ProMIS Neurosciences: Selective targeting of pathogenic misfolded proteins, based on a proprietary discovery platform Toronto Stock Exchange (TSX) ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

ProMIS Neurosciences: Selective targeting of pathogenic
misfolded proteins, based on a proprietary discovery platform
Toronto Stock Exchange (TSX) ticker: PMN.TO January 2021
OTCQB ticker: ARFXF.
1
ProMIS Neurosciences Overview
• Growing portfolio of monoclonal antibody candidates focused on neurodegenerative and
other misfolded protein diseases
• ProMIS platform allows for selective targeting of pathogenic species enhancing
efficacy and safety
1. Selective Antibodies - Unique capability and track record generating antibodies
highly selective for the toxic misfolded molecular species of proteins involved in disease
2. Advantages in Gene Therapy Vectors/Intrabodies - ProMIS highly selective
antibodies are ideally suited for vectorization and intracellular targeting of toxic protein
aggregates while sparing the normal forms of the protein. Intrabody function may
improve efficacy
3. Biomarkers for rapid clinical POC - ProMIS will capitalize on emerging fluid-based
biomarkers for rapid and cost efficient early clinical proof of concept
2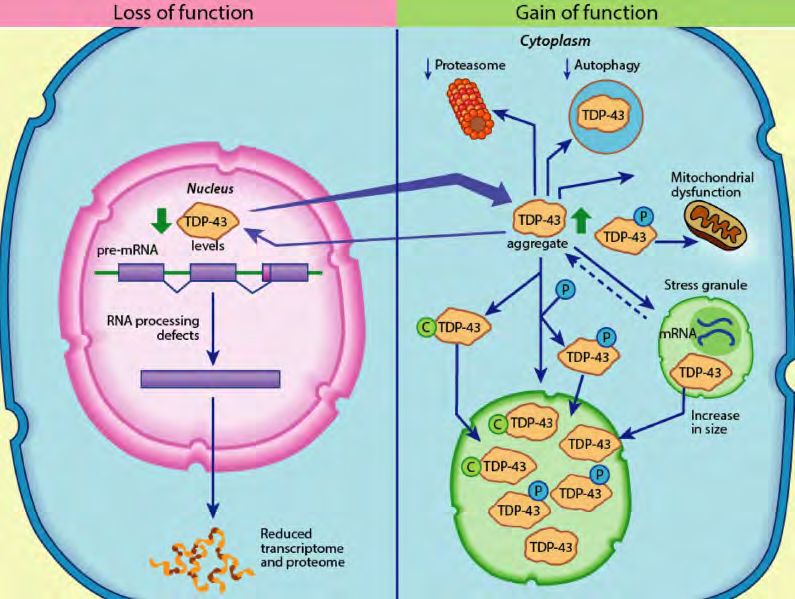
ProMIS goal: Collaboration with gene therapy capable partner,
broad scope – ALS or ALS + MSA
ALS MSA
Alpha
TDP-43 RACK1 SOD1 Ataxin2
Synuclein
Shared program design: Shared program design:
- Clinical program* - Clinical program*
- Vectorization - Vectorization
- Non-clinical proof of concept (Shared with ALS
Also, growing understanding of biologic interaction programs)
* High level draft Clinical Development Plan available – Halloran Group 3
Selectivity is critical to avoid interference with essential cellular function and to
optimize clinical safety
• Many misfolded pathogenic proteins driving neurodegenerative
disorders are primarily intracellular
• Intracellular targeting with intrabodies requires selectivity for the
toxic form in order to preserve function of the normal form of the
protein
Extracellular Antibody Delivery
Neutralization of cell-to-cell propagation
ke
Limited intracellular upta
Cell to cell transmission of
pathogenic aggregates via
exosomes and free protein
Vector delivery of intrabody
Clearance of existing
pathogenic aggregates &
Inhibition of further
propagation 4
The ProMIS platform generates antibodies/intrabodies selective for the
misfolded toxic forms of pathogenic proteins
Identification of epitopes selectively exposed on toxic misfolded form
of the target protein using predictive computational algorithm
Immunization with disease-associated epitope and screening of Normal protein
monoclonal antibodies with desired binding profile and protective activity Misfolding Conformational
epitope
Don’t interfere with the
Neutralize the toxic mis-
normal form, critical for
folded form
brain health Toxic form
Successful track record thus far in generating antibodies selective for
pathogenic forms of amyloid-beta, tau, alpha-synuclein, TDP-43, SOD1
Image adapted from Racaniello V et al, virology.ws, 2016 5
The ProMIS platform has generated tailored antibodies selective for the toxic
forms of proteins involved in neurodegenerative diseases
Antibody therapeutic
BETA-AMYLOID ALPHA-SYNUCLEIN profile optimized to:
Minimize Minimize • Focus the dose on
Bind Toxic Avoid Bind Toxic
Binding Non- Binding Non- the toxic species
Oligomers Monomers Oligomers Toxic Oligomers
Toxic Oligomers • Preserve normal
function
Avoid Minimize
Avoid Bind Soluble • Avoid “soaking up”
Avoid Plaque Physiologic Binding Lewy of therapeutic
Monomers Fibrils Bodies
Tetramers antibody by non-
toxic species
TAU TDP43 100% success rate in
Bind Seed- Bind toxic cytoplasmic generating multiple
Bind Toxic Bind Soluble
Competent aggregates antibody candidates
Oligomers Fibrils for each target protein
Monomers
following immunization
Minimize Binding Avoid Physiologic Avoid Avoid native with 3-6 predicted
Tangles Monomers monomers dimers conformational peptide
scaffolds
6
ProMIS ALS pipeline
Target Conformational Immunization Selective Intrabody
epitope antibodies constructs
TDP-43
SOD1
RACK1 In process
Ataxin-2 In process
7
Expanding beyond ALS to disorders with shared pathogenic targets
FTD HD
LATE
TDP-43 RACK1
Cancers
ALS
Ataxin2 SOD1
Hereditary
Cerebellar
Degen
LATE: Limbic-predominant age-related TDP-43 encephalopathy,
FTD: Frontotemporal dementia, HD: Huntington’s disease
Nonaka T et al. 2013. Cell Reports; Russo A et al. 2017. Hum Molec Gen; Nelson PT et al. 2019. Brain; Culver BP et al. 2012. J Biol Chem;
Ostrowski LA, Hall AC & Mekhail K. 2017. Genes; Li J-J & Xie D. 2015. Oncogene
8
Program highlights
TDP-43 RACK1 Alpha-synuclein
• RACK1 co-aggregates with TDP-43 • mAbs with unparalleled selectivity
• mAbs with selectivity for • Proof of concept studies with RACK1 for misfolded a-syn
misfolded TDP-43 in cell KD support targeting of RACK1: Ø Bind toxic oligomers, small soluble
system and patient samples Ø RACK1 KD “dissolves” TDP-43 fibrils
aggregates Ø Do not bind monomers,
• mAbs inhibit cell to cell physiologic tetramers, inert Lewy
Ø Reverses suppression of protein
transmission body deposits
synthesis by pathogenic TDP-43
• Intrabody constructs bind and • mAbs protect against toxic a-syn:
promote clearance of • Immunizations ongoing to generate Ø Inhibit killing of 1ry dopaminergic
cytoplasmic TDP-43 aggregates mAbs with selectivity for misfolded neurons by toxic oligomers
RACK1 Ø Inhibit uptake and seeding activity
of small soluble fibrils
9
TDP-43
10Misfolded TDP-43 is a driver of pathogenesis in ALS and frontotemporal
lobar degeneration (FTLD)
• Misfolded protein aggregates of TDP-43 are a pathological hallmark of ALS and FTLD1
• The stereotypical distribution of TDP-43 pathology in ALS and FTD shows a spreading pattern consistent with
progressive dissemination from neuron to neuron2,3
• Misfolded aggregates of TDP-43 are toxic to neural cells4,5
• Prion-like cell to cell propagation of TDP-43 aggregates has been demonstrated in cell culture5,6 and animal models7
In vivo spreading of TDP43 pathology from site of injection7
Empty vector sup
TDP43DNLS sup
1Neumann et al, 2006, Science; 2Braak et al, 2013, Nat Rev Neurol; 3Brettschneider et al, 2014, Acta Neuropatholl; 11
4Wang et al, 2016, Nat Med; 5Nonaka et al, 2013, Cell Reports; 6Feiler et al, 2015, J Cell Biol; 7Porta et al, 2018,
Nature CommunSelective targeting of pathogenic TDP-43 for optimal safety and efficacy
TDP-43 is essential to neuronal cell survival1
• TDP-43 is normally present in the nucleus of all cells and
performs an essential role in RNA splicing, transport and stability
• Under stress conditions (e.g. oxidative stress) normal TDP-43
also forms protective stress granules in the cytoplasm
Misfolded TDP-43 gives rise to both loss of function and toxic gain
of function1
• Loss of function: Under disease conditions, misfolding of TDP-43
causes formation of mislocalized cytoplasmic aggregates.
Nuclear depletion leads to defective splicing and transport of
mRNA.
• Toxic gain of function: Cytoplasmic aggregates of misfolded
TDP-43 are toxic and interfere with physiologic stress granule
function. They also induce misfolding of other proteins into
pathogenic aggregates2-4 - “TDP-43 Pathological Interactome”
Targeting of pathogenic TDP-43 requires stringent
selectivity for the misfolded form of the protein to avoid
Figure from de Boer et al1 safety concerns
1de Boer, EMJ et al, 2020, J Neurol Neurosurg Psychiatry; 2Pokrishevsky et al, 2016, Scientific Reports; 3Chou et al, 2018,
Nat Neurosci; 4Endo et al, 2018, Biological Psych 12ProMIS mAbs to misfolded TDP-43 react with mislocalized, aggregated
DNLS-TDP-43 but not nuclear wild type TDP-43
Misfolded DNLS-
Nuclei TDP-43 (HA Tag) ProMIS mAb Merge
• HEK-293 cells
transfected with
DNLS TDP-43 lacking
a functional nuclear
localization signal
• Cells stained for HA
tag (red) of
overexpressed DNLS
TDP-43 or with
rabbit mAb to
misfolded TDP-43
epitope at 2 µg/ml
(green).
• Nuclei stained with
DAPI (blue)
• Images analyzed by
confocal microscopy
(Z-stacks)
13Misfolded TDP-43 is recognized by ProMIS antibody in brain tissue from FTLD
patients with pathology subtypes A,B,C,E and in spinal cord from ALS patients
FTD brain, TDP-43 pathology type A FTD brain - TDP-43 pathology type B FTD brain - TDP-43 pathology type C
WM GM WM GM WM GM
FTD brain - TDP-43 pathology type E ALS patient 1 – Spinal cord ALS patient 2 – Spinal cord
WM GM
Staining with ProMIS rabbit pAb to misfolded TDP-43
WM – White matter Performed by Dept. of Pathology, Amsterdam Neuroscience, VU University Medical 14
GM = Grey matter Center, Amsterdam, The NetherlandsProMIS mAbs to misfolded TDP-43 do not recognize TDP-43 in
protective physiologic stress granules
Staining for stress
granule marker G3BP1 Staining with
ProMIS mAb Staining with pan-TDP-43
Non-stressed
HEK293 cells
Staining with a pan-reactive TDP-43
antibody confirms that normal TDP-43
is present in physiologic stress granules
-> not recognized by misfolded-specific
ProMIS mAb
HEK293 cells
stressed by
exposure to • HEK293 cells stressed by 60min exposure
to 1mM sodium arsenite for 60 min
sodium arsenite
• Cells stained with ProMIS rabbit mAb at
10 µg/ml (green) or with antibody to
G3BP1 (magenta)
• Images analyzed by confocal microscopy15Subnanomolar affinity of ProMIS monoclonal antibodies for their
cognate epitope
Mouse 262 kON kOFF KD
Mouse 263 mAb (M-1s-1) (s-1) (M)
PMN262 2.83E+06 1.76E-03 6.22E-10
PMN263 2.77E+06 2.00E-03 7.22E-10
PMN264 4.03E+05 2.50E-05 6.20E-11
PMN266 4.70E+05 4.32E-05 9.19E-11
PMN267 5.99E+05 9.04E-06 1.51E-11
Rabbit 264 Rabbit 266 Rabbit 267
16
16Antibody-Based Targeting of Pathogenic TDP-43 – Extracellular delivery
Extracellular Antibody Delivery
Neutralization of cell-to-cell propagation
ke
Limited intracellular upta
Cell to cell transmission of
pathogenic species via
exosomes and free protein1-4
Vector delivery of intrabody
Clearance of existing
pathogenic aggregates5-6 1
Feiler et al, JCB 2015: 2Porta et al, Nature Commun 2018; 3Munch et al, PNAS 2011; 4Grad et al,
Inhibition of pathological PNAS 2011; 5Tamaki et al, Scientific Reports 2018; 6Ganashyam et al, Neurobiol of Dis 2018; 7
interactome and further Pokrishevsky et al, PLoS-One 2012; 8Pokrishevsky et al, Sci Rep 2016; 9Chou et al Nat Neurosci
2018
propagation7-9 17Functional Assay: ProMIS mAbs inhibit cell-to-cell transmission of
misfolding DNLS-TDP43
HA-DNLS-TDP-43 transmission relative
dNLS-TDP-HA
to mIgG1 control
1.2
1
0.8
0.6
0.4
0.2
0
mIgG1 Ab1
3F11 Ab2
2F7 3Ab3
E8 Ab4
9C5 Ab5
5B7
48 hr ProMIS mAbs inhibit transmission of misfolding TDP-43
from the conditioned medium of donor HEK293 cells
transfected with DNLS-TDP-43 to naïve recipient cells
WB
18Antibody-Based Targeting of Pathogenic TDP-43 – Intracellular
vectorized delivery
Extracellular Antibody Delivery
Neutralization of cell-to-cell propagation
ke
Limited intracellular upta
Cell to cell transmission of
pathogenic species via
exosomes and free protein1-4
Vector delivery of intrabody
Clearance of existing
pathogenic aggregates5-6
Inhibition of pathological
1
Feiler et al, JCB 2015: 2Porta et al, Nature Commun 2018; 3Munch et al, PNAS 2011; 4Grad et al,
PNAS 2011; 5Tamaki et al, Scientific Reports 2018; 6Ganashyam et al, Neurobiol of Dis 2018; 7
interactome and further Pokrishevsky et al, PLoS-One 2012; 8Pokrishevsky et al, Sci Rep 2016; 9Chou et al Nat Neurosci
2018
propagation7-9 19ProMIS intrabodies only interact with cytoplasmic DNLS-TDP43
aggregates and not normal nuclear TDP43
HEK293 cells transfected with DNLS-TDP43 and single chain ProMIS intrabody
Intrabody DNLS-TDP43 Merged images
• Expression of ProMIS TDP43 intrabody is not toxic to cells
• Intrabody co-localizes with mislocalized, aggregated cytoplasmic DNLS-TDP43
• Intrabody does not interact with endogenous normal TDP43 in the nucleus
20TDP-43 intrabody promotes clearance of misfolded TDP-43 aggregates
DNLS –TDP-43+ EV
Misfolded TDP43-selective intrabody with lysosomal targeting signal
DNLS-TDP43 +
promotes degradation of TDP-43 aggregates without cellular toxicity
intrabody
HA intensity relative to control “dNLS+EV”
ê57.6%
HA tag – Aggregated 1.0
mutant TDP-43
Actin
0.5
0.0
Intrabody 267 Flag tag DNLS + EV DNLS + intrabody 267
EV
-5
IB
+
ak
LS
O
dN
+
LS
dN
EV = empty vector 21ProMIS selectivity for the toxic form of TDP-43 is critical for gene therapy
vectorization of intrabodies in order to preserve normal cell function
• ProMIS antibodies are highly selective for misfolded TDP-43
ü Epitope binding affinity in the sub-nanomolar range
ü Reactive and specific for aberrant cytoplasmic TDP-43 aggregates with no reactivity with
wild-type nuclear TDP-43 → preserves normal, essential TDP-43 function
ü No binding to physiological stress granules → preserves stress-protective function
ü Reactive with native pathological TDP-43 from human brain and spinal cord samples
• Potential for rapid clinical proof of concept in ALS using fluid-based biomarkers (e.g.
NfL decrease)
• Gene therapy in ALS could enable a rapid route to approval
22TDP-43 intrabody program - Next steps
• Vectorization of lead intrabody candidate
• Optimize genetic construct for lead intrabody PMN267 (potentially also PMN265 back-up)
• Insert into AAV9 gene therapy vector
• Select lead vector construct based on in vitro activity (engagement and clearance of
misfolded TDP-43) – Cell system and iPSC-derived motor neurons from ALS patients
• In vivo testing
• Confirm CNS access and expression of intrabody in motor neurons
• Confirm target engagement
• Disease models largely non-predictive of clinical efficacy
• Conduct IND-enabling studies
23RACK1
24Targeting RACK1 in ALS
• Receptor of activated protein C kinase 1 (RACK1) is a core ribosomal protein of the eukaryotic small (40S)
ribosomal subunit.1
• It is a scaffold protein that interacts with several other proteins thereby regulating a variety of signaling
pathways critical for cell proliferation, transcription and protein synthesis -> essential for neuronal function.1
• In ALS, RACK1 and TDP-43 co-localize in cytoplasmic aggregates in spinal cord motor neurons.2
• In a cell system, DNLS-TDP-43 has been reported to suppress global protein synthesis by co-aggregating
with RACK1 on polyribosomes.2 Translational repression was mitigated with a RACK1 mutant that reduced
interaction with DNLS-TDP-43.2
• These results from the literature and ProMIS’ proof of concept data using RACK1 knock-down support
intracellular targeting of RACK1 as a therapeutic approach
• Knock-down of RACK1 establishes proof of concept but is non-selective -> Intrabodies
selective for pathogenic, aggregated RACK1 will be required to preserve normal cell function
• Current status: Immunizations underway with predicted epitope of misfolded RACK1
1Adams et al, 2011, Cell Commun Signal; 2Russo et al, 2017, Hum Molec Gen 25Proof of concept - Knock-down of RACK1 “rescues” misfolded DNLS-TDP-43
RACK1 DNLS-TDP-43 (HA tag) Merge (DAPI nuclei)
RACK1
-siRNA • RACK1 in the cytoplasm co-
Normal aggregates with misfolded,
levels cytoplasmic DNLS-TDP-43
• RACK1 knock-down reduces
cytoplasmic aggregates of
DNLS-TDP-43
Nuclear
RACK1
+siRNA
Knock-down
Diffuse
26Proof of concept - RACK1 knock-down reverses suppression of
protein synthesis by DNLS-TDP-43
DNLS-TDP-43 (HA tag) Puromycin-tagged mRNA Merge
• Protein synthesis
(puromycin-tagged mRNA)
is suppressed in cells
transfected with DNLS-TDP-
43
• RACK1 knock-down reduces
cytoplasmic aggregates of
DNLS-TDP-43 and restores
protein synthesis
27Alpha-Synuclein
28Alpha-synuclein is the major driver of Parkinson’s disease and other synucleinopathies
• Synucleinopathies: Parkinson’s disease (PD) is a progressive neurodegenerative disorder characterized by loss of
dopaminergic neurons located in the midbrain and the presence of intraneuronal inclusions (Lewy bodies/Lewy
neurites) consisting mainly of aggregates of a-synuclein (a-syn). Accumulation of insoluble a-syn fibrils in the
brain is also observed in dementia with Lewy bodies (LBD) and multiple system atrophy (MSA).
• Genetic mutations (A53T, A30P, E46K, H50Q, G51D) and duplications/triplications of the a-syn gene cause
inherited familial PD1
• Polymorphisms in the a-syn gene and its promoter are associated with an increased risk of sporadic PD1
• Mice injected with aggregated a-syn from diseased patient brains develop synucleinopathy-like pathology and
symptoms2
• Aggregated a-syn propagates from cell to cell in a prion-like manner in cell culture3 and
in animals4,5 mimicking disease progression in patients
Braak, H et al, Neurobiol of Aging, 2003
1
Reviewed in Vaikath, NN et al, J Neurochem 2019; 2Reviewed in Karpowicz Jr, RJ et al, Lab Invest 2019; 3Choi et al, Cell
Reports 2018; 4Peelaerts et al, Nature, 2015; 5Froula JM et al, JBC 2019 23Therapeutic imperative: selectively target only the toxic a-synuclein aggregates
• Alpha-synuclein exists in different forms including normal,
physiologically important conformations and toxic forms:
• Monomers play an important role in the regulation of synaptic Optimum
vesicle trafficking and release as well as neuronal survival1
Therapeutic Profile
• Physiological a-syn tetramers inhibit aggregation and must be
preserved for healthy a-syn homeostasis2-4
• Lewy bodies consisting of insoluble a-syn are a marker of disease
but unlikely to have direct neurotoxicity5
• Recent evidence indicates that a-syn toxicity resides primarily Minimize
with soluble oligomers6,7 Avoid Target Toxic
targeting non-
Monomer Oligomers toxic oligomers
• Progression of disease is likely mediated by oligomers and small
soluble fibrils of a-syn that have been shown to propagate from
Avoid Minimize
cell to cell in a prion-like manner in vitro8 and in vivo9 Target Soluble
physiologic targeting Lewy
Fibrils Bodies
• Maximal efficacy and safety is expected to require selectivity tetramer
for the toxic forms of a-syn, oligomers and/or small soluble
fibrils, while avoiding physiologic forms of a-syn
1Lashuel HA et al, Nat Rev Neurosci, 2013; 2Nuber et al, Neuron, 2018; 3Dettmer et al, PNAS, 2015;
4
Foulds et al, Scientific Reports, 2013; 5Vaikath NN et al, J Neurochem 2019; 6Fusco et al, Science, 30
2017; Desplats P et al, PNAS, 2009; 8Choi et al, Cell Reports, 2018; 9Peelaerts et al, Nature, 2015Biological relevance of predicted epitopes: The misfolded portion of a-syn
(conformational epitope) is sufficient to replicate the seeding activity of pre-
formed fibrils
Thioflavin T seeding assay
1.5×105
ThT fluorescence intensity
Monomer + cyclic peptide
1.0×105 Conformational epitope induces seeding
5.0×104
Linear sequence (no conformation)
does not have seeding activity
0.0 Monomer +linear peptide
Monomer only
0 50 100 150 200 250 300
Time (hrs)
31ProMIS’ technology platform has created antibodies with greater selectivity
than other a-synuclein-directed antibodies
PMN Prothena/ BioArctic/ Neurimmune/
Target Properties Antibodies Roche ABBVIE Biogen
No binding to monomers ✔ X +/- X
No binding to physiological ✔ X +/- X
tetramers
Binding to oligomers/small ✔ ✔ ✔ ✔
soluble fibrils
Binding to native toxic a-syn in ✔ ✔ ✔ ✔
PD/LBD brain extract
Little or no binding to insoluble ✔ X X X
fibrils (Lewy bodies)
32ProMIS antibodies show a high degree of selectivity for pathogenic species of a-syn
(toxic oligomers and small soluble fibrils)
12G1 - Monomers 4000
12G1 - Toxic oligomers 1500 12G1 - Sonicated fibrils
1500
1250
Response Units
Response Units
Response Units
3000
1000 1000
750
2000
200
500
LLOQ ~1.4 ug/ml 1000 LLOQ ~ 40 pg/ml LLOQ ~8 pg/ml
100
0 0 0
0 500 1000 1500 0 5000 10000 15000 0 500 1000 1500
Concentration (nanogram/ml) Concentration (picogram/ml) Concentration (picogram/ml)
• Millipore SMC platform
• Representative antibody from
Fold selectivity oligomers vs Fold selectivity fibrils vs
several tested monomers: 35,000X monomers: 175,000X
• Microparticles coated with PMN432
antibody for capture
• Detection with pan-a-synuclein
antibody
33Misfolded tetramers of a-synuclein are neurotoxic
• In vitro evidence suggests that misfolded tetrameric species carry toxicity, unlike physiological tetramers (Nuber
S et al, Neuron 2018): Two validated preparations of synthetic oligomers of alpha-synuclein toxic to primary rat
dopaminergic neurons consist primarily of tetramers (Neuron Experts, Synaging)
Toxicity of a-synuclein oligomer preps
SEC of toxic oligomer preps shows
predominance of tetramers
100
NE toxic oligomers SA toxic oligomers
80 Blue: MW markers Tetramers
Blue: MW markers Tetramers
*
% Viability
56kDa
60
* Red: Oligomers 56kDa Green: Oligomers
40 17kDa 17kDa
20
44kDa 44kDa
0
Control NE a-syn SA a-syn
• NE = Neuron Experts a-syn oligomer prep
• SA = SynAging a-syn oligomer prep
• Both at 500 nM
• * p < 0.05 vs control
34Toxic a-syn oligomers, but not physiologic tetramers, have seeding activity comparable
to that of pre-formed fibrils
Monomers only hPFF + Monomers
1.5×107
Pre-formed
ThT Fluorescence Intensity (RFU)
fibrils (PFF)
1.0×107
5.0×106
0.0
0
24
48
72
96
120
144
168
192
216
240
Time (hrs)
Monomer only Toxic Oligomers + Monomer Monomers only Physiol Tetramers + Monomers
1.5×107 1.5×107
Toxic oligomers Physiologic tetramers
ThT Fluorescence Intensity (RFU)
ThT Fluorescence Intensity (RFU)
1.0×107 1.0×107
5.0×106 5.0×106
0.0 0.0
0
24
48
72
96
120
144
168
192
216
240
35
0
24
48
72
96
120
144
168
192
216
240
Time (hrs) Time (hrs)Only ProMIS antibodies bind toxic oligomers but not physiologic tetramers (SPR)
Physiologic tetramers* Misfolded Toxic oligomers
Target epitope of
Target epitope of PMN
PMN antibodies is Structure?
antibodies is exposed
buried in the
and available for binding
hydrophobic core
*Tetramer structure: Xu et al, Chem Commun 2018 Physiological tetramers
Toxic oligomers
30 1000
Selective binding of PMN 800
Comparator antibodies show
antibodies to toxic oligomers vs comparable binding to toxic oligomers
Binding Response (RU)
600
and physiological tetramers
Binding Response (RU)
physiological tetramers
400
20
200
60
10 40
20
0 0
mIgG1 PMN432
PMN1 PMN451
PMN2 PMN431
PMN3 Biogen BioArctic Prothena
Physiological tetramers isolated by SEC following Toxic oligomers prepared by SynAging,
spontaneous monomer aggregation neurotoxicity confirmed in in vitro toxicity assay 36ProMIS antibodies protect dopaminergic neurons
against a-synuclein oligomer toxicity in vitro CONTROL
Normal neurons
in bright red
Survival (dopaminergic neuron % of ocntrol)
120
** **
** ** **
100
* *
80
60
a-SYN OLIGOMERS
Neurons killed by
40 toxic oligomers
431 432 442 421 452 412
ol
o
)
)
)
)
)
)
F
:2
:2
:2
:2
:2
:2
ig
N
tr
(1
(1
(1
(1
(1
(1
D
ol
on
B
n
n
n
n
n
n
n
C
sy
sy
sy
sy
sy
sy
sy
a-
a-
a-
a-
a-
a-
a-
+
+
+
+
+
+
8
1
12
12
6
5
9D
G
7F
D
B
8B
10
12
12
• Multiple antibodies provide neuroprotection in the same range as the brain-derived PMN ANTIBODY +
neurotrophic factor (BDNF) positive control a-SYN OLIGOMERS
• As expected, antibodies alone had no effect on viability (not shown) Neuronal death
blocked by PMN
antibody
*pProMIS antibodies inhibit a-syn propagation: Reduced PFF uptake and formation of
aggregates
Human soluble a-syn fibrils +/- PMN antibody
Staining for human a-syn aggregates
14 days
Rat neurons Rat neurons
ProMIS antibodies significantly decrease
formation of a-syn aggregates
Ratio a-syn area/neuron number (in % of Control)
60000
20000
*
40000
*
10000
20000
CONTROL a-SYN SOLUBLE FIBRILS a-SYN SOLUBLE FIBRILS +
0 0
Control
Control PFF
PFF PFF + 411
PFF + 2E9 (0.25uM)
Control
Control PFF
PFF PFFPFF + 442
+ 12B12 (0.05uM) PMN ANTIBODY 411
Human a-syn aggregates stained red
*pProMIS antibodies inhibit a-syn propagation: decreased recruitment of
endogenous rat a-syn into pathogenic phosphorylated aggregates
Human PFF +/- PMN antibody Staining for rat phosphorylated a-syn aggregates
14 days
Rat neurons Rat neurons
ProMIS antibodies significantly decrease recruitment
into a-syn phosphorylated aggregates
Survival (dopaminergic neuron % of control)
250
200
150 * *
100
50
CONTROL a-SYN SOLUBLE FIBRILS a-SYN SOLUBLE FIBRILS +
0 PMN ANTIBODY 411
Control
Control PFF
PFF PFF PFF
+ 2E9+ (0.05uM)
411PFF + PFF + 442
12B12 (0.25uM)
Endogenous rat phospho-aggregates stained yellow (denoted by arrows)
Human a-syn aggregates stained red
*pProMIS antibodies show strong binding to native toxic a-syn in human
soluble MSA brain extract (SPR)
80
• All PMN antibodies show a
60
greater binding response
than the pan-a-syn control
Binding Response (RU)
(4D6) and the BioArctic and
Biogen antibodies
40 • The Prothena antibody
shows the highest binding
response in agreement with
its pan-reactivity and non-
20 selective binding to all forms
of a-syn
0
411 432 451 442 441 431 412 BioArctic Biogen Prothena 4D6 mIgG1
40ProMIS has generated multiple anti-a-syn antibodies with potential
competitive advantages over therapies currently in development
• Scientific literature suggests a complex binding profile for an optimum antibody
• Bind toxic oligomers and small soluble fibrils involved in propagation
• Avoid physiologic monomer and tetramer, reduce adverse event odds, enable vectorization
• Minimize binding to inert soluble aggregates and Lewy Bodies – maximize therapeutic dose
available
• ProMIS identified unique epitopes presented on target molecular species and generated
multiple antibody candidates with the target binding profile and the ability to inhibit both a-
syn neurotoxicity and propagation in vitro
• Multiple System Atrophy as an early clinical proof of concept indication – patients are rapidly
progressive with elevated biomarkers (serum NfL) at baseline
An enhanced, highly selective binding profile is expected to lead to better clinical
outcomes, and enable safe and effective vectorization
41Experienced leadership team
Name Title Years of Experience Prior Experience
§ Former SVP at Genzyme, with senior roles integrating commercialization, drug
development, and deal making
Gene Williams Executive Chairman 25+ § Recently the CEO of Dart Therapeutics, an Orphan Disease drug development
company
§ Founder and director of Adheris, which became the largest company in the
patient adherence/compliance area
§ Held positions as SVP of Strategic Product Development at SmithKline
Beecham (now GSK)
Elliot Goldstein CEO 25+ § Chief Operating Officer and Chief Medical Officer of Maxygen
§ Chief Operating Officer at DART Therapeutics
§ Holds the Canada Research Chair in Neurodegeneration and
Protein Misfolding Diseases,
Neil Cashman Chief Science Officer 25+ § Serves as the Director of the University of British Columbia ALS
Centre,
§ Awarded the Jonas Salk Prize for biomedical research in 2000
§ Professor at UBC in the Department of Physics and Astronomy since 2001
§ Appointed as the Canada Research Chair in Theoretical Molecular Biophysics
Steven Plotkin Chief Physics Officer 20 § Associate member of the Genome Sciences and Technology Program, the
Bioinformatics Program, and the Institute for Applied Mathematics at the
University of British Columbia
§ Founding Managing Director of Danforth Advisors
§ Served as the Chief financial officer of Homology, Inc, GenePeeks,
Dan Geffken CFO 25+ Inc., Transkaryotic Therapies, Inc., Cidara, Inc., Apellis, Inc. and
Stealth BioTherapeutics, Inc.
§ Former VP of Research at Genzyme
Chief Development 25+
§ Associate Immunopathologist at SmithKline Beecham where she
Johanne Kaplan Officer
established an Immunotoxicology program
§ Her work has resulted in over 60 scientific publications and multiple patents
59Scientific Advisory Board
Name Years of Experience Prior Experience Affiliations
§ Medical Director & Principal Investigator of Toronto Memory Program
Sharon Cohen, MD 20+ § FRCPC in neurology from Royal College of Physicians of Canada and a fellowship in
Behavioural Neurology from the University of Toronto
Director of the Center for Translational Research in Neurodegenerative Disease at
Todd Golde, MD, PhD 20+ §
the University of Florida
§ Dean for Neurosciences Initiatives, Distinguished Professor of Neurosciences, and
Bill Mobley, MD, PhD 25+ Florence Riford Chair for Alzheimer Disease at the University of California, San
Diego
§ Former VP, Global Clinical Leader for Parkinson’s disease, and Clinical Head of the
James Kupiec, MD 20+ Neuroscience Research Unit for Pfizer, Inc.
§ Clinical focus on development of therapies for neurodegenerative disorders
§ Previous Henry P & Georgette Goldschmidt Professor & Chairman, Department of
C. Warren Olanow, MD 25+ Neurology at Mount Sinai School of Medicine, presently Professor Emeritus
Department of Neurology & Department of Neuroscience, CEO of CLINTREX
§ Professor Institute of Psychiatry, Psychology and Neuroscience at King’s College
Andre Strydom, MD, PhD 25+ London
§ Honorary Consultant psychiatrist, South London and the Maudsley NHS Foundation
Trust
§ Professor of Neurology at Harvard University, Vice Chair of Neurology, Director of
Rudy Tanzi, PhD 20+ Genetics & Aging Research Unit, Co-Director McCance Center for Brain Health at
Mass General Hospital
§ Associate Professor of Neurology and Research Professor at Emory University
Lary Walker, PhD 20+ Yerkes National Primate Research Center
60Thank You
Please feel free to contact us with any additional questions.
Eugene Williams, Executive Chairman Elliot Goldstein, MD, CEO
eugene.williams@promisneurosciences.com elliot.goldstein@promisneurosciences.com
+1 (617) 460-0978 +1 (415) 341-5783
Website: www.promisneurosciences.com
Twitter: https://twitter.com/ProMISinc
LinkedIn:
https://www.linkedin.com/company/promis-
neurosciences
44You can also read

















































