Uterine flushing proteome of the tammar wallaby after reactivation from diapause
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
REPRODUCTION
RESEARCH
Uterine flushing proteome of the tammar wallaby after
reactivation from diapause
Florine C Martin1, Ching-Seng Ang2, David K Gardner1, Marilyn B Renfree1 and Geoff Shaw1
1
School of BioSciences and 2Bio21 Mass Spectrometry and Proteomics Facility, Bio21 Molecular Science and
Biotechnology Institute, The University of Melbourne, Melbourne, Australia
Correspondence should be addressed to G Shaw; Email: g.shaw@unimelb.edu.au
Abstract
The marsupial tammar wallaby has the longest period of embryonic diapause of any mammal, up to 11 months, during which
there is no cell division or blastocyst growth. Since the blastocyst in diapause is surrounded by acellular coats, the signals that
maintain or terminate diapause involve factors that reside in uterine secretions. The nature of such factors remains to be
resolved. In this study, uterine flushings (UFs) were used to assess changes in uterine secretions of tammars using liquid
chromatography–mass spectrometry (LC–MS/MS) during diapause (day 0 and 3) and reactivation days (d) 4, 5, 6, 8, 9, 11 and
24 after removal of pouch young (RPY), which initiates embryonic development. This study supports earlier suggestions that the
presence of specific factors stimulate reactivation, early embryonic growth and cell proliferation. A mitogen, hepatoma-derived
growth factor and soluble epidermal growth factor receptors were observed from d3 until at least d11 RPY when these secreted
proteins constituted 21% of the UF proteome. Binding of these factors to specific cellular receptors or growth factors may
directly stimulate DNA synthesis and division in endometrial gland cells. Proteins involved in the p53/CDKN1A (p21) cell cycle
inhibition pathway were also observed in the diapause samples. Progesterone and most of the oestrogen-regulated proteins
were present in the UF after d3, which is concomitant with the start of blastocyst mitoses at d4. We propose that once the p21
inhibition of the cell cycle is lost, growth factors including HDGF and EGFR are responsible for reactivation of the diapausing
blastocyst via the uterine secretions.
Reproduction (2016) 152 491–505
Introduction cellular cytoplasm (Casado-Vela et al. 2009, Scotchie
et al. 2009, Hannan et al. 2010).
The uterine environment provides a complex array
Reproduction in the tammar wallaby is characterised
of secreted factors under dynamic endocrine control
by embryonic diapause. Like all marsupials, tammars
(Clemetson et al. 1977, McRae 1988, Salleh et al. 2005).
Several studies in the 1970s used uterine flushing (UF) to give birth to an immature young that undergoes
sample uterine fluid in order to characterise the changing extended development whilst attached to the teat
composition of uterine secretions in humans and animal (Tyndale-Biscoe & Renfree 1987). Within 1 h after
models (Urzua et al. 1970, Renfree 1972, 1973, Aitken birth, the female mates (Rudd 1994), and the resulting
1974, Wolf & Mastroianni 1975, Ametzazurra et al. conceptus grows for only 7 days in the uterus until it
2009, Boomsma et al. 2009), but the techniques of the reaches about 80 cells when further cell division is
time lacked sensitivity and specificity now possible with halted as a result of the sucking stimulus of the new
modern proteomic techniques. A major contribution pouch young (Renfree & Shaw 2000). This unilaminar
to the uterine contents comes from secretions of the blastocyst is surrounded by an acellular shell coat,
glandular epithelial cells (Salamonsen et al. 2009), a mucin layer and a zona pellucida. The diapausing
including nutrients, proteases, hormones, cytokines and embryo has no measurable metabolic activity, no cell
growth factors, associated with the regulation of uterine division or cell growth (Moore 1978, Thornber et al.
function and embryonic development (Kane et al. 1997, 1981, Shaw & Renfree 1986, Spindler et al. 1998,
Hempstock et al. 2004, Dominguez et al. 2010, Hannan 1999), but removing the sucking pouch young (RPY)
et al. 2011, Binder et al. 2014, Thouas et al. 2015). removes the neuroendocrine inhibition of the corpus
Human UFs are complex, with more than 800 proteins luteum. Consequently, circulating progesterone rises
derived from epithelial cell secretions, transudates from after day 3 to a peak at day 5–6 after RPY (Hinds &
blood, and from breakdown of cells distinct from the Tyndale-Biscoe 1982), which stimulates increased
© 2016 Society for Reproduction and Fertility DOI: 10.1530/REP-16-0154
ISSN 1470–1626 (paper) 1741–7899 (online) Online version via www.reproduction-online.org
Downloaded from Bioscientifica.com at 03/10/2022 04:29:27AM
via free access
492 F C Martin and others
Increase in metabolic reactivation et al. 1986). Furthermore, progesterone alone is
Increase in cell division sufficient to reactivate diapausing embryos (Renfree &
Expansion starts
Blastocyst
Tyndale-Biscoe 1973). In some marsupial endometria,
the region around the nucleus of each cell undergoes
Days after
Removal
active protein synthesis; these secretions may include
of Pouch 0 1 2 3 4 5 6 7 8 9 10 11 24 25 26 cell compartments that continue to flow into uterine
lumen (Shorey & Hughes 1973, Walker & Hughes 1981)
Young (RPY)
and these patterns are similar to that in the tammar
Day of birth
Endometrium (Tyndale-Biscoe & Renfree 1987). However, after day
15, differences between the two uteri emerge that are
No uterine changes before due to the presence of the developing embryo, and
day 3 after RPY
appear to be the result of stimulation by the placenta
Figure 1 Schematic illustration of the sequential events from diapause (Renfree 1972, 2015) perhaps mediated by placental
to reactivation and birth at d26 RPY. Reactivation occurs after hormones (Menzies et al. 2011), demonstrating that
inhibition of the CL by the early morning pulse of prolactin which
there is maternal recognition of pregnancy (Renfree &
has been removed for three consecutive days. If a young is replaced
onto the teat at any time up to 72 h after RPY, reactivation will not Shaw 2000, Renfree 2015). Thus, only the gravid uterus
occur. On the fourth day of the removal of PY (d4 RPY), there is an maintains secretory activity as pregnancy progresses
increase in mitosis, protein synthesis and transport followed by an (Renfree 1972, 2000, Renfree & Tyndale-Biscoe 1973,
increase in RNA synthesis by d5 RPY. However, the first expansion of Tyndale-Biscoe & Renfree 1987).
the blastocyst does not occur until d8 RPY. The sampled days are In tammars, the embryo is separated from the
enclosed in boxes. (Adapted from Renfree & Shaw 2000). uterine epithelium by an acellular shell coat during
diapause and until after day 17 of the 26-day active
endometrial secretory activity and reactivation of gestation, preventing direct cellular interaction with the
the diapausing blastocyst (Renfree 1973, Renfree & endometrium, so control of diapause and reactivation
Tyndale-Biscoe 1973, Shaw & Renfree 1986, Fletcher must be achieved through soluble factors in the uterine
et al. 1988, Shaw 1996, Renfree & Shaw 2000). fluids (Renfree 1972, Renfree & Tyndale-Biscoe 1973,
Reactivation commences after day 3 (Gordon et al. Shaw 1996). Components of UF are derived from
1988, Hinds et al. 1989). By day 4 RPY, there is a secretions from the luminal epithelium and glands,
dramatic increase in glucose oxidation and glutamine proteins selectively transuded from blood, and include
uptake and oxidation by the blastocyst and the first several uterine-specific pre-albumins during reactivation
mitoses in the blastocyst are observed (Spindler et al. (Renfree 1973). Previous studies have not identified the
1998, 1999, Fig. 1). specific uterine regulatory factor(s) that control diapause
Tammars have two separate uteri, but, since they are of the embryo. In this study, we have therefore used a
monovular, only one becomes gravid; the contralateral non-biased global proteomics approach to characterise
uterus is non-gravid. Both uteri respond similarly because the proteome of the uterine fluid in diapause and
of local distribution of progesterone from the ipsilateral reactivation. We profiled tammar proteins in UF from
corpus luteum (Towers et al. 1986) until around day diapause until late gestation with the aim of identifying
15 when only the gravid uterus becomes larger and and evaluating if those proteins might potentially be
more secretory due to local embryonic signals (Renfree implicated in the maintenance of diapause or embryonic
1972, Renfree & Tyndale-Biscoe 1973, Renfree & Shaw reactivation, expansion and development.
2000, Renfree 2015). Embryo transfer experiments
(Tyndale-Biscoe 1963, 1970, Renfree 1972) confirm that
the corpus luteum (CL) does not directly stimulate the Materials and methods
quiescent blastocyst to resume development, but rather Animal maintenance
acts by inducing changes in the uterine secretions.
Embryo transfer experiments in eutherian mammals Tammars of Kangaroo Island, South Australia origin was
kept in open grassy yards with shelters provided. Their diet
have provided further evidence of the importance of
was supplemented with fresh fruits, vegetables and lucerne
the uterus during embryonic development in diapause
cubes and water supplied ad libitum. Care and treatment
and that the uterine signals regulating the embryo are
of animals conformed to the National Health and Medical
conserved (Chang 1968, Ptak et al. 2012, Cha et al. Research Council Australian Code for the Care and Use
2013, Fenelon et al. 2014, Renfree & Shaw 2014). of Animals for Scientific Purposes 2013. Animal handling
Specific components of the uterine secretions and experimentation were approved by the University of
undoubtedly play a role in regulating early embryo Melbourne Animal Experimentation Ethics Committees. In
development. There is greater rate of protein synthesis this study, adult females with a pouch young older than day
in the gravid uterus of the tammar (Renfree 1972, 8 post-partum were presumed to be carrying a diapausing
1973, Shaw & Renfree 1986) because progesterone blastocyst (Tyndale-Biscoe & Renfree 1987). Reactivation was
is preferentially delivered to the gravid uterus (Towers achieved during the seasonal breeding period (Jan–May) by
Reproduction (2016) 152 491–505 www.reproduction-online.org
Downloaded from Bioscientifica.com at 03/10/2022 04:29:27AM
via free accessUterine flushing proteome of tammar wallaby 493
removing the pouch young, and stages of pregnancy were performance liquid chromatography (HPLC) (Dionex, CA,
determined relative to the day of the removal of the pouch USA). The HPLC system was equipped with an Acclaim
young (designated day 0 after RPY). Pepmap nano-trap column (Dinoex-C18, 100 Å, 75 µm × 2 cm)
and an Acclaim Pepmap RSLC analytical column (Dinoex-C18,
Sample collection and preparation 100 Å, 75 µm × 15 cm) (Dionex, CA, USA). The tryptic peptides
were injected to the enrichment column at an isocratic flow of
Analysis of marsupial uterine fluids poses problems because 5 µL/min of 3% (v/v) acetonitrile containing 0.1% v/v formic
of their small volumes, which preclude direct collection of the acid for 5 min applied before the enrichment column was
secretions, so we used uterine flushings. Females were killed switched in-line with the analytical column. The eluants were
humanely on days 0, 3, 4, 5, 6, 8, 9, 11 and 24 after removal 0.1% v/v formic acid (solvent A) and 100% v/v acetonitrile
of pouch young. Five animals at each stage with paired gravid in 0.l% v/v formic acid (solvent B). The flow gradient was
and non-gravid UF samples except at days 3, 11 and 24, where (i) 0–5 min at 3% B; (ii) 5–6 min at 3–6% B; (iii) 6–18 min
three animals were used and at days 11 and 24 UF from only at 6–10% B; (iv) 18–38 min at 10–30% B; (v) 38–40 min at
the gravid side, were collected. Immediately post-mortem, 30–45% B; (vi) 40–42 min at 45–80% B; (vii) 42–45 min at
the reproductive tract was dissected out and gravid and non- 80% B; (vii) 45–46 min at 80–3% B; and (viii) 46–53 min at
gravid uteri were each flushed with 2 mL of 0.9% saline. The 3% B. The LTQ Orbitrap Elite spectrometer was operated in the
volume of uterine fluids is minute and effectively exists only data-dependent mode with nano ESI spray voltage of 2.0 kV,
as a moist surface, so the concentration of protein in the capillary temperature of 250°C and S-lens RF value of 55%.
flush was taken as the amount of protein in the uterine fluids All spectra were acquired in positive mode with full scan
per 2 mL. The flushings were examined under a dissecting mass spectrometry (MS) spectra scanning from m/z 300–1650
microscope and any blastocysts were retrieved. About 10 µL of in the FT mode at 240,000 resolutions after accumulating
protease inhibitors (Sigma Aldrich) was added to the collected to a target value of 1.0e6. A lock mass of 445.120025 was
flushings. It was then centrifuged at 16,000 g for 10 min at used. The top 20 most intense precursors were subjected to
4°C in a refrigerated microcentrifuge (Eppendorf, Hamburg, collision-induced dissociation (CID) with normalized collision
Germany) to remove cell debris. The supernatants were stored energy of 30 eV and activation q of 0.25. A dynamic exclusion
at −80°C until further use. This study characterises the protein duration of 45 s was applied for repeated precursors.
composition of UF in tammar from diapause (day 0 and day
3), 4, 5, 6, 8 and 9, 11 and 24 RPY. Chemicals were purchased
from Sigma Aldrich unless otherwise indicated.
Data analysis
The protein concentrations of gravid UF were determined
using a BCA Protein Assay Kit−Reducing Agent Compatible Protein concentrations in UF at the different stages in both
(Thermo Fisher Scientific) following the manufacturer’s gravid and non-gravid uteri were assessed using analysis of
instructions. variance (ANOVA). Data are presented as mean ± s.d. unless
An aliquot equivalent to 100 µg of protein was initially with otherwise indicated. All statistical analysis was carried out
reduced by the addition of 5 mM dithiothreitol (DTT) at 95°C using R (R Core Team 2015).
for an hour and alkylation with 50 mM iodoacetamide (IAA) The MS and MS/MS data were searched using Proteome
in the darkness. This was followed by overnight digestion with Discoverer (Thermo Scientific Version 1.4) with the MASCOT
sequencing grade modified trypsin (Thermo Pierce) at 37°C on search engine (Version 2.4.1) against a tammar proteome
a shaker with the ratio of enzyme to protein, 1:50. The following database generated in-house (15,344 protein sequences) from
day, the protein digestion was halted by adding formic acid to the tammar genome version 2.0 (Renfree et al. 2011). Search
final 1% (v/v). The protein digest was then purified using an criteria used were trypsin digestion, variable modifications set
Oasis HLB (polymeric reverse phase) solid phase extraction as carbamidomethylation of cysteine (Cys) and oxidation of
(SPE) cartridge (Waters Corporation, Milford, MA, USA). methionine (Met), allowance of up to two missed cleavages,
Purified peptides were then lyophilised overnight in a freeze precursor tolerance of 10 ppm and 0.6 Da on the fragment ions.
dryer (Virtis, PA, USA) and reconstituted in 0.1% formic acid A targeted false discovery threshold of 0.05, Fig. 2). Concentrations
Scientific) mass spectrometer with a nanoESI interface rose progressively after d 3 from about 25 µg/mL at days
in conjunction with an Ultimate 3000 RSLCnano high- 0 and 3 to about 150 µg/mL at d 9 (P < 0.0001, Fig. 2).
www.reproduction-online.org Reproduction (2016) 152 491–505
Downloaded from Bioscientifica.com at 03/10/2022 04:29:27AM
via free access494 F C Martin and others
250 (Stavréus-Evers et al. 2002, Wang & Dey 2006). It is likely
that proteins involved in adhesion present on the luminal
200
gravid * epithelial surface are important for the maintenance of
Protein concentrations
non gravid integrity between adjacent epithelial cells. Hepatoma-
150 derived growth factor, Transforming growth factor
* Significantly different to d0.
(TGFβ) 68 kDa protein and epidermal growth factor
µg/mL
*
receptor (EGFR) were identified at different stages of
100
* reactivated UFs and in d3 UFs but not in the d0 samples.
* UFs contain significant number of growth factors, which
50 * were found to be secreted into the lumen from the day
of reactivation (d4 RPY) and these increased up to late
0 gestation (Table 2). Proteins involved in the p53/p21
0 1 2 3 4 5 6 7 8 9
cell cycle inhibition pathway (Fig. 3A) including Septin
Days after RPY
2 (SEPT2), Septin 7 (SEPT7), mitogen-activated protein
Figure 2 The protein concentrations (mean ± s.d.) measured by BCA kinase 1 (MAPK1), proliferation-associated protein
assay (n = 5 for each stage and side) across the stages (d0–d9 RPY) in 2G4 (PA2G4), glia maturation factor beta (GMFB),
the gravid and non-gravid uteri. At no stage, there was a significant neural precursor cell expressed, developmentally
difference between the concentrations in the two sides (P > 0.05). downregulated 8 (NEDD8) protein, cullin-associated
Protein concentrations in the flushings from both uteri increased
progressively from d3 to d9 (P < 0.0001). The significant differences
NEDD8-dissociated protein 1(CAND1) and RNA binding
with reference to d0 are denoted by *. protein fused in sarcoma (FUS) were only seen in d0–d3
samples (Supplementary Table 2). Lectin, galactoside-
We identified 994 proteins in uterine flushings binding soluble 1 (LGALS1) and proliferating cell nuclear
based on MASCOT searches of the tammar genome antigen (PCNA), which regulates cell proliferation and
(Supplementary Table 1). Six hundred and three proteins differentiation, were observed only in the reactivation
were filtered based on minimum of two unique peptides samples (Supplementary Table 2).
and the protein to be present in minimum of three
biological replicates out of five (Supplementary Table 2).
Discussion
A classification based on the cellular location using
TargetP 1.1 (Emanuelsson et al. 2000) (Supplementary Earlier studies of the tammar UF using approaches
Table 2 column M – location) predicted 128 secretory such as gradient gel electrophoresis (Renfree 1973)
proteins, 24 mitochondrial proteins and 451 other and radioactive tracer studies (Shaw & Renfree 1986)
proteins. To assess the contribution of serum proteins demonstrated that there are stage-specific changes in
to UF, we conducted parallel proteomic analyses of the proteins present in the uterine secretions, but this
blood serum of tammar. We identified 47 proteins in is the first study using modern proteomics techniques
serum (Supplementary Table 3A); of these, all were also and provides new data on the presence of individual
found in UF, with albumin being the most significant proteins identified by MASCOT search (Perkins et al.
component. Although there is no tammar serum/plasma 1999) of the recently developed tammar genome. This
proteome database in the public domain, we were approach allowed us to study the qualitative changes
able to cross-reference to the human plasma proteome in protein composition in UF taking into consideration
database (Nanjappa et al. 2014) and identified 342 whether a protein was consistently found in at least
proteins that were present both in UF and the human three biological replicates during diapause, reactivation
plasma proteome database (Supplementary Table 3B). and up to late gestation.
Enzymes involved in glycolysis, gluconeogenesis, Studies in a range of eutherian and marsupial
pentose phosphate pathway, glycogenolysis, fatty acid species show that UF contains selectively transudated
metabolism, arachidonic acid metabolism, tricarboxylic serum proteins as well as proteins unique to the uterus
(TCA) cycle, galactose metabolism, creatinine and (Junge & Blandau 1958, Stevens et al. 1964, Renfree
retinol metabolism were identified in samples from d0 1973, Beier 1974). The protein repertoire of UF in
RPY until d11 RPY (Table 1) after d11 proteins involved tammar using gradient acrylamide gel electrophoresis
in adhesion, implantation and embryogenesis were identified numerous small uterine-specific pre-albumin
being identified (Supplementary Table 2). Comparison proteins (Renfree 1973) and similarly in human
of proteins in d0 and early stages of reactivation (d4–d6) secretions using two-dimensional polyacrylamide gel
showed that most of the proteins were present in all the electrophoresis (Maclaughlin et al. 1986, Hannan
stages except the cell cycle regulatory proteins that were et al. 2010). However, the studies in tammar did not
present only in d0 and growth factors only in stages reveal the identities of several UF proteins. Our study
d4–d6 (Supplementary Table 4). demonstrated the presence of many serum proteins in
Apart from the other types of proteins, by d11 RPY, the UF (Supplementary Table 3A). These may be passing
there were more proteins associated with cell adhesion though the endometrial epithelial barrier by selective
Reproduction (2016) 152 491–505 www.reproduction-online.org
Downloaded from Bioscientifica.com at 03/10/2022 04:29:27AM
via free accessTable 1 Metabolic enzymes identified across the different stages in tammar wallaby with their corresponding pathways.
Accession Protein name d0 d3 d4 d5 d6 d8 d9 d11 d24 Metabolic process
ENSMEUP00000006216 Peroxiredoxin 1 + + + + + + + + Detoxification of reactive oxygen species/selenium pathway
ENSMEUP00000007211 Carbonyl reductase NADPH 1 + + + + + + + + Arachidonic acid metabolism
ENSMEUP00000010739 Glutathione S-transferase mu 3 brain + + + + + + + + + Glutathione metabolism
ENSMEUP00000012361 Sepiapterin reductase + + + + + + + Tetrahydrobiopterin biosynthesis/folate biosynthesis
ENSMEUP00000004359 Phosphoglycerate kinase 1 + + + + + + + + Glycolysis and gluconeogenesis
ENSMEUP00000000879 Peroxiredoxin 5 + + + + Thioredoxin/selenium pathway, detoxification of ROS
ENSMEUP00000001389 Glutathione S-transferase P Glutathione metabolism/xenobiotic metabolism
www.reproduction-online.org
+ + + + + + + +
ENSMEUP00000006370 Triosephosphate isomerase 1 + + + + + + + + + Gluconeogenesis
ENSMEUP00000003207 Thioredoxin + + + + + + + Detoxification of ROS
ENSMEUP00000005432 Alpha enolase + + + + + + + + + Glycolysis
ENSMEUP00000001178 Creatine kinase B chain + + + + + + + + Urea cycle/amino acid metabolism
ENSMEUP00000000833 Carbonic anhydrase III muscle specific + Vitamin/coenzyme/sulphur compound metabolism
ENSMEUP00000007173 Phosphoglycerate kinase 1 + + + + + + + Glycolysis and gluconeogenesis
ENSMEUP00000013868 Dimethylarginine dimethylaminohydrolase 2 + + + Arginine metabolism/citrulline metabolism/nitric acid
biosynthesis
ENSMEUP00000006724 Alcohol dehydrogenase NADP + + + + + + + + Glycolysis/gluconeogenesis
ENSMEUP00000002721 Creatine kinase B chain + + + + + + + Vitamin/coenzyme/sulphur compound metabolism
ENSMEUP00000013946 Glyceraldehyde 3-phosphate dehydrogenase + + + + + + + + + Glycolysis/gluconeogenesis
Fragment EC 1.2.1.12
ENSMEUP00000012704 Glutathione peroxidase 2 gastrointestinal + + Lipid metabolism
ENSMEUP00000005468 Peroxiredoxin 2 + + + + + + + + Detoxification of reactive oxygen species/selenium pathway
ENSMEUP00000003077 Glutathione S-transferase theta2B + + + + + + + Glutathione metabolism
ENSMEUP00000004100 Pyruvate kinase muscle + + + + + + + + + Glycolysis/gluconeogenesis
ENSMEUP00000004497 Glutaredoxin thioltransferase + + + + + + + Electron transport chain
ENSMEUP00000002725 Cyclophilin B + + Prolactin signalling pathway/collagen biosynthesis
ENSMEUP00000004406 Lactate dehydrogenase B + + + + + + Carbohydrate metabolism
ENSMEUP00000013345 Catalase + + + + + + + + Electron transport chain
ENSMEUP00000005695 Sulphotransferase 1A1 + + + Steroid metabolism
ENSMEUP00000000775 Transaldolase 1 + + + + + + + + Pentose phosphate pathway
ENSMEUP00000008411 Fructosebisphosphate aldolase A + + + + + + + + Glycolysis/gluconeogenesis
ENSMEUP00000005681 Dihydropyrimidinase-like 2 + + + + + + + + BDNF signalling pathway
ENSMEUP00000001338 Lactate dehydrogenase A + + + + + TCA cycle/pyruvate metabolism
ENSMEUP00000002583 Cytosolic non-specific dipeptidase + + + + + + + Nitrogen metabolism/glutathione biosynthesis
ENSMEUP00000001121 Protein disulphide isomerase family A member 3 + + + + + + + Detoxification of ROS/cellular protein metabolism
ENSMEUP00000005305 Esterase D + + + + + + + + Oestrogen/progesterone biosynthesis
ENSMEUP00000013368 Protein phosphatase 2 regulatory subunit A alpha + + + MAPK signalling, cell cycle and mitosis
ENSMEUP00000012631 Glutathione peroxidase 3 plasma + Detoxification of ROS
ENSMEUP00000012662 Gamma-glutamyl hydrolase conjugase + Glutamine metabolism
folylpolygammaglutamyl hydrolase
ENSMEUP00000014632 Glutathione peroxidase 1 + + lipid metabolism
ENSMEUP00000009000 Isocitrate dehydrogenase 1 NADP + + + TCA cycle/phospholipid biosynthesis
ENSMEUP00000012664 Fructose bisphosphate aldolase C + Glucose metabolism/gluconeogenesis
ENSMEUP00000007804 Glutathione S-transferase theta + + + Glutathione metabolism
ENSMEUP00000011838 Phosphoglycerate mutase 1 + + + + + + Glucose metabolism/gluconeogenesis
ENSMEUP00000005036 Peroxiredoxin 4 + + + + + + + + Detoxification of reactive oxygen species
ENSMEUP00000011992 d-dopachrome decarboxylase + + + + + + Melanin biosynthetic process
ENSMEUP00000001181 Prostaglandin E synthase 3 cytosolic + + + + Arachidonic acid metabolism/prostaglandin biosynthesis
ENSMEUP00000008081 Aldehyde dehydrogenase 1 family member A + + + + + + + + Retinol metabolism
Uterine flushing proteome of tammar wallaby
ENSMEUP00000005444 Acetyl-CoA acetyltransferase 2 + + + Mevolonate pathway
ENSMEUP00000013692 Galactose mutarotase aldose 1-epimerase + + Carbohydrate metabolism
ENSMEUP00000015288 Prolyl 4-hydroxylase beta polypeptide + + Redox homeostasis
Reproduction (2016) 152 491–505
495
(Continued)
via free access
Downloaded from Bioscientifica.com at 03/10/2022 04:29:27AMTable 1 Continued. 496
Accession Protein name d0 d3 d4 d5 d6 d8 d9 d11 d24 Metabolic process
ENSMEUP00000009171 Cytidine monophosphate UMP-CMP kinase 1 + + Purine & pyrimidine metabolism
cytosolic
ENSMEUP00000009302 Glucose-6-phosphate isomerase + + + + Glycolysis
ENSMEUP00000010438 Glutamine–fructose-6-phosphate transaminase + + + UDP-N-acetylglucosamine metabolism/protein metabolism
ENSMEUP00000014039 Methylthioadenosine phosphorylase + + + + + + Purine metabolism
ENSMEUP00000007229 Enolase 3 beta muscle + + + + + + + Glycolysis/gluconeogenesis
ENSMEUP00000001514 Protein phosphatase 2 catalytic subunit alpha + + + Meiotic/mitotic cell cycle/wnt signalling pathway
isozyme
F C Martin and others
Reproduction (2016) 152 491–505
ENSMEUP00000013706 Phosphoglucomutase 1 + + + Glucose/galactose/glycogen metabolism/pentose phosphate
pathway
ENSMEUP00000003732 Peroxiredoxin 6 + + + + + Phenylalanine metabolism
ENSMEUP00000005004 Puromycin-sensitive aminopeptidase + + + + + Hydrolysis of amino acids
ENSMEUP00000009792 Acid phosphatase 1 soluble + + + + Phosphatase activity
ENSMEUP00000009128 6-Phosphogluconolactonase + + + + + Glucose metabolism/pentose phosphate pathway
ENSMEUP00000004425 N-acetylneuraminate synthase + + + Oligosaccharide/lipopolysaccharide biosynthesis
ENSMEUP00000001387 Glutathione S-transferase theta-2 + + + Glutathione metabolism
ENSMEUP00000002646 Quinoid dihydropteridine reductase + + Phenylalanine and tyrosine metabolism/folate biosynthesis
ENSMEUP00000000713 Aminoacyl-tRNA synthetase class II + + tRNA modification/processing
ENSMEUP00000015196 Carnosine synthase 1 + Carnosine biosynthesis pathway
ENSMEUP00000008483 Aldehyde dehydrogenase 1 family member L1 + + 10-Formyltetrahydrofolate catabolic process/one carbon
metabolic process
ENSMEUP00000006508 3-Hydroxybutyrate dehydrogenase type 2 + Siderophore metabolism/beta fatty acid oxidation/ketone
biosynthesis
ENSMEUP00000006320 Alanine tRNA ligase + tRNA modification/processing
ENSMEUP00000002574 Methionine adenosyltransferase II alpha + + + + S-adenosylmethionine biosynthetic process
ENSMEUP00000013099 Protease serine 8 + Positive regulation of sodium ion transport
ENSMEUP00000005092 Aspartylglucosaminidase + + + Protein deglycosylation and maturation
ENSMEUP00000005423 UDP-glucose 6-dehydrogenase + + Lipopolysaccharide biosynthesis
ENSMEUP00000012604 Prostaglandin reductase-2 + + Prostaglandin metabolism
ENSMEUP00000013328 GDP-mannose 46-dehydratase + + + Nucleotide sugar biosynthesis
ENSMEUP00000013481 Superoxide dismutase 1 soluble + + + + + Detoxification of oxygen species/activation of MAPK activity
ENSMEUP00000005428 Dihydropyrimidinase-like 3 + + + + Regulation of cell migration
ENSMEUP00000003945 Peptidase D + + Amino acid metabolism/collagen catabolism
ENSMEUP00000013977 Quiescin Q6 sulfhydryl oxidase 1 + + + + + + Growth regulation
ENSMEUP00000010353 Leukotriene A4 hydrolase + + + + Fatty acid metabolism
ENSMEUP00000005926 Fumarylacetoacetate hydrolase + + Arginine/phenylalanine/tyrosine metabolism
fumarylacetoacetase
ENSMEUP00000004615 UDP-GlcNAcbetaGal beta-13-N- + Carbohydrate/glycosaminoglycan metabolism
Acetylglucosaminyltransferase 2
ENSMEUP00000004865 Protein phosphatase 1 catalytic subunit alpha + Glycogen metabolism, triglyceride catabolism/cell division
isozyme and cell cycle
ENSMEUP00000007839 Methionine sulphoxide reductase A + Methionine metabolism
ENSMEUP00000000045 ADP-ribosylhydrolase like 2 + Cellular response to superoxide
ENSMEUP00000006354 Ubiquitin specific peptidase 5 isopeptidase T + Ubiquitin-dependent protein catabolism
ENSMEUP00000003136 Aldo-keto reductase family 1 member B1 + + + + + Carbohydrate/doxorubicin/sorbitol metabolism
ENSMEUP00000001600 Glycyl-tRNA synthetase + tRNA modification/processing
ENSMEUP00000011680 Phosphopantothenoylcysteine synthetase + Coenzyme A/pantothenate/vitamin metabolism
ENSMEUP00000007002 Protein disulphide isomerase family A member 4 + + + + Detoxification of ROS/cellular protein metabolism
ENSMEUP00000004464 Aldehyde dehydrogenase mitochondrial + + + + Carbohydrate/xenobiotic metabolism
ENSMEUP00000004313 Prenylcysteine oxidase 1 + Terpenoid synthesis
ENSMEUP00000015108 Carbonic anhydrase II + + + Vitamin/coenzyme/sulphur compound metabolism
www.reproduction-online.org
ENSMEUP00000009844 ATP citrate lyase + + TCA cycle
ENSMEUP00000011277 Phosphoglucomutase 2 + + + + + + Glycogen/carbohydrate metabolism
via free access
Downloaded from Bioscientifica.com at 03/10/2022 04:29:27AMENSMEUP00000001078 Prolyl endopeptidase + + Proteolysis
ENSMEUP00000013173 Lysosomal alpha-mannosidase + Protein modification/mannose metabolism
ENSMEUP00000002183 Phospholipase A2 group XV + Ceramide/glycerophospholipid/phosphatidylcholine
metabolism, fatty acid catabolism
ENSMEUP00000010449 Enolase-phosphatase 1 + Methionine biosynthesis
ENSMEUP00000014768 Malate dehydrogenase mitochondrial + Carbohydrate metabolism/TCA cycle
ENSMEUP00000013511 Uroporphyrinogen decarboxylase + Uroporphyrinogen III metabolism/small molecule metabolism
ENSMEUP00000000258 Aflatoxin B1 aldehyde reductase member 2 + Carbohydrate metabolism/Xenobiotic metabolism
ENSMEUP00000005261 Prostaglandin-E2 9-reductase + + Prostaglandin biosynthesis
ENSMEUP00000007181 Hexosaminidase B beta polypeptide + Carbohydrate metabolism/sphingolipid metabolism
www.reproduction-online.org
ENSMEUP00000000995 Arylacetamide deacetylase esterase 17 + Triglyceride catabolism, xenobiotic metabolism
ENSMEUP00000009032 Glutathione reductase + Glutathione metabolism/destruction of ROS
ENSMEUP00000000895 Ectonucleotide pyrophosphatasephospho + Phospholipid catabolism, cell migration
diesterase 2
ENSMEUP00000007146 Nuclear casein kinase and cyclin-dependent + Glucose homeostasis, insulin receptor signalling pathway
kinase substrate 1
ENSMEUP00000006299 Glutaminyl-tRNA synthetase + tRNA modification/processing
ENSMEUP00000002407 Alcohol dehydrogenase 5 class III chi polypeptide + Destruction of ROS
ENSMEUP00000012523 Carboxypeptidase N polypeptide 1 + Bradykinin catabolism
ENSMEUP00000013478 Aconitase 1 soluble + + + + TCA cycle
ENSMEUP00000004870 Lecithin-cholesterol acyltransferase + Cholesterol/lipoprotein metabolism
ENSMEUP00000006047 Phosphofructokinase liver + Carbohydrate metabolism
ENSMEUP00000001439 Tryptase + Endopeptidase activity
ENSMEUP00000014818 Galactosidase beta 1 + Carbohydrate metabolism/sphingolipid metabolism
ENSMEUP00000002036 Tryptophanyl-tRNA synthetase + Angiogenesis
ENSMEUP00000002335 Aldehyde dehydrogenase 1 family member A3 + + + Retinol metabolism
ENSMEUP00000000288 Phosphorylase glycogen brain + Carbohydrate metabolism
ENSMEUP00000006031 Protease serine 35 + Serine protease
ENSMEUP00000011456 X-prolyl aminopeptidase aminopeptidase P 1 + Bradykinin catabolism
soluble
ENSMEUP00000000536 Serine hydroxymethyltransferase 1 soluble + Carnitine biosynthesis, vitamin/nitrogen/folic acid metabolism
ENSMEUP00000008533 Leucine carboxyl methyltransferase 1 + Protein modification/regulation of mitotic cell cycle spindle
assembly
ENSMEUP00000001467 Peptidylprolyl isomerase F + + + + + Negative regulation of oxidative phosphorylation
ENSMEUP00000007250 Aspartyl aminopeptidase + Peptide metabolism
ENSMEUP00000003548 Glutamyl-prolyl-tRNA synthetase + Protein biosynthesis
ENSMEUP00000004150 Deoxyribose-phosphate aldolase putative + Carbohydrate/deoxyribosenucleotide catabolism
ENSMEUP00000004805 N-acylaminoacyl-peptide hydrolase + Endopeptidase activity
ENSMEUP00000011308 Dihydropyrimidine dehydrogenase + Beta-alanine biosynthesis, purine-pyrimidine catabolism
ENSMEUP00000001344 Protein disulphide isomerase family A member 6 + + Detoxification of ROS/cellular protein metabolism
ENSMEUP00000015002 Methylenetetrahydrofolate dehydrogenase + One carbon/tetrahydrofolate metabolism
ENSMEUP00000003224 Glutamate-cysteine ligase catalytic subunit + Xenobiotic/glutathione/sulphur amino acid/cysteine/glutamate
metabolism
ENSMEUP00000008144 UDP-N-acteylglucosamine pyrophosphorylase 1 + Uridylyltransferase activity
ENSMEUP00000003820 Mitogen-activated protein kinase 1 + Activation of MAPK activity/negative regulator of cell
differentiation/apoptosis/cell cycle
ENSMEUP00000015019 Histidyl-tRNA synthetase + Protein biosynthesis
ENSMEUP00000009382 Aldehyde dehydrogenase 9 family member A1 + + + + Carnitine biosynthesis/nitrogen metabolism
ENSMEUP00000010778 Tyrosyl-tRNA synthetase + Protein biosynthesis
ENSMEUP00000010557 Protein disulphide isomerase + Detoxification of ROS
ENSMEUP00000011501 Ubiquitin specific peptidase 14 tRNA-guanine + Ubiquitin-dependent protein catabolism
Uterine flushing proteome of tammar wallaby
transglycosylase
ENSMEUP00000010927 Mannosidase alpha class 1A member 1 + Protein modification
ENSMEUP00000007934 Lysyl-tRNA synthetase + Protein biosynthesis
ENSMEUP00000005076 Alpha-lactalbumin + Carbohydrate metabolism
Reproduction (2016) 152 491–505
497
via free access
Downloaded from Bioscientifica.com at 03/10/2022 04:29:27AM498 F C Martin and others
Table 2 Growth factors and associated binding proteins that may be involved in regulating diapause and later pregnancy that have been
identified from d3 RPY to d24 RPY uterine flushings of the tammar wallaby.
Growth factors Stage present (after RPY) References
Hepatoma-derived growth factor d3, d4–d5, d6–d9, d11 Gómez et al. (2012, 2014), Muñoz et al. (2014)
Granulin d8, d9 Gerton et al. (2000), Bateman et al. (2003)
Insulin-like growth factor binding proteins 1,2,3,4,5,6,7 d24 Simmen et al. (1995), Bagnell et al. (1997),
Seidel et al. (1998), Costello et al. (2014)
Transforming growth factor beta-2 d24 Li et al. (2014, 2015)
MMP-2 d24 Howe et al. (1999), Aslan et al. (2007)
Growth differentiation factor 15 d24 Trovik et al. (2014),Chudecka-Głaz et al. (2015)
sEGFR d3–d4, d8, d11 Perez-Torres et al. (2008), Adamczyk et al. (2011),
Maramotti et al. (2012)
Insulin-like growth factor 2 d24 Geisert et al. (2001), Costello et al. (2014)
entry or apocrine secretions. There is also evidence contain lipids, proteins or micro-(mi)RNA are acquired
to demonstrate localisation of some of these serum by the endometrial epithelial cells or the blastocyst to
proteins in the endometrium (DeSouza et al. 2005, promote implantation (Valadi et al. 2007, Simpson et al.
Fowler et al. 2007). In marsupials and primates, uterine- 2008). Exosomal marker proteins include CD9, CD63
specific pre-albumins may pass into the blastocyst and and CD81 along with heat-shock proteins HSP70 and
exert some effect on its growth (Renfree 1973, 1978, HSP90 (Mathivanan et al. 2010). Among the exosomal
Peplow et al. 1974, Hearn & Renfree 1975), but this proteins reported in earlier studies, glyceraldehyde
idea has not been tested directly. The predictions by phosphate dehydrogenase (GAPDH), tyrosine
TargetP 1.1 revealed that 21% of the proteins in UF are 3-monooxygenase/tryptophan 5-monooxygenase
classed as secretory proteins. activation protein zeta polypeptide (YWHAZ), HSP70
Exosomes and microvesicles derived from endometrial and high mobility group B1 (HMGB1) have been
epithelium or trophectoderm have been identified in identified in our study. However, given that the tammar
human (Kshirsagar et al. 2012, Ng et al. 2013, Tannetta blastocyst is enclosed in an acellular shell coat for the
et al. 2014) and ovine UF (Racicot et al. 2012, Burns first 18 days of pregnancy, it has yet to be determined
et al. 2014), which provide an alternative mode of whether microsome- or exosome-mediated transfer from
maternal embryo communication. Exosomes that the uterus to the blastocyst is possible in this species.
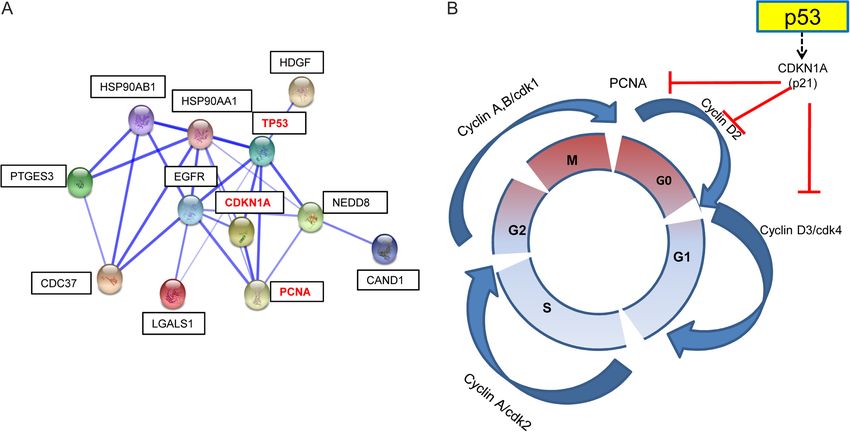
Figure 3 (A) A protein interaction map of some of the factors identified in UF. p53, PCNA and CDKN1A are the main players in this regulatory
pathway. CDKN1A induction by p53 leads to cdk2 inhibition, thereby inhibiting cellular and blocking cell cycle progression. STIP1, HSPs
and CDC37 act as molecular chaperones promoting their binding and stabilisation of protein complexes. NEDD8 and CAND1 also play
important roles in cell cycle control by degradation of cyclins and other regulatory proteins. PTGES3 in a hormone-dependent manner disrupts
receptor-mediated transcription. PCNA interaction with DNA polymerases is sensitive to changes in CDKN1A levels. (B) A kinase-dependent
p53/CDKN1A regulatory pathway controls the cell cycle.
Reproduction (2016) 152 491–505 www.reproduction-online.org
Downloaded from Bioscientifica.com at 03/10/2022 04:29:27AM
via free accessUterine flushing proteome of tammar wallaby 499
Pregnancy is also regulated by cytoskeleton-associated the uterus, this may be achieved by local action of
proteins involving cytoskeletal rearrangements, enzymes in the uterine fluid. This reducing environment
apoptosis and constant remodelling (Lee et al. 1998, is preserved by enzymes that maintain the reduction–
Miehe et al. 2005), which may in turn regulate the oxidation (REDOX) state through a constant input of
developing embryo (Jensen et al. 2013). Keratins are metabolism-derived energy (Gilbert & Colton 1999).
usually considered as a source of contamination during Cells have several mechanisms to protect against reactive
proteomic procedures, but 51 specific keratins were oxygen species (ROS). Prolonged, experimentally
detected in pre-implantation mouse embryos, and induced ROS production severely inhibits embryo
several other studies confirm the presence of keratins development (Johnson & Nasresfahani 1994, Guerin
and cytokeratins in the early mammalian embryonic et al. 2001), and its regulation is necessary for optimal
development (Lehtonen et al. 1983, Magin et al. 2000). embryo growth (Burdon 1996). The apocrine secretion
The changes in the keratin components in UF in our of enzymes identified by our study (Table 1), which
study possibly reflect changes resulting from the cellular catalyse the destruction of ROS, are transcribed in pre-
differentiation of the endometrium. Adhesion protein implantation embryos (Harvey et al. 1995, Takahashi
Vanin 3 was specifically found in d11RPY UF samples 2012, Ramos et al. 2015). REDOX enzymes also
in the tammar previously known to be present on the known to support cell cycle progression (Yamauchi &
surface of equine endometrium (Hayes et al. 2012). Bloom 1997). Glutathione peroxidase, identified in our
Furthermore, synthesis of several classes of proteins, study, is an important REDOX regulator and is found
including adhesion factors, cytokines and growth in reproductive tract fluid (Gardiner et al. 1998) and
factors, increases immediately after the oestrogen pulse decreases in concentration as early cleavage proceeds
that induces mouse implantation (Dey et al. 2004) as (Gardiner & Reed 1995). A family of peroxiredoxins
well as at reactivation from diapause in the tammar (Cha identified in our study is known to be secreted by
et al. 2013). Several ECM components associated with endometrial epithelium into the uterine lumen in mice
apocrine secretion from the glandular epithelium (Demir and play crucial roles as antioxidants in the development
et al. 2002) are up-regulated in the pre-implantation of pre-implantation mouse embryos (Wang et al. 2010,
endometrium in mice, including fibronectin, laminin Bhutada et al. 2013). Another autocrine secretory factor
and collagen type IV (Armant et al. 1986, Carson et al. cyclophilin A, commonly seen during oxidative stress
1988). These ECM proteins were observed in tammar (Jin et al. 2000), was observed in all UFs especially more
UFs during diapause and most of the reactivation stages in days d0–d3 and early reactivation stages.
and not limited to any stage. Progesterone and oestradiol induce changes in the
HMGB1 acts in an autocrine/paracrine fashion, uterine environment and the production and release
and immunohistochemical analysis of human and rat of some cytokines and growth factors from the uterus
endometrial tissue revealed the higher levels of HMGB1 that can have both autocrine and paracrine actions to
in the nuclei of glandular epithelial, stromal and luminal regulate the pre-implantation embryo and prepare the
epithelial compartments during the pre-receptive phase endometrium for implantation (Harvey et al. 1995,
compared with their counterparts in the receptive Sharkey 1998, Hannan et al. 2011, Binder et al. 2014.
phase in human (Cui et al. 2008, Bhutada et al. 2014). Nineteen proteins previously shown to be regulated by
HMGB1/p53 complex are known to regulate autophagy progesterone and oestradiol in human endometrial and
and apoptosis (Livesey et al. 2012). In the tammar, myometrial cells (Tamm et al. 2009, Soloff et al. 2011)
HMGB1 was present from d0 until d11RPY, and high were identified in tammar UF (Supplementary Table 5).
mobility group B2 (HMGB2) was present in d0–d3 and Several growth factors play important roles during
d11 of pregnancy. pre-implantation mammalian development. Leukaemia
UF contains many enzymes involved in important inhibitory factor (LIF), insulin-like growth factors (IGFs),
metabolic pathways that have been studied in the uterus epidermal growth factor (EGF), platelet-derived growth
and embryo (Murdoch & White 1968, Kirchner et al. factor (PDGF), fibroblast growth factor (FGF), platelet
1971, Denker & Petzoldt 1977, Peplow 1982, Zavy activating factor (PAF), vascular endothelial growth
et al. 1984). During diapause, the blastocyst remains factor (VEGF) and transforming growth factor-β (TGF-β)
viable but has a low metabolic rate that must provide are present in the uterus and influence the development
sufficient energy for the maintenance and homeostasis and growth of the pre-implantation embryo in several
of the embryo during the long period of arrest (Spindler species including mouse, rat, cow and sheep and tammar
et al. 1995, 1997, 1999). The first significant increase (Thouas et al. 2015). Roles of growth factors during early
in blastocyst metabolism is at day 4 RPY when the development have been demonstrated by the addition
first mitoses are seen in the blastocyst, subsequent to of purified growth factors to culture medium or by
activation of the CL and progesterone stimulation of the molecular genetic techniques that interfere with gene
uterus (Spindler et al. 1995, 1997). To maintain viability expression and necessary for successful development of
during diapause, it is important for the blastocyst to avoid the blastocyst. A few of the previously identified proteins
damage from reactive oxygen species (ROS). Within like progestagen-associated endometrial protein (PAEP),
www.reproduction-online.org Reproduction (2016) 152 491–505
Downloaded from Bioscientifica.com at 03/10/2022 04:29:27AM
via free access500 F C Martin and others
LIF and uteroglobin found in UF (Renfree 1973, Li et al.
1993) were not identified by our study likely due to them Growth factors like HDGF
Blastocyst
and sEGFR
below the detection limit of this technique. Epidermal
growth factor receptor identified in our study is important
in intricate signalling and transcriptional networks hormones &
growth factors
(Large et al. 2014), which regulate diverse cellular
functions, promoting cell proliferation, differentiation, Secretion of
enzymes & nutrients
Transudation of Plasma
& byproducts of cellular
migration, cell growth, and survival. These receptors differentiation
bind epidermal growth factors (EGFs), a mitogen known
to terminate diapause in ovariectomized rats in the
absence of the oestrogen pulse (Johnson & Chatterjee
1993). Oestrogen stimulates the expression of the EGF
family, and EGF is a potent mitogen expressed in the
uterus during implantation in the mouse (Dey et al. Uterine Stromal tissue
2004). EGF receptors are present in truncated forms
Figure 4 Summary of the uterine embryo interactions that occur
on porcine endometrium during d9–d11 of pregnancy during reactivation from diapause in the tammar. The blastocyst is
(Kliem et al. 1998) and in dormant carnivore embryos surrounded by an acellular coat, so uterine control of the blastocyst
(Paria et al. 1994). Upregulation of epidermal growth must be mediated by soluble factors that can pass through this
factor receptor (EGFR) signalling is often observed in barrier. Uterine flushings contain significant quantities of enzymes,
carcinomas and promotes uncontrolled cell proliferation products of cellular differentiation and growth factors from the day
and metastasis. The soluble forms (sEGFR) are diagnostic of reactivation (d4 RPY), which increase up to late gestation.
The absence or low concentrations of growth factors during diapause
and/or prognostic cancer biomarkers (Perez-Torres et al. and presence of cell cycle regulatory proteins that causes cell cycle
2008, Adamczyk et al. 2011, Maramotti et al. 2012). arrest at G0/G1 phase may maintain diapause, whilst the surge of
The soluble secretory forms of EGFR were present in the progesterone from the CL at reactivation induces the release of
tammar UF from d3 onwards correlating with the start of specific growth regulators from the endometrium that lead to
cell proliferation. reactivation of the blastocyst.
Hepatoma-derived growth factor (HDGF), a
component of p53/p21 cell cycle control pathway, cycle arrest. A p53/p21 cell cycle inhibition pathway
mediates cell proliferation, is activated by mitotic was evident in all diapause samples (d0 and d3) by
phosphorylation (Everett 2011) and secretion of HDGF the presence of p21-associated proteins that mediate
requires processing of the N-terminus (Thakar et al. cell cycle arrest at G0/G1 phase. This cell cycle
2010). HDGF was absent at d0 but was identified from arrest pathway was illustrated by STRING 10 using
day 3 RPY onwards. Thus, it is the earliest specific equivalent proteins from human (Szklarczyk et al.
secretory change identified, and may be associated 2010) (Fig. 3A). Different domains of p21 interact
with the start of cell proliferation (Fig. 3A). HDGF in with Cdks and PCNA, and both of these domains can
bovine UF promotes embryonic development and cell independently inhibit DNA replication when present
proliferation and is synthesized by the endometrium in cells (Fig. 3B) (Cayrol et al. 1998). The presence
and embryo (Gomez et al. 2014), therefore performing of LGALS1 at d3–d4 and PCNA at d6 by autocrine
a dual role by receptor-mediated action or directly by secretions indicates their roles in DNA replication
DNA binding. and cell cycle control of the endometrium during
Fibroblast growth factors activate mink embryos from reactivation. These findings are consistent with those
diapause and there is a gradual increase from day 3 after identified in the mouse embryo in which dormancy
activation (Desmarais et al. 2004). Most of the growth is associated with the decrease in DNA replication
factors identified have roles in malignancy due to their genes (Hamatani et al. 2004).
involvement in proliferation and metastasis (Witsch et al. Future studies of the validation of these growth
2010). The absence, or low concentrations, of specific factors and other molecules of interest in the UF
growth factors and the presence of cell cycle inhibitors could be carried out using MALDI (Matrix Absorption
during lactation and up to d3 RPY in the tammar may be Laser Desorption Ionisation Time of Flight) Imaging
a potent reason the blastocyst remains dormant (Fig. 4). Mass Spectrometry (MALDI IMS) (Caprioli et al.
Reduction or cessation of mitotic activity in 1997) coupled to LC–MS/MS experiments. IMS is
the embryo is controlled by cell cycle regulatory advantageous over other techniques since it does
mechanisms (Lopes et al. 2004). The cell cycle arrest not require target-specific labelling reagents such
markers may cause activation of mitogen-activated as antibodies or tissue homogenization and utilizes
protein (MAP) kinase signal pathway (Moscatello intact tissue, which enables to correlate molecular
et al. 1998), downregula tion of cyclins or cyclin- information with histological details. This technique
dependent kinases (cdks) (Kim et al. 1999, Wang et al. can help to localise specific protein signatures
2013), which leads to upregulation of p21 and cell within different tissue compartments, thus preserving
Reproduction (2016) 152 491–505 www.reproduction-online.org
Downloaded from Bioscientifica.com at 03/10/2022 04:29:27AM
via free accessUterine flushing proteome of tammar wallaby 501
the spatial localisation of the molecules of interest References
(Balluff et al. 2011, Fehniger et al. 2014). Comparison Adamczyk KA, Klein-Scory S, Tehrani MM, Warnken U, Schmiegel W,
of the peak list from MALDI IMS for identification Schnölzer M & Schwarte-Waldhoff I 2011 Characterization of soluble
of proteins using a parallel liquid chromatography and exosomal forms of the EGFR released from pancreatic cancer cells.
Life sciences 89 304–312. (doi:10.1016/j.lfs.2011.06.020)
(LC)–MS approach enables the identification of Aitken RJ 1974 Delayed Implantation in Roe deer (Capreolus-
hundreds of proteins (Schober et al. 2012, Cole et al. capreolus). Journal of Reproduction and Fertility 39 225. (doi:10.1530/
2013, Franck et al. 2013). jrf.0.0390225)
Ametzazurra A, Matorras R, García-Velasco JA, Prieto B, Simón L,
Martínez A & Nagore D 2009 Endometrial fluid is a specific and
non-invasive biological sample for protein biomarker identification
Conclusion in endometriosis. Human Reproduction 24 954–965. (doi:10.1093/
humrep/den450)
The proteomics approach using multiple biological Armant DR, Kaplan HA & Lennarz WJ 1986 Fibronectin and laminin
replicates to characterise the UF proteome from d0 promote invitro attachment and outgrowth of mouse blastocysts.
until late gestation coupled with LC–MS/MS analysis Developmental Biology 116 519–523. (doi:10.1016/0012-1606(86)
90152-1)
provided a global profile of proteins in the UFs and Balluff B, Schöne C, Höfler H & Walch A 2011 MALDI imaging mass
provides evidence for autocrine, paracrine and spectrometry for direct tissue analysis: technological advancements
apocrine pathways. The data from our study collectively and recent applications. Histochemistry and Cell Biology 136 227–244.
suggest that p21 may be responsible for the inhibition (doi:10.1007/s00418-011-0843-x)
Beier HM 1974 Oviducal and uterine fluids. Journal of Reproduction and
of the cell cycle in the uterine epithelium, thus Fertility 37 221–237. (doi:10.1530/jrf.0.0370221)
preventing growth factor synthesis, but once diapause Bhutada S, Katkam RR, Nandedkar T, Metkari SM, Chaudhari UK,
is terminated, numerous growth factors including Varghese S, Kholkute SD & Sachdeva G 2013 Uterine secretome and
HBGF and EGFR may have a role in reactivation of the its modulation in rat (Rattus norvegicus). Reproduction 146 13–26.
(doi:10.1530/REP-12-0461)
diapausing blastocyst. Bhutada S, Basak T, Savardekar L, Katkam RR, Jadhav G, Metkari SM,
Chaudhari UK, Kumari D, Kholkute SD, Sengupta S et al. 2014 High
mobility group box 1 (HMGB1) protein in human uterine fluid and
its relevance in implantation. Human Reproduction 29 763–780.
Supplementary data (doi:10.1093/humrep/det461)
This is linked to the online version of the paper at http://dx.doi. Binder NK, Evans J, Gardner DK, Salamonsen LA & Hannan NJ 2014
Endometrial signals improve embryo outcome: functional role of
org/10.1530/REP-16-0154. vascular endothelial growth factor isoforms on embryo development
and implantation in mice. Human Reproduction 29 2278–2286.
(doi:10.1093/humrep/deu211)
Declaration of interest Boomsma CM, Kavelaars A, Eijkemans MJC, Amarouchi K, Teklenburg G,
Gutknecht D, Fauser BJCM, Heijnen CJ & Macklon NS 2009 Cytokine
The authors declare that there is no conflict of interest that profiling in endometrial secretions: a non-invasive window on
could be perceived as prejudicing the impartiality of the endometrial receptivity. Reproductive BioMedicine Online 18 85–94.
(doi:10.1016/S1472-6483(10)60429-4)
research reported. Burdon RH 1996 Control of cell proliferation by reactive oxygen species.
Biochemical Society Transactions 24 1028–1032. (doi:10.1042/
bst0241028)
Funding Burns G, Brooks K, Wildung M, Navakanitworakul R, Christenson LK &
Spencer TE 2014 Extracellular vesicles in luminal fluid of the ovine
This research was funded by the Australian Research Council uterus. Plos One 9 e90913. (doi:10.1371/journal.pone.0090913)
(ARC Discovery project grant # DP110101727, awarded to Caprioli RM, Farmer TB & Gile J 1997 Molecular imaging of biological
samples: localization of peptides and proteins using MALDITOF MS.
G S and D K G). Analytical Chemistry 69 4751–4760 (doi:10.1021/ac970888i)
Carson DD, Tang JP & Gay S 1988 Collagens support embryo attachment and
outgrowth invitro – effects of the Arg-Gly-Asp sequence. Developmental
Acknowledgements Biology 127 368–375. (doi:10.1016/0012-1606(88)90323-5)
Casado-Vela J, Rodriguez-Suarez E, Iloro I, Ametzazurra A, Alkorta N,
The authors thank Scott Brownlees, Helen Clark and the García-Velasco JA, Matorras R, Prieto B, González S, Nagore D et al.
entire wallaby research group for assistance with the 2009 Comprehensive proteomic analysis of human endometrial fluid
aspirate. Journal of Proteome Research 8 4622–4632. (doi:10.1021/
wallabies. They thank their collaborators Associate Professor pr9004426)
Andrew Pask, Professor Asao Fujiyama (National Institute of Cayrol C, Knibiehler M & Ducommun B 1998 p21 binding to PCNA
Informatics, Japan), Professor Rachel O’Neill and Mr Tom causes G1 and G2 cell cycle arrest in p53deficient cells. Oncogene 16
Heider (University of Connecticut) for their contributions 311–320. (doi:10.1038/sj.onc.1201543)
Cha JXS, Bartos A, Fenelon J, Lefèvre P, Daikoku T, Shaw G, Maxson R,
to the generation of an updated tammar genome that was
Murphy BD, Renfree MB & Dey SK 2013 A new role for muscle segment
used to derive the proteome database used in this study. They homeobox genes in mammalian embryonic diapause. Open Biology 3
are also grateful to David Perkins at the proteomics facility 130035. (doi:10.1098/rsob.130035)
Bio21 institute for setting up the tammar protein database Chang MC 1968 Reciprocal insemination and egg transfer between ferrets
and enabling automated BLAST searches. and mink. Proceedings of the 12th international Congress of Genetics,
Tokyo 1 279–279.
The mass spectrometry proteomics data can be publicly Clemetson CAB, Verma UL & De Carlo SJ 1977 Secretion and reabsorption
accessed at the ProteomeXchange Consortium via the PRIDE of uterine luminal fluid in rats. Journal of Reproduction and Fertility 49
database. 183–187. (doi:10.1530/jrf.0.0490183)
www.reproduction-online.org Reproduction (2016) 152 491–505
Downloaded from Bioscientifica.com at 03/10/2022 04:29:27AM
via free accessYou can also read

















































