Tidal Wetlands after Hurricane Sandy: Baseline restoration assessment and future conservation planning
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Tidal Wetlands after Hurricane Sandy:
Baseline restoration assessment and future
conservation planning
Final Report, February 2018
Submitted By:
Co-PIs
Chris S. Elphick1, Brian J. Olsen2, W. Gregory Shriver3 and Jonathan Cohen4
Post-doctoral researchers and staff
Maureen D. Correll2, Wouter Hantson2, Brian Klingbeil1, and Elizabeth Tymkiw3
1. Department of Ecology & Evolutionary Biology and Center for Conservation and Biodiversity, University of
Connecticut, 75 North Eagleville Road, U-43, Storrs, Connecticut 06269
2. School of Biology and Ecology, Climate Change Institute, University of Maine, 200 Clapp Greenhouse, Orono,
Maine 04469
3. Department of Entomology and Wildlife Ecology, University of Delaware, 250 Townsend Hall, Newark,
Delaware 19716
4. Department of Environmental and Forest Biology, SUNY College of Environmental Science and Forestry, 1
Forestry Drive, Syracuse, NY 13210
Acknowledgments
This work was primarily funded by a grant from the US Department of the Interior, Fish and Wildlife
Service and North Atlantic Landscape Conservation Cooperative through the Disaster Relief
Appropriations Act of 2013 (award # F14AC00965) and relied heavily on data collected using a
Competitive State Wildlife Grant (U2-5-R-1) via the US Fish and Wildlife Service, Federal Aid in Sportfish
and Wildlife Restoration award to the states of Delaware, Maryland, Connecticut, and Maine. We
received additional funding that supported our work on tidal marsh bird responses to Hurricane Sandy
from the US Fish and Wildlife Service (Cooperative Agreement Award Fl5AC00163) and the National
Science Foundation (DEB-1340008). Additional support for portions of the work come from the United
States Department of Agriculture (National Institute of Food and Agriculture, Project Number ME0-
21710). The findings and conclusions in this report are those of the authors and do not necessarily
represent the views of the funding agencies.
All work conducted by the Saltmarsh Habitat and Avian Research Program (SHARP) is a collaborative
venture and we thank our co-PIs Adrienne Kovach and Tom Hodgman; other SHARP postdocs Britt Cline,
Chris Field, and Kate Ruskin; and graduate students, especially Sam Apgar, Bri Benvenuti, Logan Cline,
Meaghan Conway, Tim Freiday, Laura Garey, Alison Kocek, and Sam Roberts. We also thank the many
Saltmarsh Habitat and Avian Research Program (SHARP) field technicians who collected data for this
effort, and all participating landowners and land managers that allowed access to their properties or
logistic support in the field. In particular, we thank the many points-of-contact for coastal resiliency
project sites at which we worked: R. Albers, K. Bennet, J. Burger, P. Castelli, C. Chaffee, P. Comins, D.
Curson, A. Drohan, L. Duff, N. Ernst, B. Finn, C. Folsom-O’Keefe, B. Gaspar, D. Golden, S. Guiteras, H.
Hanlon, J.M. Hartman, K. Holcomb, E. Jordan, M. Katkowski, M. Larson, J. Lister, J. Luk, G. Macdoland, J.
Mattei, L. Mitchell, L. Niles, J. Ong, N. Pau, J. Quispe, C. Salazar, J. Smith, M. Whitbeck, J. White, M.
Williams, and R. Wolfe. We also thank Janet Leese for countless hours spent digitizing training polygons
in the lab and Erika Lentz for reclassifying coastal response raster layers to reflect IPCC ranges.
Comments from E. Legaard, K. Legaard, D. Rosco, N. Hanson, members of the PIs respective lab groups,
and the larger SHARP team have substantially improved the work described here.
ii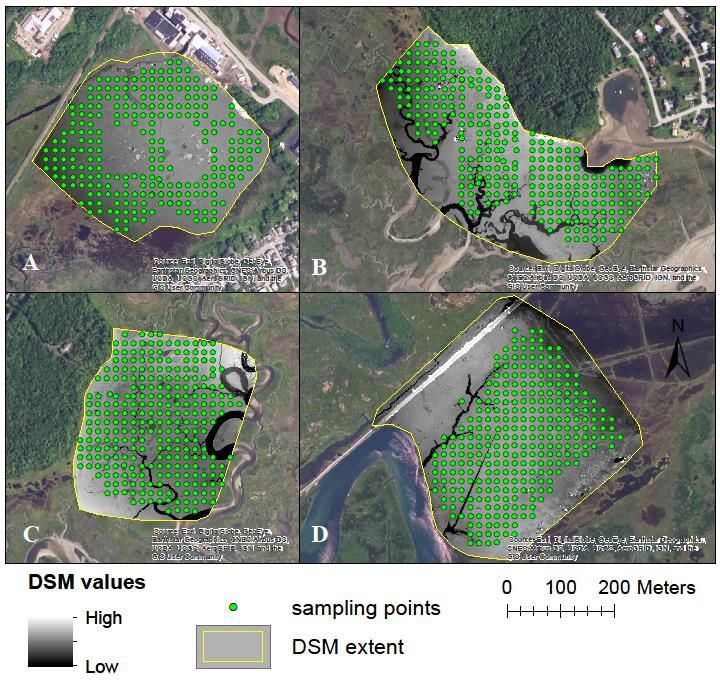
Contents
I. Executive Summary …………………………………………………………………………………………………………………. iv
II. Manuscript 1: Measuring elevation in northeastern USA tidal marshes …………………………………… 1
III. Manuscript 2: Fine-scale mapping of coastal plant communities in the northeastern USA ……… 14
IV. Manuscript 3: Using systematic conservation planning to identify a focal species for tidal marsh
bird conservation in the northeastern United States ………………………………………………………………. 35
V. Manuscript 4: Characterizing uncertainty to inform tidal marsh conservation in response to sea-
level rise ………….……………………………………………………………………………………………………………………… 60
VI. Hurricane Sandy coastal resiliency program: restoration project summaries ……...….……………… 69
Thin-layer sediment additions and vegetation planting at Sachuest Point, RI.
iii
I. Executive summary
Hurricane Sandy has fundamentally altered the way in which society thinks about coastal resilience in
the mid-Atlantic and New England states. Considerable funds are being invested in restoring coastal
resources and the need to systematically plan how we invest is increasingly apparent. A thorough
understanding of the effectiveness of Hurricane Sandy restoration projects on tidal marshes and the
wildlife they support is thus needed to direct conservation actions where they will have the greatest
benefits toward increasing the resilience of green infrastructure.
Many species of conservation concern use tidal marshes and face additional threats from sea-level rise.
The Saltmarsh Habitat and Avian Research Program (SHARP) has assembled a large collection of
historical and contemporary measures of plant and bird communities in tidal marshes from Virginia to
Maine. In the current project, we used this platform to begin to assess the efficacy of restoration
activities and to provide planning guidance to enhance the future resiliency of natural coastal assets.
Our primary focus was to collected elevation, vegetation, and bird data to quantify baseline conditions
at National Fish and Wildlife Foundation (NFWF) funded Hurricane Sandy resiliency sites throughout the
Northeast and mid-Atlantic states and at paired control sites. These data have been combined with
similar data collected at Department of the Interior-funded resiliency sites across the same region into a
common database, and will form the basis for long-term investigations into the efficacy of coastal
restoration and management work.
In addition to baseline data collection at resiliency sites, our work has had three specific research goals.
First, we investigated whether elevation data collected in the field via real-time kinetic (RTK) GPS
methods can be accurately replicated using remote sensing data products. Collection of RTK elevation
data is both time consuming and expensive, and thus difficult to obtain for large areas with high spatial
resolution. Remote sensing data, by contrast, are available across vast areas and are often publically
available for free. We tested a variety of remote sensing data at both local and regional scales and found
that the 1/9 arc-second (~1 m resolution) data layer from the National Elevation Dataset (NED)
produced high quality predictions of the RTK measurements. These predictions were especially good for
high elevation marsh, a habitat of particular interest to land managers. Other resolution NED data, and
data collected used an unmanned aircraft system, were far less satisfactory and were poor substitutes
for field-collected data in our tests.
Second, we compared several remote sensing techniques to develop a tool that accurately maps high-
and low-marsh zonation for use in management and conservation planning. We found that random
forests outperformed other classifier tools when applied to the most recent National Agricultural
Imagery Program (NAIP) imagery, NAIP derivatives, and elevation data between coastal Maine and
Virginia, USA. We then used these methods to classify plant zonation within a 500-m buffer around
coastal marsh delineated in the National Wetland Inventory. We found mean classification accuracies of
94% for high elevation marsh, 76% for low marsh zones, and 90% overall map accuracy. The detailed
output is a 3-m resolution continuous map of tidal marsh vegetation communities and cover classes that
can be used in habitat modeling of marsh-obligate species or to monitor changes in marsh plant
communities over time.
Third, we used the systematic conservation planning software Marxan to identify priority areas for the
conservation of five species that rely on tidal marsh for breeding: clapper rail Rallus crepitans, eastern
iv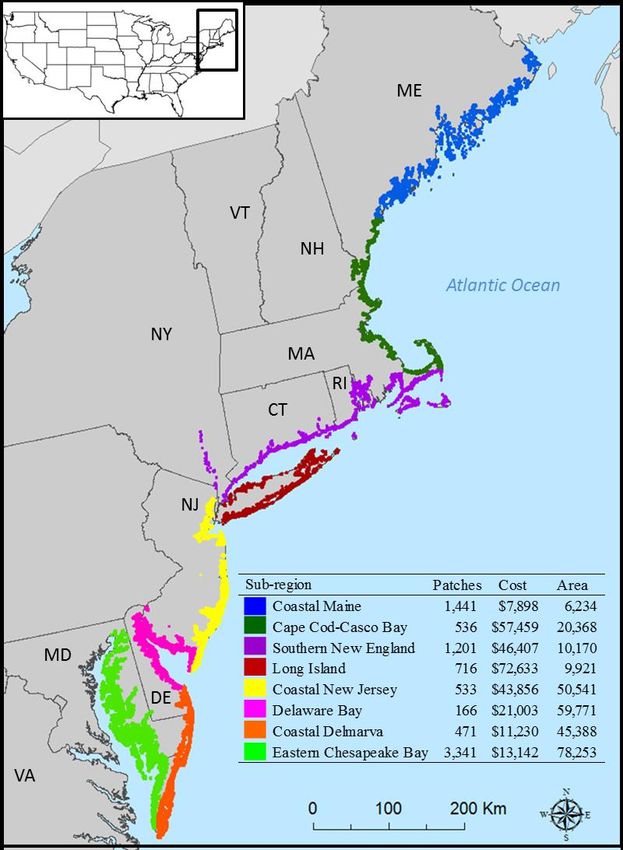
willet Tringa semipalmata semipalmata, Acadian Nelson’s sparrow Ammodramus nelsonii subvirgatus,
saltmarsh sparrow Ammodramus caudacutus, and seaside sparrow Ammodramus maritimus. We
compared the spatial prioritization of sites and cost-effectiveness of alternative protection scenarios
that considered individual species, groups of species, and all species simultaneously. Scenarios that
prioritized areas for conservation based on different single species targets were poorly correlated with
each other. Scenarios based on saltmarsh sparrow conservation were most strongly related (rs = 0.76) to
site prioritizations that consider all five tidal marsh specialists simultaneously. These results suggest that
although no species is a good surrogate for another single species, saltmarsh sparrow may be a viable
focal species for conservation planning to protect tidal marsh birds as a group. When comparing multi-
species combinations to prioritizations based on saltmarsh sparrow alone, the estimated costs, area of
land protection, and number of individuals of each species on protected land were similar, suggesting
that it is the best option considered. We were also able to identify areas where conservation is likely to
have little or no effect for the target species. Eliminating these sites from consideration for the
conservation of these species would reduce the risk of misdirecting limited conservation funds.
Because sea-level rise is likely to alter where suitable habitat lies, long-term conservation planning
should account for future conditions. With that in mind, we assessed the likelihood that each marsh
patch considered in our initial conservation planning exercise would persist by 2050. To accommodate
uncertainty in future conditions, we used both results from sea-level projections that assume static
inundation and those that assume dynamic adaptation by marshes in response to sea-level under
alternative carbon emissions scenarios. The results suggest that, at most, ~15% of current tidal marsh is
secure (>66% chance of persisting), but that much of the remainder has potential to persist via dynamic
adaptation (e.g., marsh building). Substantial declines of all tidal marsh specialist breeding birds are
predicted, commensurate with the predicted losses. In light of these projections, we identified marsh
patches with the greatest chance of persisting in the future and recommend a conservation planning
strategy that accounts for both current conservation value and the probability of long-term habitat
persistence.
This report consists of four manuscripts, written for submission to the peer review literature, and
dealing with (1) the use of remote sensing to estimate marsh elevation, (2) description of marsh
vegetation throughout the northeastern USA, (3) current conservation planning for focal marsh birds,
and (4) future projections for marsh area and focal marsh birds. We also include baseline data
summaries for seventeen coastal resilience projects distributed throughout the northeastern USA.
v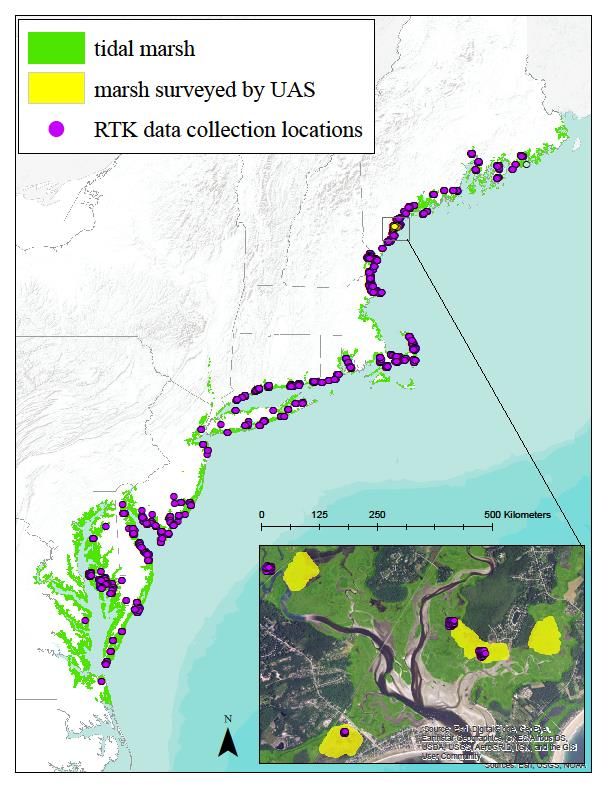
II. Measuring elevation in northeastern USA tidal marshes
Maureen D. Correll, Chris S. Elphick, Wouter Hantson, Brittany B. Cline, Elizabeth L. Tymkiw, W. Gregory
Shriver, and Brian J. Olsen
Sea levels are rising worldwide (Church et al. 2013, Chen et al. 2017), raising questions about the future
of coastal ecosystems and built infrastructure (Hinkel et al. 2014, Garner et al. 2017). Rates of sea-level
rise vary regionally due to differences in such things as prevailing winds and shifting ocean currents, and
their impact is further affected by factors such as isostatic rebound following past glaciation, and
groundwater and oil extraction, which affect the behavior of the land surface (Rhamstorf 2017). Our
ability to predict how the world’s coastlines will change is thus limited, yet vitally important given the
concentration of human populations in coastal areas (Kummu et al. 2016) and the wide range of services
that coastal ecosystems provide (Barbier et al. 2011).
The rate of sea-level rise is particularly high along the Atlantic seaboard of North America,
especially in mid-Atlantic and southern New England states, where rates are substantially higher than
the global average (Sallenger et al. 2012). Portions of this region also contain extensive salt marshes and
the consequences of higher sea levels for these marshes has received considerable attention. Models
suggest potential for marshes to build elevation via a combination of increased plant production and
greater rates of sediment deposition, and some evidence suggests that this might be happening (Kirwan
et al. 2016). Sediment flow, however, is declining in many river systems (Syvitski et al. 2005; Weston
2014) and the accretion rates needed to maintain marshes appear to be insufficient in many areas
(Crosby et al. 2016). Accretion rates are higher for lower-elevation marshes, however, and are more
likely to be sufficient to match current rates of sea-level rise in these areas (Kirwan et al. 2016).
Consequently, even where marshes can persist, their elevation relative to sea-level may decline, shifting
the nature of the marsh towards lower-elevations. Evidence for a drop in marsh elevation relative to
sea-level comes from studies that show consistent shifts towards plant species that are associated with
greater tidal flooding (Field et al. 2016; Raposa et al. 2017).
Losses in the overall extent of marshes might also be mitigated, at least somewhat, by migration
of marsh vegetation into adjacent upland areas (Kirwan et al. 2016). Current evidence, however,
suggests that these changes are not happening as quickly as changes within current marshes (Field et al.
2016). Even if some form of marsh can persist over the long term, higher elevation marsh and any
species or ecosystem services that it supports are placed at risk due to the lag time needed for
ecosystems to transition (e.g., Field et al. 2017a). This problem is exacerbated in areas where patterns
of human land use and attitudes towards land protection impose additional constraints on the ability of
marshes to migrate (Field et al. 2017b).
Given the importance of marsh elevation relative to sea-level to discussions about the future of
coastal marshes, it is important that we have good baseline data on current elevations. Locally, this
information can be obtained through the use of surface elevation tables (SETs; Lynch et al. 2015)
installed in the marsh. At a local scale, use of Geographic Positioning Systems (GPSs) in the form of Real
Time Kinematic positioning (RTK) allows for the manual measurement of elevation using a handheld unit
and antenna. Increasingly, local scale information also is being collected using unmanned aircraft
systems (UASs), which can provide contiguous, high resolution data, over moderately large patches of
marsh (Drummond et al. 2015, Ma et al. 2016). Measurements over larger regional scales are harder to
come by without making inferences from remote sensing data, either based on assumptions about
relative elevation derived from vegetation data (Hladik et al. 2013; Correll et al. submitted), or via direct
measurements. Light Detection and Ranging data (Lidar) collected via aircraft offers a high-resolution
(albeit high-cost) and large-scale elevation point cloud which when processed can predict elevation at
1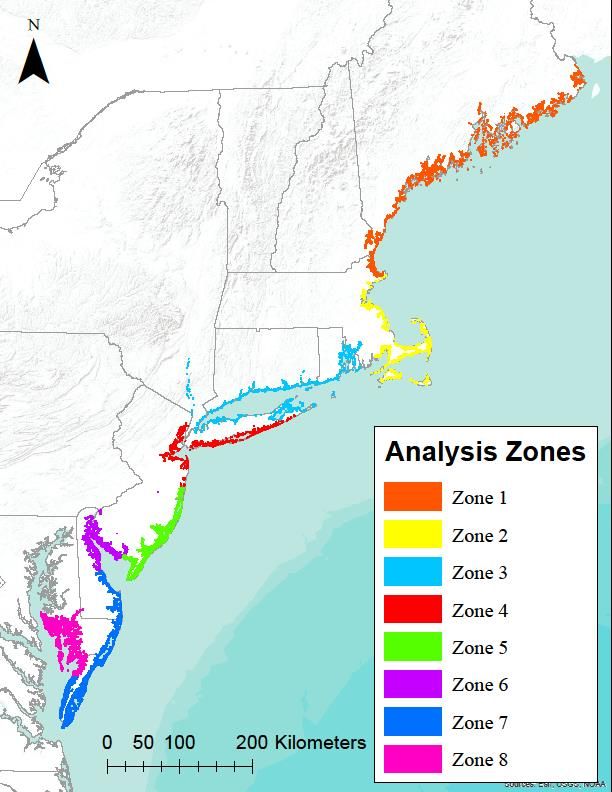
both terrain and vegetation levels (e.g. Hladik et al. 2013, Buffington et al. 2016). The United States
Geological Survey National Elevation Dataset (NED; USGS 2015) also provides a set of elevation data
products at several different resolutions (2, 1, 1/3, 1/9 arc-seconds and 1 m) derived from source data of
topographic maps, manual profiling through photogrammetry, and compiled Lidar. Ideally we would
understand how detailed measurements made at local scales relate to those obtained from these larger-
scale data sets that can be applied over large regions. With this information, we would be better placed
to develop a baseline understanding of current elevation patterns and to track changes over time as sea-
levels rise.
With these needs in mind, we set out to compare measurements of elevation made with
different methods and at different scales. First, we conducted a local-scale study in an extensively
studied coastal wetland near Scarborough, Maine, USA. We compared measurements taken using RTK
units with elevation data created through UAS imagery, and at three different resolutions (1, 1/3, and
1/9 arc-second layers) provided by the NED. Second, we scaled analysis up to compare RTK data
collected at 12,864 points around 655 survey locations between the Canada-Maine border and the
Chesapeake Bay, Virginia, to NED data collected at the same three resolutions previously mentioned.
Third, we describe the elevation profile of marshes throughout the Northeast and mid-Atlantic USA to
provide a general overview of current baseline conditions that can be used for future comparisons.
Finally, we illustrate the relationship between elevation estimates obtained during this study and marsh
habitat types obtained during a parallel study designed to describe vegetation classes across the same
region.
Methods
Data collection
To explore the efficacy of different elevation measurement techniques for tidal marsh, we measured
~10 ha plots (n = 4, Figure 1, 2) developed by Ruskin et al. (2017) in and around Scarborough, ME as part
of a study monitoring demographic rates of tidal marsh birds (Figure 1). Within each plot delineation, we
overlaid a 20 x 20 m grid using ArcGIS 10.3 (ESRI 2016) before navigating to each plot in the field. Once
in the marsh, a trained technician measured all grid points that fell within plot boundaries using a
Trimble R10 RTK sensor system.
In addition to RTK data collection we flew a DJI Phantom 3 Advanced UAS over each marsh plot
in August 2016 to collect imagery for use with photogrammetry software. We programmed the UAS
using the Pix4D Capture mission planning application to fly a grid over each plot at 100 m altitude. The
programmed grid provided images collected with ~80% overlap using a MAPIR 16-megapixel red-green-
blue aerial mapping camera affixed to the base of the UAS with the camera pointing directly downward.
All photographs were collected at wind speeds of less than 15 mph and within 2 hours of local low tide.
We placed 8-9 black and white 20 x 28 cm ground control points on each plot during flights to
georeference digital mosaics during processing. We recorded the location of each ground control point
using a Trimble GEO7X unit, accurate to the decimeter. All imagery was processed using Pix4D
photogrammetry software (Pix4D SA, Switzerland) to create a digital orthomosaic and Digital Surface
Model (DSM) for each marsh plot.
For our regional effort we selected marsh survey sites based on a two-stage cluster sampling
scheme (Wiest et al. 2016) to comprehensively survey tidal marshes for bird populations between
Maine and Virginia. We navigated to a subset of bird survey locations visited by Wiest et al. (n = 655,
Figure 1) between April and September 2015 to sample elevation in tidal marshes within the same study
extent. At each survey location we overlaid a 20 x 20 m point grid on the marsh patch, centering the grid
around the survey location as closely as possible given the orientation of the marsh patch. Technicians
attempted to measure points resulting in a square grid with 5 points on each side, covering a total of 1
ha around each survey location. If a grid point fell in non-marsh habitat, we measured another point
2from the surrounding grid instead. When a marsh patch was too small or narrow for a this size grid to fit
we used either a 5 x 5 m or 1 x 1 m point grid instead (n = 3004 and 530 elevation measurements,
respectively), choosing the resolution that provided the largest grid that could be fit within the marsh
patch. In total, we measured elevation at 12,864 points associated with the 655 survey locations in tidal
marsh between Maine and Virginia.
Once we measured elevation at all points in the field, we overlaid the point for each marsh
elevation reading from both our small-scale and regional study on the 1, 1/3, and 1/9 arc-second layers
(~30 m, ~10 m, and ~1 m resolution, respectively) from the NED (USGS 2015). We then extracted values
from these layers for each sampling point. For our small-scale study we also extracted values from the
UAS-derived DSMs for each RTK sampling point.
To describe elevation data by community type for our regional and fine-scale elevation data, we
used a vegetation layer produced by the Saltmarsh Habitat and Avian Research Program (Correll et al.
submitted, available at https://nalcc.databasin.org/galleries/46d6e771dd6f4fdb8aa5eb46efffffa7)
describing six marsh cover types (high marsh; low marsh; Phragmites australis, hereafter Phragmites;
mudflat; pool/panne; terrestrial border) and two bordering cover types (open water; upland) in tidal
marshes from Maine to Virginia. We extracted the cover type for each RTK point using ArcGIS 10.3 (ESRI
2016).
Statistical analysis
We analyzed our plot-level data using linear and quantile regression using base R and the lqmm package
(Geraci, 2014). Quantile regression allows users to specify the percentage of data points included
beneath the regression line; for reference, an ordinary linear regression maintains a tau = 0.5. We varied
tau from 0.1 to 0.9 (Table 1) and compared these models to linear regressions to identify the best
regression structure to model the data we collected. To account for variation among plots, we included
a plot “site” as a variable in all models. To explore the predictive power of our independent variables
and marsh cover type, we compared models including additive and interactive terms for marsh cover
types with over 10 data points (high marsh, low marsh, pool/panne). We assessed variation explained by
our linear models using adjusted R2. Due to the difficulties in assessing R2 for quantile regression models
we compared models using Akaike’s Information Criterion (AIC, Burnham and Anderson 2004) instead.
We analyzed regional data in a linear mixed-modeling framework (LMM) using the lme4 package
(Bates et al 2015) in Program R (R Core Team 2016). We modeled the accuracy with which NED data can
be used to predict RTK measurements at each spatial scale. We also modelled the relationship between
RTK measurement and latitude using identical modelling methods. Because RTK grids comprised of
individual data points were always grouped around pre-established survey locations, we included a
random variable for “site” in all models. We ran models with and without zero values after examination
of our data because there is an obvious concentration of zero values in the NED dataset. False-zero
values are commonly produced in remotely sensed datasets occurring near water or extremely wet soil.
To explore the utility of measuring additional RTK points to further strengthen the relationships
measured in our regional analysis, we subsetted our data by increments of 10% by (1) removing all
points taken around randomly-selected survey locations and (2) randomly selecting and removing values
from individual points. To explore the predictive power of our independent variables by marsh cover
type, we compared models including additive and interactive terms for marsh cover type using a dataset
of all points falling within the marsh layer. We assessed variation explained and directly compared
models using marginal R2 (Nakagawa and Schielzeth 2013) and AIC.
Results
We measured RTK elevation at a total of 1230 points across the four local plots (Figures 1, 2) in
Scarborough Marsh. Measurements ranged from -0.11 m to 2.0 m ASL ( = 1.31 m). Elevation values
3extracted from the DSM produced from UAS imagery (Figure 2) ranged from -7.2 m to 3.3 m ( = 0.74
m). All points surveyed fell within the marsh cover type layer. We collected 979 overlapping photos for
photogrammetry analysis at these four plots (Figure 2). The processed DSM rasters display elevation
data at a 2 cm resolution.
We measured RTK elevation at a total of 12,864 points centered around 655 survey locations
between Maine in Virginia in 2015. Values measured using RTK ranged from -1.14 m to 2.76 m above
sea level (ASL, = 0.85). Values extracted at these locations ranged from -0.67 m to 5.63 m ASL ( = 1.04
m) for the 1 arc-second NED, -0.67 m to 5.63 m ASL ( = 1.04 m) for the 1/3 arc-second NED, and -0.87
m to 3.18 m ( = 0.84 m) for the 1/9 arc-second NED. Of the RTK points surveyed, 11,328 points fell
within the marsh cover type layer.
For our plot-level data from Maine, we found that RTK elevation measurements were only
weakly predicted using the 1 arc-second ( = 0.14, R2 = 0.27) and 1/3 ( = 0.14, R2 = 0.27) arc-second
NED. The 1/9 arc-second NED had much better predictive power ( = 0.63, R2 = 0.64, Figure 3). RTK
elevation measurements were weakly predicted by DSM values, and fit improved when excluding zero
values ( = 0.14, R2 = 0.28), although most of this variation was explained by the site variable in our
models. Quantile regression models performed better than linear models, with the model using tau =
0.7 ( = 0.07) performing best (Table 1, Figure 4). In our model comparison including marsh cover type,
the best performing model included the additive and interactive term for marsh cover. Generally
speaking, DSM values were higher than corresponding RTK measurements, with the discrepancy greater
at higher elevations.
In our regional analysis, we found that the 1 arc-second ( = 0.49, Rm2 = 0.24, Figure 5a) and 1/3
arc-second ( = 0.20, Rm2 = 0.11, Figure 5b) NED produced only weak predictions of RTK elevation
measurements. This relationship was much improved with the 1/9 arc-second data layer ( = 0.83, Rm2 =
0.85, Fig 5c), especially when excluding zero values ( = 0.87, Rm2 = 0.90, Fig 5d). We found that this
relationship was insensitive to data sub-setting and we obtained similar estimates and explained similar
variance when using as little as 30% of the full dataset ( = 0.81, Rm2 = 0.85). Including both additive and
interactive terms for marsh cover produced a model that out-performed all others (AIC = 29.94
compared to the next best model). We found that most zero-value NED measurements belong to the
“Upland” cover type (Figure 6a), and that regression slopes differed among cover types (Figure 6b). RTK
measurements were generally better predicted by the NED for high marsh and terrestrial border, with
the worst predictions for mudflat. Finally, we found a strong relationship between RTK measurement
and latitude ( =0.19, Rm2 = 0.42, Figure 7).
Discussion
In the northeastern United States, tidal marshes dominate much of the coastal landscape.
Understanding and measuring change in this ecosystem as sea level rise progresses is critical to the
conservation of the system, and the biodiversity and services it supports. In our comparison of elevation
measurements in tidal marshes across different methods and scales, we found that elevation of tidal
marshes was well predicted by the 1/9 arc-second layer of the NED both locally and regionally, however
other data sources (UAS, 1 arc-second NED, 1/3 arc-second NED) provided limited predictive power
(Figure 5). This key finding can help researchers explore local and regional change in tidal marshes
without the need for manual mapping and sampling using RTK units, which can be costly in the
sometimes scarce resources afforded conservation professionals and researchers.
We also found that elevation measurements from the 1/9 arc-second NED diverged least from
RTK measurements in high marsh at both the local and regional scale, and that the relationship was
generally stronger for higher elevation cover types. This result, combined with the improved
performance of models when excluding zero-values, suggest that the NED predicts elevation more
4accurately for higher elevations marsh cover types than those closer to the marine interface. Those
wishing to focus study on lower elevations, therefore, may benefit more from investing in specialized
equipment to manually measure marsh elevation instead of relying on the NED, although predictions
were generally good for most habitat types at a regional scale.
It is important to note that there are known limitations to the NED which should be considered
before using this dataset in any analysis. We found that the 1/9 arc-second NED under-predicts RTK at
both the local and regional scales (regression slope = ~0.85 for both scales). While it is a good predictor
of relative elevation, it should be used cautiously to predict absolute elevation above MSL. Further, the
elevation data that comprise the NED were collected on different dates (2010-2016 for the 1/9 arc-
second layer) and there are no current plans to refresh the 1/9 arc-second layer in the near future.
Instead resources are currently allocated for further development of the spatial extent and temporal
resolution of the 1 m layer, which currently covers a much smaller area than the 1, 1/3, or 1/9 arc-
second layers. Those wishing to measure change over time annually or across 2-3 years may still want to
invest in RTK data collection for best results.
The elevation profile we produced through shows a strong latitudinal trend in tidal marsh
elevation, decreasing from north to south (Figure 7). Tidal marshes occur at higher elevations in the
north and at steadily lower elevations as one moves south along the coastline. This pattern may be
caused by the similar latitudinal gradient in tidal amplitude along the coast, which is driven
overwhelmingly by local bathymetry and geography. The Bay of Fundy in eastern Maine regularly
exhibits tides nearing 10 m, where the Chesapeake Bay experiences tides of only 1-2 m. Because
zonation in plant communities is directly connected to the timing and duration of inundation by
incoming and outgoing tides (Bertness 1991), a larger amplitude tide could result in the once-weekly to
once-monthly inundation window being located in a higher elevation relative to MSL.
While the primary goal of this work was to compare elevation measurements from the NED to
those collected using RTK, we also explored the use of UASs in measuring elevation in tidal marshes. We
found that the use of raw, UAS-derived DSMs do not work well to predict ground-level elevation in tidal
marshes. We believe this is because of the inherent differences in DSM and DEM datasets; DSMs depict
the vegetation surface, while DEMs represent ground-level elevation without consideration of
vegetation. As a result, even our top model shows that our UAS-derived DSM predicts elevations equal
to or higher than the ground elevation estimated using RTK methods. The recent development of Digital
Terrain Models (DTMs) using photogrammetry software such as Pix4D is promising and has the potential
to address the shortcomings we encountered with UAS-derived DSMs. The ease with which a localized
DSM is produced with a UAS is promising, and we expect more accurate tidal marsh elevation profiles to
emerge as the technology and software continue to improve.
Conclusions
The effective measurement of ground elevation is an important step in the effective study and
monitoring of tidal marshes, yet the methods and technology required to produce this type of data are
costly and time-consuming to implement. We found that in tidal marshes of the northeastern USA,
elevation measurements available from the publicly accessible 1/9 arc-second NED were effective in
predicting RTK elevation measurements, explaining 90% of the variation in our regional dataset.
Prediction accuracy was highest for high marsh and terrestrial border, but was good (>80%) for most
cover types. This finding is key for scientists and land managers wishing to measure and monitor marsh
elevation but who are limited by cost and labor availability. Use of the 1/9 arc-send NED can, with
minimal preprocessing, be a suitable stand-in for RTK methods for estimating ground elevation of
coastal marsh, particularly when measuring elevation in high marsh, a cover type of particular
conservation interest due to the ecosystem services it provides for wildlife and humans alike.
5Literature Cited
Barbier EB, Hacker SD, Kennedy C, Koch EW, Stier AC, Silliman BR (2011) The value of estuarine and
coastal ecosystem services. Ecological Monographs 81:169–193.
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effect models using lme4. Journal of
Statistical Software 67:1–48.
Bertness MD (1991) Zonation of Spartina patens and Spartina alterniflora in New England salt marsh.
Ecology 72:138–148.
Bertness MD, Ellison AM (1987) Determinants of pattern in a New England salt marsh plant community.
Ecological Monographs 57:129–147.
Buffington K J, Dugger BD, Thorne KM, Takekawa JY (2016) Statistical correction of lidar-derived digital
elevation models with multispectral airborne imagery in tidal marshes. Remote Sensing of
Environment 186:616–625.
Burnham KP, Anderson RP (2004) Multimodel inference: Understanding AIC and BIC in model sSelection.
Sociological Methods and Research 33:261–304.
Chambers RM, Meyerson LA, Saltonstall K (1999) Expansion of Phragmites australis into tidal wetlands of
North America. Aquatic Botany 64:261–273.
Chen X, Zhang X, Church JA, Watson CS, King M, Monselesan D, Legresy B, Harig C (2017) The increasing
rate of global mean sea-level rise during 1993–2014. Nature Climate Change 7:492–495.
Church JA, Clark PU, Cazenave A, Gregory JM, Jevrejeva S, Levermann A, Merrifield MA, Milne GA,
Nerem RS, Nunn PD, Payne AJ, Pfeffer WT, Stammer D, Unnikrishnan AS (2013) Sea level change.
In: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth
Assessment Report of the Intergovernmental Panel on Climate Change [Stocker TF, Qin D, Plattner
G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM, eds.]. Cambridge
University Press, Cambridge.
Chu-Agor ML, Muñoz-Carpena R, Kiker G, Emanuelsson A, Linkov I (2011) Exploring vulnerability of
coastal habitats to sea level rise through global sensitivity and uncertainty analyses. Environmental
Modelling and Software 26:593–604.
DJI (2016) The Phantom 3 Advanced quadcopter Unmanned Aircraft System. Available at:
https://www.dji.com/phantom-3-adv
Crosby SC, Sax DF, Palmer ME, Booth, HS, Deegan LA, Bertness MD, Leslie HM (2016) Salt marsh
persistence is threatened by predicted sea-level rise. Estuarine, Coastal and Shelf Science 181:93–
99.
Day JW, Christian RR, Boesch DM, Yáñez-Arancibia A, Morris J, Twilley RR, Naylor L, Schaffner L,
Stevenson C (2008) Consequences of climate change on the ecogeomorphology of coastal
wetlands. Estuaries and Coasts 31:477–491.
Donnelly JP, Bertness MD (2001) Rapid shoreward encroachment of salt marsh cordgrass in response to
accelerated sea-level rise. Proceedings of the National Academy of Sciences, USA 98:14218–14223.
Drummond CD, Harley MD, Turner IL, A Matheen, A Nashwan, Glamore WC (2015) UAV applications to
coastal engineering. Australasian Coasts & Ports Conference 2015: 22nd Australasian Coastal and
Ocean Engineering Conference and the 15th Australasian Port and Harbour Conference. Engineers
Australia and IPENZ 2015: 267-272.
Emery NC, Ewanchuk PJ, Bertness MD (2001) Competition and salt-marsh plant zonation: stress
tolerators may be dominant competitors. Ecology 82:2471–2485.
ESRI (2016) ArcGIS Desktop: Release 10.3. Redlands, CA: Environmental Systems Research Institute.
Ewanchuk PJ, Bertness MD (2004) Structure and organization of a northern New England salt marsh
plant community. Journal of Ecology 92:72–85.
Field, CR, Gjerdrum C, Elphick CS (2016) Forest resistance to sea-level rise prevents landward migration
of tidal marsh. Biological Conservation 201:363–369.
6Field CR, Bayard T, Gjerdrum C, Hill JM, Meiman S, Elphick CS (2017a) High-resolution tide projections
reveal extinction threshold in response to sea-level rise. Global Change Biology 35:2058–2070.
Field CR, Dayer AA, Elphick CS (2017b) Landowner behavior can determine the success of conservation
strategies for ecosystem migration under sea-level rise. Proceedings of the National Academy of
Sciences, USA 114:9134–9139.
Garner AJ, Mann ME, Emanuel KA, Kopp RE, Lin N, Alley RB, Horton BP, DeConto R, Donnelly JP, Pollard
D (2017) Impact of climate change on New York City’s coastal flood hazard: Increasing flood heights
from the preindustrial to 2300 CE. Proceedings of the National Academy of Sciences, USA
114:11861–11866.
Geraci M (2014) Linear quantile mixed models: the lqmm package for laplace quantile regression.
Journal of Statistical Software 57:1–29.
Hinkel J, Lincke D, Vafeidis AT, Perrette M, Nicholls RJ, Tol RSJ, Marzeion B, Fettweis X, Ionescu C,
Levermann A (2014) Coastal flood damage and adaptation cost under 21st century sea-level rise.
In: Proceedings of the National Academy of Sciences, USA 111: 3292–3297.
Hladik C, Schalles J, Alber M (2013) Salt marsh elevation and habitat mapping using hyperspectral and
LIDAR data. Remote Sensing of Environment 139:318–330.
IPCC (2013) IPCC Fifth Assessment Report (AR5). IPCC.
Kirwan ML, Guntenspergen GR (2010) Influence of tidal range on the stability of coastal marshland.
Journal of Geophysical Research: Earth Surface 115:F02009.
Kirwan ML, Temmerman S, Skeehan EE, Guntenspergen GR, Fagherazzi S (2016) Overestimation of
marsh vulnerability to sea level rise. Nature Climate Change 6:253–260.
Kummu M, Moel H de, Salvucci G, Viviroli D, Ward PJ, Varis O (2016) Over the hills and further away
from coast: global geospatial patterns of human and environment over the 20th–21st centuries.
Environmental Research Letters 11:034010.
Lynch JC, Hensel P, Cahoon DR (2015) The surface elevation table and marker horizon technique: a
protocol for monitoring wetland elevation dynamics. Natural Resource Report NPS/NCBN/NRR -
2015/1078, National Park Service.
Ma Y, Zhang J, Zhang J (2016) Analysis of Unmanned Aerial Vehicle (UAV) hyperspectral remote sensing
monitoring key technology in coastal wetland. Proceedings volume 9796, Selected Papers of the
Photoelectronic Technology Committee Conferences held November 2015, 97962S. doi:
10.1117/12.2229746.
Miller W, Egler F (1950) Vegetation of the Wequetequock-Pawcatuck tidal-marshes, Connecticut.
Ecological Monographs 20:143–172.
Nakagawa S, Schielzeth H (2013) A general and simple method for obtaining R2 from generalized linear
mixed-effects models. Methods in Ecology and Evolution 4:133–142.
Pennings S, Callaway R (1992) Salt marsh plant zonation: the relative importance of competition and
physical factors. Ecology 73:681–690.
Philipp KR, Field RT (2005) Phragmites australis expansion in Delaware Bay salt marshes. Ecological
Engineering 25:275–291.
R Core Team (2016) R: A language and environment for statistical computing.
Rhamstorf, S (2017) Rising hazard of storm-surge flooding. Proceedings of the National Academy of
Sciences, USA 114:11806–11808.
Ruskin KJ, Etterson MA, Hodgman TP, Borowske AC, Cohen JB, Elphick CS, Field CR, Longenecker RA, King
E, Kocek AR, Kovach AI, O'Brien KM, Pau N, Shriver WG, Walsh J, Olsen BJ (2017). Demographic
analysis demonstrates systematic but independent spatial variation in abiotic and biotic stressors
across 59 percent of a global species range. Auk: Ornithological Advances 134:903–916.
Sallenger AHS, Jr, Doran KS, Howd PA (2012) Hotspot of accelerated sea-level rise on the Atlantic coast
of North America. Nature Climate Change 2:884–888.
7Syvitski JPM, Vörösmarty CJ, Kettner AJ, Green P (2005) Impact of humans on the flux of terrestrial
sediment to the global coastal ocean. Science 308:376–380.
USGS (2015) National Elevation Dataset (NED). Available at: https://nationalmap.gov/elevation.html.
Accessed January 2016.
Weston NB (2014) Declining sediments and rising seas: an unfortunate convergence for tidal wetlands.
Estuaries and Coasts 37:1–23.
Wiest WA, Correll MD, Olsen BJ, Elphick CS, Hodgman TP, Curson DR, Shriver WG (2016) Population
estimates for tidal marsh birds of high conservation concern in the northeastern USA from a
design-based survey. Condor: Ornithological Applications 118:274–288.
Wilson CA, Hughes ZJ, FitzGerald DM, Hopkinson CS, Valentine V, Kolker AS (2014) Saltmarsh pool and
tidal creek morphodynamics: Dynamic equilibrium of northern latitude saltmarshes?
Geomorphology 213:99–115.
Table 1. Model structure, AIC values, and model weights for quantile and linear regressions between
elevation measurements in tidal marshes in Scarborough, Maine, USA, collected using real-time
kinematic (RTK) and digital surface models (DSMs) produced through photogrammetric analysis of
images collected via Unmanned Aircraft System (UAS).
model regression type tau AIC model weight
rtk ~ dsm + site quantile 0.7 0.00 1.00
rtk ~ dsm + site quantile 0.6 12.66 0.00
rtk ~ dsm + site quantile 0.8 44.04 0.00
rtk ~ dsm + site quantile 0.5 68.03 0.00
rtk ~ dsm + site quantile 0.4 164.46 0.00
rtk ~ dsm + site quantile 0.9 206.77 0.00
rtk ~ dsm + site linear . 237.96 0.00
rtk ~ dsm + site quantile 0.3 316.29 0.00
rtk ~ site (null) linear . 426.09 0.00
rtk ~ dsm + site quantile 0.2 537.81 0.00
rtk ~ dsm + site quantile 0.1 883.36 0.00
8Figure 1. Study area and data collection locations for elevation measurement in tidal marshes in the
northeastern USA using real-time kinematic (RTK, purple points) and unmanned aircraft systems (UAS,
yellow polygons, inset).
9Figure 2. Maps and digital surface models (DSMs) of four marsh study plots and elevation measurement
points (green dots) using real-time kinematic (RTK) and unmanned aircraft system (UAS) methods in
Scarborough Marsh, Maine.
10Figure 3. Results of linear regression model comparing tidal marsh ground elevation estimated using
real-time kinematic (RTK) measurements and the National Elevation Dataset (NED) 1/9 arc-second layer.
Data come from four plots in Scarborough, Maine, USA.
Figure 4. Results of quantile regression (tau = 0.7) visualized through A) scatterplot and B) estimates and
confidence intervals of the regression slopes comparing tidal marsh ground elevation estimated using
real-time kinematic (RTK) measurements and Digital Surface Models (DSMs) created via an unmanned
aircraft system (UAS), stratified by cover type. Data come from four plots in Scarborough, Maine, USA.
11Figure 5. Results of mixed-effect linear regressions comparing tidal marsh ground elevation estimated
using real-time kinematic (RTK) measurements and the National Elevation Dataset (NED) from A) 1 arc-
second data layer (Rm2 = 0.24), B) 1/3 arc-second data layer (Rm2 = 0.11), C) 1/9 arc-second data layer
(Rm2 = 0.85), and D) 1/9 arc-second data layer excluding zero values (Rm2 = 0.90). Data come from sites
throughout the northeastern USA, from Virginia to Maine.
12Figure 6. Results of mixed-effects linear regression visualized through A) scatterplot and B) estimates
and confidence intervals of regression slopes comparing tidal marsh ground elevation estimated using
real-time kinematic (RTK) measurements and the National Elevation Dataset (NED) stratified by marsh
community type. Data come from sites throughout the northeastern USA, from Virginia to Maine.
Figure 7. Results of mixed-effect linear regressions comparing tidal marsh ground elevation estimated
using real-time kinematic (RTK) measurements across latitudes between coastal Maine and Virginia,
USA.
13III. Fine-scale mapping of coastal plant communities in the
northeastern USA
(submitted manuscript)
Maureen D. Correll, Wouter Hantson, Thomas P. Hodgman, Brittany B. Cline, Chris S. Elphick, W. Gregory
Shriver, Elizabeth L. Tymkiw, and Brian J. Olsen
Coastal marshes are among the world’s most productive ecosystems and provide significant services to
humans across the globe. These marshes serve as a gateway between land and sea for humans and
wildlife alike, act as a buffer against coastal storms, and provide critical nutrients to marine food webs
(Barbier et al. 2011). Tidal marshes also support and protect biodiversity by providing habitat to marine
and estuarine fish, crustacean populations, and migratory birds (Boesch and Turner 1984; Master 1992;
Brown et al. 2002).
Within the world’s tidal marsh systems, those located along the Atlantic coast of the United
States support the highest number of terrestrial vertebrate specialists described worldwide (Greenberg
et al. 2006). This suite of species includes herpetofauna and mammals, but the majority of described
vertebrate specialists are birds. Several species are limited completely to these marshes during the
breeding season, several of which are in decline (Correll et al. 2017), with extinction predicted for the
saltmarsh sparrow within 50 years (Correll et al. 2017a; Correll et al. 2017b; Field et al. 2017a; Field et al.
in press).
These declining species nest predominantly within the high-marsh zone, one of several
vegetation communities found within coastal marshes. High marsh differs from other marsh areas in
elevation, salinity, and frequency of inundation (Bertness and Ellison 1987; Pennings and Callaway 1992;
Ewanchuk and Bertness 2004) and is characterized by flooding during spring tides linked to the lunar
cycle. In the northeastern United States, the plant species Spartina patens, short-form S. alterniflora,
Distichlis spicata, and Juncus gerardii characterize high-marsh zones, which also include Salicornia spp.,
Glaux maritima, and Solidago sempervirens (Nixon and Oviatt 1973; Bertness 1991; Emery et al. 2001,
Ewanchuk and Bertness 2004). Conversely, low marsh is characterized by daily tidal flooding and is a
near monoculture of tall form S. alterniflora. The surrounding terrestrial border experiences infrequent
inundation by salt water during extreme tides and storms, and is characterized by a more diverse flora
that is often dominated by Iva frutescens and Typha spp. (Miller and Egler 1950; Ewanchuk and Bertness
2004). Introduced Phragmites australis (hereafter Phragmites) also occurs within this ecosystem,
especially around the borders of disturbed marshes (Chambers et al. 1999; Philipp and Field 2005).
These plant community zones can be quickly altered by both natural and anthropogenic
stressors such as sea-level rise, nutrient run-off from adjacent uplands, and the spread of introduced
species (Day et al. 2008). The significant increase in sea level during recent decades poses one of the
largest threats to these marsh ecosystems. As sea levels encroach on the marshes’ seaward side and
upland marsh migration is limited by human-developed coastal infrastructure (Field et al. 2017b) and
upland habitats (Field et al. 2016), a “pinching effect” can occur, resulting in marsh loss. Coastal marshes
can combat rising sea levels through vertical growth, or accretion (Kirwan et al. 2016), but when the rate
of sea-level rise exceeds the rate of accretion, marsh area will decline (Crosby et al. 2016). Rising sea
levels can also drive invasion of high marsh areas with flood-tolerant low marsh species (Donnelly and
Bertness 2001; Field et al. 2016), causing transition from high to low marsh (Kirwan et al. 2016). This
pattern, however, is not ubiquitous to all marshes (Kirwan and Guntenspergen, 2010; Wilson et al.
2014). In addition to sea-level rise, extreme storm events that flood the coastline have been shown to
permanently alter marsh structure within days (Day et al. 2008) and can have a lasting effect on plant
community structure and saltmarsh degradation.
14Marsh degradation and rapid change is likely to continue into the future due to the paired effects of
climate change and human development. Sea levels are expected to rise substantially between 2013 and
2100 (IPCC 2014), and continuing storm events affecting coastal regions are also predicted. The future
distribution of high- and low-marsh habitat therefore remains uncertain (Chu-Agor et al. 2011; Kirwan et
al. 2016). It is essential to develop tools to identify coastal marsh plant communities, particularly high
marsh, on a biologically relevant timescale to protect existing ecosystem services and to inform the
adaptive management of coastal wetlands as habitat for high-marsh specialist species.
The physical and biological characteristics that differentiate high marsh from low marsh and
other marsh plant communities are potentially detectible using remotely-sensed multispectral and
hyperspectral imagery. Both types of imagery can record wavelengths of light outside of the visible
range for humans, with hyperspectral imagery recording reflectance values in much finer detail and
precision (hundreds of individual bands recorded) than multispectral imagery (several wide-ranging
bands recorded, e.g. red green blue, or RGB, imagery). Several studies have previously demonstrated
distinct spectral differences between tidal marsh species using hyperspectral imagery (Rosso et al. 2005;
Belluco et al. 2006; Yang 2009). Such imagery, when combined with elevation data, has previously
produced high-accuracy classifications of tidal marsh vegetation communities, albeit at smaller spatial
scales (e.g. Hladik et al. 2013).
Hyperspectral imagery is costly, however, especially across large landscapes (Adam et al. 2010);
Belluco et al. (2006) compared several aerial and satellite sensors with changing spatial and spectral
resolution, and although hyperspectral imagery performed slightly better than the multispectral, spatial
resolution was the most important factor in classifier performance. Belluco et al. (2006) recommend the
use of multispectral satellite imagery for the mapping of marsh vegetation. Beside the visible spectrum
(RGB), multispectral imagery should also include infrared (IR) reflectance values to allow differentiation
of vegetation types, calculation of vegetation indices (e.g. the Normalized Difference Vegetation Index
or NDVI, Rouse, Haas, & Schell 1974) and detection of soil moisture differences (Jin and Sader 2005;
Pettorelli et al. 2005), particularly in tidal wetlands (Klemas 2011). The IR spectrum has previously been
used as a tool to predict tidal marsh communities both in smaller regions within the northeastern United
States (Gilmore et al. 2008; Hoover et al. 2010; Meiman et al. 2012) and elsewhere (Isacch et al. 2006;
Liu et al. 2010). An exception to this has been in the classification of invasive Phragmites that often
borders tidal marshes. Large-scale classification of this wetland class has met with some success (e.g.
Borgeau-Chavez et al. 2015, Long et al. 2017), however success is limited when using RGB and IR inputs
alone (Samiappan et al. 2017). A large-scale effort to map coastal Phragmites in the northeastern US has
not yet been attempted.
Due to the large spatial scale at which northeastern tidal marshes occur, publicly-available and
low-cost imagery datasets offer the most promising option for repeatedly delineating large swaths of
marsh along the coast. Landsat satellite imagery provided by the National Aeronautic and Space
Administration (NASA) is publicly available multispectral imagery including RGB and IR bands provided at
a 30 x 30 m resolution and is often used for classifying coarse cover types across large landscapes. In the
case of tidal marshes, however, the heterogeneity in tidal marsh vegetation often occurs at scales
smaller than 30 m pixels, and large-scale classifications of tidal marsh plant communities using this
imagery have so far been unfruitful (e.g. Correll 2015). There is thus a clear need for an alternative path
to create a regional classification of tidal marsh vegetation.
Recent advances in high-resolution airborne imagery provide new opportunities to develop
large-scale classifications of coastal plant communities in the northeast, including Phragmites (e.g. Xie et
al. 2015). The National Agricultural Imagery Program (NAIP) from the US Department of Agriculture
(USDA 2016) captures 3-band, high-resolution RGB orthophotos during the growing season. Since 2007
most states have added a Near-InfraRed (NIR) band to the image requirements to aid in the accurate
classification of vegetative cover. The image resolution is 1 m with 6-m horizontal accuracy and a
15maximum of 10% cloud cover. The imagery, freely available for governmental agencies and the public, is
an affordable alternative to commercial aerial and high-resolution multispectral satellite imagery.
Recent applications of NAIP imagery include mapping of tree cover (Davies et al. 2010), forest clearings
(Baker et al. 2013), isolated trees (Meneguzzo et al. 2013), land cover classification (Baker et al. 2013)
and mining activity (Maxwell et al. 2014).
In this study we compare several remote sensing techniques applied to NAIP imagery, elevation
data from the National Elevation Dataset (NED) provided by the US Geological Survey (USGS 2015), and
local tidal information records from the National Oceanic and Atmospheric Administration (NOAA 2016)
to develop an affordable tool capable of repeated classification of high-marsh zones in tidal marshes in
the northeastern United States. We then use the best-performing classifier to categorize marsh
vegetation communities with a 3-m resolution from coastal Maine to Virginia, USA.
Methods
Study site and community types
Our marsh-mapping effort encompasses all salt marshes of the Northeast Atlantic coast of the USA,
from northern Maine to Virginia. To define our classification extent, we applied a 500-m buffer to all
coastal, tidal marsh as delineated by the National Wetland Inventory (NWI, USFWS 2010) estuarine
emergent wetland (E2EM) layer. The study site is further split into 8 subzones (Figure 1) to
accommodate data management and processing.
These coastal marshes vary substantially from north to south. Due to local bathymetric
structure, tidal amplitudes in the Gulf of Maine are among the highest in the world (Garrett 1972), while
those farther south experience much less variation between high and low tides. Similarly, a
preponderance of rocky or highly sloped shorelines in the north limits marshes to small (~10 - 100 ha)
patches, while southern marshes form larger patches of marsh along the coast. Across our study area,
however, tidal marsh ecosystems can be reliably separated into six distinct cover types, plus two
bordering cover types, which we included in our marsh mapping effort:
1. High marsh: Area flooded during spring tides related to the lunar cycle and dominated by
Spartina patens, Distichlis spicata, Juncus gerardii, and short form Spartina alterniflora. Other
species include Juncus roemerianus, Scirpus pungens, Scirpus robustus, Limonium nashii, Aster
tenuifolius, and Triglochin maritima.
2. Low marsh: Area flooded regularly by daily tides and dominated by tall form Spartina
alterniflora.
3. Salt pools/pannes: Depressed, bare areas with sparse vegetation cover and extremely high soil
salinities. Generally, pools retain water between high tides while pannes do not.
4. Terrestrial border: Area infrequently flooded by storm and spring tides and can include areas of
marsh with fresh/brackish water due to a high water table and/or runoff from impervious
surfaces. Typical plant species include Typha angustifolia, Iva frutescens, Baccharis halimifolia,
Solidago sempervirens, Scirpus robustus, and Spartina pectinata.
5. Phragmites: The exotic invasive form of Phragmites australis (subspecies australis). This
subspecies is of considerable management interest (Saltonstall 2002), especially in marshes with
freshwater input, upland development, and/or increased nutrients (Dreyer and Nierling 1995;
Bertness et al. 2002; Silliman and Bertness 2004).
6. Mudflat: Exposed muddy areas free of vegetation.
7. Open water (bordering cover type): Channels and bays leading to open ocean included within
the 500-m buffer.
8. Upland (bordering cover type): All non-marsh terrestrial cover included within the 500-m
buffer.
16Data sources
We collected training data for marsh vegetation classes both in the field and remotely using aerial
imagery, depending on the cover type. We collected training polygons for high marsh, low marsh, and
Phragmites between May and August of 2015 and 2016. Technicians collecting polygon data were
collectively trained at the beginning of the season in salt marsh vegetation identification. Phragmites
polygons mapped were of the invasive Phragmites australis australis and not of the native North
American form Phragmites australis americanus. Tecnnicians used a GEO 7X Trimble GPS (Trimble 2015)
without an external antenna for all community delineation. Horizontal accuracy of this unit without the
external antenna is estimated at < 1m by the manufacturer.
We used a generalized random tessellation stratified (GRTS) sampling framework designed to
sample tidal marsh bird communities across all ownership types (Wiest et al. 2016) to select randomly-
located delineation sites for training data across our study area. Technicians navigated to bird survey
points and located contiguous patches of high marsh, low marsh, and Phragmites larger than 10 x 10 m
as they traversed to each bird survey location. At each patch, they placed a stake flag or other highly
visible marker on the ground to indicate the beginning of polygon delineation and then delineated the
outer boundary of the patch on foot by walking the outer perimeter with the GEO 7X. We collected
training data for open water, pools and pannes, and mudflat cover classes using manual digitization of
2014-2015 1-m NAIP imagery using ArcGIS 10.3 (ESRI 2016) since these cover classes are easily
identifiable in visible wavelength imagery.
We used the most recent digital ortho-photography (RGB and NIR) available from the NAIP
collected during the growing season from 2014 or 2015 as imagery predictor data (see Appendix A for
acquisition year by state). We resampled raw 1 m NAIP imagery to 3 m resolution to match the spatial
scale of the NED, which was used as the digital elevation model (DEM) for this analysis. We also
calculated NAIP imagery derivatives using ArcMap 10.3 using the raw band values. We refer to them as
‘pseudo’-vegetation indices because we used the raw band values instead of reflectance values. In total,
we used the following data inputs as predictor variables: DEM, Raw NAIP Band 1, Raw NAIP Band 2, Raw
NAIP Band 3, Raw NAIP band 4, NDVI, the Normalized Difference Water Index (McFeeters 1996), the
Difference Vegetation Index (Richardson and Wiegand 1977), and the first three principle components
from a principal components analysis (Fung and Ledrew 1987) of the four NAIP bands, which collectively
explained > 95% of the variance.
Marsh habitats are often influenced by their elevation and topographic context and therefore
can often be successfully mapped using elevation data (e.g. Hladik et al. 2013, Maxwell et al. 2016).
Elevational data is particularly helpful in tidal marshes, where topography can drive tidal flooding
frequency and thus can influence plant species zonation (Silvestri et al. 2005). We used the NED for all
elevation predictor data. The NED is derived from different contributed datasets and then processed by
the USGS into a near-continuous DEM at various resolutions across the US. We used 1/9-second (~3 m
resolution) data when available for our classification. When no 1/9 arc-second imagery was available, we
used 1/3 arc-second data (~10 m resolution). To account for the large differences in tidal inundation
across our study area, we collected tidal data for the study area from the closest NOAA tidal gauge
station, creating 29 different tidal zones (NOAA 2016). For each of the stations we collected the
following tidal datums: HAT, MHHW, MHW, MSL, NAVD88 and MAX (Table 1). We resampled the NED
data to an exact 3 m resolution to match the upscaled NAIP imagery and clipped the resulting imagery
with the 500 m buffer around all coastal tidal marsh in the NWI. We further clipped the NED by the 29
tidal gauge zones of the study area, and rescaled each zone to the NAVD88 datum using the NOAA tidal
amplitude data. To calibrate the DEM across the entire study site we used the Mean High Tide (MHT)
divided by Mean Highest High Tide (MHHT) value for each tidal gauge zone as a basis for elevational
differences between marsh zone types.
17You can also read

















































