Preliminary Investigation of Schmalhausen's Law in a Directly Transmitted Pathogen Outbreak System
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
viruses
Communication
Preliminary Investigation of Schmalhausen’s Law in a Directly
Transmitted Pathogen Outbreak System
Antoine Filion * , Mekala Sundaram and Patrick R. Stephens
Department of Integrative Biology, 501 Life Sciences West, Oklahoma State University, Stillwater, OK 74078, USA
* Correspondence: afilion90@gmail.com
Abstract: The past few decades have been marked by drastic modifications to the landscape by
anthropogenic processes, leading to increased variability in the environment. For populations that
thrive at their distributional boundaries, these changes can affect them drastically, as Schmalhausen’s
law predicts that their dynamics are more likely to be susceptible to environmental variation. Recently,
this evolutionary theory has been put to the test in vector-borne disease emergences systems, and has
been demonstrated effective in predicting emergence patterns. However, it has yet to be tested in
a directly transmitted pathogen. Here, we provide a preliminary test of Schmalhausen’s law using
data on Marburg virus outbreaks originating from spillover events. By combining the two important
aspects of Schmalhausen’s law, namely climatic anomalies and distance to species distributional
edges, we show that Marburgvirus outbreaks may support an aspect of this evolutionary theory, with
distance to species distributional edge having a weak influence on outbreak size. However, we failed
to demonstrate any effect of climatic anomalies on Marburgvirus outbreaks, arguably related to the
lack of importance of these variables in directly transmitted pathogen outbreaks. With increasing
zoonotic spillover events occurring from wild species, we highlight the importance of considering
ecological variability to better predict emergence patterns.
Keywords: disease ecology; Schmalhausen’s law; ecological boundaries; spillover; filovirus; disease
emergence
Citation: Filion, A.; Sundaram, M.;
Stephens, P.R. Preliminary
1. Introduction
Investigation of Schmalhausen’s Law
in a Directly Transmitted Pathogen Anthropogenic changes have greatly altered the delicate balance that dictates species
Outbreak System. Viruses 2023, 15, distributions worldwide. By either introducing new species, encroaching on natural habi-
310. https://doi.org/10.3390/ tats, or changing the landscape, human activities have completely modified expected
v15020310 species distributions and interactions in many ecosystems [1]. For species that already live
in physiologically stressful conditions in areas of greater variability, for instance at the edge
Academic Editor: James Strong
of their distributional range, these modifications can affect them drastically. For instance,
Received: 27 November 2022 many mechanistic processes, such as predation, competition, and others, are often height-
Revised: 16 January 2023 ened in ecotones [2,3]. Following the same rationale, greater variability in the environment
Accepted: 20 January 2023 has been demonstrated to increase the chances of disease spillover and subsequent zoonotic
Published: 22 January 2023 outbreaks [4]. For example, mammalian hosts of Lyme disease (Borrelia burgdorferi) are
more abundant in ecotones and contribute to larger outbreaks in these areas [5]. With the
potential for more zoonotic outbreaks, gaining a deeper understanding of the influence
of environmental variability in spillover events would prove as a valuable interest for
Copyright: © 2023 by the authors.
conservation management from a One Health perspective.
Licensee MDPI, Basel, Switzerland.
This article is an open access article
Schmalhausen’s law holds that species pushed to the edges of their ecophysiological
distributed under the terms and
tolerances in any dimension will express unusual phenotypes, as genotypic expression
conditions of the Creative Commons
occurs in conditions that have been rare or absent during species’ evolutionary history [6].
Attribution (CC BY) license (https:// Applied to diseases, Schmalhausen’s law predicts that temporal or spatial boundaries
creativecommons.org/licenses/by/ of ecological systems are close to the edge of species physiological tolerances, and there-
4.0/). fore susceptible to minor environmental changes potentially leading to unusual outbreak
Viruses 2023, 15, 310. https://doi.org/10.3390/v15020310 https://www.mdpi.com/journal/viruses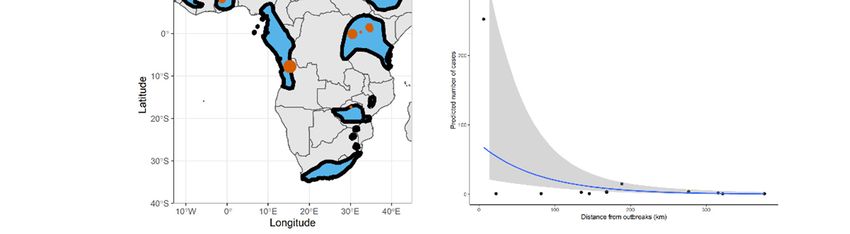
Viruses 2023, 15, 310 2 of 7
dynamics [7]. The key advantage of working within this evolutionary theory is that it
integrates both spatial and temporal variability into a unified conceptual framework to
apply toward disease outbreaks. For example, [8] showed that increased variability in
both temperature and rainfall was positively related to mosquito emergencies, contributing
to outbreaks of the Western Nile Virus in Texas. So far, Schmalhausen’s law has been
applied to vector-borne disease systems, such as avian malaria [7] and leishmaniasis [9],
presumably because conditions that are close to the physiological limits of either the vectors
or the pathogen itself allow for unusual outbreak dynamics with minor environmental
changes. There is some evidence that directly transmitted parasite prevalence is higher at
their host’s range edge [10]. However, whether Schmalhausen’s law may affect outbreak
characteristics remains to be tested in a directly transmitted pathogen.
There are multiple mechanistic ways in which individuals or populations of a host
species being pushed to the edge of their ecophysiological tolerances will potentially result
in an increased frequency of spillover. For instance, immunity to any given pathogen
can be a function of environmental tolerance, in which a harsher environment limits an
individual’s resource allocation to the immune system, in turn making them more prone
to infection [11,12]. Local landscape features are also known to change immune functions
between individuals of the same species [13]. As such, increased variation in immunity at
species range edges might lead to increase spillover risk toward other populations. Another
mechanistic path that would result in an increased spillover rate is an increased rate of
contacts at the human/wildlife interface, particularly in cases when species range edges
occur near transitions zones between biomes such as forest edges, resulting in spillover
event due to an increase in contact with natural hosts shedding their pathogens [14].
Here we present a preliminary test of Schmalhausen’s law in a directly transmitted
zoonotic pathogen, Marburg virus (Marburgvirus marburgvirus), a filovirus related to the
Ebola genera of viruses Though outbreaks of this pathogen are relatively rare, they also
generally show extremely high mortality [15]. For example, the largest outbreak to date
showed a case fatality rate of 88% among over 100 human cases [16–18], suggesting that
Marburg virus could post a significant threat were it to ever spread widely. Thus, there
is an urgent need to better understand the mechanisms behind this pathogen’s outbreaks.
Marbug virus outbreaks are an ideal system for preliminary analyses of Schmalhausen’s law
in a directly transmitted pathogen since it is a relatively simple system, with one reservoir
being associated with outbreak events, the Egyptian fruit bat (Rousettus aegyptiacus) [19,20].
This allowed us to test Schmalhausen’s law by focusing on geographic range effects in a sin-
gle well-characterized wild host species, simplifying our analyses considerably. Moreover,
it has been demonstrated that roosting Egyptian fruit bats release the Marburg virus in
seasonal pulses, suggesting a temporal influence in spillover events [21]. Recent advances
in remote sampling [22] have also made it possible to obtain month-to-month climatic
anomalies for the locality of every documented Marburg virus outbreak. As such, we feel
confident that we are able to incorporate the spatio-temporal conceptual framework of
Schmalhausen’s law into a disease dynamic that is extremely important from a One Health
perspective.
Arguably, as opposed to vector-borne pathogens, small environmental changes would
have far less impact on a non-vector host species: endothermic mammalian hosts in
particular are likely far less influenced by the types of minor climatic events that prove
impactful to vectors. On the other hand, species at the edge of their distributional range
will still be affected by multiple environmental stressors. If Schmalhausen’s law applies
to directly transmitted pathogens, we predict that minor climatic anomalies, represented
by events outside the normal standard deviation for an area, will affect outbreak size. In
addition, we predict that outbreaks at the edge of the Egyptian fruit bat spatial range will
be more severe than those in the center of this species’ distribution. If we can detect any
evidence of the influence of Schmalhausen’s law in this relatively rare pathogen maintained
by a single endothermic reservoir, it is likely that this evolutionary law would apply to
other directly transmitted pathogens as well.allowing us to work with 13 outbreaks ranging from 1975 to 2021, inclusively.
Viruses 2023, 15, 310 3 of 7
2. Materials and Methods
2.1. Data Collection
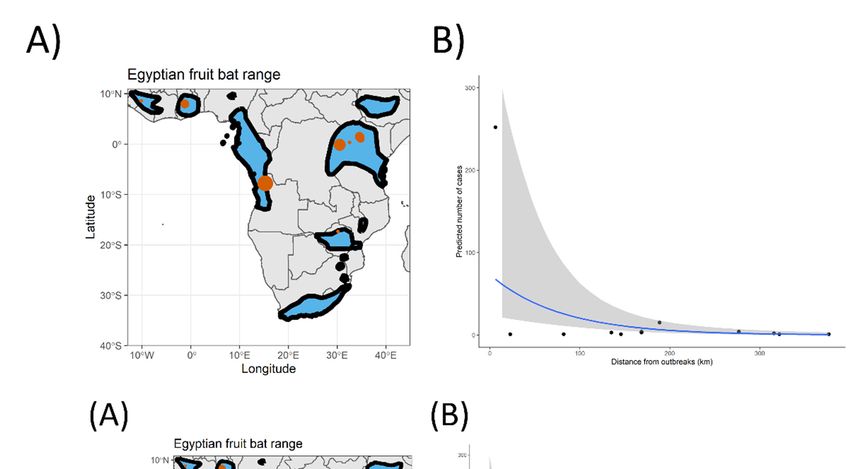
All outbreaks included in our study (Figure 1A) were compiled from [23], CDC
outbreak data [20], and ProMED [24]. We then pruned the dataset to include only confirmed
Marburg virus outbreaks that originated from a wild source (i.e., we excluded all data
points from accidental laboratory spillover events). For each outbreak, we compiled the
date of the outbreak, its geographical original location, and the number of human cases,
allowing us to work with 13 outbreaks ranging from 1975 to 2021, inclusively.
Figure Figure 1. (A) Distributional
1. (A) Distributional range
range of of the Egyptian
the Egyptian fruit batfruit bat (Rousettus
(Rousettus aegyptiacus).
aegyptiacus). Black borders
Black borders
represent
represent the edgetheofedge of this species’
this species’ distribution
distribution in eachin each territory.
territory. Red
Red dots dots represent
represent Marburg Marburg
virus virus
outbreaks in human populations, independent of the year of the outbreak. Note that multiple out-
outbreaks in human populations, independent of the year of the outbreak. Note that multiple
breaks occurred at some of these locations. The size of the red dot represents the size of the largest
outbreaks occurred at some of these locations. The size of the red dot represents the size of the largest
outbreak that occurred at each location. Range data for the Egyptian fruit bat from IUCN red list
outbreak that occurred at each location. Range data for the Egyptian fruit bat from IUCN red list
repository. (B) Predicted risk of Marburg virus infection based on our Bayesian model prediction.
repository.
Grey(B) Predicted
polygon risk of Marburg
represents virus
95% credible infection based on our Bayesian model prediction.
intervals.
Grey polygon represents 95% credible intervals.
We obtained monthly rainfall temperatures from the Historical monthly weather
We obtained monthly rainfall temperatures from the Historical monthly weather
data [25], downscaled with WorldClim 2.1; [22]. We then extracted monthly rainfall and
data [25], downscaled with WorldClim 2.1; [22]. We then extracted monthly rainfall and
minimum monthly temperature for each set of coordinates in our database (N = 13, pack-
minimum monthly temperature for each set of coordinates in our database (N = 13, package:
age: raster; [26]. We focused on minimum monthly temperature somewhat ar bitrarily
raster; [26]. We focused on minimum monthly temperature somewhat ar bitrarily from
from among several highly correlated measures of temperature we could have focused on
among several highly correlated measures of temperature we could have focused on (e.g.,
(e.g., maximum monthly temperature, mean monthly temperature). However, we specu-
maximum monthly temperature, mean monthly temperature). However, we speculate that
late that
in a tropical in a tropical
environment environment
cold temperaturecold temperature
anomalies anomalies
seem to seem to
be particularly be particularly
stressful to
hosts and their pathogens. We focused on this measure of environmental variation rather
than historical anomalies derived from timeseries analyses (e.g., [27]) as it closely mirrors
the measures of climatic variation used in previous studies that supported the influence of
Schmalhausen’s law in vector-borne disease systems [7–9].
We obtained the spatial range of the Egyptian fruit bat as a polygon shapefile from
the IUCN red list repository [28]. Using this information, we then extracted the minimum
distance between each location and the edge of the closest polygon, representing a mini-
mum distance between a Marburg virus outbreak and the distributional edge of the natural
reservoir of the Marburg virus using ArcGIS V10.4 [29].
2.2. Data Analysis
All analyses were performed using R version 4.0.2 [30]. We provide our R code as
Supplementary Materials to allow readers to fully reproduce our results and provide a
useful starting point for similar analyses in other systems. A known source of bias that
could influence our response variable is human population density [31,32], where larger
outbreaks occur in areas of higher density. To ensure that our results were not influenced
by this, we downloaded the Population Density Grid, v3 (SEDAC) for the year 2000, and
extracted human population density for our geographic coordinates. We then performed a
Bayesian regression analysis with a negative binomial distribution (package: brms; [33]),Viruses 2023, 15, 310 4 of 7
where our response variable was the outbreak size and the predictor was human population
density. We found no relationship between these two variables (effect size (slope) = −0.001,
lower Credible interval = −0.002, upper Credible interval = 0.001), indicating that our
results are unlikely to have been driven by variation in human population density.
First, to understand the importance of our climatic variables on outbreak severity, we
calculated the moving average and standard deviation for both rainfall and temperature for
each sample site with a window of 30 days (package: tidyquant; [34]). We then randomly
subsampled 100 data points for which we knew there were no reported outbreaks, thereby
creating a pseudo-absence data frame. Afterward, we calculated the 95% confidence
intervals of the standard deviation of environmental variables for our pseudo-absence data
frame, and checked if the moving standard deviation for up to a year before outbreaks at
each site fell inside it, allowing us to check for anomalies in the time series for each Marburg
virus outbreak. We considered it unlikely that the ecophysiological effects of climate on
outbreak risk predicting Schmalhausen’s law would exhibit a temporal lag greater than one
year. These analyses showed no evidence of any influence of climatic variation on outbreak
risk (see results), and so no additional analyses of these variables were conducted.
To determine whether there was a significant overall influence of host species spatial
range in the Marburg virus outbreak, we used a Bayesian population-level model (package:
brms; [33]) with weakly informative priors set for our only coefficient. We used four chains,
with 10,000 iterations per chain (5000 for warmup, 5000 for sampling) We used the human
outbreak size (count data) as a response variable with a negative binomial distribution (link
function: log) and the closest distance to the edge of the Egyptian fruit bat distributional
range as our only population-level effect (see Figure 1B). Lastly, we checked for convergence
of our predictor using the RHAT indicator (at convergence, RHAT = 1).
3. Results
The moving monthly standard deviation for rainfall did not deviate from the expected
95% confidence intervals, and neither did the monthly standard deviation for temperature.
In contrast, we detected a relevant effect from the distance to our target species distribu-
tional range on the Marburg virus outbreak, with a mean effect size (slope) of −0.01 (lower
Credible interval: −0.02, Upper Credible interval: −0.01). However, keeping in mind that
this result may be driven by an outlier in a small dataset, we emphasize caution in the
interpretation of this result.
4. Discussion
Schmalhausen’s law predicts that minor environmental changes should strongly
affect outbreak risks in variable areas, particularly for species living at the edge of their
distributional range or environmental tolerances in general [6]. While this prediction
has been tested in multiple vector-borne pathogen systems ([7–9], there is a clear lack of
literature regarding Schmalhausen’s law indirectly transmitted pathogens. In terms of
large-scale disease dynamics, Schmalhausen’s law predicts that species near the edges of
conditions they can tolerate due either to spatial or temporal variation in environmental
conditions have the potential to generate outbreaks with unusual characteristics. Here,
we show how the potential influence of Schmalhausen’s law can be tested for both in the
context of temporal climatic anomalies and distance to species distributional edge (i.e., to
the spatial limits of conditions a species is able to survive and reproduce) using Marburg
virus outbreak as our study system. Directly transmitted pathogens, even those similar
to the Marburg virus such as Ebolaviruses and hemorrhagic fevers in general [35], tend
to have numerous potential wild reservoirs [36]. In contrast, Marburg virus is a relatively
simple system with a single primary reservoir [19,20]. This makes it an ideal choice for a
“proof of concept” study.
While we failed to detect any direct influence of climatic variability on the Marburg
virus outbreak, we still believe that there could be an underlying mechanism dictating
the effects of climatic variability on disease outbreaks in the context of Schmalhausen’sViruses 2023, 15, 310 5 of 7
law. For instance, the pulse of resource hypothesis would prove a valuable alternative
explanation for any climatic patterns related to Schmalhausen’s law, with species gathering
in resource-rich environments after a minor climatic disturbance [37] thus enhancing the
potential for pathogen spillover.
The main finding of this study is that distance to species distributional edge seems
to be a key driver in outbreaks severity of Marburg virus in Africa. Schmalhausen’s law
predicts that when species are pushed to the limits of their ecophysiological tolerances such
as in the boundaries of an ecosystem [6], minor environmental changes can lead to unusual
outbreak dynamics. Here, we show that the Marburg virus outbreak pattern seems to follow
this aspect of Schmalhausen’s law, with outbreaks having an increased severity and the
edge of the Egyptian fruit bat distribution. However, the mechanisms behind this remain
a matter for debate and there are few tests of any of the predictions of Schmalhausen’s
law in directly transmitted pathogens (but see [10]). We posit that the pattern observed
here could be due to the presence of either natural or artificial ecotones, increasing the rate
of contact between the Egyptian fruit bat and human settlements, resulting in a higher
probability of a spillover event [4,38]. This phenomenon has been demonstrated in multiple
systems, for instance in intensified agricultural landscape (see systematic review by [39],
and we believe this could provide a plausible explanation for what we observe here as well.
Another equally suitable explanation for our result would be a change in R0 of the Marburg
virus in stressful areas. For instance, [40] showed that many directly transmitted pathogen
transmissions were increased with climatic perturbations, mainly due to changes in habitat
range and increased rates of contact between species. Lastly, it is possible that individuals
present at the edge of their ecophysiological tolerance, and hence in stressful conditions,
will not be able to allocate sufficient resources toward their immunity, resulting in increased
chances of spillover in these environments [13]. As such, we bring forward that an increase
in transmission rates of the Marburg virus at the edge of the reservoir species distribution
would create larger outbreaks.
Of course, the result presented here should be interpreted carefully. The obvious
caveat of this study is the small sample size: we decided to use the Marburg virus outbreak
to test Schmalhausen’s law in a directly transmitted pathogen since it remains a simple
system of critical interest from a One Health perspective. However, it is also a relatively
rare pathogen, with only 13 documented natural outbreaks to date (Figure 1A), which
limited our statistical power. In addition, the main result of our study may be biased by
an outlier, with the only large (100+ cases) outbreak of the Marburg virus happening at
the edge of the Egyptian fruit bat range in Angola. This remains an odd, but nevertheless
interesting, an observation which would benefit from increased research. Despite these
limitations, we were able we show preliminary support for the influence of Schmalhausen’s
law on directly transmitted pathogens. A major improvement in this system would be
to measure viral prevalence in species edge compared to the center of their distribution,
as this help distinguish whether mechanisms related to hosting physiology and immune
function or alternative mechanisms such as increased contact rates between the reservoir
and other species including human play a role.
The purpose of our study was to determine whether we could detect evidence of the
disease dynamics predicted by Schmalhausen’s law in this simple one-reservoir system
in which vector-borne transmission does not play a role, as a proof-of-concept. We thus
focused on factors expected to push populations of the primary reservoir of the Marburg
virus to the edges of their physiological tolerances. Our results demonstrate that distance to
the range edges of known hosts may be an important factor to include in future studies of
outbreak risk, even for directly transmitted pathogens with endothermic reservoirs. How-
ever, previous studies have shown that a number of other factors including non-anomalous
climatic variation, patterns of deforestation, and human population density [41,42] are also
important predictors of filovirus outbreak risk. In addition to host range effects, these
would be important to consider in future studies focused on building the most accurate
possible predictive models of outbreak risk of filoviruses or other pathogens.Viruses 2023, 15, 310 6 of 7
With an increase in pathogen spillover leading to zoonotic events in many terrestrial
systems around the world, we believe that there is a strong need to better understand the
drivers of past and current outbreaks in order to mitigate the impact of future pathogen
spillover events.
Supplementary Materials: The following supporting information can be downloaded at: https:
//www.mdpi.com/article/10.3390/v15020310/s1.
Author Contributions: A.F. and P.R.S. conceived the idea and designed the study. M.S. collected
the data. A.F. wrote the first draft of the manuscript. All authors were involved in data analyses,
contributed critically to the drafts, and gave final approval for publication. All authors have read and
agreed to the published version of the manuscript.
Funding: This work was supported by NIH R01Al156866 “Spillover of ebola and other filoviruses at
ecological boundaries.” awarded to Patrick R. Stephens.
Institutional Review Board Statement: Not applicable.
Informed Consent Statement: Not applicable.
Data Availability Statement: Upon acceptance of this manuscript, dataset and code will be provided
via DRYAD repository. The code is not novel code but is there to help other researcher achieve
reproducibility with our dataset.
Acknowledgments: The authors would like to thank 2 anonymous reviewers for their comments
and efforts in improving this manuscript.
Conflicts of Interest: The authors declare no conflict of interest.
References
1. Gottdenker, N.L.; Streicker, D.G.; Faust, C.L.; Caroll, R. Anthropogenic Land Use Change and Infectious Diseases: A Review of
the Evidence. EcoHealth 2014, 11, 619–632. [CrossRef]
2. Risser, P.G. The Status of the Science Examining Ecotones. Biosci. 1995, 45, 318–325. [CrossRef]
3. Kark, S. Ecotones and Ecological Gradients. In Ecological Systems; Leemans, R., Ed.; Springer: New York, NY, USA, 2013; pp.
147–160.
4. Despommier, D.; Ellis, B.R.; Wilcox, B.A. The role of Ecotones in Emerging Infectious Diseases. EcoHealth 2007, 3, 281–289.
[CrossRef]
5. LoGiudice, K.; Ostfeld, R.S.; Schmidt, K.A.; Keesing, F. The ecology of infectious disease: Effects of host diversity and community
composition on Lyme disease risk. Proc. Natl. Acad. Sci. USA 2003, 100, 567–571. [CrossRef]
6. Lewontin, R.; Levins, R. Schmalhausen’s law. Capital. Nat. Social. 2000, 11, 103–108. [CrossRef]
7. Chaves, F.L.; Koenraadt, C.J.M. Climate Change and Highland Malaria: Fresh Air for a Hot Debate. Q Rev. Biol. 2010, 85, 27–55.
[CrossRef]
8. Poh, K.C.; Chaves, F.L.; Reyna-Nava, M.; Roberts, C.M.; Fredregill, C. The influence of weather and weather variability on
mosquito abundance and infection with West Nile virus in Harris County, Texas, USA. Sci. Total Environ. 2019, 625, 260–272.
[CrossRef]
9. Chaves, F.L.; Cohen, J.M.; Pascual, M.; Wilson, M.L. Social Exclusion Modifies Climate and Deforestation Impacts on a Vector-
Borne Disease. PLoS Negl. Trop. Dis. 2008, 2, e176. [CrossRef]
10. Briers, R.A. Range limits and parasite prevalence in a freshwater snail. Proc. R. Soc. London Ser. B Biol. Sci. 2003, 270 (Suppl. S2),
S178–S180. [CrossRef]
11. Ardia, D.R.; Parmentier, H.K.; Vogel, L.A. The role of constraints and limitation in driving individual variation in immune
response. Funct. Ecol. 2011, 25, 61–73. [CrossRef]
12. Becker, D.J.; Down, C.J.; Martin, L.B. Multi-Scale Drivers of Immunological Variation and Consequences for Infectious Disease
Dynamics. Integr. Comp. Biol. 2019, 59, 1129–1137. [CrossRef] [PubMed]
13. Merrill, L.T.E.; Barger, A.M.; Benson, T.J. Avian Health across the Landscape: Nestling Immunity Covaries with Changing
Landcover. Integr. Comp. Biol. 2019, 59, 1150–1164. [CrossRef] [PubMed]
14. Borremans, B.; Faust, C.; Manlove, K.R.; Sokolow, S.H.; Lloyd-Smith, J.O. Cross-species pathogen spillover across ecosystem
boundaries: Mechanisms and theory. Philos. Trans. R. Soc. 2019, B37420180344. [CrossRef]
15. World Health Organisation. Marburg Virus Disease. 2022. Available online: https://www.who.int/health-topics/marburg-virus-
disease#tab=tab_1 (accessed on 12 September 2022).
16. Red Cross. Angola: Marburg Haemorrhagic Fever Outbreak Final Report. DREF Bulletin 2006. Available online: https:
//reliefweb.int/sites/reliefweb.int/files/resources/2EF170EDC6219323C12571FE004C9D3D-ifrc-ang-05oct.pdf (accessed on 12
September 2022).Viruses 2023, 15, 310 7 of 7
17. Ajelli, M.; Merler, S. Transmission potential and design of adequate control measures for Marburg haemorrhagic fever. PLoS ONE
2012, 7, e50948. [CrossRef] [PubMed]
18. Berger, S. GIDEON Guide to Outbreaks; Global Ecology and Infectious Disease Network: Los Angeles, CA, USA, 2017.
19. Towner, J.S.; Amman, B.R.; Sealy, T.K.; Carroll, S.A.R.; Comer, J.A.; Kemp, A.; Swanepoel, R.; Paddock, C.D.; Balinandi, S.;
Khristova, M.L.; et al. Isolation of genetically diverse Marburg viruses from Egyptian fruit bats. PLoS Pathog. 2009, 5, e1000536.
[CrossRef]
20. Center for Disease Control and Prevention. Marburg (Marburg Virus Disease). 2022. Available online: https://www.cdc.gov/
vhf/marburg/about.html (accessed on 12 September 2022).
21. Amman, B.R.; Carroll, S.A.; Reed, Z.D.; Sealy, T.K.; Balinandi, S.; Swanepoel, R.; Kemp, A.; Erickson, B.R.; Comer, J.A.; Campbell,
S.; et al. Seasonal Pulses of Marburg Virus Circulation in Juvenile Rousettus aegyptiacus Bats Coincide with Periods of Increased
Risk of Human Infection. PLoS Pathog. 2012, 8, e1002877. [CrossRef]
22. Fick, S.E.; Hijmans, R.J. WorldClim 2, New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37,
4302–4315. [CrossRef]
23. Kuhn, J.S. Filoviruses: A Compendium of 40 Years of Epidemiological, Clinical, and Laboratory Studies; Springer: Wien, New York, NY,
USA, 2008; p. 413.
24. Madoff, L.C. ProMED-mail: An early warning system for emerging diseases. Clin. Infect. Dis. 2004, 39, 227–232. [CrossRef]
25. Harris, I.; Jones, P.D.; Osborn, T.J.; Lister, D.H. Updated high-resolution grids of monthly climatic observations—The CRU TS3.10
Dataset. Int. J. Climatol. 2014, 34, 623–642. [CrossRef]
26. Hijmans, R.J. raster: Geographic Data Analysis and Modelling; R Package Version 3.1-5; 2020. 2020.
27. Sillmann, J.; Kharin, V.; Zwiers, F.; Zhang, X.; Bronaugh, D. Climate extremes indices in the CMIP5 multi-model ensemble. Part 1:
Model evaluation in the present climate. J. Geophys. Res. 2013, 118, 1716–1733. [CrossRef]
28. IUCN. The IUCN Red List of Threatened Species. Version 2022-1. 2022. Available online: https://www.iucnredlist.org (accessed
on 8 September 2022).
29. ESRI ArcGIS Desktop: Release 10.4; Environmental Systems Research Institute: Redlands, CA, USA, 2016.
30. R Core team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria,
2020.
31. Hu, H.; Nigmatulina, K.; Eckhoff, P. The scaling of contact rates with population density for the infectious disease models. Math.
Biosci. 2013, 244, 125–134. [CrossRef] [PubMed]
32. Li, R.; Richmond, P.; Roehner, B.M. Effect of population density on epidemics. Phys. A Stat. Mech. Its Appl. 2018, 510, 713–724.
[CrossRef]
33. Bürkner, P.C. Brms: An R Package for Bayesian Multilevel Models Using Stan. J. Stat. Softw. 2017, 80, 1–28. [CrossRef]
34. Dancho, M.; Vaughan, D. Tidyquant: Tidy Quantitative Financial Analysis; R Package Version 1.0.5; 2022.
35. Center for Disease Control and Prevention (2022b) Viral Haemorrhagic Fevers Fact Sheet. Available online: https://www.cdc.
gov/ncidod/dvrd/spb/mnpages/dispages/fact_sheets/viral_hemorrhagic_fevers_fact_sheet.pdf (accessed on 13 October 2022).
36. Haydon, D.T.; Cleaveland, S.; Taylor, L.H.; Laurenson, M.K. Identifying reservoirs of infection: A conceptual and practical
challenge. Emerg. Infect. Dis. 2002, 8, 1468–1473.
37. Chase, J.M.; Knight, T.M. Drought-induced mosquito outbreaks in wetlands. Ecology 2003, 6, 1017–1024. [CrossRef]
38. Wilkinson, D.A.; Marshall, J.C.; French, N.P.; Hayman, D.T.S. Habitat fragmentation, biodiversity loss and the risk of novel
infectious disease emergence. Phil. Trans. R. Soc. B 2018, 374, 20180344. [CrossRef]
39. Jones, B.A.; Grace, D.; Kock, R.; Alonso, S.; Rushton, J.; Said, M.Y.; McKeever, D.; Mutua, F.; Young, J.; McDermott, J.; et al.
Zoonosis emergence linked to agricultural intensification and environmental change. Proc. Natl. Acad. Sci. USA 2013, 110,
8399–8404. [CrossRef]
40. Mora, C.; McKenzie, T.; Gaw, I.S.; Dean, J.M.; von Hammerstein, H.; Knudson, T.A.; Setter, R.O.; Smith, C.Z.; Webster, K.M.; Patz,
J.A.; et al. Over half of known human pathogenic diseases can be aggravated by climate change. Nat. Clim. Chang. 2022, 12,
869–875. [CrossRef]
41. Schmidt, J.P.; Park, A.W.; Kramer, A.M.; Han, B.A.; Alexander, L.A.; Drake, J.M. Spatiotemporal fluctuations and triggers of Ebola
virus spillover. Emerg. Infect. Dis. 2017, 23, 415–422. [CrossRef]
42. Olivero, J.; Fa, J.E.; Real, R.; Márquez, A.L.; Farfán, M.A.; Vargas, J.M.; Gaveau, D.; Salim, M.A.; Park, D.; Suter, J.; et al. Recent
loss of closed forests is associated with Ebola virus disease outbreaks. Sci. Rep. 2017, 7, 1–9. [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual
author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to
people or property resulting from any ideas, methods, instructions or products referred to in the content.You can also read