Species Status Assessment for the Egyptian Tortoise (Testudo kleinmanni) - Photo Credit: Saint Louis Zoo
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Species Status Assessment for the Egyptian
Tortoise (Testudo kleinmanni)
Photo Credit: Saint Louis Zoo
May 2019
U.S. Fish and Wildlife Service
Ecological Services Program
Branch of Delisting and Foreign Species
Falls Church, VA
1
Species Status Assessment for the Egyptian Tortoise (Testudo kleinmanni)
Prepared by the Ecological Services Branch of Delisting and Foreign Species
U.S. Fish and Wildlife Service
EXECUTIVE SUMMARY
This species status assessment report provides the results of a comprehensive review by the U.S.
Fish and Wildlife Service (Service) for the Egyptian tortoise (Testudo kleinmanni) and an
account of the species’ overall viability.
The Egyptian tortoise is a terrestrial species restricted to a narrow coastal zone in Libya, Egypt,
and Israel. It is the only dwarf tortoise occurring in the northern hemisphere, the second smallest
species of tortoise in the world, and the least-known tortoise species in the genus Testudo
inhabiting the Mediterranean basin. The species occurs in the fairly arid region along the
southeastern coast of the Mediterranean Sea, and is mostly found in desert and semi-desert areas,
shoreline grasses and at the edges of salt lakes or salt marshes, and scrub thorn areas. The current
distribution is highly fragmented due to human impacts and natural factors.
Stressors influencing the Egyptian tortoise are similar across the species’ range, although the
magnitude of stressors may vary between Libya, Egypt, and Israel. Severe habitat loss and
degradation caused by expansion of agriculture, overgrazing, urban development, and military
activities affect the Egyptian tortoise throughout its range. Habitat loss may also occur due to
changing environmental conditions from climate change, i.e., increasing temperatures and
decreased precipitation. Collection for the pet trade is a significant stressor in Libya. The
Egyptian tortoise is nearly extirpated from Egypt partially because of historical collection for the
pet trade. Egyptian tortoises are not substantially collected in Israel.
The Egyptian tortoise is afforded some protection based on existing regulations in each of the
range countries. However, these regulations have had varying success conserving the species and
its habitat from habitat loss and degradation and collection of the species for the pet trade.
Egyptian tortoises are actively traded internationally despite their status in Appendix I of CITES
(Convention on International Trade in Endangered Species of Wild Fauna and Flora). Protected
areas in Egypt at Zaranik Protected Area and in Israel at Holot Agur provide some protection to
the Egyptian tortoise from habitat conversion and collection pressure. The IUCN Red List has
assessed the conservation status of the Egyptian tortoise as Critically Endangered, indicating a
≥80% risk of extinction.
To evaluate the biological status of the Egyptian tortoise, we assessed a range of conditions to
allow us to consider the species’ resiliency, redundancy, and representation (together, the 3Rs)
now and into the future. The Egyptian tortoise needs populations resilient enough to withstand
stochastic events. The species also needs redundancy of multiple populations distributed across
its range, in each country, and on both sides of the Nile River, to withstand catastrophic events.
2
Maintaining representation on both sides of the Nile River is essential because there is some
ecological diversity between the habitats west and east of the river.
We assessed the level of resiliency, redundancy, and representation currently and into the future
using three potential scenarios for the species based on the current stressors and population trend.
The three scenarios we developed are: (1) Status quo scenario in which the stressors and
population levels would continue on the same trajectory; (2) Reduced collection scenario in
which the collection of Egyptian tortoises for the pet trade in Libya would decrease; and (3) In
situ scenario in which the reduced collection scenario (Scenario 2) is combined with an in situ
program that reintroduces the species into existing protected areas and includes habitat
management of protected areas in Egypt. These areas would be managed similar to the
management of the wild population at Zaranik Protected Area
Overall, the results of our analysis indicate the Egyptian tortoise has low to moderate resiliency
due to small fragmented populations, habitat loss and degradation throughout the species range,
and collection pressure for the pet trade, particularly in Libya. We assessed the potential for
implementing conservation actions for the species, such as reducing collection pressure in Libya
and implementing habitat management and in situ programs in existing protected areas.
However, there is uncertainty of the successfulness of conservation actions given resource
limitations and ongoing impacts to the habitat and tortoise populations.
3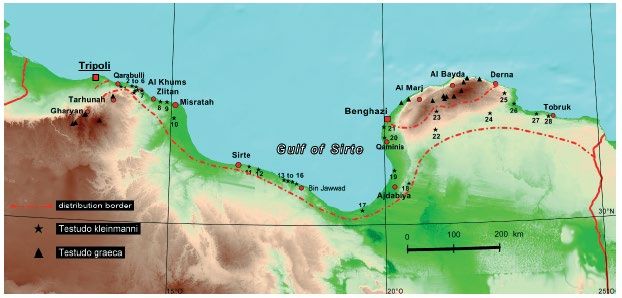
Table of Contents
EXECUTIVE SUMMARY ............................................................................................................ 2
CHAPTER 1. INTRODUCTION ................................................................................................... 6
CHAPTER 2. LIFE HISTORY AND SPECIES NEEDS .............................................................. 8
Taxonomy.................................................................................................................................... 8
Species Description ..................................................................................................................... 9
Habitat and Shelter .................................................................................................................... 10
Burrows and Shrubs .............................................................................................................. 11
Life History ............................................................................................................................... 12
Behavior................................................................................................................................. 12
Life Cycle of Egyptian tortoise ............................................................................................. 13
Life Span and Survival .......................................................................................................... 15
Individual and Species Needs ................................................................................................... 16
Population and Species Needs................................................................................................... 17
Historical and Current Distribution ....................................................................................... 17
Historical and Current Population Size ................................................................................. 22
Population and Species Needs for Viability.............................................................................. 27
Resiliency .............................................................................................................................. 28
Representation ....................................................................................................................... 28
Redundancy ........................................................................................................................... 28
CHAPTER 3. CURRENT CONDITION ..................................................................................... 29
Description of Stressors ............................................................................................................ 29
Agriculture and Grazing ........................................................................................................ 29
Urban Encroachment and Development ................................................................................ 32
Military Activities and Vehicles ............................................................................................ 33
Desertification ....................................................................................................................... 33
Collection............................................................................................................................... 34
Predation ................................................................................................................................ 38
Effects of Climate Change ..................................................................................................... 39
Current Conservation Efforts .................................................................................................... 40
4
International ........................................................................................................................... 40
Egypt...................................................................................................................................... 40
Libya ...................................................................................................................................... 43
Israel ...................................................................................................................................... 44
Summary of Current Condition ................................................................................................. 45
Resiliency .............................................................................................................................. 46
Representation ....................................................................................................................... 48
Redundancy ........................................................................................................................... 48
CHAPTER 4: FUTURE CONDITIONS ...................................................................................... 49
Future Scenario 1 – Status Quo ................................................................................................. 51
Resiliency .............................................................................................................................. 51
Representation ....................................................................................................................... 51
Redundancy ........................................................................................................................... 52
Future Scenario 2 – Reduced Collection from Libya ................................................................ 52
Resiliency .............................................................................................................................. 52
Representation ....................................................................................................................... 52
Redundancy ........................................................................................................................... 53
Future Scenario 3 – In situ Programs and Reduced Collection................................................. 53
Resiliency .............................................................................................................................. 53
Representation ....................................................................................................................... 53
Redundancy ........................................................................................................................... 54
CHAPTER 5: UNCERTAINTY ................................................................................................... 54
Population Estimate................................................................................................................... 54
Future Scenarios ........................................................................................................................ 55
South Sinai Population .............................................................................................................. 55
Appendix A. Species Status Assessment Summary for Egyptian tortoise ................................... 56
Appendix B. Resiliency: Current and Future Condition Tables (See Table 5 for scoring
methodology) ................................................................................................................................ 58
LITERATURE CITED ................................................................................................................. 60
5
CHAPTER 1. INTRODUCTION
We received a petition on June 9, 2014, from Friends of the Animals to list the Egyptian tortoise
as threatened or endangered under the Endangered Species Act (Act). On April 10, 2015, we
published in the Federal Register (80 FR 19259) a 90-day finding that found that the petition
presented substantial scientific and commercial information indicating that the petitioned action
may be warranted.
This report summarizes the results of a species status assessment (SSA) for the Egyptian tortoise
(Testudo kleinmanni). The SSA framework is intended to be an in-depth review of a species’
biology and threats, an evaluation of its biological status, and an assessment of the resources and
conditions needed to maintain long-term viability. An SSA does not result in a decision by the
Service on whether this taxon should be proposed for listing as a threatened or endangered
species under the Act, but instead provides a review of the available information strictly related
to the biological status of the Egyptian tortoise. The Service will make a decision after reviewing
this document and all relevant laws, regulations, and policies. We will publish any proposed
listing decision in the Federal Register with opportunities for public input.
Using the SSA framework (Figure 1), we considered what the species needs to maintain viability
by characterizing the status of the species in terms of the 3Rs: Resiliency, Redundancy, and
Representation (Service 2016, entire; Smith et al. 2018, entire). For the purpose of this SSA, we
generally define viability as the ability of species to sustain populations in the wild over time.
Figure 1. Species Status Assessment Framework
6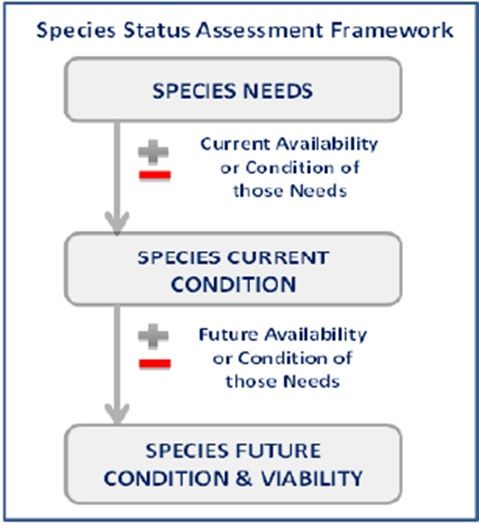
• Resiliency describes the ability of populations to withstand stochastic events (i.e., those that
arise from random factors). For example, we can measure resiliency based on metrics of
population health, birth versus death rates, or population size. Highly resilient populations
are better able to withstand random disturbances such as drought or the effects of
anthropogenic activities.
• Representation describes the ability of a species to adapt to changing environmental
conditions. We measure representation by the breadth of genetic or environmental diversity
within and among populations. Representation gauges the probability that a species is
capable of adapting to environmental changes. The more representation (or diversity) a
species has, the more it is likely capable of adapting to changes (natural or human caused) in
its environment. In the absence of species-specific genetic and ecological diversity
information, we evaluate representation based on the extent and variability of habitat
characteristics across the geographical range.
• Redundancy describes the ability of a species to withstand catastrophic events. Measured by
the number of populations, their resiliency, and their distribution (and connectivity),
redundancy gauges the probability that the species has a margin of safety to withstand or
bounce back from catastrophic events involving many populations.
To evaluate the biological status of the Egyptian tortoise, both currently and into the future, we
assessed a range of conditions to allow us to consider the species’ resiliency, redundancy, and
representation (the 3Rs). This SSA report provides a thorough assessment of biology and natural
history and assesses demographic risks, stressors, and limiting factors in the context of
determining the viability and risks of extinction for the species.
The SSA report discusses (1) life history and species needs for the Egyptian tortoise (Chapter 2);
(2) the current condition for the Egyptian tortoise (Chapter 3); the (3) future condition scenarios
for the Egyptian tortoise, including a description of the viability in terms of resiliency,
redundancy, and representation (Chapter 4), and uncertainties in the analysis (Chapter 5). It
represents a compilation of the best available scientific and commercial information and a
description of past, present, and likely future risk factors to the Egyptian tortoise.
7
CHAPTER 2. LIFE HISTORY AND SPECIES NEEDS
Taxonomy
The Egyptian tortoise (Testudo kleinmanni) is a reptile species of the order Testudines (formerly
Chelonia) (i.e., turtle, terrapin, or tortoise) and the family Testudinidae (Integrated Taxonomic
Information System (ITIS) 2014, unpaginated). The Egyptian tortoise was recorded from Egypt
in 1875 near Alexandria, Egypt and subsequently named after the collector; as Testudo
kleinmanni (Anderson 1898, p. 29; Buskirk 1985, p. 35; Buskirk 1993, unpaginated).
A specimen, which was believed to be the type specimen, was collected from Sindh, Baluchistan
(now northwestern Pakistan) that was initially presented in 1869 and described as Testudo leithii
(Gunther 1869, p. 502). However, no other example of this species has been recorded from Sindh
or from any locality to the east of southern Syria (Anderson 1898, p. 29; Perälä 2001, p. 567).
The fact that no other example of the species has been recorded in Sindh, Baluchistan, or in any
other part of India, renders it probable that the specimen was not a native of its reputed type-
locality (Flower 1933, p. 745; Anderson 1898, p. 29).
The Egyptian tortoise named as Testudo kleinmanni was considered a junior synonym of Testudo
leithii until it was determined that the name Testudo leithii was previously allocated to a fossil
freshwater turtle from India, found in 1852; therefore, the name of Testudo leithii was invalid
(Loveridge and Williams 1957, p. 277; Perälä 2001, p. 567; Connor 1993, unpaginated). A new
subgenus was proposed for the Egyptian tortoise, Pseudotestudo (Loveridge and Williams 1957,
p. 276) as opposed to the genus Testudo, but this new proposed subgenus was not accepted
(Delfino et al. 2009, p. 858). Therefore, the specimen collected in Alexandria, Egypt, retained
the scientific name of Testudo kleinmanni.
In 2001 the Egyptian tortoise (Testudo kleinmanni) was proposed to be split into two distinct
species: T. kleinmanni west of the Nile River (African population) and T. werneri east of the Nile
River (Israeli population), based on shell morphology and geographic separation (Perälä 2001, p.
570). The strong morphological heterogeneity of the two allopatric (related species occurring in
separate non-overlapping) populations was interpreted as clear evidence for a divergent
evolutionary history and the formation of two genealogical lineages within T. kleinmanni (Perälä
2001, p. 570). However, other studies do not consider these populations west and east of the Nile
River to be distinct species because:
1. No diagnostic features exist that can separate populations east and west of the Nile Valley
consistently (e.g., abdominal and pectoral plastron markings);
2. Differences could be due to shell wear because of more abrasive substrates;
3. The statistical methods used to evaluate the difference in shell dimensions have several
technical errors, rendering most of no significance, and vulnerable to experiment-wise
Type 1 error;
4. Shell morphology has proven repeatedly to be of limited taxonomic value in Chelonians
at large due to the elasticity of the shell in response to a variety of factors, including
environment and diet;
5. Molecular analysis has shown a very small genetic distance between the two populations,
much smaller than that expected between two species; and
86. The ecology and biology of the two populations are identical (Attum et al. 2007a, pp.
397-400; Baha El Din 2006, p. 324).
The subtle morphological differences of the western and eastern populations represent merely
another example of population-specific variation and phenotypic plasticity in Testudo species
(Siroky and Fritz 2007, p. 230). Therefore, the eastern and western populations of the Egyptian
tortoise should be considered as a single species. In addition, the Nile River has not acted as a
continuous biogeographical barrier (Attum et al. 2007a, pp. 399–400), allowing ample
opportunities for terrestrial fauna to move from one side to the other over an extended period of
time (Attum et al. 2007a, p. 400).
Considering the lack of differentiation between the populations west and east of the Nile River,
and that mitochondrial DNA variation reasonably reflects species borders in other Testudo
species (Fritz et al. 2005, 2006, 2007, in Siroky and Fritz 2007, p. 3), the eastern population
referred to as T. werneri was determined to be a junior synonym of T. kleinmanni (Baha el Din
2006, p. 324; Siroky and Fritz 2007, p. 230; Attum et al. 2007a, p. 399). In zoological
nomenclature, a synonym is each of two or more scientific names of the same taxonomic rank
that applies to the same taxon; the earliest published name is the senior synonym and the later
name is the junior synonym (International Code of Zoological Nomenclature 2019, unpaginated).
Therefore, the valid taxonomic status of the Egyptian tortoise is Testudo kleinmanni (ITIS 2014,
unpaginated; Perälä 2003, unpaginated). Testudo leithii is accepted as a synonym (IUCN 2014,
p. 1) and Testudo werneri is accepted a junior synonym (Attum et al. 2007a, p. 399; see also
Turtle Taxonomy Workling Group, Rhodin et al. 2017).
Species Description
The Egyptian tortoise is the only dwarf tortoise occurring in the northern hemisphere, the second
smallest species of tortoise in the world (Woodland Park Zoo 2014, p. 1), and the smallest and
least-known tortoise species inhabiting the Mediterranean basin (Buskirk 1985, p. 35, 37). In
appearance, the Egyptian tortoise resembles a small Greek tortoise (Testudo graeca), the species
to which the Egyptian tortoise appears most closely related (Buskirk 1985, p. 36). The most
distinguishing characteristic of the Egyptian tortoise is its remarkably small size (Highfield and
Martin 2014, p. 1).
Figure 2. (a) Egyptian tortoise; (b) V-shaped markings on the abdominal scutes
9Females are generally a bit larger than males (Woodland Park Zoo, p. 1; Buskirk 1985, p. 36)
with the sexes of adult tortoises most easily known by the tail; the female has a short pointed tail
and the male has a longer tail (Flower 1933, p. 746). The carapace (top shell) length in a straight
line (SCL) for adult males is almost always between 90 and 100 mm (3.54–3.93 in); adult
females have carapaces usually over 110 mm (4.33 in) (Flower 1933, p. 746). New hatchlings
have a SCL of about 33 mm (1.3 in) (Flower 1933, p. 746). The weight of females is
approximately 300–350 g (10.6–12.4 oz) and males is 160–250 g (5.6–8.8 oz) (Woodland Park
Zoo, p. 1; Buskirk 1985, p. 36). Individuals in captivity may be larger (Flower 1933, p. 746).
The head, neck, limbs, feet, nails, and tail vary from yellow to yellowish-brown to ivory colored
(Loveridge and Williams 1957, p. 280; Flower 1933, p. 748; Highfield and Martin 2014, p. 1;
Ernst et al. 2014, p. 1). Hatchlings are completely yellow (Loveridge and Williams 1957, p. 279)
The button-like eyes are strikingly black and the irides in both sexes are shiny black and very
conspicuous (Loveridge and Williams 1957, p. 280; Flower 1933, p. 748).
The shell (carapace – top shell and plastron – bottom shell) is composed of hard, bone plates
covered by scutes (bony plates) made of keratin. Both parts are fused together at the sides by a
bridge. The high-domed carapace is pale yellow with lemon and yellow-green shades, with each
scute edged with brown or black (Buskirk 1985, p. 36; Loveridge and Williams 1957, p. 279;
(Woodland Park Zoo 2014, p. 1). These marks vary in individuals, regardless of sex or locality;
they may be strong and broad, wide or narrow, or merely outlines to the shields (Flower 1933, p.
749; Loveridge and Williams 1957, p. 279; Ernst et al. 2014, p. 1). The centers of the carapacial
scutes bear no dark pigment (Buskirk 1985, p. 36). The Egyptian tortoise usually has much less
dark pigment on the carapace than any other Testudo species (Buskirk 1985, p. 36).
The plastron is greenish to yellow (Loveridge and Williams 1957, p. 279). The vast majority of
specimens feature two V-shaped brown or black markings upon the abdominal scutes, which is a
symmetrical dark triangular figure with the base being along the abdomino-pectoral seam and the
apex toward the tail (Buskirk 1985, p. 36) (see Figure 2b, above). This feature is quite different
from the abdominal marks seen on the plastron of other Palaearctic land-tortoises – Greek
tortoise, Hermann’s tortoise (Testudo hermanni), Marginated tortoise (Testudo marginata), and
Russian tortoise (Testudo horsfieldii) (Flower 1933, p. 749; Highfield and Martin 2014, p. 1).
Habitat and Shelter
The Egyptian tortoise occurs in the fairly arid region along the southeastern coast of the
Mediterranean Sea (Baha El Din 1994, p. 1, Turtle Taxonomy Working Group, Rhodin et al.
2017) and is mostly found in desert and semi-desert areas, shoreline grasses and at the edges of
salt lakes or salt marshes, and areas of scrub thorn (Lortet 1887, and Werner 1982, in Buskirk
1985, p. 40; Maryland Zoo 2015, p. 1; Ernst et al 2014, p. 1). The species prefers areas of sandy
soils and dunes with a fair amount of cover of bushes and small shrubs and solidified sands with
denser plant cover. However, it does not appear to be restricted to soft sand habitats (Baha El
Din 1994, p. 4). In parts of Egypt, the species was recorded from regions with very little soft
sand or dunes, or none at all. Past records from the North Coast of Egypt include areas
dominated by compacted sand and gravel plains with scattered rocks and shallow sandy wadis
(valley, ephemeral ravine, or channel), very different from the dune fields of North Sinai, Egypt
and the western Negev Desert in Israel (Baha El Din 1994, p. 4). The species also invades
cultivated fields and gardens (Lortet 1887, in Buskirk 1985, p. 40). The Egyptian tortoise occurs
10at elevations below 380 m (1,247 ft), which is the highest altitude in Africa at which the tortoise
has been found, about 30 km (18.6 mi) south of Täknis, on the southern slopes of the Jebel
Akhdar (Green Mountains) in Libya (Schneider and Schneider 2008, p. 144).
Throughout much of the Egyptian tortoise’s habitat, the woody plant Artemisia monosperma (no
common name) is the dominant plant providing the basic habitat complexity and cover that the
species requires (Baha El Din 1994, p. 23). Artemisia monosperma is widespread in the desert
plains and wadis, both inland and in the Mediterranean coastal region (Badr et al. 2012, p. 67). It
is the most important dwarf shrub of the sand vegetation in the coastal sands, the western Negev,
and North Sinai (Danin 2019, unpaginated). Other common shrubs and grasses include white
broom (Retama raetam), desert grass (Panicum turgidum), sabat (Stipagrostis scoparia),
European tea-tree (Lycium europaeum), bushy bugloss (Echiochilon fruticosum), creeping thorn
rose (Neurada procumbens), and callous-leaved gromwell (Moltkiopsis ciliate) (Geffen and
Mendelssohn 1989, p. 404). In Libya, the dunes contain shrubs such as A. monosperma, Ephedra
sp. and Atriplex sp., and in the early spring, numerous annual flowering plants carpet the dune
valleys (Schneider and Schneider 2008, pp. 147–148). Because of human impacts, most of the
areas where Egyptian tortoises occur have about 5–10 percent plant cover, which is unusually
open habitat for a tortoise species, although plant cover may reach up to 40 percent in areas not
degraded by overgrazing (Mendelssohn 1982, p. 134; Buskirk 1985, p. 46;) and up to 70 percent
in areas distant from human impacts (Zahran and Willis 1992, in Baha El Din 1994, p. 15).
Burrows and Shrubs
Bushes, shrubs, and rodent burrows are essential to the survival of the Egyptian tortoise (Geffen
and Mendelssohn 1989, p. 408; Mendelssohn 1982, p. 134). The use of burrows for cover is well
known in small land tortoises (Voigt and Johnson 1976, entire; Douglass and Layne 1978,
entire). The Egyptian tortoise uses existing rodent burrows if available, or may dig their own
when necessary in areas where rodent burrows are scarce; these burrows are always dug under
bushes or shrubs (Groombridge 1982, p. 134; Mendelssohn 1982, p. 134). The small body size of
the Egyptian tortoise provides a great advantage, considering the energetic cost of burrow
excavation (Woodbury and Hardy 1948, in Geffen and Mendelssohn 1989, p. 407). During
inactive periods, most Egyptian tortoises in Israel were located in rodent (Gerbil (Gerbilus) and
(Meriones) spp.) burrows, which were especially abundant in the valleys, either under bushes or
in harder soil (Geffen and Mendelssohn 1989, pp. 405–406). Although in Libya, tortoises hide in
the root area of shrubs and half-bury themselves in the sand and are rarely found sheltered in the
entrances of rodent burrows (Schneider and Schneider 2008, p. 148).
Thermal buffering of shrubs is essential to the Egyptian tortoises’ long-term survival. Larger
shrubs serve as thermal refuges that stabilize micro-climatic variation, and are especially
important during aestivation because smaller shrubs are not adequate thermal refuges in the
summer (Attum et al. 2013, p. 76). The summer months are characterized by daily, potentially
lethal maximum daytime temperatures of approximately 32 degrees Celsius (°C) (90 degrees
Fahrenheit (°F)) along the Mediterranean Coast and even hotter in the desert and other interior
areas (Weather Channel 2019, unpaginated; Weather and Climate 2019, unpaginated). The
diameter of shrubs that tortoises use during aestivation increases to approximately 1 m (3.3 ft) or
greater (Attum et al. 2013, p. 77). The use of large shrubs helps the tortoise to reduce the range
of temperature fluctuation, which in turn maintains relatively stable body temperatures and
reduces metabolic costs of prolonged inactivity (Beck and Jennings 2003, and Kennet and
11Christian 1994, in Attum et al. 2013, p. 77). Therefore, the benefits of shade provided by large
shrubs as thermal refuges is more valuable during this period when the tortoises aestivate (Attum
et al. 2013, p. 77).
During active periods when Egyptian tortoises are not aestivating or in prolonged dormancy (fall,
winter, and spring seasons), they use smaller shrubs as refuge when the threat of overheating is
reduced due to overall cooler ambient air temperatures (Attum et al. 2013, p. 77). The use of
small shrubs as refuge outside of the summer months may also be advantageous for
thermoregulatory purposes because it allows tortoises to bask and be exposed to sunlight while
being partially concealed from predators (Attum et al. 2013, p. 77). The small size of Egyptian
tortoises allows them to warm quickly after cold desert nights, and their reflective, rather than
absorptive, carapace color allows them to extend their time foraging during the day before the
desert heat drives them under cover (Tabaka and Senneke 2003, unpaginated). No inactive
tortoises are found in the open (Geffen and Mendelssohn 1989, p. 406).
Life History
Behavior
Egyptian tortoises’ activity is largely confined to the cooler part of the year, roughly October
through May or early June. However, by April activity is much reduced. The peak in activity is
from December to March, but tracks are occasionally seen as early as October and as late as May
(Geffen and Mendelssohn 1989, p. 405; Mendelssohn 1982, p. 134). Advantages to being active
during the cooler part of the year include (1) An abundance of food because annual plants are
available during winter and spring (Geffen and Mendelssohn 1989, p. 407); (2) lower potential
for predation by species such as the short-toed eagle (Circaetus galicus), the Egyptian vulture
(Neophren percnopterus) that are summer visitors (Mendelssohn, 1982, p. 135; Geffen and
Mendelssohn 1989, p. 407), and the tortoises’ main predator the gray monitor (Varanus griseus)
that is active during the day and begins its activity in April (Mendelssohn 1982, p. 134; Geffen
and Mendelssohn 1989, p. 407); and (3) the tortoises’ small body size allow them to reach
optimal temperature rapidly, even on cool winter days (Geffen and Mendelssohn 1989, p. 407).
Activity ceases on cold, cloudy, or rainy days in the winter and during overcast and extremely
hot, dry days in spring (Geffen and Mendelssohn 1989, p. 406).
During the summer tortoise activity decreases (Geffen and Mendelssohn 1989, p. 406). Egyptian
tortoises typically aestivate or experience prolonged dormancy starting from mid-May or early
June through the end of September, a period characterized by extremely high ambient
temperatures, no rainfall, and the lowest food availability (Attum et al. 2006, 2007b, 2008, in
Attum et al. 2013, p. 74; Geffen and Mendelssohn 1989, p. 406).
Figure 3: Months during the year when Egyptian tortoises are active (blue) and aestivate (red).
Month Jan Feb Mar Apr May Jun Jul Aug Sep Oct Nov Dec
Active
Aestivate
The decrease in activity following rising mean maximum ambient temperatures over 30°C
(86°F) reflects the strong influence of environmental temperature on the activity of the Egyptian
12tortoise, as is found in other desert species (Rose and Judd, 1975, p. 453; Voigt and Johnson
1976, in Geffen and Mendelssohn 1989, p. 408). However, active tortoises have been found at air
temperatures ranging from 15.3 to 35.4°C (59.5 to 95.7°F) and a larger range of surface
temperatures of 17 to 45°C (62.6 to 113°F) (Mendelssohn 1982, p. 134; Geffen and Mendelssohn
1989, p. 406; Attum et al. 2007b, p. 401). More than 50 percent of tortoises were active within
air temperatures of 21 to 27°C (69.8 to 80.6°F), although large numbers of individuals were
inactive in the same air temperature range (Geffen and Mendelssohn 1989, p. 406).
Two major factors that seem to stimulate the onset of aestivation in the Egyptian tortoise are
rising ambient temperature and withering of food plants (Ernst et al. 2014, p. 1; Geffen and
Mendelssohn 1989, p. 408). Captive individuals at the Canadian Center of Ecological Zoology at
Tel-Aviv University in Israel remained active during summer with occasional ambient air
temperatures over 30°C (86°F) as long as succulent food was offered, which may suggest that
withering of annual plants in late spring to early summer is the major factor influencing the onset
of aestivation in the Egyptian tortoise (Geffen and Mendelssohn 1989, p. 408).
Figure 4. (a) Simple life cycle diagram of Egyptian tortoise, which consists of five main life
stages: egg, hatchling, juvenile, subadult, and adult. Included (b) are the months in which wild
and captive Egyptian tortoises are known to mate, lay eggs, and hatch (Geffen and Mendelssohn
1991; Mendelssohn 1982; Highfield and Martin 2014).
Life Cycle of Egyptian tortoise
(a)
Adult
Mature at
10 years
Egg
Subadult
Hatch after
3 -10 years
70-90 days
Hatchling
Juvenile post
1-3 years hatching to
1 year
(b)
January February March April May June July August September October November December
Mating/Copulation
Wild
Captive
Egg Laying
Wild
Captive
Hatching
Wild
Captive
13Testudines are characterized by oviparity (eggs laid with little or no other embryonic
development within the mother), lack of parental care, high and variable egg and juvenile
mortality, and high adult survival rates (Wilbur and Morin 1987, p. 389; Reptiles of the World
2015, unpaginated). Testudines are slow-maturing populations that depend on long reproductive
lifespans of adults, which produce rates of population increases that are among the lowest
observed in vertebrates (Wilbur and Morin 1987, in Díaz-Paniagua et al. 2001, p. 707).
The Egyptian tortoise breeding strategy can be categorized as type II, characterized by a small
number of relatively large-size eggs, a prolonged nesting period, and solitary nesting site that
does not require specific location or structure (Geffen and Mendelssohn 1991, p. 576). Little is
known about the reproduction of the Egyptian tortoise in the wild (Mendelssohn 1982, p. 136;
Geffen and Mendelssohn 1991, p. 574). Much of the information comes from captive breeding
but that is also limited (Geffen and Mendelssohn 1991, p. 576; Flower 1933, p. 746;
Mendelssohn 1982, p. 136; Highfield and Martin 2014, pp. 2–3).
Mating has only been observed in March in the wild (Geffen and Mendelssohn 1991, p. 575). In
captivity, mating has been observed February through April as well as August through
November (Woodland Park Zoo 2014, p. 1; Geffen and Mendelssohn 1991, p. 574; Mendelssohn
1982, p. 136; Mendelssohn 1982, p. 136). Laying eggs normally occurs at monthly intervals;
clutches of 1 to 5 eggs may be deposited (Highfield and Martin 2014, pp. 2–3; Geffen and
Mendelssohn 1991, p. 575). The female digs a hole the depth of her outstretched hindleg,
preferring sunlight exposure under a bush (Ernst et al. 2014, p. 1). In the wild, eggs are laid from
March through the end of June (Geffen and Mendlessohn 1991, p. 576). In captivity, eggs are
laid between the end of April and end of July (Mendelssohn 1982, p. 136; Flowers 1933, p. 746)
(see Figure 4c).
In the wild, eggs are incubated 70 to 90 days (Geffen and Mendelssohn 1991, p. 575). In
captivity, incubation lasts longer with a range of 78 to 119 days at 30ºC (86ºF), which is the
normal suggested temperature for most Testudo species (Mendelssohn 1982, p. 136; Highfield
and Martin 2014, p. 3). However, this incubation temperature may not be the ideal one, as
hatchlings can turn out to be all males (Mendelssohn 1982, p. 136; Highfield and Martin 2014, p.
3). The influence of incubation temperature on the sex of the hatchlings is well known for
several turtles, with higher temperatures producing more females (Bull and Vogt 1979, pp.
1186–1187).
Hatching in the wild occurs in summer and autumn (Mendelssohn 1982, p. 134; Geffen and
Mendelssohn 1991, p. 575). In captivity, hatching is observed to occur in August through
November and also in January and February. The hatchlings are believed to remain largely
inactive and take shelter from the heat until the weather cools (Tortoise Trust 2014,
unpaginated). Hatchlings may also wait inside the egg to absorb the yolk sac (Highfield and
Martin 2014, p. 3).
In the wild, it is likely that Egyptian tortoises do not reproduce at all during drought years
(Mendelssohn 1982, p. 136). Sperm storage for long periods is well documented in terrestrial
tortoises (Moll, 1979, in Geffen and Mendelssohn 1991, p. 576). Females may use the stored
14sperm to fertilize later egg clutches, and sperm storage can serve as a means for successful
mating if reproductive cycles are asynchronous (Perce and Avise 1998, p. 209; Gist and
Congdon 1998, p. 532; Palmer et al. 1998, pp. 703–705). Sperm storage may be especially
important in species with relatively low-density populations, such as the Egyptian tortoise,
because it ensures reproduction when males are not encountered (Geffen and Mendelssohn 1991,
p. 576).
Egyptian tortoises in captivity become mature at about 10 years old (Woodland Park Zoo 2014,
unpaginated; Perälä 2005, p. 892; Perälä 2006, p. 60). A rapid juvenile growth phase, until 8 to
10 years, is followed by a continuous, slow adult growth phase (Macale et al. 2009, pp. 141–142;
Mendelssohn 1982, p. 136). Because the growth rate slows down, the growth rings of adults
added are very narrow, making age estimates difficult (Mendelssohn 1982, p. 136). The growth
rate in the wild and in captivity seems to be similar, but the growth rings are more pronounced in
the wild because wild tortoises feed intensely during their active season and do not feed during
aestivation, unlike captive specimens that may feed throughout the year (Mendelssohn 1982, p.
136). Age at maturity in the wild is not known, and may be longer than 10 years.
Life Span and Survival
The life span of the Egyptian tortoise in the wild is mostly unknown. One generation in the wild
is estimated to be about 20 years (Perälä 2006, p. 60; Macale et al. 2009, p. 143). Although, at
Zaranik Protected Area most tortoises did not exceed 15 years old and average age was 8 years
old (Egyptian Environmental Affairs Agency (EEAA) 2009, p. 222). In captivity, at the Giza
Zoo, the life span was more than 20 years (Woodland Park Zoo 2014, p. 1; Buskirk 1985, p. 43);
at the Maryland Zoo the species was reported to live around 50 years (Maryland Zoo 2015, p.1).
Survivorship of turtles suggests that they are characterized by a type III survivorship curve, with
mortality rates inversely related to age (Iverson 1991, p. 386). Survival rates of Testudines are
reasonably well documented for the more easily accessible egg and adult stages, but hatchlings
are rarely encountered in the field (Dunham et al. 1988, in Keller et al. 1998, p. 238). The
probability of detecting an animal increases with age as juveniles and immature tortoises have
lower daily activity than adults (Henry et al. 1998, p. 189).
Survival rate specific to Egyptian tortoises is lacking. However, survivorship data are available
for other closely related Testudo species, such as the Greek tortoise and Hermann’s tortoise.
These species of Testudo are distributed in countries bordering the Mediterranean Sea and in
Asia Minor (Lambert 1982, entire; Henry et al. 1998, entire; Iverson 1991, entire; Diaz-Paniagua
et al. 2001, entire). The rate of survival for a Greek tortoise hatchling has been reported at 39
percent (Keller et al. 1998, p. 239). However, average juvenile survivorship between 0 and 6
years old ranged from 1 to 10 percent, and adult survivorship was 90 percent (Diaz-Paniagua et
al. 2001, p. 713). Similarly, survivorship of juvenile Hermann’s tortoise was 52 percent and
adults was 95 percent (Henry et al. 1998, p. 192). Generally, survivorship during the egg stage
and first year of life is significantly lower than during later stages (Iverson 1991, p. 385; Henry
et al. 1998, p. 192).
Factors that may affect survivorship of the life stages of tortoises are predation, quality and
quantity of food, trampling, and environmental conditions (e.g., temperature and precipitation).
Hatchling survival rates of the Greek tortoise may also decline in arid conditions when there is
15competition for green food with grazing animals (e.g., goats) (Berry 1976, p. 47: Lambert 1982,
pp. 181, 187). The Egyptian tortoise may see similar declines, as it competes with grazing
animals throughout its range.
Individual and Species Needs
In this section we outline the resource needs of individual Egyptian tortoises. The table below
describes the needs of each life stage of the Egyptian tortoise.
Table 1. Egyptian tortoise resource needs by life stage. Juvenile and subadults are combined.
Resource function refers to whether the needs affect breeding (B), feeding (F), sheltering (S), or
dispersal (D).
Life Stage Resources and/or circumstances needed for Resource
individuals to complete each life stage Function
(B, F, S, D)
Preferred air temperature: 15.3 to 35.4ºC (59.5 B, F, D
to 95.7ºF) and preferred surface temperatures:
17 to 45ºC (62.6 to 113ºF)
Adults
Annual plants F
Perennial bushes and shrubs B, F, S
Rodent burrows S
Preferred air temperature: 15.3 to 35.4ºC (59.5 F, D
to 95.7ºF) and preferred surface temperatures:
Juveniles and 17 to 45ºC (62.6 to 113ºF)
sub-adults Annual plants F
Perennial bushes and shrubs F, S
Rodent burrows S
Preferred air temperature: 15.3 to 35.4ºC (59.5 F, D
to 95.7ºF) and preferred surface temperatures:
Hatchlings 17 to 45ºC (62.6 to 113ºF)
Perennial bushes and shrubs F, S
Annual Plants F
Air Temp of at least 30ºC (86ºF) for hatching S
Eggs success
Shrubs S
The Egyptian tortoise is an herbivore (Maryland Zoo 2015, p. 1), although the diet of wild
tortoises is not well understood. Saltwort (Salsola sp.) and sea lavender (Limonium sp.) were
reported to be plants that Egyptian tortoises consumed (Loveridge and Williams 1957, p, 282).
Egyptian tortoises in Zaranik Protected Area in North Sinai, Egypt, were observed eating 13
species of plants, with two perennial shrubs, bindweed (Convolvulus lanatus) and Bushy
bugloss, being the most common (Attum et al. 2007b, p. 401). In Libya, the Egyptian tortoise
16used 20 different plants as food (Schneider and Schneider 2008, p. 147). Additional field
observations indicate that Egyptian tortoises feed primarily on bindweed and the annuals Sleisla
(Eremobium aegyptiacum) and desert campion (Silene villosa) (Attum pers. comm., in Baha El
Din 2006, p. 326).
Because food is likely to be most abundant when Egyptian tortoises are active, wild tortoises
feed intensely during this period and mainly on annual vegetation and leaves of perennial bushes
and shrubs, although most parts of the shrubs may be out of reach (Mendelssohn 1982, p. 134;
Groombridge 1982, p. 134). Precipitation between 100–200 mm (3.94–7.87 in) facilitates the
growth of short-lived annual vegetation, and this relatively high level of precipitation may be the
main factor restricting the species to areas along the Mediterranean coast because coastal areas
receive higher rainfall than areas further inland (Mendelssohn 1982, p. 134).
In captivity, Egyptian tortoises are known to eat grasses, fruits, and vegetables (Maryland Zoo
2015, p. 1); they did not eat sea lavender but used its large leaves as shelter (Connor 1993,
unpaginated). High-protein foods and overfeeding have shown to produce lumpy shells and long-
term renal problems (Highfield and Martin 2014, p. 2). Egyptian tortoises get most of their liquid
from food (Maryland Zoo 2015, p. 1). They consistently refused to drink water when offered in a
dish (Connor 1993, unpaginated). In captivity, when placed outside on dew-laden or water-
sprayed grass they move through the short grass sipping up dew drops as they go (Connor 1993,
unpaginated).
Population and Species Needs
In this section we consider the Egyptian tortoise’s historical and current distribution, its historical
and current population size, and what the species needs for viability.
Historical and Current Distribution
The Egyptian tortoise is restricted to a narrow coastal zone in North Africa and Israel in the
southeast Mediterranean. From west to east, this species occurs in Tripolitania in Libya, east
through northern Egypt, and into the western Negev Desert in Israel (Groombridge 1982, p. 133;
Mendelssohn 1982, p. 133; Baha El Din 1994, p. 1; Baha El Din et al. 2003, p. 648; Schneider
and Schneider 2008, p. 144; Perälä 2001, pp. 570, 573). Historically, the Egyptian tortoise
ranged inland 50 to 120 km (37.3–74.6 mi) from the Mediterranean Sea (Baha El Din 1994, p. 1;
Buskirk 1985, p. 40; Mendelssohn 1982, p. 133; Schneider and Schneider 2008, p. 145; McGrath
2011, unpaginated). The range was possibly much greater in the past, with localities further
inland, such as those individuals reported from Siwa Oasis, Egypt and the Tripolitanian range in
Libya, respectively, which may represent the last remnants of a more extensive distribution that
could have decreased with the natural aridification (i.e., process in which a region becomes
increasingly dry) and expansion of the Saharan Desert (Baha El Din 1994, p. 3; Perälä 2005, p.
892). However, isolated occurrences of the species further inland may represent human
introductions or be extralimital (i.e., outside its natural area) and should be viewed with caution
(Perälä 2005, p. 892).
The current range is highly discontinuous because of the human impact on habitat and natural
factors such as low precipitation (Buskirk 1985, pp. 37–39). The species generally occurs closer
to the coast because it cannot withstand extremely arid conditions further inland (Mendelssohn
1982, p. 133).
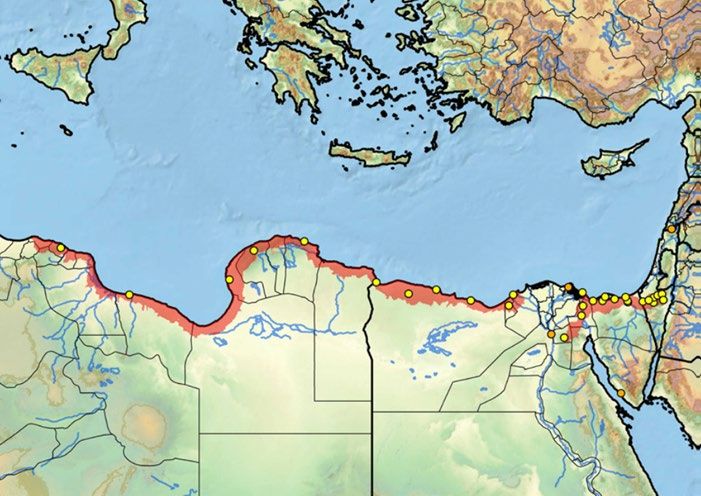
17Figure 5. Distribution of Egyptian tortoise, from Libya through Israel is shaded red. Yellow dots
represent historical and/or current occurrences and the orange dots are probable trade or
extirpated (Turtle Taxonomy Working Group, Rhodin et al. 2017).
Nile River
Israel
Libya Egypt
18Figure 6a. Libyan Range: Black stars represent Egyptian tortoise occurrences and are numbered.
Between the red-dashed line and the Mediterranean Sea is the range of the tortoise in Libya.
However, in Cyrenaica, all confirmed historical records are from immediately east, south, and
west of the Jebel Akhdar (Green Mountains) (see black stars numbered 20–28). Triangles in
northern Cyrenaica represent Greek tortoise. Red dots and squares are Libyan cities (Schneider
and Schneider 2008).
Tripolitania
Cyrenaica
Sirte
19Figure 6b. Egyptian Range: Red dots represent historical distribution of Egyptian tortoise. Solid
red area in upper right, in North Sinai, is current distribution in Egypt. Solid red area in upper
left is part of the Libyan range of the species (Tortoise Trust 2014).
Sinai
Peninsula
North Coast
Nile River
20Figure 6c. Israel Distribution: Shaded area in southwestern Israel is the range of Egyptian
tortoise in western Negev Desert. Stars represent Egyptian tortoise. Stars in black circles are
Greek tortoise. The two stars numbered 6 and 7 are misidentified as Egyptian tortoise and not
part of the species’ range (Bringsoe and Buskirk 1998).
Negev Desert
Based on limited historical data, the extent of occurrence was calculated for the Egyptian tortoise
throughout its range (Perälä 2005, pp. 892–894, Perälä 2006, pp. 57–61). The extent of
occurrence (EOO), as defined by the International Union for Conservation of Nature and Natural
Resources (IUCN), is the area contained within the shortest continuous imaginary boundary that
can be drawn to encompass all the known, inferred, or projected sites of present occurrence of a
taxon, excluding cases of vagrancy. This measure may exclude discontinuities or disjunctions
within the overall distributions (e.g. large areas of obviously unsuitable habitat) and can often be
measured by a minimum convex polygon (the smallest polygon in which no internal angle
exceeds 180 degrees and which contains all the sites of occurrence) (IUCN 2012, pp. 11–12).
Table 2. Historical and current estimates of extent of occurrence (i.e., distribution) for the
Egyptian tortoise; numbers in parentheses include the potential that the species occurred in
21southern Sinai because of possible human introductions (Perälä 2005, p. 894; Perälä 2006, p. 61).
Note that the estimates of area do not include Sirte, Libya where a population is known to occur
(Perälä 2005, p. 894). Surveys for the species in Sirte were conducted in 2005–2007, after the
extent of occurrence was estimated, and the historical or current distribution in Sirte has not been
quantified (Schneider and Schneider 2008). Additionally, the current area of occurrence in
Tripolitania is likely not the same as the historical range due to human impacts. However, it is
the only estimate of area available for the population in Tripolitania.
Population Name Extent of Occurrence (km2)
Historical Current
Libya (Cyrenaica) 50,250 11,100
Libya (Tripolitania) 5,500 5,500
Egypt (North Coast) 67,680 0
Egypt (Sinai) and 21,000 3,625
Israel (40,000)
Total Area 144,430 20,225
(163,430)
Historical and Current Population Size
A population density estimate in Israel was reported at 27 individuals per km2 in the early 1980s,
based on a small study area of 3 km2 (1.16 mi2) (Geffen 1990, in Perälä 2006, p. 61). However,
the average population density in western Israel was estimated at 4 to 5 specimens per km2 in
1982, with about 25 percent of the total population being young and immature individuals
(Perälä 2006, p. 61; Mendelssohn 1982, p. 133). This is extremely low in comparison to small
tortoises elsewhere and may be the result of habitat degradation (i.e., overgrazing) (Mendelssohn
1982, p. 133). No population density assessments based on local field data are available for the
Egyptian tortoise in Egypt or Libya. Nevertheless, densities are thought to be very low (Pieh,
pers. comm., in IUCN 2014, p. 3 and Perälä 2005, p. 894; Mendelssohn 1982, p. 133).
The population in 1982 was estimated to be approximately 10,000 individuals (Mendelssohn
1982, p. 134), and includes the populations of Egyptian tortoise west of the Nile River in Libya
and North Coast, Egypt and east of the Nile River in North Sinai, Egypt and Israel (IUCN 2014,
p. 3). However, the 1982 population estimate was likely an underestimate based on current
populations and statistics on trade (IUCN 2014, p. 3; Perälä 2005, p. 894). Additionally, the
estimate of 10,000 did not take into account tortoises in northwest Libya (Perälä 2005, p. 894).
Because no fieldwork-based data exist for the Egyptian tortoise, the average population density
of 4.5 individuals per km2, based on Mendelssohn’s 1982 study, was assumed to be applicable to
Egyptian tortoises throughout its entire range. This density was used to calculate the population
two to three generations ago (30–50 years ago) and to calculate the current population in 2005
and 2006 (see Table 3, below). Additionally, it was assumed that population sizes are calculated
relative to an estimated area of occupancy, defined as 10 percent of the extent of occurrence
(Perälä 2005, p. 894; Perälä 2006, p. 61; IUCN 2012, entire). The area of occupancy (AOO) is
defined as the area within its 'extent of occurrence' that is occupied by a taxon, excluding cases
22of vagrancy. The measure reflects the fact that a taxon will not usually occur throughout the
entirety of its extent of occurrence (IUCN 2012, pp. 12–13).
Based on this information the total historical population was estimated to be 100,680 individuals
and 118,375 individuals if individuals further inland from southern Sinai are included (see Table
3, below). These tortoises further inland may represent human introductions or be extralimital
and should be viewed with caution (Perälä 2005, p. 892).
Since the late 1950s, or approximately the last three generations, as judged by counts of all
known Egyptian tortoise localities in Libya (including Sirtica (i.e., Sirte)) and the North Coast of
Egypt, the reduction in population size west of the Nile River has been approximately 85 percent
(Perälä 2005, p. 895). Similarly, based on past and present population sizes for Egyptian tortoise
in North Sinai and Israel, the reduction in population size east of the Nile River has been around
93 percent assuming no populations existed in southern Sinai, and a reduction of 95 percent
assuming historical populations in southern Sinai (Perälä 2006, p. 61). Combining all areas west
and east of the Nile River where the Egyptian tortoise is known to occur, the population has
decreased about 90 percent since the 1950s.
The population estimated in 2005 and 2006 was approximately 10,650 individuals (see Table 3,
below). However, because no accurate, fieldwork-based data on population sizes exist, average
population densities have been assumed based on surveys in the early 1980s, and because several
well-known threats and stressors affect the population throughout its range, the population
estimates may be overestimates (Perälä 2005, p. 895). Additionally, taking trade factors into
account, the current population of Egyptian tortoise in the wild might be less than a few thousand
individuals, perhaps only hundreds, but hard evidence is lacking (Perälä 2005, p. 895; McGrath
2011, unpaginated).
Table 3. Estimates of the historical and current populations for the Egyptian tortoise; numbers in
parentheses include the potential that the species occurred in southern Sinai because of possible
human introductions (Perälä 2005, p. 894; Perälä 2006, p. 61). Only one small population
(approximately 100 individuals) exists in North Sinai, Egypt within the Zaranik Protected Area
and on the periphery of this area (Attum 2019, pers. comm.). Note that this table does not include
the population of Egyptian tortoises in Sirte, Libya. This population was not quantified, although
it was reported to be widespread along the coastal area (Schneider and Schneider 2008, p. 148).
Additionally, the current population in Tripolitania is likely not the same as the historical
population due to human impacts.
Population Name Historical Individuals Current Individuals
Libya (Cyrenaica) 22,600 5,000
Libya (Tripolitania) 2,500 2,500
Egypt (North Coast) 30,500 0 (was previously reintroduced
in El Omayed Protected Area)
Egypt North Sinai (South 45,000 (62,775) 3,150, which are mostly in
Sinai) and Israel Israel. The population in North
Sinai is about 100
Total Individuals 100,600 (118,375) 10,650
23You can also read

















































