Uterine Glands: Developmental Biology and Functional Roles in Pregnancy
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
REVIEW
Uterine Glands: Developmental Biology and
Functional Roles in Pregnancy
Andrew M. Kelleher,1 Francesco J. DeMayo,2 and Thomas E. Spencer1,3
1
Division of Animal Sciences, University of Missouri, Columbia, Missouri 65211; 2Reproductive and
Downloaded from https://academic.oup.com/edrv/article/40/5/1424/5487358 by guest on 18 November 2021
Developmental Biology Laboratory, National Institute on Environmental Health Sciences, Research Triangle
Park, Durham, North Carolina 27709; and 3Department of Obstetrics, Gynecology, and Women’s Health,
University of Missouri, Columbia, Missouri 65211
ORCiD numbers: 0000-0003-2815-766X (T. E. Spencer).
ABSTRACT All mammalian uteri contain glands in the endometrium that develop only or primarily after birth. Gland development or
adenogenesis in the postnatal uterus is intrinsically regulated by proliferation, cell–cell interactions, growth factors and their inhibitors, as well as
transcription factors, including forkhead box A2 (FOXA2) and estrogen receptor a (ESR1). Extrinsic factors regulating adenogenesis originate
from other organs, including the ovary, pituitary, and mammary gland. The infertility and recurrent pregnancy loss observed in uterine gland
knockout sheep and mouse models support a primary role for secretions and products of the glands in pregnancy success. Recent studies in mice
revealed that uterine glandular epithelia govern postimplantation pregnancy establishment through effects on stromal cell decidualization and
placental development. In humans, uterine glands and, by inference, their secretions and products are hypothesized to be critical for blastocyst
survival and implantation as well as embryo and placental development during the first trimester before the onset of fetal–maternal circulation.
A variety of hormones and other factors from the ovary, placenta, and stromal cells impact secretory function of the uterine glands during
pregnancy. This review summarizes new information related to the developmental biology of uterine glands and discusses novel perspectives on
their functional roles in pregnancy establishment and success. (Endocrine Reviews 40: 1424 – 1445, 2019)
T he uterus is part of the female reproductive tract
(FRT) and consists of two different layers (en-
dometrium and myometrium). The endometrium con-
sheep and mice devoid of uterine glands provide direct
evidence that their secretions and products impact
conceptus survival and growth, uterine receptivity, stro-
tains many different types of cells (epithelium, stroma, mal cell decidualization, and placental growth. Con-
immune, endothelium), with the epithelium comprised of temporary studies of human implantation sites discovered
two distinctive cell types [luminal epithelium (LE) and that conceptus trophoblast cells associate with uterine
glandular epithelium (GE)]. The idea that the glands of glands and direct their secretions into the developing
the uterus produce factors essential for pregnancy success placenta. These observations support the hypothesis that
evolved over many centuries [see Refs. (–)]. During the gland-derived histotroph is critical for human conceptus
past few decades, evidence has accumulated in primate survival and growth during the first trimester prior to the
and subprimate species supporting an unequivocal role establishment of maternal blood flow toward the placenta.
for uterine glands and their secretions, termed histotroph, This review summarizes new information related to the
as principal regulators of conceptus (embryo/fetus and developmental biology of uterine glands and discusses
ISSN Print: 0163-769X
associated extraembryonic placental membranes) survival, novel perspectives on their functional roles in pregnancy
ISSN Online: 1945-7189 development, implantation, and placentation. Studies on establishment and success.
Printed: in USA
Copyright © 2019
Endocrine Society
Developmental Biology of Uterine Glands (–). However, the uterus is neither fully developed
Received: 13 December 2018
nor differentiated at birth in domestic and labora-
Accepted: 15 April 2019
First Published Online: The organogenesis and differentiation of most FRT tory animals as well as humans (, , –). Events
10 May 2019 organs are completed during prenatal development common to postnatal uterine development in laboratory
1424 https://academic.oup.com/edrv doi: 10.1210/er.2018-00281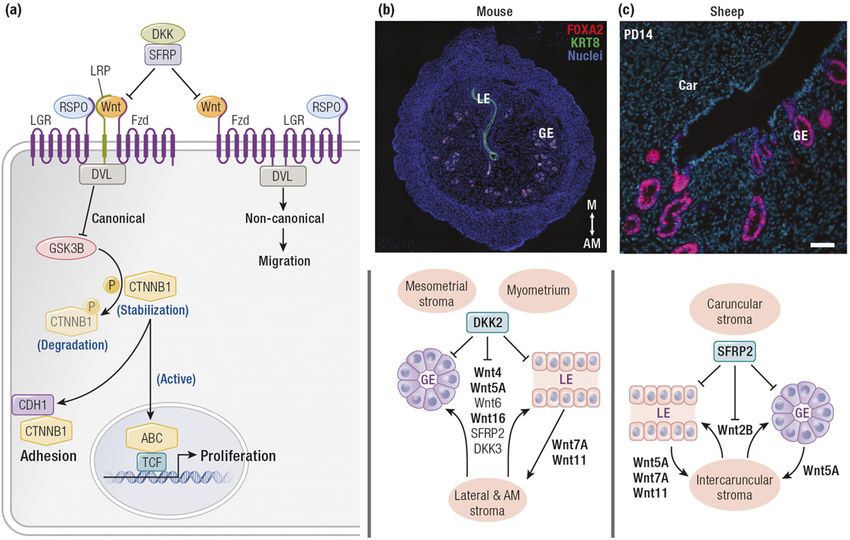
REVIEW
ESSENTIAL POINTS
· Uterine gland development or adenogenesis occurs primarily after birth and is initially ovary- and steroid-independent,
but particularly susceptible to programming by exposure to steroid hormones or endocrine-disrupting compounds
· Uterine adenogenesis is intrinsically regulated by proliferation, cell–cell interactions, growth factors, such as WNTs and
their inhibitors, as well as forkhead box A2 (FOXA2) and estrogen receptor a (ESR1) transcription factors
· Extrinsic factors regulating uterine adenogenesis originate from the ovary and pituitary as well as maternal mammary
gland
· Uterine glands are essential for fertility and pregnancy success in the adult, as their secretions and products impact
embryo survival and implantation in domestic animals and stromal cell decidualization and placental development in
mice
· Analysis of human implantation sites supports a role for uterine gland secretions or histotroph for embryo and placental
development during the first trimester, prior to the onset of fetal–maternal circulation
Downloaded from https://academic.oup.com/edrv/article/40/5/1424/5487358 by guest on 18 November 2021
· Defects in uterine gland development and/or function may be causative factors for pregnancy loss and later pregnancy
complications such as preeclampsia and fetal growth retardation
· Increased knowledge of uterine gland biology is critical to understanding and diagnosing pregnancy loss, developing
therapies for the restoration of endometrial function, and treating fertility problems and disease in women
rodents and domestic animals include: organization and domestic animals support the ideas that postnatal
stratification of the mesenchyme and/or endometrial uterine development is multifaceted and governed
stroma; myometrial differentiation and growth; and by intrinsic genetic cascades, cell proliferation, and
differentiation and development of glands within the cell–cell interactions as well as extrinsic factors from
endometrium (Fig. ). the ovary, pituitary, and mammary gland that coop-
The original descriptions of uterine gland devel- erate to achieve a fully developed and mature organ
opment or adenogenesis after birth and the architecture capable of supporting a pregnancy. The identification
of glands in the adult uterus relied on two-dimensional of genes and gene networks involved in organogen-
histology (Fig. ). During the first or second week of esis and morphogenesis of the uterus has greatly
postnatal life, adenogenesis is initiated by differentiation benefited from mouse genetic models and, more re-
of the GE from precursor LE. Nascent glands elongate cently, conditional deletion approaches employing Cre
into the stroma and then coil and slightly branch as they drivers to delete floxed alleles in almost all uterine cell
develop through the stroma toward the inner circular types or only the epithelia or stroma. In particular,
layer of the myometrium. Distinctively, gland devel- the progesterone receptor (Pgr) Cre (Pgr-Cre) mouse
opment is limited to intercaruncular areas of the en- has been invaluable to study postnatal uterine de-
dometrium of ruminants (sheep, cattle, deer), as the velopment, because Cre excision activity occurs only
aglandular caruncles are the site of placentation (). In after birth and is restricted to cells that express the Pgr,
mice and rats, adenogenesis occurs primarily in the including the uterus, ovary, oviduct, pituitary gland,
lateral and antimesometrial areas of the uterus, as the and mammary gland (). In the neonatal uterus of
upper mesometrial area of the endometrium is devoid Pgr-Cre mice, Cre excision activity begins in the LE
of glands. Recent advances in tissue clearing and mi- of the uterus by P prior to initiation of adenogenesis
croscopy have provided three-dimensional (D) models (). Thus, Pgr-Cre mice are particularly useful to
of uterine gland architecture in prepubertal and adult study genes in the postnatal or adult uterus that are
mice (–). At birth [postnatal day (P)], no rudi- embryonic lethal.
mentary gland buds are present (Fig. ). Bud- and
teardrop-shaped epithelial invaginations become evi- Intrinsic regulatory mechanism
dent between P and P and form elongated tubes by Development of the uterus depends on cell–cell in-
P. As the new glands elongate, they become coiled teractions for local control and coordination of
and sinuous. By P, the histoarchitecture of the uterus morphogenetically important cell behaviors, including
resembles that of the adult, containing glands that are differentiation, proliferation, and movement (–).
coiled and slightly branched. Proliferation of the LE, as well as nascent and budding
Our understanding of genetic and physiological GE, is observed in the neonatal uterus of mice, sheep,
pathways that regulate the formation, patterning, and and pigs (, ). A recent study suggested that a
differentiation of the uterus and, in particular, ade- critical epithelial population in the uterus must be
nogenesis has significantly relied on the study of model reached for adenogenesis to proceed normally in mice
organisms given the difficulty and ethical concerns of (). Growth of budding glands into the stroma also
studying the human (, ). Findings in laboratory and involves alterations in the basal lamina that permit and
doi: 10.1210/er.2018-00281 https://academic.oup.com/edrv 1425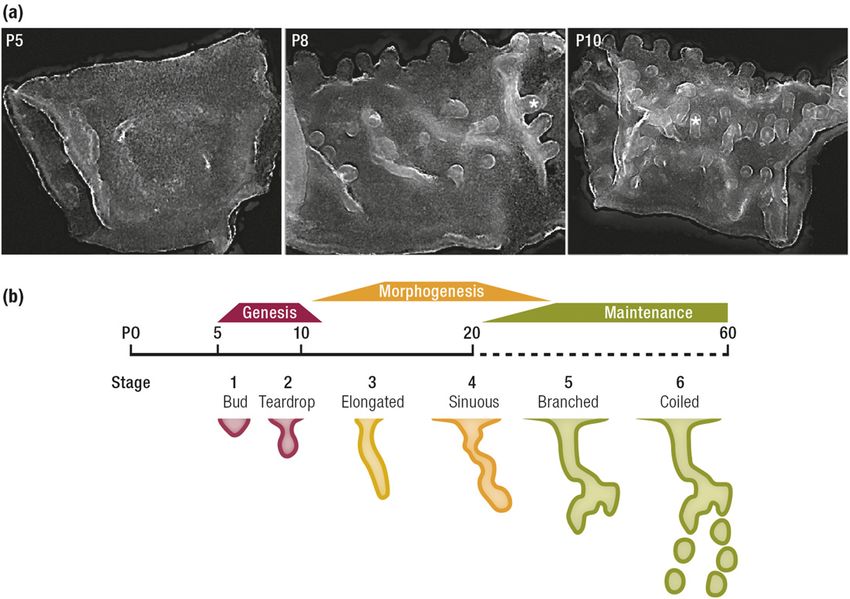
REVIEW
direct GE cell migration into the underlying stroma support the idea that Wnta coordinates a variety of
(–). A number of intrinsic growth factors have cell and developmental pathways that guide postnatal
been implicated in postnatal uterine adenogenesis, uterine differentiation and growth ().
including wingless-type mouse mammary tumor virus Wnta-null mice have short and coiled uterine
integration site family (WNT), hepatocyte growth horns of normal diameter at birth (). Given that the
factor, fibroblast growth factors (FGFs), IGFs, and Wnta-null mice perish immediately after birth, their
matrix metalloproteinases and tissue inhibitors of reproductive tract was xenografted into adult hosts to
matrix metalloproteinase [see (–, , , , )]. assess postnatal development. Although the oviduct,
uterus, and cervix of the FRT developed in the absence
WNTs of Wnta, the xenografted mutant uterus failed to
The Wnt family of genes encodes a group of highly form glands. Wnt-null females lack Müllerian and
conserved, secreted signaling molecules that are ho- Wolffian ducts and die shortly after birth (). The
mologous to the Drosophila melanogaster segment Pgr-Cre:Wnt cKO uterus possessed a pseudos-
Downloaded from https://academic.oup.com/edrv/article/40/5/1424/5487358 by guest on 18 November 2021
polarity gene wingless and critical regulators of cell fate, tratified LE with decreased gland number (). Cell-
growth, and differentiation as well as cell–cell in- specific conditional and inducible deletion studies in
teractions in many different organs (). Defects in the developing mouse uterus are needed to precisely
uterine gland development occur in Wnta, Wnta, investigate the function of Wnts and other mor-
and Wnt mutants as well as in mice harboring phoregulatory genes because adenogenesis in the
mutation in Wnt downstream effectors, catenin uterus is inhibited when the LE changes identity
(cadherin-associated protein) b (Ctnnb) and lym- and becomes stratified during the first week after
phoid enhancer binding factor (Lef) (–). WNT birth ().
signaling is categorized as either canonical or non- The canonical WNT signaling pathway is partic-
canonical () [Fig. (a)]). Both pathways are initiated ularly crucial for uterine adenogenesis. Using Pgr-Cre
by Wnt ligands binding to frizzled (Fzd) receptors. The mice, Jeong et al. () found that expression of a
canonical Wnt pathway leads to Ctnnb:T-cell factor/ dominant stabilized form of CTNNB resulted in
Lef–mediated gene expression, which regulates pro- endometrial gland hyperplasia, whereas ablation of
liferation and differentiation of cells [Fig. (a)]. Ctnnb inhibited endometrial gland development and
CTNNB is also involved in regulation and co- induced squamous cell metaplasia. Both Lef and
ordination of cell–cell adhesion as a component of cyclin D are expressed in nascent and budding glands
adherens junctions. Of note, conditional deletion of of the neonatal mouse uterus, and null Lef mutants
Ctnnb or cadherin (Cdh), a critical component fail to develop glands in their uterus after birth ().
of adherens junctions, in the mouse uterus after birth The porcupine homolog (Porcn) is a membrane-
using Pgr-Cre completely inhibited gland develop- bound O-acyltransferase that acylates all mamma-
ment (, ). The noncanonical Wnt pathway is lian WNTs (, ), which is required for WNT
mediated by intracellular calcium ion and c-Jun secretion and binding to their cognate FZD receptor
N-terminal kinase. This signaling leads to nuclear on target cells (, ). To inactivate all secreted
factor of activated T-cells (NFAT), a critical tran- WNTs, Pgr-Cre mice were used to conditionally delete
scriptional factor regulating gene expression, and ac- Porcn (). The Pgr-Cre:Porcn cKO mice displayed
tivation of the planar cell polarity pathway regulating normal prepubertal uterine development, but defects
the actin cytoskeleton that modulates cell polarity and in gland maintenance and stromal cell proliferation
motility. Noncanonical Wnt signaling is required for in the adult. Intriguingly, intrauterine injections of
tissue formation in many organs (). Wnta into the uterine horn of adult Pgr-Cre:Porcn
In the developing mouse uterus, Wnta is only cKO mice increased gland formation, whereas in-
expressed in the endometrial LE, whereas Wnt and trauterine injections of Wnta did not stimulate gland
Wnta are predominantly expressed in the endome- development, but they did stimulate proliferation of
trial stroma (–). Wnta-null mice are viable, but the LE and stroma (). Of note, Pgr-Cre:Wnta cKO
mutant females display abnormal morphogenesis mice exhibit aberrant glandular development on the
along the anterior–posterior and radial axes of the mesometrial side of the uterus ().
uterine horn after birth (). The adult Wnta-null Leucine-rich repeat–containing G protein–coupled
uterus has a stratified squamous LE (in contrast to receptor (Lgr) and its homolog Lgr were identified
simple columnar) surrounded by a small stromal layer as receptors of roof plate–specific spondins (R-
that does not contain glands (, ). Because Wnta is spondins), which are secreted Wnt pathway agonists
expressed in the Müllerian duct epithelium, the Pgr- and potentiators of canonical Wnt signaling (, ).
Cre mouse model was used to produce a conditional In the adult mouse uterus, Lgr is expressed in the LE,
knockout (cKO) of Wnta (). The uteri of Pgr-Cre: GE, and myometrium (). Conditional deletion of
Wnta cKO mice contained no endometrial glands, Lgr was conducted using keratin –Cre mice that
whereas all other cell types appeared normal with no deletes genes in the LE and GE of the uterus. Few if any
evidence of a stratified squamous LE. These studies glands were observed in the P keratin –Cre:Lgr
1426 Kelleher et al Uterine Gland Development and Role in Pregnancy Endocrine Reviews, October 2019, 40(5):1424–1445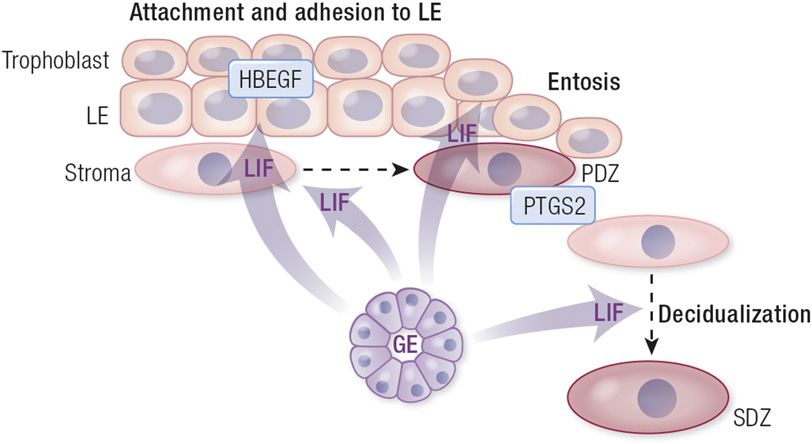
REVIEW
cKO uterus, and no glands were observed in the adult longevity in several organs and tissues (–). In
(). Thus, canonical and noncanonical WNT sig- mice, Foxa is the only family member expressed in
naling have exclusive and cooperative roles in uterine the uterus, and it is expressed explicitly in nascent,
epithelial morphogenesis and growth in the neonatal developing, and differentiated glands (, ) (Figs.
mouse. and ). Foxa-null mice die at embryonic day or
Differential WNT signaling activity limits uterine
gland development to discrete areas of the endome-
trium in both mice and sheep [Fig. (b) and (c)]. In
the neonatal mouse uterus, active WNT signaling
activity was discovered to be progressively restricted
to epithelia in the lateral and antimesometrial areas
of the uterus (). Expression of dickkopf (Dkk), a
secreted potent WNT inhibitor that antagonizes
Downloaded from https://academic.oup.com/edrv/article/40/5/1424/5487358 by guest on 18 November 2021
canonical WNT signaling by inhibiting WNT ligand
interactions with the coreceptors low-density lipo-
protein receptor–related protein (LRP) and LRP
(), is confined to the stroma of the aglandular
mesometrial area and myometrium of the uterus ()
[Fig. (b)]. Of note, progesterone treatment of
newborn mice induced Dkk in the antimesometrial
area of the uterus in concert with inhibition of gland
genesis ().
In the neonatal sheep uterus, in situ hybridiza-
tion studies found that WNTA, WNTA, and
WNT were expressed in uterine epithelium, whereas
WNTB was expressed in the stroma () [Fig. (c)].
The WNT receptors FZD and FZD were detected
in all uterine cell types, with FZD being particularly
abundant in endometrial epithelia. In the same regard,
coreceptor LRP was detected in all uterine cells.
Expression of secreted FZD-related protein (SFRP) , a
soluble WNT antagonist, was abundant in the stroma
of the aglandular areas of the endometrium, termed
caruncles, between P and P. Thus, caruncular
SFRP expression may provide a spatial mechanism to
restrict active WNT signaling and adenogenesis to the
intercaruncular areas of the uterus. Of note, estrogen-
induced disruption of adenogenesis was associated
with aberrant induction of SFRP and decreased WNT
expression in the uterus (). Thus, WNT signaling
appears to define morphogenetically active areas of
the endometrium that are permissive to uterine
adenogenesis in the developing uterus of both sheep
and mice.
Transcription factors
Forkhead box A2. FOX (forkhead box) A
transcription factors comprise a subfamily of forkhead
transcription factors that contain high homology in
the winged helix DNA-binding domain and are
termed “pioneer” transcription factors, as they can
facilitate the binding of other transcription factors to
compacted chromatin (). The FOXA family of
transcription factors includes FOXA, FOXA, and
FOXA (previously termed HNFa, HNFb, and
HNFg, respectively) (), and these factors have
essential roles in regulating expression of genes in-
volved in cell growth, proliferation, differentiation, and
doi: 10.1210/er.2018-00281 https://academic.oup.com/edrv 1427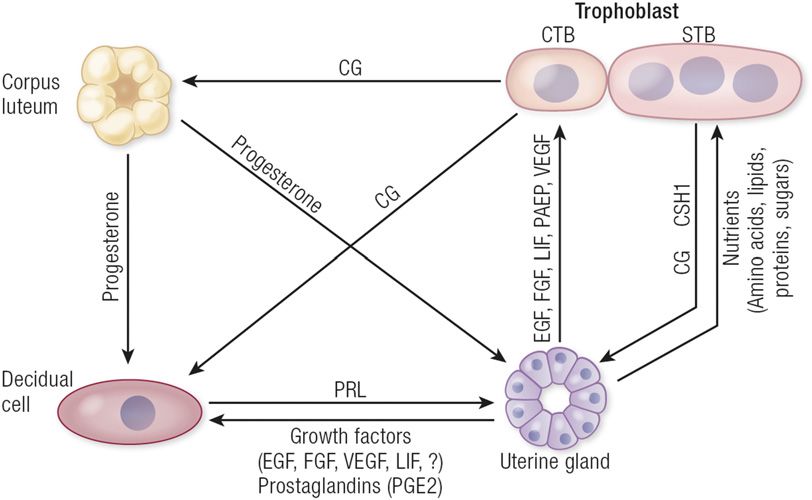
REVIEW
due to severe defects in the notochord, neural tube, Distal-less homeobox genes. The distal-less
and gut tube formation (). To obviate embryonic homeobox (Dlx) gene family includes three bigenic
lethality, Foxa was conditionally ablated in the uterus clusters (Dlx/, Dlx/, Dlx/) that comprise a highly
immediately after birth using the Pgr-Cre mouse (). conserved family of homeobox genes homologous to
In the Pgr-Cre:Foxa cKO uterus, gland differentiation the distal-less (Dll) gene of Drosophila. Genetic dis-
was inhibited, rendering adult mice glandless and ruption of Dxl/ leads to perinatal death, limb
infertile. Of note, the original report of Jeong et al. () malformations, and defects in craniofacial develop-
found that some Pgr-Cre:Foxa cKO mice had glands, ment (, ). Dlx and Dlx are expressed in the LE
but recent studies with those mice found no evidence and developing GE in the postnatal mouse uterus as
of glands (–). The differences in the phenotype of well as in the GE of the human adult uterus ().
the Pgr-Cre:Foxa cKO mice could be due to genetic Conditional deletion of Dxl and Dxl using Pgr-Cre
background and the environment as well as changes in mice severely impaired gland development in the
the diet and microbiota (, ). neonate and resulted in complete infertility in the
Downloaded from https://academic.oup.com/edrv/article/40/5/1424/5487358 by guest on 18 November 2021
FOXA is expressed solely in the glands of the adult (). Genes involved in postnatal uterine ade-
developing and adult uterus of both mice and sheep nogenesis (Foxa, Wnta) were dysregulated in Dxl/
[Fig. (b) and (c)] as well as pigs, cattle, and humans cKO mice. Thus, Dxl and Dxl may be upstream of
(, ). Thus, FOXA is postulated to be a conserved key developmental genes controlling epithelial mor-
critical transcription factor controlling the differenti- phogenesis in the neonatal mouse uterus.
ation and function of uterine GE in all mammals. In Estrogen receptor a. Adenogenesis in neo-
many other organs, such as the liver, lung, and pan- natal rodent (–), pig (, ), and sheep () uteri
creas, Foxa has an essential role in epithelial budding involve coordinated changes in epithelial phenotype
and morphogenesis (). Integrated chromatin im- that are marked by estrogen receptor a (ESR) ex-
munoprecipitation sequencing and cell-specific gene pression in nascent and proliferating glands. Neither
expression profiling have identified many potential Esr or Esr is essential for organogenetic development
Foxa-regulated genes and biological pathways (cell and patterning of the Müllerian duct in the fetus
cycle, cell junction, focal adhesion, and WNT sig- (–); however, Esr is essential for uterine growth
naling) in the developing neonatal mouse uterus (, and development in mice after birth (). The initial
). FOXA regulates expression of multiple WNTs in stages of uterine development and adenogenesis from
other organs, including Wnta, Wntb, and Wnta birth (P) to P occur normally in Esr-null mice
(–). Furthermore, CTNNB can promote Foxa (). Uterine epithelial proliferation and gland de-
expression in the prostate, indicating a reciprocal velopment were also similar in Esr-null and wild-type
interaction between FOXA and WNT signaling (). mice from P to P (). After a nadir of proliferation
Recently, ablation of sex determining region Y (SRY)- around P, uterine cells begin to proliferate again and
box (Sox) using Pgr-Cre mice was found to uterine weight increases (), presumably as a result of
phenocopy Foxa ablation, indicating that it may increased estrogen secretion between P and P (,
regulate Foxa expression in the nascent GE cells ). However, Esr-null mice exhibited little LE
within the LE, but the underlying mechanism is yet to proliferation and a considerable loss of glands by P
be defined (, ). Determination of the mechanism compared with wild-type mice (). Thus, Esr is
of how FOXA is transcriptionally regulated as well as dispensable for the initiation of gland genesis in the
the genes regulated by FOXA should reveal critical neonate, but necessary for the maintenance of de-
biological pathways governing postnatal uterine gland veloped uterine glands after P and further increases
morphogenesis and function (). in gland number as the mice attain puberty.
In sheep treated with the antiestrogen EM-, a
pure ESR antagonist, from birth, uterine growth was
not affected on P (). However, the intercaruncular
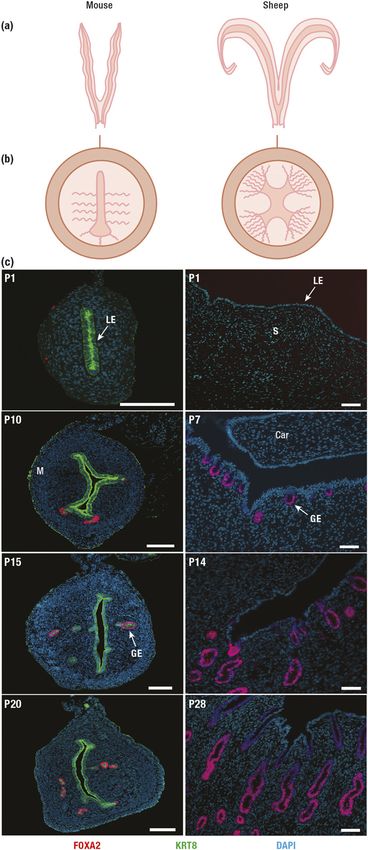
Figure 1. Uterine morphology, radial patterning, and postnatal development in mice and sheep. endometrium contained fewer ductal gland invagi-
(a) Diagrams of ideal frontal sections of uterine types. The drawings cut the oviducts off near the nations, and endometrial glands were less coiled and
uterotubal junctions, and the vagina just caudal to the cervix. Rodents (rats and mice) have a long
branched in EM-–treated ewes as compared with
duplex type of uterus with dual cervices. Sheep have a medium-length bicornuate type of uterus
and a short uterine body and a single cervix. (b) Diagrams of ideal radial patterns of the uterine wall. control ewes. In contrast to mice and sheep, ESR has
The curved lines in the endometrium denote the tubular, coiled, and slightly branched glands that a crucial regulatory role in uterine development in
extend from the uterine lumen to the inner layer of myometrium. The mouse uterus lacks the newborn pig. Administration of the antiestrogen
appreciable glands in the upper third of the endometrium on the mesometrial area of the uterus. ICI ,, a potent ESR antagonist, from birth
The sheep uterus contains large number of glands in the intercaruncular areas of the endometrium, inhibited uterine adenogenesis and overall growth
whereas the caruncles are glandless. (c) Immunofluorescent localization of forkhead box a2
at P in neonatal gilts (). Thus, uterine ESR ex-
(FOXA2) and keratin (KRT8) was performed for sections of the postnatal mouse uterus, whereas
only FOXA2 was localized in the sheep uterus. Note the expression of FOXA2 in nascent and
pression and activation are important elements of
developing glands. Sections were counterstained with 49,6-diamidino-2-phenylindole (DAPI) to the organizational program that determines patterns
visualize all nuclei. Scale bar, 100 mm (mouse) and 50 mm (sheep). Car, caruncle; M, myometrium; of uterine growth and adenogenesis in an age- and
P, postnatal day; S, stroma. species-dependent manner.
1428 Kelleher et al Uterine Gland Development and Role in Pregnancy Endocrine Reviews, October 2019, 40(5):1424–1445REVIEW
Figure 2. Overview of
uterine gland development
in the mouse before
puberty. (a) Dark-field
images of the uterine
epithelium isolated before
(P5) and during genesis and
budding of the glands from
the LE (denoted by an
asterisk). (b) Overview
of uterine gland
morphogenesis based on
postnatal age, two-
dimensional histology, and
3D imaging as proposed
Downloaded from https://academic.oup.com/edrv/article/40/5/1424/5487358 by guest on 18 November 2021
by Vue et al. (18).
Elegant tissue recombination studies involving differentiation of many epithelial-mesenchymal or-
mouse uterine stroma and epithelium indicate that gans, including the mammary gland (–). Cir-
epithelial ESR is neither necessary nor sufficient to culating levels of PRL in ewe lambs are relatively high
mediate the mitogenic actions of estrogen (, ). on P, increase to a maximum on P, and then
Instead, the proliferative effects of estrogen on epi- decline slightly to P (, ). Expression of the PRL
thelium appear to be mediated primarily by stromal receptor (PRLR) is restricted to nascent GE buds on
ESR via production of paracrine-acting, stromal- P, and proliferating and differentiating GE from P
derived growth factors such as epidermal growth to P (). Hyperprolactinemia, induced in neonatal
factor (EGF), IGF, and IGF (, –). The ewes by treatment with recombinant ovine PRL from
precise biological roles and significance of ligand- birth to P, increased uterine gland number by .%
dependent and ligand-independent actions of ESR (). Furthermore, induction of hypoprolactinemia
and stromal–epithelial interactions in uterine gland using bromocriptine, an inhibitor of pituitary PRL
morphogenesis and growth remain to be determined. secretion, in neonatal ewes from birth to P reduced
uterine glands by % (). Those effects of PRL on
the neonatal ovine uterus were mediated by activa-
Extrinsic Regulatory Mechanisms tion of the Janus kinase–signal transducer and acti-
vator of transcription (JAK-STAT) as well as MAPK
Evidence in domestic and laboratory animals estab- pathways that impact cell growth and differentiation.
lished that postnatal uterine development and ade- How PRL and its receptor impact gland develop-
nogenesis are regulated by extrinsic factors that ment and function in other mammals is not well
originate from a number of sources, including the understood. The PRLR is expressed explicitly in the
pituitary and ovary as well as the maternal mammary glands of the human uterus (), but not in the mouse
gland. uterus ().
Pituitary prolactin Ovary and its hormones
Prolactin (PRL) is a member of a unique hormone Data for several species, including the mouse (), rat
family that includes chorionic somatomammotropin (), sheep (, ), and pig (, ), indicate that
hormone (CSH; originally termed placental lac- uterine growth and morphogenesis, marked by the
togen) and GH based on genetic, structural, binding, genesis and development of nascent glands, proceed
receptor signal transduction, and function studies normally for a period of time after bilateral ovari-
(). These hormones regulate the growth and ectomy at birth. Although initial gland genesis is
doi: 10.1210/er.2018-00281 https://academic.oup.com/edrv 1429REVIEW
ovary-independent, studies in sheep and mice support and circulating estrogen levels are negligible (,
the concept that estrogens and other unknown factors , ).
from the ovary act in an endocrine manner to impact Ovariectomy of ewe lambs at birth did not affect
prepubertal uterine adenogenesis and growth. initial adenogenesis on P (), whereas ovariectomy
on P reduced uterine growth as well as the number of
Sheep glands on P (). Candidate ovarian-derived fac-
The newborn sheep ovary contains significant num- tors from the growing and antral follicles include
bers of growing and antral ovarian follicles at birth follistatin, activins, and inhibin as well as IGFs (,
(~ and per ovary, respectively) that increase in ). Of note, the activin–follistatin system is present
number by P ~ and per ovary), and then in both the uterus and ovary of neonatal ewes (,
decline in number by P (~ and per ovary) ). The Inverdale gene mutation (FecXI) in sheep
(, ). There is no evidence that those ovarian results in an increased ovulation rate in heterozygous
follicles secrete appreciable amounts of estrogens ewes, but homozygous Inverdale ewes (II) are infertile
Downloaded from https://academic.oup.com/edrv/article/40/5/1424/5487358 by guest on 18 November 2021
between birth and puberty, as aromatase (CYPA) with “streak” ovaries that lack normal developing
expression is not detectable in the granulosa cells preantral and antral follicles (). When compared
Figure 3. Proposed source and actions of the WNT signaling system in postnatal uterine development in mice and sheep. (a) Schematic
illustrating the canonical and noncanonical WNT signaling pathways and inhibition of those pathways by DKK and SFRP. Activation of
the canonical signaling stimulates epithelial adhesion and proliferation as well as stromal cell proliferation in the intercaruncular
endometrium. Activation of the noncanonical pathway would stimulate epithelial cell migration and movement. (b) Autocrine and
paracrine actions of WNTs and their inhibitors in the neonatal mouse uterus. The upper panel displays immunofluorescent localization
of forkhead box A2 (FOXA2) and keratin (KRT8) in the adult mouse uterus; note that FOXA2 is only expressed in the glands. In the
lateral and antimesometrial (AM) stroma, WNTs expressed in the LE and stroma may have autocrine or paracrine actions on the LE,
stroma, and/or GE to promote uterine adenogenesis. However, the WNT inhibitor DKK2 is expressed predominantly by the stroma of
the mesometrial area (M) and myometrium that inhibits WNT signaling and thus inhibiting GE growth and development in the upper
mesometrial area of the endometrium as well as the myometrium. Original magnification, 310. (c) Autocrine and paracrine actions of
WNTs and their inhibitors in the neonatal ovine uterus. The upper panel displays immunolocalization of FOXA2 in the uterus of a P14
ewe; note that FOXA2 is only expressed in the glands. In the intercaruncular endometrium, WNTs expressed in the LE (WNTs 5A, 7A,
and 11) may have autocrine or paracrine actions on the LE or stroma, respectively. WNT2B is expressed only in the stroma and may have
autocrine or paracrine actions on the stroma or the LE and GE, respectively. WNT5A is expressed predominantly by the GE and may have
autocrine or paracrine actions on the GE or stroma, respectively. SFRP2 is expressed predominantly by the stroma of the caruncles as well as
by the intercaruncular stroma between the tips of the glands and the inner circular layer of the myometrium. Binding of the FZD receptors
for the WNTs on the epithelia and stroma inhibit epithelial growth and development into the caruncular areas of the endometrium as well
as the myometrium. Scale bar, 50 mM. ABC, active b-catenin; Car, caruncle; CDH1, cadherin 1; DKK, dickkopf; DVL, dishevelled; FZD, frizzled
receptor; GSK3B, glycogen synthase kinase 3b; LGR, leucine-rich repeat–containing G protein–coupled receptor; LPR, lipoprotein receptor-
related protein; RSPO, roof plate–specific spondin; SFRP, secreted FZD-related protein; TCF, transcription factor.
1430 Kelleher et al Uterine Gland Development and Role in Pregnancy Endocrine Reviews, October 2019, 40(5):1424–1445REVIEW
with wild-type ewes on P, uterine growth of ho- maintenance after P is complex and involves actions
mozygous Inverdale ewes was reduced, and the en- of extrinsic ovarian-derived factors, such as estrogen
dometrium contained fewer glands (). Although and activin A, as well as peripheral-derived Kiss-
circulating concentrations of estrogen in neonatal ewes dependent factors from other organs.
were low to undetectable, homozygous Inverdale ewes
had lower concentrations of testosterone and inhibin- Mammary gland and the lactocrine hypothesis
a between P and P. Thus, developing preantral Lactation, the secretion of milk by the mammary
and/or antral ovarian follicles secrete endocrine-acting glands, is a defining characteristic of mammals, and
factors that stimulate uterine growth and adenogenesis maternal milk contains a complex array of bioactive
in neonatal ewes. The precise nature of those factors is factors (, ). Findings in the pig strongly support
not known, but they are not estrogens and could be that bioactive factors communicated from mother to
activins or follistatin from the large number of ovarian nursing offspring in first milk (colostrum) may
follicles. provide a vital source of extraovarian, uterotrophic
Downloaded from https://academic.oup.com/edrv/article/40/5/1424/5487358 by guest on 18 November 2021
support (, ). The term lactocrine refers to a
Rodents mechanism by which milk-borne bioactive factors are
In the neonatal rat, normal uterine growth and dif- communicated from mother to offspring in colostrum
ferentiation are independent of the ovaries and ad- during nursing (, , ).
renals before P, and uterine growth and In the pig, uterine glands are absent or rudimentary
adenogenesis are not appreciably affected by ovari- at birth (). Between birth and P, nascent GE cells
ectomy on P when assessed on P (). Similarly, differentiate from LE and begin to proliferate rapidly,
normal uterine growth and differentiation were ob- forming uterine glands (, ). This neonatal period
served in neonatally ovariectomized mice until P coincides with the time during early lactation when
(). However, a recent study found that uterine concentrations of bioactive factors in colostrum are
growth and gland maintenance after P requires the highest (, , ). Within hours after birth,
ovary (). The nature of the ovarian-derived factors macromolecular components of colostrum are no
impacting uterine growth and development after longer able to diffuse readily across the intestinal
weaning is not known, but a strong candidate factor is epithelium (, ). Lactocrine effects on neonatal
estrogen or an estrogen-regulated growth factor given porcine uterine development occur immediately after
postnatal changes in circulating estrogen and the in- birth upon first ingestion of colostrum. A single
fluence of Esr on uterine gland maintenance (). Of feeding of colostrum at birth increased endometrial
note, Bigsby et al. () grafted whole pieces of the cell proliferation at hours postnatally. In the same
neonatal uterus from wild-type or Esr-null mice into regard, nursing for hours from birth supported the
syngeneic wild-type or Esr-null adult host mice. establishment of the uterine developmental program
Using an ovariectomy and hormone replacement as reflected by patterns of uterine cell proliferation and
approach, estrogen-stimulated proliferation of Esr- expression of morphoregulatory genes (). Impo-
null uterine grafts was apparent, but only when they sition of a lactocrine-null state in the neonatal gilt by
were grown in wild-type hosts. These observations feeding milk replacer for days from birth decreased
support the idea that estrogen stimulates the pro- uterine gland development and endometrial thickness
duction of a systemic growth factor in an unknown on P (). Consistently, both glandular ESR ex-
organ through an Esr-dependent mechanism that pression and cell proliferation were reduced on P in
impacts uterine growth and adenogenesis. uterine tissues obtained from lactocrine-null gilts
Recently, global ablation of KiSS- metastasis (). Further studies established that lactocrine
suppressor (Kiss) or its receptor (Kissr) in mice was support from birth alters the postnatal uterine de-
also observed to impair uterine growth and decreased velopmental program and trajectory of uterine de-
gland number by % at P (). Although circu- velopment in the neonatal pig and contributes to the
lating levels of estrogen were not different, both adult programming of endometrial tissues necessary to
Kiss- and Kissr-null mice had smaller uteri with few support optimal embryotrophic function of the uterus
or no Foxa-positive glands (). Estrogen re- into adulthood (, , ).
placement of Kiss-null mice from to weeks of age
for weeks restored uterine growth, but it only Progesterone as an endocrine disruptor
partially rescued adenogenesis. Similarly, reexpression The developing and adult FRT is highly sensitive to
of Kissr in the hypothalamus of adult Kiss-null mice synthetic chemicals, in particular, those mimicking
restored uterine growth, but it only partially rescued hormones utilizing nuclear receptors. Endocrine-
gland development based on Foxa-positive gland disrupting compounds [see Ref. ()] are either
numbers. Alterations in ovarian inhibin (Inhba) and natural or synthetic exogenous compounds that in-
follistatin gene expression were observed in Kissr- terfere with the physiology of normal endocrine-
repleted Kiss-null mice. Collectively, these recent regulated events, such as reproduction and growth
studies support the idea that uterine growth and gland (), because they are agonists and antagonists of
doi: 10.1210/er.2018-00281 https://academic.oup.com/edrv 1431REVIEW
nuclear hormone receptors. Although ovarian steroids progesterone are not observed in Ihh-null mice or
and their receptors (Esr, Pgr) are not required for fetal mice lacking Pgr in the uterine epithelium ().
FRT organogenesis or initial neonatal uterine devel- Similar to UGKO sheep, progesterone-induced
opment, their receptors are expressed in the prenatal UGKO (PUGKO) mice are infertile and exhibit re-
and postnatal uterus during critical developmental current early pregnancy loss due to failure in embryo
windows. Thus, the uteri of humans (), rodents implantation (, ). Studies with UGKO sheep
(–), cattle (), pigs (, –), and sheep and mice established that uterine glands and, by in-
(, ) are particularly vulnerable to developmentally ference, their secretions and products are primary
disruptive effects of natural or synthetic endocrine- determinants of pregnancy success.
disrupting compounds.
Studies in rodents (, ) and sheep (, )
established that onset of adenogenesis in the neonatal Functional Roles of Uterine Glands
uterus occurred independently of a requirement for
Downloaded from https://academic.oup.com/edrv/article/40/5/1424/5487358 by guest on 18 November 2021
ovarian support or steroid hormones and that pro- Uterine glands directly synthesize and secrete or se-
gestins could suppress uterine epithelial cell pro- lectively transport from serum transudate a wide
liferation. Consequently, it was hypothesized that birth variety of substances, collectively termed histotroph,
provides an endocrine cue for the initiation of uterine into the lumen of the uterus (). Histotroph is complex
gland genesis (). Subsequently, exposure of new- and comprised of many different substances, includ-
born sheep to norgestomet, a nonmetabolizable and ing ions, sugars (glucose and fructose), amino acids,
potent synthetic form of progesterone, from birth was extracellular vesicles (exosomes and microvesicles),
found to inhibit uterine adenogenesis (). Removal lipids, and proteins. Recent studies of UGKO sheep
of norgestomet on P permitted glands to develop and mice established that uterine glands and, by in-
by P, although those glands were underdeveloped ference, their products and secretions are essential for
and histologically abnormal. This original obser- pregnancy and have biological roles in conceptus
vation served as the foundation for the idea that survival and growth, uterine receptivity, embryo im-
prolonged exposure of neonatal ewes to progestins plantation, stromal cell decidualization, and placental
during the critical period of adenogenesis would development (Figs. and ) ().
permanently inhibit uterine gland differentiation ().
Subsequently, exposure of neonatal ewes to norges- UGKO models
tomet from birth to weeks of life was discovered as a
method to permanently inhibit postnatal differentia- Sheep
tion of uterine glands, thereby creating a uterine gland The uteri of adult UGKO ewes are histoarchitecturally
knockout (UGKO) phenotype in adults (–). In normal except for the lack of GE and reduced LE ().
neonatal sheep, norgestomet inhibition of adeno- Notably, development and function of the brain
genesis involved inhibition of LE proliferation and (hypothalamus and pituitary), ovary, and other FRT
expression of ESR and FGFR (, ). Adult structures (oviducts, cervix, and vagina) were not al-
UGKO ewes exhibited complete infertility due to tered in UGKO ewes (, ). UGKO ewes exhibit
recurrent early pregnancy loss stemming from defects complete infertility due to recurrent pregnancy loss
in blastocyst survival and growth (). (, , ). Morphologically normal blastocysts
In neonatal mice, transient exposure to pro- were present in UGKO ewes after mating on gesta-
gesterone from P to P permanently inhibited tional day (GD) , but conceptuses were absent or
uterine adenogenesis and produced a UGKO phe- severely growth retarded when assessed on GD or
notype in the adult (, ). Progesterone treatment GD. Moreover, transfer of blastocysts from normal
decreased uterine LE proliferation and increased ewes was unable to rescue pregnancy in the UGKO
stromal cell proliferation (, , ). The anti- ewes. Thus, failure of conceptus elongation in UGKO
proliferative actions of progesterone on the LE, some ewes was attributed to the absence of specific,
of which serve as GE progenitors, may be the un- embryotrophic secretions that emanate primarily from
derlying cause of failure in endometrial adenogenesis the glands (, ).
(). Increased stromal cell proliferation could also Embryotrophic factors produced by the uterine
disrupt stromal–epithelial crosstalk necessary for GE glands that are required for conceptus survival and
differentiation (, , , ). In the adult mouse elongation have not been defined in sheep. Pro-
uterus, Indian hedgehog (Ihh) is a progesterone- gesterone drives changes in expression of genes by the
upregulated gene expressed in the LE and GE (, endometrial epithelium (LE and GE) that modulate
, ). Ihh acts in a paracrine manner through transport of ions, amino acids, glucose, and lipids from
smoothened (SMO) in the stroma to regulate LE serum and produce proteins, extracellular vesicles, and
cell proliferation, and overexpression of Smo using other bioactive substances (prostaglandins and corti-
Pgr-Cre mice caused defects in adenogenesis (, sol) (, –). Arginine is one of the only com-
). Interestingly, the antiadenogenic effects of ponents of histotroph demonstrated in vivo to be
1432 Kelleher et al Uterine Gland Development and Role in Pregnancy Endocrine Reviews, October 2019, 40(5):1424–1445REVIEW
functionally important for blastocyst growth in sheep immune cells, vascular endothelium, and basal aspect
(). of the LE. In contrast, apical secretions of GE cells
would primarily enter the uterine lumen and impact
Mice embryo and placental development.
UGKO mice can be created by exposure of neonatal
mice to progesterone from P to P (, , ) or Biological roles of uterine glands in pregnancy
conditional deletion of Foxa using Pgr-Cre mice establishment: lessons from Foxa2 cKO mice
(–). Uterine receptivity is defined as a temporally unique
Except for the uterus, the FRTs of PUGKO and sequence of factors that make the endometrium re-
Pgr-Cre:Foxa cKO mice were histoarchitecturally ceptive to embryo implantation and are mainly reg-
indistinguishable from those of wild-type mice. Both ulated by ovarian steroid hormones estrogen and
PUGKO and Pgr-Cre:Foxa cKO mice cycled and progesterone (, ). In mice, the uterus becomes
mated normally but were infertile due to an embryo receptive on day of pregnancy (day represents
Downloaded from https://academic.oup.com/edrv/article/40/5/1424/5487358 by guest on 18 November 2021
implantation defect. Histological assessment of their observation of a postcoital vaginal plug), whereas by
uteri found hatched blastocysts apposed to an intact the afternoon of day , it spontaneously becomes
LE without evidence of embryo implantation or the refractory to blastocyst implantation (–). When
onset of stromal cell decidualization (, ). Ex- synchrony between an implantation-competent blas-
pression of leukemia inhibitory factor (Lif), a critical tocyst and receptive endometrium is achieved, blas-
implantation factor of GE origin (), was absent in tocyst implantation commences (, ). The outer
the uteri of UGKO mice, whereas steroid hormone trophectoderm (TE) cells of the blastocyst then attach
receptors (Esr, Pgr) and their regulated genes were and adhere to the LE. Penetration into the stroma
not different with the exception of Lif. Of note, Lif-null involves removal of the LE and migration of the TE.
mice are also infertile due to defective embryo im- Implantation triggers robust stromal cell deciduali-
plantation (). Blastocysts from PUGKO mice could zation, which is critical for pregnancy success in mice
be successfully transferred into the uterus of wild-type and humans (–).
recipients, where they implanted and established In its broadest sense, decidualization can be viewed
pregnancy (). Thus, the infertility observed in as the postovulatory process of endometrial remod-
UGKO mice was attributed to faulty uterine re- eling in preparation for pregnancy that includes
ceptivity resulting from the absence of LIF and perhaps coordinated proliferation and differentiation of en-
other specific uterine gland–derived products. dometrial stromal cells into large epithelioid decidual
As in domestic animals, the essential factors pro- cells, secretory transformation of the uterine glands,
duced by the uterine glands that impact pregnancy influx of specialized uterine natural killer cells, and
establishment in mice are not known with the ex- vascular remodeling (, , –). Many of the
ception of LIF. Sequential effects of ovarian and cellular and genetic changes in decidualizing stromal
progesterone alter expression of genes in the endo-
metrial epithelium (LE and GE), but the biological
roles of most of those genes have not been determined
(, , ). Moreover, few studies have characterized
components of histotroph in the uterine lumen of
mice. Carbohydrates and amino acids in the uterine
lumen were initially profiled only from mice at estrus
(). Recently, glucose, amino acids, and proteins
were profiled in the uterine lumen of pseudopregnant
mice during the acquisition of uterine receptivity
(). Different from domestic animals and humans,
dynamic changes in those histotroph components
were not observed. Surprisingly, many proteins and
most amino acids were more abundant in uterine
luminal fluid from PUGKO than wild-type mice,
indicating that uterine glands have a primarily bi-
ological role in homeostasis of the uterine lumen ().
Of note, LIF and serine peptidase inhibitor Kazal type Figure 4. Uterine glands secrete factors that impact pregnancy establishment in the mouse. In
(Spink; also known as Spink), which are GE- response to the nidatory surge in ovarian estrogen on day 4 of pregnancy, LIF is secreted from
FOXA2-positive uterine glands, causing HBEGF expression for trophoblast attachment and
specific gene products (), were not identified in
adhesion to the LE. Following attachment, unknown gland-derived factors are involved in
the uterine lumen of wild-type mice. That study coordinating the removal of the LE (entosis) within the implantation chamber, allowing for direct
supports the idea that the polarized GE can secrete contact between the trophoblast and decidualizing stromal cells (PTGS2). Uterine glands are
proteins and other substances in a basolateral direction involved in secondary stromal cell decidualization (SDZ), but the identification of factors involved
that primarily act in a paracrine manner on the stroma, in mediating those processes requires the establishment of new in vivo and in vitro model systems.
doi: 10.1210/er.2018-00281 https://academic.oup.com/edrv 1433REVIEW
cells are conserved between mice and humans (, embryo death (–). Transporters for glucose and
). In mice, initiation of embryo attachment re- amino acids are expressed in the uterine glands and
action, occurring near midnight on GD, is followed may impact trophoblast activation and motility that
by extensive stromal cell proliferation surrounding the are necessary for embryo implantation (, , ).
implanting blastocyst that is evident by the morning of In species exhibiting embryonic diapause (skunks,
GD. Between GD and GD, stromal cells, adjacent mice, bears), uterine glands are thought to modulate
to the implanted blastocyst in the antimesometrial area embryo quiescence and reactivation by altering bio-
of the uterus, cease proliferating and undergo differ- active substances, such as selected amino acids and
entiation, forming the primary decidual zone (PDZ), polyamines, in the uterine lumen (, ).
an area that is avascular and epithelioid (). Stromal
cells adjacent to the PDZ continue to proliferate and Uterine receptivity and implantation
differentiate to form the secondary decidual zone Uterine glands and their secretions have long been
(SDZ). In humans, decidualization commences during implicated in uterine receptivity and blastocyst im-
Downloaded from https://academic.oup.com/edrv/article/40/5/1424/5487358 by guest on 18 November 2021
the mid-secretory phase ( to days after ovulation) in plantation in mice and humans (, –).
both nonpregnant and pregnant women and prog- Studies on LIF in mice provided the first unequivocal
resses significantly in the area beneath and sur- evidence that uterine glands secrete a factor required
rounding the embryo implantation site (). for uterine receptivity and blastocyst implantation
(). Lif is expressed explicitly on GD by the
Blastocyst activation uterine GE in response to the “nidatory” surge of
Blastocyst activation is the process by which the estrogen from the ovary (, , ). Lif-null mice
blastocysts acquire implantation competency and can are infertile owing to defects in embryo implantation,
attach to receptive uterine LE (). Amino acid and but IP injections of recombinant mouse LIF initiated
glucose transport into the uterine lumen is hypoth- embryo implantation and rescued pregnancy in Lif-null
esized to be essential for blastocyst activation and mice (). Moreover, conditional uterine knockout
implantation based on in vitro studies. Both leucine of Gp and Stat, downstream mediators of LIF
and arginine are required in culture media for blas- signaling, results in implantation failure phenocopying
tocyst activation and trophoblast motility in the that is seen in Lif-null mice (). Implantation defects
preimplantation mouse embryo (–). Precise in other null mutant (Lef) and conditional mutant
regulation of glucose within the uterine lumen is likely mice (Ctnnb, Foxa, Lgr, Wnt, Wnta) lacking
an essential component for embryo activation and uterine glands are also attributed to diminished or
implantation. Mouse and human embryos require absent Lif expression (, , , , ).
glucose in vitro, but increased glucose concentration Recently, Foxa was conditionally deleted in the
can result in developmental delays and increased epithelium of the adult uterus after puberty using the
new and very useful lactotransferrin (Ltf)-iCre mouse
model (). In both gland-containing Ltf-iCre:Foxa
cKO and glandless Pgr-Cre:Foxa cKO mouse models,
blastocysts failed to attach and adhere to the LE and
initiate stromal cell decidualization for pregnancy
establishment. Steroid receptors and most established
hormone-regulated uterine receptivity genes were not
different in the Foxa cKO and wild-type mice (, ,
, ). As expected, uteri of glandless Pgr-Cre:
Foxa cKO mice lacked Lif expression on GD (,
, , ). Surprisingly, uteri of Ltf-iCre:Foxa cKO
mice also lacked Lif expression despite having uterine
glands (). Both Foxa cKO mouse models pheno-
copy Lif-null and PUGKO mice in that they exhibited
defects in blastocyst implantation. Studies of Ltf-iCre:
Foxa cKO mice established that Foxa regulates the
ability of estrogen and Esr to induce Lif and perhaps
other GE genes (, ). Both FOXA and FOXA
influence ESR transcriptional activity in other organs
Figure 5. Hypothesis on the interrelationships of the ovarian corpus luteum, conceptus and cancer cells (, ).
trophoblast, uterine glands, and decidual cells during early pregnancy in humans. See text for
Remarkably, IP injections of LIF on GD was able
detailed description of hypotheses and supporting data. [Adapted from Burton GJ, Jauniaux E,
Charnock-Jones DS. Human early placental development: potential roles of the endometrial glands.
to rescue implantation in both gland-containing Ltf-
Placenta. 2007;28(Suppl A):S64–S69. This is an open access article distributed under the Creative iCre:Foxa cKO and glandless Pgr-Cre:Foxa cKO
Commons Attribution License, which permits unrestricted use, distribution, and reproduction in mice (, ). Heparin-binding EGF (Hbegf) induction
any medium, provided the original work is properly cited.] in the LE adjacent to the blastocyst is an early
1434 Kelleher et al Uterine Gland Development and Role in Pregnancy Endocrine Reviews, October 2019, 40(5):1424–1445REVIEW
molecular marker of the attachment reaction in mice The first evidence that endometrial glands may
(). Hbegf expression was not observed in the LE impact stromal cell decidualization originated from
adjacent to the blastocysts of both types of Foxa cKO studies of deciduoma formation in glandless mice ().
mice on the night of GD; however, LIF repletion was An artificial model of decidualization can be employed
sufficient to trigger blastocyst attachment to the LE, in mice that involves ovariectomy, hormone re-
induction of Hbegf expression, and blastocyst im- placement, and intrauterine administration of oil as a
plantation (). These studies with Foxa cKO mice deciduogenic stimulus (). Using that artificial
and others firmly establish that LIF is a critical factor model, PUGKO mice exhibited a distinct lack of
derived and secreted by the GE that acts on the LE to uterine stromal cell decidualization termed a decid-
establish uterine receptivity and permit activated uoma (). Similarly, mice with conditional ablation
blastocysts to initiate TE attachment and implantation of Foxa, Lgr, or Wnt have a uterus with much
in mice. reduced or absent glands and also exhibit defects in
In rodents, implantation crypt formation is es- blastocyst implantation and stromal cell decidualiza-
Downloaded from https://academic.oup.com/edrv/article/40/5/1424/5487358 by guest on 18 November 2021
sential for pregnancy success (, , ). The crypts tion (, , , ).
form in the gland-containing antimesometrial area of Morphological and mechanistic studies of UGKO
the uterus on GD, and blastocysts enter those crypts mice (PUGKO and Pgr-Cre:Foxa cKO) support the
before the onset of the attachment reaction. Im- concept that uterine glands secrete factors that impact
plantation crypt formation is independent of the stromal cell decidualization. In UGKO mice, there is no
preimplantation surge in estrogen from the ovary, as it evidence of LE cell removal and stromal cell decidu-
also occurs in a delayed implantation mouse model alization adjacent to the blastocyst (, , , , ).
(). Several mutant mouse lines (Msx/, Wnta, In LIF-replaced Foxa cKO mice, GD implantation
Vangl) exhibit aberrant crypt formation resulting in sites appeared normal based on morphology (decidual
increased embryo resorption (, , ). Histo- swellings), histology, cell proliferation (Ki), and
logical studies of Pgr-Cre:Foxa cKO mice established upregulation of prostaglandin-endoperoxide synthase
that implantation crypt formation and localization of (Ptgs), a molecular marker of PDZ formation (, ).
blastocysts within those crypts were not different from However, by GD, defects in embryo development and
wild-type mice (). decidual regression were evident in LIF-replaced
New D visualization studies revealed that epi- glandless mice, with full embryo loss and resorption
thelial evaginations forming implantation chambers by GD (). Transcriptome analysis of GD im-
(crypts) consistently arise with preexisting glands, plantation sites revealed numerous genes and pathways
suggesting direct access of glands to embryos and altered in the uterus of control as compared with LIF-
decidual cells within the chamber (). While the replaced glandless Pgr-Cre:Foxa cKO mice. In-
lobular domains of the glands become more de- terestingly, many known decidualization-related genes
veloped, the ductal regions continue to elongate and are increased in implantation sites of glandless Pgr-Cre:
progressively stretch following embryo implantation. Foxa cKO mouse uteri (Bmp, Cdh, Cebpb, Ptx,
Thus, glands remain in the implantation site sur- Wnt) (), which may signify premature stromal cell
rounding the embryo during implantation, decidual- differentiation into decidual cells (). The increased
ization, and pregnancy establishment. Notably, the expression of genes crucial to decidualization on GD
mechanisms allowing for the selective removal of the and persistence of stromal proliferation on GD suggest
LE but not the GE by the TE during early implantation perturbed decidual progression and premature decidual
have not been investigated. senescence in Pgr-Cre:Foxa cKO mice. These findings
support the idea that uterine glands produce paracrine-
Stromal cell decidualization acting factors that govern stromal cell decidualization by
The concept that uterine glands secrete paracrine- modulating proliferation, differentiation, and/or poly-
acting factors into the stroma to promote deciduali- ploidization () (Figs. and ). The concept of gland-
zation is relatively new (). Studies of Spink, a derived factors influencing decidualization is novel and
gland-specific gene in the uterus, support this concept supported by evidence that LIF enhanced mouse and
(). Although Spink mRNA is present only in the human endometrial stromal cell decidualization in vitro
GE of the mouse uterus, Spink protein was detected ().
in the LE and decidual cells as well as glands of the In LIF-replaced gland-containing Ltf-iCre:Foxa
pregnant mouse uterus. Furthermore, Lif, Spink, and cKO mice, stromal cell decidualization progresses
other GE-specific proteins were not identified in a with the formation of a functional SDZ and placenta
proteomic analysis of the uterine lumen of mice (). based on histology (). There is no difference in
These studies support the idea that vectorial secretion implantation site number in LIF-replaced gland-
of proteins in a basolateral direction by the polarized containing Ltf-iCre:Foxa cKO mice and control
GE has paracrine effects on other cell types within the mice on either GD or GD or number of live pups at
endometrium, including the stroma, immune cells, term. Although no overt histological differences were
and LE (Figs. and ). observed in the decidua and placenta, the expression of
doi: 10.1210/er.2018-00281 https://academic.oup.com/edrv 1435You can also read

















































