoding, China functions. However, to understand the extent of genome‐mediated microbial
3
Laboratorio de Biodiversidad y responses to environmental selections, we require studies that observe genome
Funcionamiento Ecosistemico, Instituto size distributions along environmental gradients representing different
de Recursos Naturales y Agrobiología de
conditions that soil bacteria normally encounter. In this study, we used
Sevilla (IRNAS), CSIC, Sevilla, Spain
4
Unidad Asociada CSIC‐UPO (BioFun),
surface soils collected from 237 sites across the globe and analyzed how
Universidad Pablo de Olavide, Sevilla, environmental conditions (e.g., soil carbon and nutrients, aridity, pH, and
Spain temperature) affect soil bacterial occurrences and genome size at the
5
Global Centre for Land‐Based
community level using bacterial community profiling. We used a joint species
Innovation, Western Sydney University,
Penrith, New South Wales, Australia distribution model to quantify the effects of environments on species
occurrences and found that aridity was a major regulator of genome size
Correspondence
with warmer and drier environments selecting bacteria with smaller genomes.
Haiyang Zhang, Hawkesbury Institute for
the Environment, Western Sydney Drought‐induced physiological constraints on bacterial growth (e.g., water
University, Penrith, NSW 2753, Australia. scarcity for cell metabolisms) may have led to these correlations. This finding
Email: haiyang.zhang@westernsydney.
edu.au
suggests that increasing cover by warmer and drier ecosystems may result in
bacterial genome simplifications by a reduction of genome size.
KEYWORDS
aridity, climate impacts, genome size, global soil, microbial trait, soil nutrients
Highlights
• A joint species distribution model was used to quantify the environmental
effects on occurrences of bacterial species.
• Warmer and drier environments favored bacteria with smaller genomes.
• Higher soil fertility (e.g., phosphorous) favored bacteria with smaller
genomes.
Hongwei Liu and Haiyang Zhang contributed equally to this study.
This is an open access article under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in any medium, provided
the original work is properly cited.
© 2023 The Authors. iMeta published by John Wiley & Sons Australia, Ltd on behalf of iMeta Science.
iMeta. 2023;e70. wileyonlinelibrary.com/journal/imeta | 1 of 7
https://doi.org/10.1002/imt2.702770596x, 0, Downloaded from https://onlinelibrary.wiley.com/doi/10.1002/imt2.70 by Readcube (Labtiva Inc.), Wiley Online Library on [12/01/2023]. See the Terms and Conditions (https://onlinelibrary.wiley.com/terms-and-conditions) on Wiley Online Library for rules of use; OA articles are governed by the applicable Creative Commons License
2 of 7 | LIU ET AL.
Trait‐based approaches have revolutionized ecology and one‐end of the gradients) can exist in scaled‐up/global
provided demonstrable value to our ecological under- scale environmental investigations [15]. Such environ-
standing (e.g., community assembly) of many taxa. mental effects on microbial species occurrences can be
However, knowledge of the importance of microbial either positive, negative, or neutral (Figure 1A), and the
traits lags behind other organisms (e.g., plants). In magnitude of the effect differs among bacterial species.
particular, we know little about the biogeographic To this end, four types of relationships between
pattern of microbial traits across the globe and the extent environmental effects and bacterial genome size are
to which they are under environmental selection [1, 2]. proposed: (i) trade‐offs (Figure 1B), (ii)–(iii) uni-
Among microbial traits, genome size is fundamental for directional effects (Figure 1C,D), and (iv) no correlations
the prediction of microbial physiology, their ecological (Figure 1E). In this study, we aimed to investigate
relationships with other microbes and the environment, whether genome size could mediate the effects of
and their evolutionary histories. This is because (i) environmental gradients on bacterial occurrences at the
genome size is linked to other bacterial traits related to global scale. We hypothesized that environments with
life history strategies, for example, growth rate and less favorable conditions such as drier soils (Figure 1F),
tolerance to extreme conditions [3, 4], (ii) variation in warmer climates (Figure 1G), and reduced availability of
genome size among microbial taxa can reflect evolu- soil nutrients (Figure 1H) favor occurrences of bacterial
tionary events such as genome expansion and reduction species with smaller genomes. These hypotheses are
that may be associated with emerging of new species, based on the large genome constraint hypothesis [6], and
functions or lifestyles [5, 6], and (iii) organisms with previous findings that nutrient limitation/drought‐
large genomes require more resources to grow and induced physiological constraints on growth (e.g., water
reproduce and can have a more restricted ecological scarcity for cell metabolisms) may lead to genome
distribution (large genome constraint hypothesis) [7]. In streamlining (e.g., gene loss) [18].
other words, smaller bacterial genomes require lower To test the above hypotheses, we analyzed bacterial
numbers of AGCT, and they can be more advantageous communities of soil samples collected from 237 locations
in nutrient‐limited (e.g., nitrogen, N and phosphorus, P) across the globe (Supporting Information: Figure S1),
environments [7]. Therefore, the distribution of genomic covering different ecosystem types (forests, grasslands, and
traits in environments may exhibit unique patterns shrublands) and diverse environmental gradients such as soil
reflecting nutrient conditions. aridity, nutrients, and air temperature [19]. The soil
Soil is a complex and heterogeneous ecosystem, and properties showed a wide range of variations, with soil pH
involves dynamic interactive bacterial communities that ranging from 4.04 to 9.21, soil C ranging from 0.15% to
play key roles in regulating terrestrial nutrient cycling 34.77%, soil N ranging from 0.02% to 1.57%, soil P ranging
and plant productivity. Its function is strictly dependent from 75.10 to 4111.04 mg P kg−1 soil. These sites spanned a
on critical factors like water, light, and biodiversity, gradient of −11.4°C to 26.5°C mean annual temperature and
which provide the sustenance of humanity [8]. Recent 67–3085 mm mean annual precipitation. In total 135,175 soil
studies have investigated microbial genome size in bacterial features (2315 genera, sequence variants analyzed
response to ecosystem type, trophic strategy, and oxygen by QIIME2 software, http://qiime2.org/) were detected using
use [9, 10], but investigations in soil microbial genomes 16S ribosomal RNA (rRNA) gene amplicon sequencing, and
and their responses to environmental factors along large bacterial features annotated at the genus level were extracted
spatial gradients are still limited. Current uncertainties in (see Supporting Information methods, Figures S2, S3, and
the biogeography of soil microbial genome size are due to S4). Those unidentified and less identified bacterial genera,
two main reasons. First, investigations often focus on the such as bacterium culture clones were removed from the
relationship between a single environmental factor and genera feature table. To evaluate whether aggregation of
genome size, yet to identify the most important environ- these data at the genus level was appropriate, we surveyed
mental factors requires multifactorial investigation. For bacterial genome size in the bacteria trait database [20],
example, recent studies only investigated microbial where data for both culturable and unculturable bacterial
genome size interactions with the environment in species are included for these genera. We found that across
response to warming [11–13] and nutrient availability the species within most (95%) bacterial genera (n = 2238)
[14]. Second, these studies investigated only a narrow had a low coefficient of variation (CV) (2770596x, 0, Downloaded from https://onlinelibrary.wiley.com/doi/10.1002/imt2.70 by Readcube (Labtiva Inc.), Wiley Online Library on [12/01/2023]. See the Terms and Conditions (https://onlinelibrary.wiley.com/terms-and-conditions) on Wiley Online Library for rules of use; OA articles are governed by the applicable Creative Commons License
GLOBAL SOIL BACTERIAL ASSEMBLY ALONG ENVIRONMENTAL GRADIENTS | 3 of 7
(B) (C)
(A)
(D) (E)
(F) (G) (H)
(I)
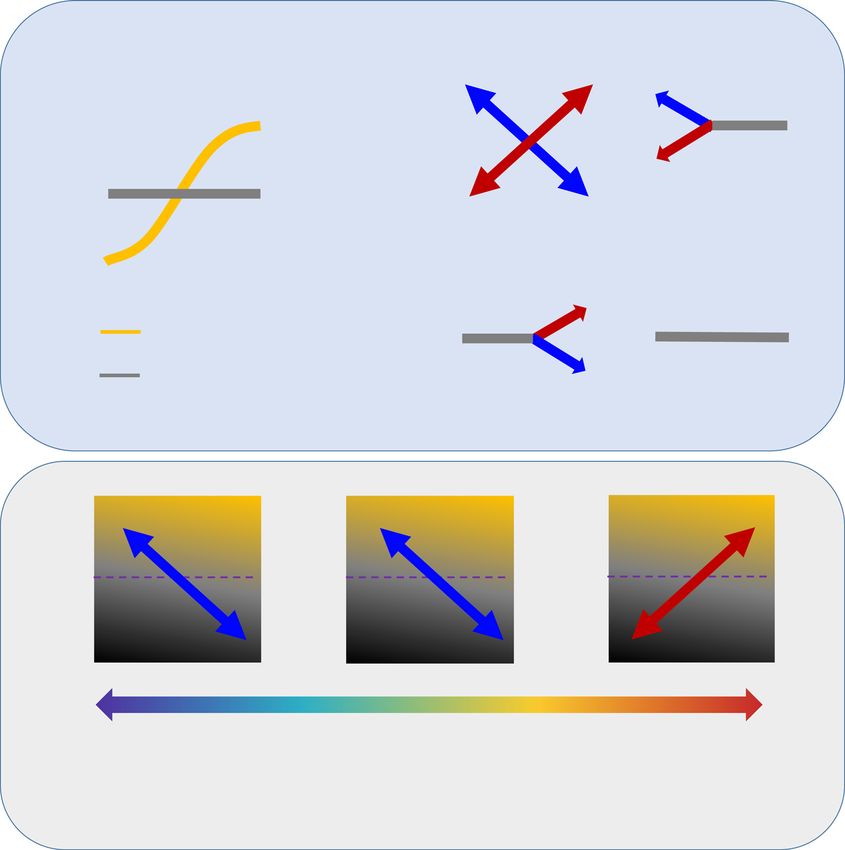
F I G U R E 1 Conception and hypotheses. The ecological responses of the probabilities of bacterial occurrences to environmental
gradients: positive (yellow), negative (black), and neutral (gray) (A). A positive (or negative) effect size indicates bacteria tend to occur with
higher (or lower) probability when specific environmental conditions increase. An effect size is the slope of the relationship between
bacterial occurrence probability and environmental conditions, calculated by beta parameters from a joint species distribution model
[16, 17]. We provide four types of correlations between microbial traits and the effects size of environments. These are (i) trade‐offs where
trait effects are bidirectional: the effect of the environmental gradients on the probability of species occurrence positively (red) or negatively
(blue) shifts along the trait gradient (B), (ii) and (iii) the trait only exhibits an effect at one end of the environmental gradient, with only a
unidirectional effect when trait values are low (C) or high (D); and (iv) no environmental effects across all trait levels (E). We hypothesized
that shifting from favorable soil conditions to stressful environments (i.e., warmer and drier climates) favors bacteria with small GS in soil.
In detail, we hypothesized that nutrients (carbon, nitrogen, and phosphorous) are positively associated with bacterial genome sizes (F),
while maximum annual temperature (MAXT) (G) and aridity (H) are negatively associated with bacterial genome sizes. Overall, we
hypothesized that stress conditions select bacteria with small genome sizes (I).
sensitivity analysis, details in Supporting Information: We then investigated relationships between the environ-
Figure S3). Thus, we set a CV threshold of 11% across mental gradients and bacterial genome size by fitting the
observations within a genus to select bacterial genera for data set with a joint species distribution model‐hierarchical
inclusion in subsequent analyses, with 1237 bacterial genera modeling of species communities (HMSC) [16, 17] (Support-
being retained. We then matched our global soil microbiome ing Information: Figure S6). After model fitting and
data with these bacterial genera [20], resulting in 143 evaluation, we extracted the effect size, that is, the slope of
bacterial genera with genome size values, ranging from the relationship between bacteria occurrence probability and
1.58 Mbp (Rickettsiella) to 16.04 Mbp (Minicystis) (Supporting environmental conditions, which was the beta‐parameters
Information: Figure S4). Our selected genera included both from HMSC with at least 95% posterior probability. For the
abundant and rare genera (Supporting Information: correlation between genome size and the magnitude of
Figure S5). environmental effects on bacterial occurrence probability, we2770596x, 0, Downloaded from https://onlinelibrary.wiley.com/doi/10.1002/imt2.70 by Readcube (Labtiva Inc.), Wiley Online Library on [12/01/2023]. See the Terms and Conditions (https://onlinelibrary.wiley.com/terms-and-conditions) on Wiley Online Library for rules of use; OA articles are governed by the applicable Creative Commons License
4 of 7 | LIU ET AL.
(A) (B) (C)
(D) (E) (F)
F I G U R E 2 Correlations between bacterial genome size (log‐transformed; Mbp) and the effects of environmental conditions on bacterial
occurrence. Y axis indicates effect sizes of an environmental condition, which are slopes (beta‐parameters) of the relationship between
bacterial occurrence probability and environmental conditions, calculated from the Hierarchical Modeling of Species Communities (HMSC).
A positive (or negative) effect size indicates bacterial genera tend to occur with higher (or lower) probability when specific environmental
conditions increase. Solid blue lines indicate correlations being significant (p < 0.05) and were fitted considering phylogenetic correlations
among bacteria genera. Aridity index reflects soil water availability (i.e., mean annual precipitation/evapotranspiration). MAXT, mean max
annual temperature.
performed phylogenetic least squares regression to account in arid ecosystem may result in bacterial genome
for phylogenetic autocorrelation among genera given that simplification. Consistently, a recent study also reported a
bacterial genome size is linked with their evolutionary positive correlation between bacterial community weight
history [21]. We found that the distance from the equator mean genome size with mean annual precipitation [23].
(absolute latitude) had a strong impact on genome size However, it is unclear in their study whether the positive
distribution in the soil (Figure 2A, R2 = 0.22, p < 0.00001), in correlation was caused by the bacteria with large genome
that bacteria with small genomes were more likely to have a sizes being positively selected by increasing water availa-
positive association with increasing latitude while bacteria bility or bacteria with small genome sizes being excluded
with large genomes exhibited the opposite. This result was in by increasing water availability, or both. Our results
line with the geographical patterns previously reported [22], support the second scenario, where bacterial species with
where the average‐genome size of bacterial communities small genome sizes had more negative responses to
decreased as distance from the equator increased. increased water availability than those with large genome
Importantly, we found that bacterial genome size size species. Similarly, warmer climates, as indicated by
significantly correlated with aridity (Figure 2B, R2 = 0.27, higher mean maximum temperature (MAXT), favored the
p < 0.00001), in that bacteria with small genomes exhibited occurrence of bacteria with small genome size by excluding
positive responses to increasing aridity (negative response large genome‐sized bacteria (Figure 2C, R2 = 0.06,
to an increasing aridity index) while bacterial genera with p = 0.003). This result is consistent with findings in
large genomes tended to not exhibit any response to aridity previous reports [12, 24], and supported by our further
(the effect sizes related these bacteria were close to zero) analyses, which showed large genome‐sized bacterial
(Figure 2B). This trend of bacterial selection with small species had a lower optimum temperature (Supporting
genomes was consistent across different taxonomic levels Information: Figure S8B, R2 = 0.07, p = 0.001). This means
such as the order and phylum level (Supporting Informa- that an increased temperature can induce higher selective
tion: Figure S7). Our further analyses demonstrated that pressures on bacteria with large genome sizes than on
large genome‐sized bacterial species contain a larger bacteria with small genome sizes.
number of genes (Supporting Information: Figure S6A, We also found that bacterial genome sizes weakly but
R2 = 0.97, p < 0.00001) (Supporting Information: significantly correlated with soil carbon (C, Figure 2D,
Figure S8A), which suggests that a genome size reduction R2 = 0.07, p = 0.001) and P (Figure 2E, R2 = 0.02,2770596x, 0, Downloaded from https://onlinelibrary.wiley.com/doi/10.1002/imt2.70 by Readcube (Labtiva Inc.), Wiley Online Library on [12/01/2023]. See the Terms and Conditions (https://onlinelibrary.wiley.com/terms-and-conditions) on Wiley Online Library for rules of use; OA articles are governed by the applicable Creative Commons License
GLOBAL SOIL BACTERIAL ASSEMBLY ALONG ENVIRONMENTAL GRADIENTS | 5 of 7
p = 0.035) concentrations, in that bacteria with small (Supporting Information: Figure S10), which may high-
genomes exhibited less negative responses to increasing light the strength of the joint species distribution
soil C or more positive responses to increasing soil P, approach in detecting soil bacterial responses to envir-
suggesting that higher soil fertility favors bacteria with onmental changes. Recent studies indicated that the
smaller genomes. Soil nitrogen did not exhibit a model‐based approach outperforms the CWM‐based
significant correlation (Figure 2F). While contradicting approaches in terms of results accuracy and statistical
our hypothesis (based on the large genome constraint power [28, 29].
theory), our results are supported by previous studies Overall, our study provides novel evidence that
where enrichments of soil nutrients (e.g., P) decreased environmental stress associated with increases in
the mean genome sizes of bacteria at the community aridity and warmer climates favors bacteria with
level in grassland soil [14]. These findings suggest that smaller genomes in soils. Drought‐induced physiolog-
larger genome‐sized bacterial species can dominate in ical constraints on growth (e.g., water scarcity for cell
environments where resources are scarce but diverse, metabolisms) may have led to these correlations.
and where there is little penalty for slow‐growth soil Bacterial community change can lead to changes in
bacteria, in terms of their colonization and competition soil ecological processes such as carbon sequestration.
in soils [22, 25]. It would be also important to investigate For example, previous studies showed that bacteria
how genome size responses to soil nutrient availability with small genomes are associated with high carbon
are linked with their r/K strategy in future studies. For use efficiency (CUE, i.e., the ratio of net carbon gain
example, previous studies showed that soil bacteria from to gross carbon assimilation, a key factor that controls
copiotrophic (r strategy) Firmicutes increased in relative carbon sequestration) [30]. Drought stress reduces
abundance by nutrient addition while oligotrophic (K plant carbon assimilation and availability to soil
strategy) bacteria such as Actinobacteria decreased in microbes. Shifting to a small genome‐size‐dominated
abundance [26]. Consistently, we found that Firmicutes community is important to increase CUE at the
had relatively small and Actinobacteria had large community level and help the ecosystem to maintain
genomes (Supporting Information: Figure S5), and, functions under stressful conditions. Favoring bacte-
therefore, our results (higher soil fertility favors bacteria rial species with small genome sizes may also have
with small genomes) can be explained by the r/K consequences on the redundancy of microbial func-
selection theory [27]. We also found that genome size tions and resilience. Soil microbes make up a complex
did not affect the sensitivity of bacterial occurrence gene pool; reducing or discarding genes for redundant
probability to changes in soil pH and mean minimum metabolic burdens may be a strategy for microbes to
annual temperature (Supporting Information: Figure S9). survive stresses [13]. Now we are facing climate
By contrast, bacterial genome size significantly correlated change that is more rapid than ever before. Despite
with net primary production (NPP, the carbon amount the unknown consequences of such microbial changes
retained by plants or other primary producers) and to ecosystem functions, future climate change
ultraviolet (UV) light, in that bacteria with large genomes research and policy‐making should consider soil
exhibited positive correlations to increasing NPP and UV bacteria and their functional traits as they play
light while bacterial genera with small genomes tended important roles in soil carbon sequestration and
to not exhibit any correlations with NPP and UV light maintaining ecosystem functions. Our study also
(Supporting Information: Figure S9). Increasing NPP suggests that lower soil fertility (soil C and P) and
favored bacteria with large genomes was in line with our higher UV light levels favor bacteria with larger
results that warming and drier ecosystems selected for genomes, which can dominate in soils where
smaller bacterial genomes, because NPP is generally resources are scarce but without major restrictions
lower under stressed environments (e.g., drought). A on slow‐growing bacteria. These findings point
positive response to UV light intensity also indicates that toward fundamental mechanisms underpinning
large genome‐sized bacteria are more tolerant to strong nucleotide selection in bacterial communities in
UV light. Mechanisms for these findings are still unclear global soils. The knowledge is also critical for efforts
and require future studies to investigate in‐depth. to predict biogeochemical feedback in the future
We acknowledge that the community‐weighted trait climate system, where drought events are expected
means (CWM), which is the average of trait values for to occur more frequently and to last longer at a global
species at each site weighted by species abundance, is scale. Our study employed bacterial analyses at the
also commonly used to study trait–environment relation- genus level by obtaining genome size data from a
ships. However, we did not observe significant patterns publicly available database. This method is reliable
between CWM genome size and environmental gradients in testing our hypothesis, and it provides a useful2770596x, 0, Downloaded from https://onlinelibrary.wiley.com/doi/10.1002/imt2.70 by Readcube (Labtiva Inc.), Wiley Online Library on [12/01/2023]. See the Terms and Conditions (https://onlinelibrary.wiley.com/terms-and-conditions) on Wiley Online Library for rules of use; OA articles are governed by the applicable Creative Commons License
6 of 7 | LIU ET AL.
tool to investigate soil genomes based on amplicon REFER ENCES
sequencing that is still vastly used for microbiome 1. Green, Jessica L., Brendan J. M. Bohannan, and Rachel J.
research. It has shortfalls in that only a part of the soil Whitaker. 2008. “Microbial Biogeography: From Taxonomy to
bacterial community can be included in analyses due Traits.” Science 320: 1039–43. https://doi.org/10.1126/science.
1153475
to the inherent limitations of 16S rRNA gene
2. Martiny, Jennifer B. H., Stuart E. Jones, Jay T. Lennon, and
amplicon sequencing in profiling bacterial communi- Adam C. Martiny. 2015. “Microbiomes in Light of Traits: A
ties (e.g., 16S rRNA gene is conserved and it may Phylogenetic Perspective.” Science 350: aac9323. https://doi.
mask certain trait variations within bacterial commu- org/10.1126/science.aac9323
nities). Future research is warranted to validify our 3. Westoby, Mark, Daniel A. Nielsen, Michael R. Gillings,
findings by investigating bacterial community assem- Elena Litchman, Joshua S. Madin, Ian T. Paulsen, and
bly in soil and other ecosystems (e.g., marine Sasha G. Tetu. 2021. “Cell Size, Genome Size, and Maximum
Growth Rate are Near‐Independent Dimensions of Ecological
ecosystems) by determining genome size from meta-
Variation Across Bacteria and Archaea.” Ecology and
genomes across environmental gradients. Evolution 11: 3956–76. https://doi.org/10.1002/ece3.7290
4. Li, Junhui, Rebecca L. Mau, Paul Dijkstra, Benjamin J. Koch,
AUTHOR CONTRIBUTIONS Egbert Schwartz, Xiao‐Jun Allen Liu, Ember M. Morrissey,
Hongwei Liu, Haiyang Zhang, Brajesh Singh, and Manuel et al. 2019. “Predictive Genomic Traits for Bacterial Growth in
Delgado‐Baquerizo conceived the idea, and Haiyang Zhang, Culture Versus Actual Growth in Soil.” ISME Journal 13:
Hongwei Liu, Jeff Powell, and Juntao Wang analyzed the 2162–72. https://doi.org/10.1038/s41396-019-0422-z
5. Morris, J. Jeffrey, Richard E. Lenski, and Erik R. Zinser. 2012.
data. Hongwei Liu and Haiyang Zhang wrote the paper and
“The Black Queen Hypothesis: Evolution of Dependencies
all authors have significantly contributed to the manuscript Through Adaptive Gene Loss.” mBio 3: e00036–12. https://doi.
revision. org/10.1128/mBio.00036-12
6. Spanu, Pietro D., James C. Abbott, Joelle Amselem,
ACKNOWLEDGMENTS Timothy A. Burgis, Darren M. Soanes, Kurt Stüber,
This study was supported by the Australian Research Emiel Ver Loren van Themaat, et al. 2010. “Genome
Council (DP190103714, DP210102081, and FT190100590). Expansion and Gene Loss in Powdery Mildew Fungi Reveal
Tradeoffs in Extreme Parasitism.” Science 330: 1543–46.
M.D‐B. acknowledges support from the Spanish Ministry
https://doi.org/10.1126/science.1194573
of Science and Innovation for the I+D+i project PID2020‐ 7. Knight, C. A. 2005. “The Large Genome Constraint Hypothe-
115813RA‐I00 funded by MCIN/AEI/10.13039/5011000 sis: Evolution, Ecology and Phenotype.” Annals of Botany 95:
11033. M.D‐B. is also supported by a project of the Fondo 177–90. https://doi.org/10.1093/aob/mci011
Europeo de Desarrollo Regional (FEDER) and the 8. Fitter, A. H., C. A. Gilligan, K. Hollingworth, A. Kleczkowski,
Consejería de Transformación Económica, Industria, R. M. Twyman, and J. W. Pitchford, Members of the Nerc Soil
Conocimiento y Universidades of the Junta de Andalucía Biodiversity Programme. 2005. “Biodiversity and Ecosystem
Function in Soil.” Functional Ecology 19: 369–77. https://doi.
(FEDER Andalucía 2014‐2020 Objetivo temático “01—
org/10.1111/j.0269-8463.2005.00969.x
Refuerzo de la investigación, el desarrollo tecnológico y la 9. Rodríguez‐Gijón, Alejandro, Julia K. Nuy, Maliheh Mehrshad,
innovación”) associated with the research project Moritz Buck, Frederik Schulz, Tanja Woyke, and Sarahi L. Garcia.
P20_00879 (ANDABIOMA). 2022“A Genomic Perspective Across Earth's Microbiomes Reveals
That Genome Size in Archaea and Bacteria is Linked to
CONFLI CT OF I NTER EST Ecosystem Type and Trophic Strategy.” Frontiers in Microbiology
The authors declare no conflict of interest. 12: 761869. https://doi.org/10.3389/fmicb.2021.761869
10. Nielsen, Daniel A., Noah Fierer, Jemma L. Geoghegan,
Michael R. Gillings, Vadim Gumerov, Joshua S. Madin,
DATA A VAILABILITY S TATEMENT
Lisa Moore, et al. 2021. “Aerobic Bacteria and Archaea Tend
The data that supports the findings of this study are to Have Larger and More Versatile Genomes.” Oikos 130:
available in the supplementary material of this article. 16S 501–11. https://doi.org/10.1111/oik.07912
rDNA data and metadata used in this study are available in 11. Dove, Nicholas C., Margaret S. Torn, Stephen C. Hart, and
Figshare (DOI: 10.6084/m9. figshare.5611321). Supporting Neslihan Taş. 2021. “Metabolic Capabilities Mute Positive
Information materials (figures, tables, scripts, graphical Response yo Direct and Indirect Impacts of Warming
abstract, slides, videos, Chinese translated version, and Throughout the Soil Profile.” Nature Communications 12:
2089. https://doi.org/10.1038/s41467-021-22408-5
update materials) may be found in the online DOI or iMeta
12. Huete‐Stauffer, Tamara M., Nestor Arandia‐Gorostidi, Laura
Science http://www.imeta.science/.
Alonso‐Sáez, and Xosé Anxelu G. Morán. 2016. “Experimental
Warming Decreases the Average Size and Nucleic Acid Content of
ORCID Marine Bacterial Communities.” Frontiers in Microbiology 7: 1–13.
Hongwei Liu http://orcid.org/0000-0002-8200-8801 https://doi.org/10.3389/fmicb.2016.007302770596x, 0, Downloaded from https://onlinelibrary.wiley.com/doi/10.1002/imt2.70 by Readcube (Labtiva Inc.), Wiley Online Library on [12/01/2023]. See the Terms and Conditions (https://onlinelibrary.wiley.com/terms-and-conditions) on Wiley Online Library for rules of use; OA articles are governed by the applicable Creative Commons License
GLOBAL SOIL BACTERIAL ASSEMBLY ALONG ENVIRONMENTAL GRADIENTS | 7 of 7
13. Sorensen, Jackson W., Taylor K. Dunivin, Tammy C. Tobin, Communities.” bioRxiv preprint 2021.2011.2017.469016.
and Ashley Shade. 2019. “Ecological Selection for Small https://doi.org/10.1101/2021.11.17.469016
Microbial Genomes Along a Temperate‐to‐Thermal Soil 24. Sabath, Niv, Evandro Ferrada, Aditya Barve, and Andreas
Gradient.” Nature Microbiology 4: 55–61. https://doi.org/10. Wagner. 2013. “Growth Temperature and Genome Size in
1038/s41564-018-0276-6 Bacteria are Negatively Correlated, Suggesting Genomic Stream-
14. Leff, Jonathan W., Stuart E. Jones, Suzanne M. Prober, lining During Thermal Adaptation.” Genome Biology and
Albert Barberán, Elizabeth T. Borer, Jennifer L. Firn, Evolution 5: 966–77. https://doi.org/10.1093/gbe/evt050
W Stanley Harpole, et al. 2015. “Consistent Responses of Soil 25. Konstantinidis, Konstantinos T., and James M. Tiedje. 2004.
Microbial Communities to Elevated Nutrient Inputs in Grass- “Trends Between Gene Content and Genome Size in
lands Across the Globe.” Proceedings of National Academy of Prokaryotic Species with Larger Genomes.” Proceedings of
Sciences of United States of America 112: 10967–72. https://doi. National Academy of Sciences of United States of America 101:
org/10.1073/pnas.1508382112 3160–65. https://doi.org/10.1073/pnas.0308653100
15. Ferenci, Thomas. 2016. “Trade‐off Mechanisms Shaping the 26. Su, Jiao, Haiyang Zhang, Xingguo Han, Ruofei Lv, Li Liu,
Diversity of Bacteria.” Trends in Microbiology 24: 209–23. Yong Jiang, Hui Li, Yakov Kuzyakov, and Cunzheng Wei.
https://doi.org/10.1016/j.tim.2015.11.009 2022. “5300‐year‐old Soil Carbon is Less Primed Than Young
16. Tikhonov, Gleb, Øystein H. Opedal, Nerea Abrego, Soil Organic Matter.” Global Change Biology 29: 260–75.
Aleksi Lehikoinen, Melinda M. J. Jonge, Jari Oksanen, and https://doi.org/10.1111/gcb.16463
Otso Ovaskainen. 2020. “Joint Species Distribution Modelling 27. Yin, Qidong, Yuepeng Sun, Bo Li, Zhaolu Feng, and
With the R‐Package Hmsc.” Methods in Ecology and Evolution Guangxue Wu. 2022. “The R/K Selection Theory and its
11: 442–47. https://doi.org/10.1111/2041-210X.13345 Application in Biological Wastewater Treatment Processes.”
17. Ovaskainen, Otso, Gleb Tikhonov, Anna Norberg, Science of the Total Environment 824: 153836. https://doi.org/
F. Guillaume Blanchet, Leo Duan, David Dunson, 10.1016/j.scitotenv.2022.153836
Tomas Roslin, and Nerea Abrego. 2017. “How to Make 28. Peres‐Neto, Pedro R., Stéphane Stéphane, and Cajo J. F.
More Out of Community Data? A Conceptual Framework ter Braak. 2017. “Linking Trait Variation to the Environment:
and its Implementation as Models and Software.” Ecology Critical Issues With Community‐Weighted Mean Correlation
Letters 20: 561–76. https://doi.org/10.1111/ele.12757 Resolved by the Fourth‐Corner Approach.” Ecography 40:
18. Simonsen, Anna K. 2022. “Environmental Stress Leads to 806–16. https://doi.org/10.1111/ecog.02302
Genome Streamlining in a Widely Distributed Species of Soil 29. Miller, Jesse E. D., Ellen I. Damschen, and Anthony R. Ives.
Bacteria.” ISME Journal 16: 423–34. https://doi.org/10.1038/ 2019. “Functional Traits and Community Composition: A
s41396-021-01082-x Comparison Among Community‐Weighted Means, Weighted
19. Delgado‐Baquerizo, Manuel, Angela M. Oliverio, Tess E. Brewer, Correlations, and Multilevel Models.” Methods in Ecology and
Alberto Benavent‐González, David J. Eldridge, Richard D. Evolution 10: 415–25. https://doi.org/10.1111/2041-210X.13119
Bardgett, Fernando T. Maestre, Brajesh K. Singh, Noah Fierer. 30. Saifuddin, Mustafa, Jennifer M. Bhatnagar, Daniel Segrè, and
2018. “A Global Atlas of the Dominant Bacteria Found in Soil.” Adrien C. Finzi. 2019. “Microbial Carbon Use Efficiency
Science 359: 320–25. https://doi.org/10.1126/science.aap9516 Predicted From Genome‐Scale Metabolic Models.” Nature
20. Madin, Joshua S., Daniel A. Nielsen, Maria Brbic, Ross Corkrey, Communications 10: 3568. https://doi.org/10.1038/s41467-019-
David Danko, Kyle Edwards, Martin K. M. Engqvist, et al. 2020. 11488-z
“A Synthesis of Bacterial and Archaeal Phenotypic Trait Data.”
Scientific Data 7: 170. https://doi.org/10.1038/s41597-020-0497-4
21. Martinez‐Gutierrez, Carolina A., and Frank O. Aylward. 2022. SU PPOR T ING INF OR MATI ON
“Genome Size Distributions in Bacteria and Archaea are
Additional supporting information can be found online
Strongly Linked to Evolutionary History at Broad Phylogenetic
in the Supporting Information section at the end of this
Scales.” PLoS Genetics 18: e1010220. https://doi.org/10.1371/
journal.pgen.1010220 article.
22. Lear, Gavin, Kelvin Lau, Anne‐Marie Perchec, Hannah L.
Buckley, Bradley S. Case, Martin Neale, Noah Fierer, et al. 2017.
“Following Rapoport's Rule: The Geographic Range and Genome How to cite this article: Liu, Hongwei, Haiyang
Size of Bacterial Taxa Decline at Warmer Latitudes.”
Zhang, Jeff Powell, Manuel Delgado‐Baquerizo,
Environmental Microbiology 19: 3152–62. https://doi.org/10.1111/
Juntao Wang, and Brajesh Singh. 2023. “Warmer
1462-2920.13797
23. Chuckran, Peter F., Cody Flagg, Jeffrey Propster,
and drier ecosystems select for smaller bacterial
William A. Rutherford, Ella Sieradzki, Steven J. Blazewicz, genomes in global soils.” iMeta. e70.
Bruce Hungate, et al. 2021. “Edaphic Controls on Genome https://doi.org/10.1002/imt2.70
Size and GC Content of Bacteria in Soil MicrobialYou can also read

















































