Two New Species of Batrisini (Coleoptera: Staphylinidae: Pselaphinae) from Nanling Mountain Area, China
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

insects
Communication
Two New Species of Batrisini (Coleoptera: Staphylinidae:
Pselaphinae) from Nanling Mountain Area, China †
Wen-Xuan Zhang and Zi-Wei Yin *
Laboratory of Systematic Entomology, College of Life Sciences, Shanghai Normal University, 100 Guilin Road,
Xuhui District, Shanghai 200234, China; zhangwx@protonmail.com
* Correspondence: pselaphinae@gmail.com
† This published work and the nomenclatural acts it contains have been registered in ZooBank, the online
registration system for the ICZN (International Code of Zoological Nomenclature). The LSID (Life Science
Identifier) for this publication is: urn:lsid:zoobank.org:pub:63804F59-A030-4EF4-A242-33C2BDAC6010.
Simple Summary: The Nanling Mountains are the largest mountain range and an important phys-
ical geographical boundary in Southeast China, but their insect diversity has not been sufficiently
documented up to now. Through recent investigations of the local staphylinid fauna at Nanling,
more than 4500 adult pselaphine beetles were collected and most of the samples belong to the highly
diverse tribe Batrisini. Here, two new species of the genera Batrisceniola and Physomerinus, both
poorly represented in the Chinese pselaphine fauna, are described.
Abstract: Two new species of the Batrisocenus complex of genera, e.g., Batrisceniola nanlingensis sp.
nov. and Physomerinus clavipes sp. nov., are described, diagnosed, and illustrated from the Nanling
Mountain Area. Both represent a second species of the respective genus from the Chinese mainland.
Keywords: Batrisceniola; Physomerinus; new species; Nanling; China; taxonomy
Citation: Zhang, W.-X.; Yin, Z.-W.
Two New Species of Batrisini
(Coleoptera: Staphylinidae:
Pselaphinae) from Nanling Mountain 1. Introduction
Area, China. Insects 2022, 13, 119.
As one of the 32 inland territorial and aquatic biodiversity conservation priority
https://doi.org/10.3390/
areas [1], Nanling Mountain Area is the largest mountain system as well as an important
insects13020119
geographical boundary in southern China. It is also the largest oasis near a latitude of
Academic Editor: Paulo A. V. 25 degrees north and harbors a high diversity of plants and animals [2]. Through a joint
Borges project led by Prof. Xing-Ke Yang, we had an opportunity to collect more than 4500 adult
pselaphine beetles in most conservation areas of Nanling. Approximately 68% of the
Received: 28 December 2021
Accepted: 18 January 2022
sampled specimens belong to the diverse tribe Batrisini. In contrast to this number, only
Published: 24 January 2022
16 species classified in 9 genera of the tribe are currently known to occur in Nanling, which
indicates that the true diversity of this group still remains poorly documented.
Publisher’s Note: MDPI stays neutral The Oriental genus Batrisceniola Jeannel, 1958 is represented by four species from
with regard to jurisdictional claims in
China (1 sp.) and Japan (3 spp.): B. dissimilis (Sharp, 1874), B. semipunctulata (Raffray, 1909),
published maps and institutional affil-
B. hiranoi Nomura, 1991, and B. fengtingae Yin and Li, 2014 [3–7]. Members of Batrisceniola
iations.
are characterized by the abdominal tergite 4 (VII) bearing a median bunch of erect setae
in both sexes. On the other hand, the genus Physomerinus Jeannel, 1952, defined by the
presence of sexual characters on male metafemora and a constricted basal capsule of the
Copyright: © 2022 by the authors.
aedeagus, includes 11 species distributed in China (3 spp.), Japan (including the Ryukyu
Licensee MDPI, Basel, Switzerland. Islands) (3 spp.), Indonesia (3 spp.), Thailand (2 spp.), Vietnam (1 sp.), Malaysia (1 sp.),
This article is an open access article Borneo (1 sp.), Singapore (1 sp.), New Guinea (1 sp.), and Myanmar (1 sp.) [3,6,8–17]. An
distributed under the terms and examination of the material collected from Nanling revealed one new species of each genus,
conditions of the Creative Commons which are formally described in the present paper.
Attribution (CC BY) license (https://
creativecommons.org/licenses/by/
4.0/).
Insects 2022, 13, 119. https://doi.org/10.3390/insects13020119 https://www.mdpi.com/journal/insects
Insects 2022, 13, 119 2 of 9
2. Materials and Methods
The material treated in this study is housed in the Insect Collection of Shanghai Normal
University (SNUC). The label data of the material are quoted verbatim.
Dissected parts were preserved in Euparal on plastic slides that were placed on the
same pin as the specimens. The habitus images were taken using a Canon 5D Mark III
camera (Ōita, Japan) in conjunction with a Canon MP-E 65 mm f/2.8 1-5X Macro Lens (Ōita,
Japan), and a Canon MT-24EX Macro Twin Lite Flash (Ōita, Japan) was used as the light
source. Images of morphological details were produced using a Canon G9 camera (Zhuhai,
China) mounted to an Olympus CX31 microscope (Ina-Shi, Japan) under transmitted light.
Zerene Stacker (version 1.04) was used for image stacking. All images were optimized and
grouped into plates using Adobe Photoshop CS5 Extended.
The abdominal tergites and sternites are numbered following Chandler (2001) [18] in
Arabic (starting from the first visible segment) and Roman (reflecting true morphological
position) numerals, e.g., tergite 1 (IV), or sternite 1 (III).
Measurements were taken as the following: the total body length was measured from
the anterior margin of the clypeus to apex of the abdomen; head length was measured
from the anterior margin of the clypeus to head base, excluding occipital constriction; head
width was measured across the eyes; the length of the pronotum was measured along the
midline, width equals its maximum width; the length of the elytra was measured along
the suture; the width is the maximum width across both elytra; the length of the abdomen
is the length of the dorsally exposed part of the abdomen along the midline; the width is
abdomen’s maximum width.
3. Results
Genus Batrisceniola Jeannel, 1958
Batrisceniola Jeannel, 1958: 65. Type species: Batrisus dissimilis, Sharp, 1874
(original designation).
Jeannel placed this genus in his fifth division of Batrisina, and defined it mainly
based on the male frons bearing a pair of large excavations [4], a criterion apparently of
little phylogenetic importance beyond species level. In a recent work, Nomura redefined
Batrisceniola on the basis of the large clypeus, and tergite 4 (VII) of the abdomen with a
median bunch of setae in both sexes [6]. Batrisceniola resembles Arthromelodes, Jeannel,
1954, in the similar configurations of the aedeagus and female genitalia, but can be readily
separated by the tergal setose brush. The first and the only Chinese congener, B. fengtingae,
Yin and Li, was described in 2014 [7], which possesses a simple head and a modified
tergite 1 (IV).
Batrisceniola nanlingensis Zhang & Yin, sp. nov.
Type material (20 exx.). Holotype: CHINA: ♂, ‘China: Guangdong, Shaoguan,
Ruyuan, Nanling N. R., Pubuqun (Falls Scenic Zone), 24◦ 540 9” N, 113◦ 20 53.8” E, 660–850 m,
3.V.2021, sifting, Hu, Lin, Zhou and Li leg.’ (SNUC). Paratypes: CHINA: 2 ♀♀, same
locality as holotype (SNUC); 2 ♂♂, same locality as holotype except ‘Xiaohuangshan,
24◦ 530 44.7” N, 113◦ 10 26.9” E, 1270–1570 m, 2.V.2021’, (SNUC); 1 ♂, 2 ♀♀, same locality
as holotype except ‘Qinshui Valley, 24◦ 550 42.9” N, 113◦ 00 59.05” E, 680–780 m, 5.V.2021’,
(SNUC); 10 ♂♂, same locality as holotype except ‘Guang-dong-di-yi-feng, 24◦ 550 29” N,
112◦ 590 31” E, 1538–1784 m, 28.VI.2020, Xia, Zhang, Yin, Lin leg’. (SNUC); 2 ♂♂, ‘China:
Guangdong Prov. Ruyuan County, Nanling N. R. Sta. alt. 1100 m 14, VIII, 2008, QI N and
YIN Z-W leg.’ (SNUC).
Diagnosis. Male. Body length is approximately 2.0 mm. The head is sub-rectangular,
with indistinct vertex sulcus connecting large vertexal foveae, with a thin mediobasal
carina extending from head base anteriorly and passing the level of the anterior margin
of the eyes; antennae elongate, lacking distinct club, antennomere 11 largest. Pronotum
with distinct median sulcus and pair of lateral longitudinal sulci, with broad transverse
antebasal impression. Discal striae of elytra shallow, extending posteriorly to apical 1/3
of elytral length, with small, angulate humeral prominence. Protibia with apical tuft of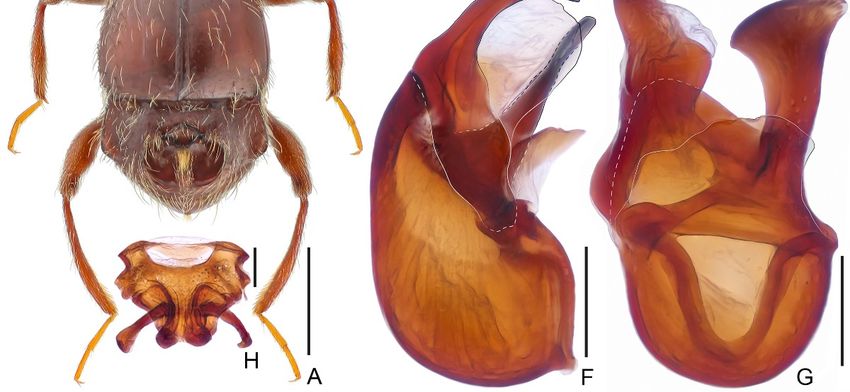
Insects 2022, 13, 119 3 of 9
setae at mesal margin; mesotibia with distinct apical spine. Tergite 4 (VII) with a large
central cavity and expanded laterally. Aedeagus strongly asymmetric, stout; ventral stalk
of median dilated at apex; dorsal lobe in lateral view narrowing toward apex, with broad
membranous ventral lamella; parameres fused to broad membrane. Female. Body length
approximately 2.0–2.1 mm; antennae slightly shorter; legs lacking apical tuft of setae or
spine; genital complex as in Figure 1H.
Description. Male. Body (Figure 1A) length 1.98–2.03 mm; color reddish-brown, tarsi
and mouthpart lighter.
Head (Figure 1B) sub-rectangular, slightly wider than long, length from anterior
margin of clypeus to head base 0.42–0.43 mm, width across eyes 0.48 mm; vertex shallowly
and roughly punctate at middle, punctation much coarser posterior antennal tubercles,
with large asetose vertexal foveae (dorsal tentorial pits), with indistinct U-shaped sulcus
connecting foveae, mediobasal carina thin, extending from head base anteriorly and passing
level of anterior margin of eyes, approaching transverse, arcuate carina between antennal
tubercles, lateral carina extending from base to posterior margin of antennal tubercle;
posterolateral margin round; lacking frontal-clypeal ridge, clypeus with carinate and
moderately raised anterior margin; ocular-mandibular carina complete, distinct. Venter
with rough surface; small gular foveae (posterior tentorial pits) in shared round opening,
with faint median carina extending from opening anteriorly to mouthpart. Compound
eyes prominent, each composed of approximately 30 ommatidia. Antenna elongate, length
1.01–1.03 mm, lacking distinct club; antennomere 1 thick, subcylindrical, with dense setae
on anterolateral margin, 2 elongate, 3–5 each slightly elongate, 6 smallest, 7 slightly wider
than 5, 8 moniliform, much smaller than 7, 9–11 each moderately enlarged, 11 largest,
approximately as long as antennomeres 9 and 10 combined.
Pronotum (Figure 1B) approximately as long as wide, length 0.45 mm, width
0.47–0.48 mm, widest at middle; sides rounded, convergent basally, disc slightly convex,
with shallow, rough punctures, median longitudinal sulcus with carinate sides, with pair of
lateral longitudinal sulci; broad antebasal sulcus connecting large, asetose lateral antebasal
foveae; with small outer and inner pair of basolateral foveae. Prosternum with anterior part
shorter than coxal part, with large lateral procoxal foveae; thin hypomeral ridge extending
from base to middle of anterior part; margins of coxal cavities obviously carinate.
Elytra wider than long, length 0.69–0.7 mm, width 0.81–0.86 mm; each elytron with
two moderately large, asetose basal foveae; thin discal stria extending posteriorly from
outer basal fovea to apical 1/3 of elytral length; with small, angulate humeral prominence,
subhumeral fovea present, carinate marginal stria extending from fovea to posterior margin
of elytron.
Mesoventrite short, demarcated from metaventrite by transverse carina; median
mesoventral foveae widely separated, in shared opening, large lateral mesoventral foveae
forked internally, marginal stria complete. Metaventrite slightly prominent, weakly im-
pressed at middle, with well-developed lateral mesocoxal and two lateral metaventral
foveae; posterior margin with small and narrow split at middle.
Legs elongate; protibia with apical tuft of setae, protrochanter with small ventral
denticle near apex, procoxa with long seta at ventral margin; mesotibia (Figure 1C) with
distinct apical spine.
Abdomen widest at mesal margins of tergite 1 (IV), length of abdomen 0.64–0.65 mm,
width 0.79–0.82 mm. Tergite 1 (IV) (Figure 1D) approximately three times as long as
2–4 (V–VII) combined, expanded on both sides, widest near middle; discal carina short but
distinct, thin basal sulcus separated by mediobasal and large basolateral foveae, lacking
inner marginal carina, outer one complete, with large, deep oval cavity at middle of tergite,
anterior margin of cavity ridged, posterior margin with large, setiferous oval nodule; tergite
2 (V) as long as 3 (VI), 4 (VII) longer than tergites 2 and 3 combined, posterior half of tergite
4 (VII) with bunch of erect setae at middle; tergite 5 (VIII) semicircular, transverse, posterior
margin roundly emarginate at middle. Sternite 2 (IV) with one pair of mediobasal and one
pair of basolateral foveae, with short lateral carina; midlength of sternites 3–5 (V–VII) short,Insects 2022, 13, x FOR PEER REVIEW 4 of 9
Insects 2022, 13, 119 4 of 9
transverse, posterior margin roundly emarginate at middle. Sternite 2 (IV) with one pair
of mediobasal
each and one
with one pair pair basolateral
of small of basolateral foveae,
foveae; with short
sternite lateral
6 (VIII) carina; midlength
transverse, of
posterior margin
sternites emarginate;
broadly 3–5 (V–VII) short, each
sternite with(Figure
7 (IX) one pair of semi-membranous,
1E) small basolateral foveae; sternite
apical 6 (VIII)
portion rounded
transverse,
and posteriorsclerotized.
more strongly margin broadly emarginate; sternite 7 (IX) (Figure 1E) semi-mem-
branous, apical (Figure
Aedeagus portion 1F,G)
rounded 0.37and
mmmore
long,strongly
stronglysclerotized.
asymmetric; median lobe with rounded
basalAedeagus
capsule and (Figure 1F,G)
roundly 0.37 mmbasal
triangular long,foramen,
stronglyventral
asymmetric; median lobe
stalk elongate, with at
broadened
rounded
apex; basal
dorsal capsule
lobe and dorsoventrally
elongate, roundly triangular
widest basal foramen,
at apical third, ventral stalk elongate,
then narrowing apically,
broadened at apex; dorsal lobe elongate, dorsoventrally widest at apical third,
strongly curved to ventral side at apex, with broad membranous ventral lamella; parameres then nar-
rowingtoapically,
fused strongly
single, broad curved to ventral
membranous side at apex, with broad membranous ventral
structure.
lamella; parameres fused to single, broad membranous
Female. Similar to male in external morphology; antenna structure.slightly shorter; protibia lack-
Female. Similar to male in external morphology; antenna
ing long apical tuft of setae, protrochanter lacking ventral denticle, slightly shorter;lacking
mesotibia protibia
apical
spine; each compound eye composed of approximately 25 ommatidia; tergite lacking
lacking long apical tuft of setae, protrochanter lacking ventral denticle, mesotibia 1 (IV) with-
apical spine; each compound eye composed of approximately 25 ommatidia; tergite 1 (IV)
out median cavity. Measurements (as for male): body length 2.03–2.06 mm; length/width of
without median cavity. Measurements (as for male): body length 2.03–2.06 mm;
head 0.43–0.44/0.49–0.50 mm, pronotum 0.45–0.47/0.48 mm, elytra 0.62–0.63/0.80–0.81 mm;
length/width of head 0.43–0.44/0.49–0.50 mm, pronotum 0.45–0.47/0.48 mm, elytra 0.62–
abdomen 0.64–0.68/0.75–0.78 mm; length of antenna 0.95–0.97 mm; maximum width of
0.63/0.80–0.81 mm; abdomen 0.64–0.68/0.75–0.78 mm; length of antenna 0.95–0.97 mm;
genitalia (Figure 1H) 0.35 mm.
maximum width of genitalia (Figure 1H) 0.35 mm.
Figure 1.
Figure 1. Batrisceniola
Batrisceniolananlingensis
nanlingensissp.
sp.nov.
nov.((A–G). Male.
((A–G). (H).(H).
Male. Female). (A) (A)
Female). Dorsal habitus;
Dorsal (B) Head
habitus; (B) Head
and pronotum; (C) mesotibia; (D) Tergite 1–4 (IV–VII); (E) sternite 7 (IX); (F,G) Aedeagus, in lateral
and pronotum; (C) mesotibia; (D) Tergite 1–4 (IV–VII); (E) sternite 7 (IX); (F,G) Aedeagus, in lateral
(F) and ventral (G) view; (H) Female genitalia. Scale bars: 0.5 mm in (A); 0.3 mm in (B); 0.2 mm in
(F) and ventral (G) view; (H) Female genitalia. Scale bars: 0.5 mm in (A); 0.3 mm in (B); 0.2 mm in
(C,D); 0.1 mm in (E–H).
(C,D); 0.1 mm in (E–H).
Comparative notes. The new species morphologically resembles the Japanese Batris-
Comparative
ceniola semipunctata notes. The1909)
(Raffray, new inspecies morphologically
the presence of a largeresembles
cavity on the 1 (IV),Batrisce-
Japanese
tergite as
niola semipunctata (Raffray, 1909) in the presence of a large cavity on tergite 1 (IV),
well as the shape of the aedeagus. They can be separated by the different structure of asthe
well as
the shape of the aedeagus. They can be separated by the different structure of the cavity,
the lack of lateral circular setiferous patches of tergite 1 (IV), and the dorsal lobe of the
aedeagus lacks a subapical protuberance in B. nanlingensis sp. nov.Insects 2022, 13, 119 5 of 9
Distribution. China: Guangdong.
Etymology. The new species is named after its type locality, i.e., Nanling National
Forest Park.
Key to species of Batrisceniola from China (male)
1 Tergite 1 (IV) with small cavity located near posterior margin [Reference [7]: Figure 1F],
lateral margins rounded; aedeagus with extended ventral stalk apically narrowing
and greatly bent [Reference [7]: Figure 1K] . . . . . . . . . . . . B. fengtingae Yin and Li 2014
- Tergite 1 (IV) with large cavity located near middle (Figure 1A,D), lateral margins
expanded; aedeagus with short ventral stalk apically greatly dilated (Figure 1G) . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . B. nanlingensis sp. nov.
Genus Physomerinus Jeannel, 1952
Physomerinus Jeannel, 1952b: 96. Type species: Batrisus septemfoveolatus Schaufuss, 1877
(original designation).
Similar to many Asian genera created by Jeannel which are often characterized by
male sexual characters, the genus Physomerinus is defined based on the sexually modified
hind femora of the male, as well as by the aedeagus with a constricted basal capsule [6].
The genus Btriscenaulax Jeannel, with five species from Japan, also has the aedeagus with a
small basal capsule, and under current definition is separated from Physomerinus by the
modified protibiae and tergite 1 (IV) in the male. The relationship between these two genera
is evidently close, and needs to be further investigated.
Physomerinus clavipes Zhang & Yin, sp. nov.
Type material (2 exx.). Holotype: CHINA: ♂, ‘China: Guangxi, Guilin, Huaping N.
R., 25◦ 370 39.83” N, 109◦ 540 20.23” E, 780 m, 18.VIII.2020, streamside, Qiu Lu leg’. (SNUC).
Paratype: CHINA: 1 ♀, same locality as holotype (SNUC).
Diagnosis. Male. Body length approximately 1.8 mm. Head sub-rectangular, with
distinct vertex sulcus connecting large, asetose vertexal foveae, with thin mediobasal carina
extending from head base to anteriorly beyond level of posterior margin of eyes, antennae
elongate, lacking distinct club, antennomere 11 largest. Pronotum with median sulcus
and pair of lateral longitudinal sulci, with broad antebasal sulcus. Discal striae extending
posteriorly from outer basal fovea to apical 1/5 of elytral length with small, angulate
humeral prominence. Protibia with apical tuft of setae; lateral margin of metafemur strongly
swollen. Aedeagus strongly asymmetric; median lobe stout, ventral stalk dorsoventrally
broadened at middle and apex; dorsal lobe in lateral view broad; parameres fused to broad
membrane. Female. Body length approximately 1.9 mm; antennae slightly shorter; legs
lacking apical tuft of setae or modification; genital complex as in Figure 2G.
Description. Male. Body (Figure 2A) length 1.78 mm; color yellowish-brown.
Head (Figure 2B) sub-rectangular, wider than long, length from anterior margin of
clypeus to head base 0.34 mm, width across eyes 0.41 mm; vertex sparsely and finely
punctate, with distinct U-shaped sulcus connecting large, asetose foveae (dorsal tentorial
pits), mediobasal carina short and thin, extending from head base anteriorly beyond level
of posterior margin of eyes, lateral carina extending from base to posterior margin of
antennal tubercle; posterolateral margin round; lacking frontal-clypeal ridge, clypeus
with carinate and moderately raised anterior margin; ocular-mandibular carina complete,
distinct. Venter with small gular foveae (posterior tentorial pits) in shared round opening,
with shallow median carina extending from opening anteriorly to mouthpart. Compound
eyes prominent, each composed of approximately 35 ommatidia. Antenna elongate, length
0.97 mm, lacking distinct club; antennomere 1 thick, subcylindrical, 2–7 elongate, 7 slightly
wider than 6, 8 smallest, 9–11 each moderately enlarged, 11 largest, slightly shorter than 9
and 10 combined.Insects 2022, 13, 119 6 of 9
Insects 2022, 13, x FOR PEER REVIEW 6 of 9
Figure 2.
Figure 2. Physomerinus
Physomerinusclavipes sp.sp.
clavipes nov. ((A–F).
nov. Male.
((A–F). (G). Female).
Male. (A) Dorsal
(G). Female). habitus;habitus;
(A) Dorsal (B) Head(B)
and
Head
pronotum; (C) metafemur; (D) sternite 7 (IX); (E,F) Aedeagus, in lateral (E) and ventral (F) view; (G)
and pronotum; (C) metafemur; (D) sternite 7 (IX); (E,F) Aedeagus, in lateral (E) and ventral (F) view;
Female genitalia. Scale bars: 0.5 mm in (A); 0.2 mm in (B,C); 0.1 mm in (G); 0.05 mm in (E,F); 0.03
(G)
mmFemale
in (D). genitalia. Scale bars: 0.5 mm in (A); 0.2 mm in (B,C); 0.1 mm in (G); 0.05 mm in (E,F);
0.03 mm in (D).
Pronotum (Figure 2B) approximately as long as wide, length 0.42 mm, width 0.41
mm,Pronotum (Figure 2B)
widest at middle; approximately
sides as long as
rounded, convergent wide, disc
basally, length 0.42 mm,
slightly width
convex, 0.41 mm,
sparsely
widest
with minute punctures, median longitudinal sulcus from anterior l/3 to posterior l/8, withwith
at middle; sides rounded, convergent basally, disc slightly convex, sparsely
minute punctures,
pair of lateral mediansulci;
longitudinal longitudinal sulcus sulcus
broad antebasal from anterior
connecting l/3large,
to posterior l/8, with
asetose lateral
pair of lateral longitudinal sulci; broad antebasal sulcus connecting
antebasal foveae; with small outer and inner pair of basolateral foveae. Prosternum with large, asetose lateral
antebasal
anterior part shorter than coxal part, with large lateral procoxal foveae; thin hypomeral an-
foveae; with small outer and inner pair of basolateral foveae. Prosternum with
terior
ridge part shorter
extending thanbase
from coxal
to part,
middle with large lateral
of anterior part;procoxal
margins of foveae;
coxal thin hypomeral
cavities obviously ridge
extending
carinate. from base to middle of anterior part; margins of coxal cavities obviously carinate.
Elytra widerthan
Elytra wider thanlong,
long, length
length 0.610.61
mm,mm,widthwidth
0.69 mm;0.69eachmm;elytron
each with
elytron
twowith
mod-two
moderately large, asetose basal foveae; thin discal stria extending posteriorly
erately large, asetose basal foveae; thin discal stria extending posteriorly from outer basal from outer
basal
fovea fovea to apical
to apical 1/5 of1/5 of elytral
elytral length;length; with angulate
with small, small, angulate
humeralhumeral
prominence,prominence,
sub-
subhumeral
humeral fovea fovea present,
present, carinate
carinate marginal
marginal striastria extending
extending from fromfoveafovea to posterior
to posterior margin
margin
of
ofelytron.
elytron.
Mesoventrite short,
Mesoventrite short, demarcated
demarcatedfrom frommetaventrite
metaventrite byby transverse
transverse carina; median
carina; median
mesoventral foveae widely
mesoventral widelyseparated,
separated,ininshared
shared opening,
opening, lateral
lateralmesoventral
mesoventral foveae large,
foveae large,
without internal
without internal fork,
fork, marginal
marginalstria
striacomplete.
complete.Metaventrite
Metaventrite slightly
slightlyprominent,
prominent,weakly
weakly
impressed at
impressed at middle,
middle, with
withwell-developed
well-developedlateral
lateral mesocoxal
mesocoxal and andtwotwolateral metaventral
lateral metaventral
foveae; posterior margin with small and narrow split
foveae; posterior margin with small and narrow split at middle. at middle.
Legs elongated; protibia with long apical tuft of setae; metafemur (Figure 2C) thick,
strongly dilated at apical 2/5, dorsal surface of dilation with narrow cavity, with lamellar
projection at bottom of cavity.Insects 2022, 13, 119 7 of 9
Abdomen widest at anterior margins of tergite 1 (IV), length of abdomen 0.46 mm,
width 0.58 mm. Tergite 1 (IV) much longer than 2–4 (V–VII) combined, discal carina shot
but distinct, thin basal sulcus separated by mediobasal and large basolateral foveae, lacking
inner marginal carina, outer one thin, short, extending posteriorly from anterior margin of
tergite 1 (IV) to basal 2/3 of tergite length; tergites 2–3 (V–VI) very short, tergite 4 (VII) twice
as long as 2–3 (V–VI) combined in posterior view; tergite 5 (VIII) ovoid, convex, posterior
margin roundly emarginate at middle. Sternite 2 (IV) with 2 pairs of mediobasal and one
pair of basolateral foveae, with short lateral carina; midlength of sternites 3–5 (V–VII) short,
lacking basolateral foveae; sternite 6 (VIII) transverse, posterior margin greatly emarginate;
sternite 7 (IX) (Figure 2D) semi-membranous, apical portion narrowed and more strongly
sclerotized, with small split at right.
Aedeagus (Figure 2E,F) 0.19 mm long, strongly asymmetric; median lobe with trans-
versely extended basal capsule, broad foramen, and distinct basoventral projection; ventral
stalk elongate, broadened at middle and apex; dorsal lobe in lateral view broad through
entire length; parameres fused to single, broad membranous structure.
Female. Similar to male in external morphology; antenna slightly shorter; protibia
lacking long apical tuft of setae, metafemur lacking large, swollen modification at apical
2/5; each compound eye composed of approximately 30 ommatidia. Measurements (as for
male): body length 1.86 mm; length/width of head 0.37/0.42 mm, pronotum 0.43/0.42 mm,
elytra 0.57/0.69 mm; abdomen 0.59/0.62 mm; length of antenna 0.99 mm; maximum width
of genitalia (Figure 2G) 0.29 mm.
Comparative notes. The new species morphologically resembles P. hasegawai Nomura,
1991 in the similar form of the femoral modification, but can be readily separated by the
different configuration of the aedeagus.
Distribution. China: Guangxi.
Etymology. The specific epithet refers to the strongly clavate metafemoral modification
of the new species.
Key to species of Physomerinus from China
1 Median longitudinal sulcus relatively much longer, approaching both anterior and
posterior margin of pronotum . . . . . . . . . . . . . . . . . . . . . P. schenklingi (Raffray, 1912)
- Median longitudinal sulcus relatively much shorter, never approaching anterior
margin of pronotum . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2 Metafemur with ventral oblique furrow . . . . . . . . . . . . . . . P. cruralis (Raffray, 1914)
- Metafemur with cavity and projection/expansion . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
3 Metafemur [Reference [6]: Figure 133A,D] hardly clavate, with deep excavation on
dorsal surface and mushroom-shaped projection at bottom of excavation . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . P. pedator (Sharp, 1883)
- Metafemur (Figure 2C) greatly clavate, with excavation on dorsal surface of expansion
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . P. clavipes sp. nov.
4. Discussion
The Nanling Mountain Range was identified as one of China’s biodiversity hotspots
in terms of endemic plants and mammals [19]. Two studies on the local species richness
of Argidae (Hymenoptera) and butterflies (Lepidoptera) indicated the insect fauna is
predominated by Oriental elements [20,21]. Regardless, summarized knowledge regarding
species diversity of most insect groups, including the subject of the present paper, the
Pselaphinae, is largely unavailable. Prior to the present study, merely 16 species of Batrisini
inhabiting several types of micro-habitats have been known to occur in the vast Nanling
Mountain Area: two cavernicolous species each of Araneibatrus Yin and Li, 2010, and
Tribasodites Jeannel, 1960, one myrmecophilous species each of Dendrolasiophilus Nomura,
2010 and Songius Yin & Li, 2010, and the others live freely in association with leaf litter
layer or decomposing logs [22–26]. Aside from the two new species described here, we
have recognized at least 100 additional species belonging to various groups of Batrisini
from our sample, most of which was collected in the past three years and will be dealt withInsects 2022, 13, 119 8 of 9
elsewhere. The true diversity of Pselaphinae from Nanling is difficult to evaluate at this
moment due to the lack of taxonomic and distribution data, but the need in accelerating
species description is evident when considering the ecosystems of the Nanling Mountain
Range are fragile and sensitive to environmental change [27,28].
Author Contributions: Conceptualization, Z.-W.Y.; resources, W.-X.Z. and Z.-W.Y.; writing—original
draft preparation, W.-X.Z.; writing—review and editing, Z.-W.Y.; visualization, W.-X.Z.; project
administration, Z.-W.Y. All authors have read and agreed to the published version of the manuscript.
Funding: This work was funded by the National Natural Science Foundation of China (31872965),
the Shanghai Municipal Science and Technology Commission, China (19QA1406600), and
GDAS Special Project of Science and Technology Development (2020GDASYL–20200102021,
2020GDASYL–20200301003).
Institutional Review Board Statement: Not applicable.
Informed Consent Statement: Not applicable.
Data Availability Statement: Data sharing is not applicable to this article.
Acknowledgments: We are grateful to Xing-Ke Yang (Guangdong Entomological Institute,
Guangzhou, China), who leads the Nanling project, for his encouragement and support to our
work. Lu Qiu (Mianyang Normal University, Mianyang, Sichuan, China) collected and kindly do-
nated the material of P. clavipes to the SNUC collection. Jia-Yao Hu (Shanghai Normal University,
Shanghai, China) led the 2021 trip to Nanling, and Xing-Min Wang (South China Agricultural Univer-
sity, Guangdong, China) offered various kinds of help in the field. Four anonymous reviewers read
the draft manuscript and provided helpful comments which improved the paper.
Conflicts of Interest: The authors declare no conflict of interest.
References
1. Ministry of Ecology and Environment of the People’s Republic of China (MEE). China National Biodiversity Conservation Strategy
and Action Plan (2011–2030). 2015. Available online: https://www.cbd.int/doc/world/cn/cn-nbsap-v2-en.pdf (accessed on 20
November 2021).
2. Wang, Y.-Y.; Dong, Y.-X. Geographical detection of regional demarcation in the Nanling Mountains. Trop. Geogr. 2018, 38, 337–346.
[CrossRef]
3. Newton, A.F. StaphBase: Staphyliniformia World Catalog Database (Version Nov. 2018). In Species 2000 & ITIS Catalogue of
Life, 2020-12-01; Roskov, Y., Ower, G., Orrell, T., Nicolson, D., Bailly, N., Kirk, P.M., Bourgoin, T., DeWalt, R.E., Decock, W.,
van Nieukerken, E.J., et al., Eds.; Naturalis Biodiversity Center: Leiden, The Netherlands; ISSN 2405-8858. Available online:
www.catalogueoflife.org (accessed on 10 December 2021).
4. Jeannel, R. Révision des Psélaphides du Japon. Mém. Mus. Natl. d’Histoire Nat. 1958, 18, 1–138.
5. Raffray, A. Nouvelles espèces de Psélaphides. Ann. Soc. Entomol. Fr. 1909, 78, 15–52.
6. Nomura, S. Systematic study on the genus Batrisoplisus and its allied genera from Japan (Coleoptera, Pselaphidae). Esakia 1991,
30, 1–462. [CrossRef]
7. Yin, Z.-W.; Li, L.-Z. Batrisceniola fengtingae sp. nov., the first record of the genus in China (Coleoptera: Staphylinidae: Pselaphinae).
Acta Ent. Mus. Nat. Pra. 2014, 54, 233–236.
8. Reitter, E. Beitrag zur Pselaphiden- und Scydmaeniden-Fauna von Java und Borneo. Verh. K. K. Zool.-Bot. Ges. Wien 1883, 32,
283–302.
9. Löbl, I. Beitrag zur Kenntnis der Pselaphidae (Coleoptera) der Koreanischen Volksdemokratischen Republik, Japans und des
Ussuri Gebietes. Ann. Zool. 1973, 30, 319–334.
10. Raffray, A. Nouvelles études sur les Psélaphides et les Clavigérides. Ann. Soc. Entomol. Fr. 1897, 65, 227–284.
11. Raffray, A. Genera et catalogue des Psélaphides. Ann. Soc. Entomol. Fr. 1904, 73, 1–400.
12. Raffray, A. Sauter’s Formosa-Ausbeute. Pselaphidae (Col.) II. Suppl. Entomol. 1914, 3, 1–5.
13. Motschulsky, V. Énumération des nouvelles espèces de Coléoptères. Bull. Soc. Imp. Nat. Moscou 1851, 24, 479–511.
14. Schaufuss, L.W. Pselaphiden Siam’s; Ferdinand Thomass: Dresden, Germany, 1877; pp. 1–25.
15. Schaufuss, L.W. Pselaphiden und Scydmaeniden der Niederländischen Besitzungen auf den Sunda-Inseln, in Reichsmuseum zu
Leyden. Tijdschr. Entomol. 1882, 25, 65–76.
16. Schaufuss, L.W. Neue Pselaphiden im Museo Civico di Storia Naturale zu Genua. Ann. Mus. Civ. Stor. Nat. Genova 1882, 18,
349–399.
17. Jeannel, R. Psélaphides de Saigon. Rev. Fr. Entomol. 1952, 19, 69–113.Insects 2022, 13, 119 9 of 9
18. Chandler, D.S. Biology, morphology, and systematics of the antlike litter beetles of Australia (Coleoptera: Staphylinidae:
Pselaphinae). Mem. Entomol. Int. 2001, 15, 1–560.
19. Tang, Z.-Y.; Wang, Z.-H.; Zheng, C.-Y.; Fang, J.-Y. Biodiversity in China’s mountains. Front. Ecol. Environ. 2006, 4, 347–352.
[CrossRef]
20. You, Q. Species and fauna of Argidae (Hymenoptera) in Nanling Mountains, China. Acta Pratacult. Sin. 2009, 18, 130–135, (In
Chinese with English abstract).
21. Zhou, G.-Y.; Gu, M.-B.; Gong, Y.-N.; Wang, S.-K.; Wu, Z.-M.; Xie, G.-G. Diversity and fauna of butterflies in Nanling National
Nature Reserve. J. Environ. Entomol. 2016, 38, 971–978.
22. Yin, Z.-W.; Jiang, R.-X.; Steiner, H. Revision of the genus Araneibatrus (Coleoptera: Staphylinidae: Pselaphinae). Zootaxa 2016,
4097, 475–494. [CrossRef]
23. Yin, Z.-W.; Li, L.-Z.; Zhao, M.-J. Discovery in the caves of Guangxi, China: Three new troglobitic species of Tribasodites Jeannel
(Coleoptera, Staphylinidae, Pselaphinae). Zootaxa 2011, 3065, 49–59. [CrossRef]
24. Yin, Z.-W.; Nomura, S.; Li, L.-Z. Ten new species of cavernicolous Tribasodites from China and Thailand, and a list of East Asian
cave-inhabiting Pselaphinae (Coleoptera: Staphylinidae). Acta Ent. Mus. Nat. Pra. 2015, 55, 105–128.
25. Yin, Z.-W.; Li, L.-Z. Dendrolasiophilus Wenhsini (Coleoptera: Staphylinidae: Pselaphinae), A New Myrmecophilous Batrisine
Associated with Lasius (Dendrolasius) spathepus from Guangxi, South China. In Memory of Mr. Wenhsin Lin; Lin, M.-Y., Chen, C.-C.,
Eds.; Formosa Ecological Company: Kaohsiung City, China (Taiwan), 2013; pp. 183–187.
26. Yin, Z.-W.; Li, L.-Z. New and little known species of the genus Songius (Coleoptera: Staphylinidae: Pselaphinae) in China. Zootaxa
2015, 3905, 293–300. [CrossRef] [PubMed]
27. Stone, R. Ecologists report huge storm losses in China’s forests. Science 2008, 319, 1318–1319. [CrossRef] [PubMed]
28. Li, C.-H.; Yu, S.-Y.; Yao, S.-C.; Shen, J.; Ji, M.; Chen, R.; Sun, W.-W.; Ling, C.-H. Response of alpine vegetation to climate changes in
the Nanling Mountains during the second half of the Holocene. Quat. Int. 2019, 522, 12–22. [CrossRef]You can also read

















































