Non-canonical argonaute loading of extracellular vesicle-derived exogenous single-stranded miRNA in recipient cells
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
© 2021. Published by The Company of Biologists Ltd | Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914
RESEARCH ARTICLE
Non-canonical argonaute loading of extracellular vesicle-derived
exogenous single-stranded miRNA in recipient cells
Bartika Ghoshal1, Edouard Bertrand2 and Suvendra N. Bhattacharyya1,*
ABSTRACT 2014). EVs, and exosomes in particular, are known to carry different
MicroRNAs (miRNAs), the tiny regulators of gene expression, can be cargoes, including mRNAs, microRNAs (miRNAs), proteins and
transferred between neighbouring cells via extracellular vesicles macromolecules, to the recipient cell to ensure intercellular
(EVs) to control the expression of genes in both donor and recipient communication and exchange of materials.
cells. How the EV-derived miRNAs are internalized and become miRNAs are 20–22-nucleotide-long gene regulatory RNAs,
functional in target cells is an unresolved question. We have which by base pairing to target messages can trigger translational
expressed a liver-specific miRNA, miR-122, in non-hepatic cells for repression or degradation of target RNAs (Fabian et al., 2010;
packaging in released EVs. With these EVs, we have followed the Filipowicz et al., 2008). miRNAs target important regulatory genes
trafficking of miR-122 to recipient HeLa cells that otherwise do not and play crucial roles in disease pathogenesis. miRNAs have been
express this miRNA. We found that EV-associated miR-122 is found to be present in the body fluids either as ‘free’ entities
primarily single-stranded and, to become functional, is loaded onto (sometimes in complex with the argonaute proteins) or in enclosed
the recipient cell argonaute proteins without requiring host Dicer1. vesicles, such as exosomes (Arroyo et al., 2011; Kosaka et al.,
Following endocytosis, EV-associated miR-122 is loaded onto the 2010a).
host cell argonaute proteins on the endosomal membrane, where the Transfer of functional miRNAs by EVs to recipient cells has been
release of internalized miRNAs occurs in a pH-dependent manner, observed to evoke a physiological response both in cancer and
facilitating the formation of the exogenous miRNP pool in the recipient immune cells (Kogure et al., 2011; Mittelbrunn et al., 2011;
cells. Endosome maturation defects affect EV-mediated entry of Montecalvo et al., 2012; Pegtel et al., 2010; Salido-Guadarrama
exogeneous miRNAs in mammalian cells. et al., 2014; Valadi et al., 2007). EV-mediated crosstalk between
cells having different miRNA expression profiles has also been
This article has an associated First Person interview with the first reported (Basu and Bhattacharyya, 2014). Specific sets of proteins
author of the paper. have been observed to selectively sort and package certain miRNAs
into MVBs. These MVBs then fuse with the cell membrane to
KEY WORDS: miRNA, Extracellular vesicles, Argonaute, miRNA release the intraluminal EVs, containing specific miRNAs, in a
loading, miRNA endocytosis, Endosomes, Dicer1 context-dependent manner (Cha et al., 2015; Mukherjee et al., 2016;
Shurtleff et al., 2016; Villarroya-Beltri et al., 2013).
INTRODUCTION The internalization of EVs into recipient cells occurs either
Intercellular communication in metazoan cells, within the same or by endocytosis (Svensson et al., 2013; Tian et al., 2010),
different tissues, can be achieved via exchange of membrane- macropinocytosis (Tian et al., 2014), phagocytosis (Feng et al.,
enclosed vesicles that can carry proteins or nucleic acids (Maas 2010) or engulfment with the help of filopodia (Heusermann et al.,
et al., 2017). This exchange of materials is considered as a very 2016). However, despite recent advances in the study of EV-
important physiological phenomenon in animals (Stahl and Raposo, mediated transport of miRNA, the mode of uptake of these miRNAs
2019). These extracellular vesicles (EVs), as they are collectively and the factors that render them functional in recipient cells are
known, either originate from the multivesicular bodies (MVBs) or entirely unexplored. Using an ectopic miRNA expression system in
the plasma membrane and are classified according to their origin human HeLa cells, we have observed functional miRNA transfer to
and physical status as microvesicles, ectosomes, microparticles or the recipient cell by EVs. This exogenous miRNA enters the
exosomes (van Niel et al., 2018). Exosomes are 40–100 nm-sized recipient cell in a single-stranded form. Internalized miRNAs
vesicles that are positive for the tetraspanin protein CD63 and become associated with the recipient cell argonaute proteins in a
are formed during membrane invagination of late endosomes to Dicer1-independent manner. We have observed that the internalized
generate MVBs. Following MVB fusion with the cell membrane, miRNAs utilize the endocytic pathway to reach the endoplasmic
exosomes are released into the extracellular space (Colombo et al., reticulum (ER) of target cells to elicit their repressive action. We
Journal of Cell Science
have substantiated our data in an in vitro assay to show that a
1
decrease in endosomal pH causes the release of the miRNA from the
RNA Biology Research Laboratory, Molecular Genetics Division, CSIR-Indian
Institute of Chemical Biology, Kolkata 700032, India. 2Institut de Gé né tique
internalized EVs – a prerequisite process for functional loading of
Molé culaire de Montpellier, Université de Montpellier, CNRS, Montpellier 34293, internalized miRNAs onto argonaute 2 (Ago2) of the recipient cells.
France.
*Author for correspondence (suvendra@iicb.res.in) RESULTS
Exogenously expressed liver-specific mature miR-122 is
B.G., 0000-0003-0216-2348; E.B., 0000-0002-9642-7994; S.N.B., 0000-0001- packaged in the EVs released by non-hepatic HeLa cells
9697-3824
To understand the mechanistic aspects of uptake of EV-enclosed
Handling Editor: Maria Carmo-Fonseca
miRNAs, we used HeLa cells ectopically expressing a ‘foreign’
Received 3 September 2020; Accepted 19 March 2021 liver-specific miR-122 as donor cells that would produce EVs
1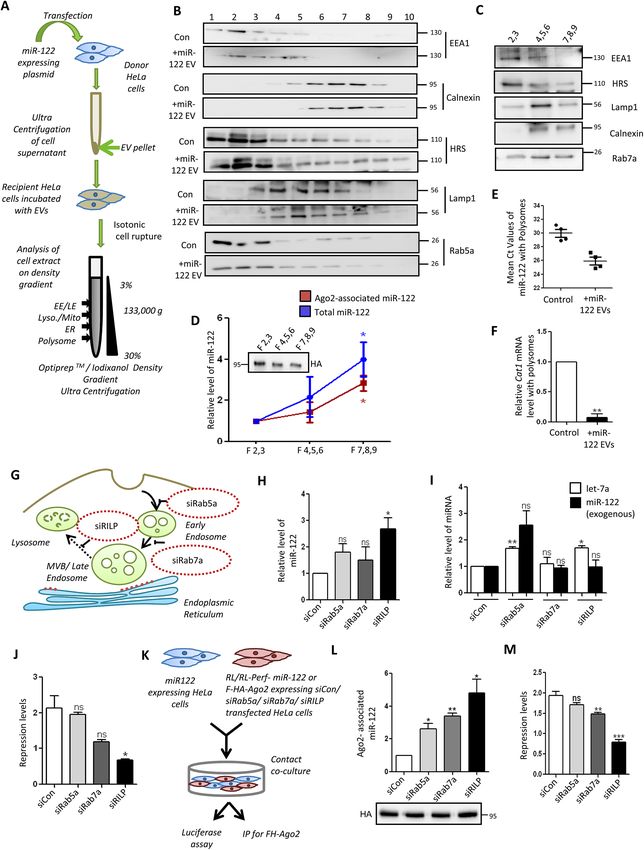
RESEARCH ARTICLE Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914
containing miR-122. This miRNA is expressed abundantly in the The EVs were isolated from these donor cells and added to recipient
liver cells, and is otherwise not detectable in HeLa cells (Landgraf cells (Fig. 1A, left panel). Confocal microscopy analysis suggested
et al., 2007). HeLa cells without the miR-122 expression cassette internalization of CD63–GFP EVs in recipient cells compared to
thus could serve as recipient cells to measure the EV-mediated control cells. Three-dimensional (3D) reconstitution from several
uptake and functional activity of transferred miR-122. To z-plane confocal slices further demonstrated the internalization of
characterize the miR-122-containing EVs, HeLa cells transfected GFP-tagged CD63-containing EVs (Fig. 1A, right panels). The
with miR-122 expression plasmid were subjected to EV isolation by CD63–GFP EVs, used for the purpose of microscopy, were isolated
ultracentrifugation of culture supernatants (Fig. S1A). The EVs using a commercial exosome isolation reagent. Thus, we compared
isolated from either control or miR-122-expressing cells were them with the EVs obtained from ultracentrifugation and found no
subjected to nanoparticle tracking analysis (NTA) to obtain an difference in size or concentration (Fig. S2A,B,D,E). This was
estimate of size and number of the two groups of EVs. No difference consistent with a previous report (Cheng et al., 2019). However,
in size or concentration was observed for the two EV groups (Fig. upon western blot analysis, protein levels were found to be higher in
S1B,C). Atomic force microscopy (AFM) revealed that there was the CD63–GFP-positive EVs than in the control set, suggesting an
also no difference in their shapes (Fig. S1D). The EV protein increase in protein content upon expression of CD63–GFP in donor
content showed no major change between the control and miR-122- cells (Fig. S2C). Upon incubating the recipient cells with CD63–
containing EVs. The isolated EVs were found to be positive for GFP positive EVs for 0, 4 and 16 h, we found that the number of
CD63, flotillin 1, Alix (also known as PDCD6IP) and HRS (also internalized GFP-positive EVs per cell increased at 16 h (Fig. 1B
known as HGS), and were devoid of most cellular markers, including and Fig. 1C, left panel). Consistent with this, we measured the levels
calnexin, Lamp1 and GAPDH. Furthermore, the EVs used for of EV-derived miR-122 in the recipient cells over time upon
analysis were free from the apoptotic body marker cytochrome c as treatment with miR-122-positive EVs isolated by ultracentrifugation
well as lipoprotein contaminants like ApoE (Fig. S1E). and found an increase in the internalized miRNA content at 16 h
We then further confirmed the presence of mature miR-122 in the (Fig. 1C, right panel).
EVs isolated from miR-122-expressing donor HeLa cells compared The miR-122 antisense strand (miR-122*) was not detected in
to EVs from the control HeLa cells. The levels of mature miR-122 recipient cells incubated with miR-122-positive EVs (Fig. 1D, left
detected in EVs from miR-122-expressing donor cells remained panel), suggesting exclusive transfer of mature miR-122 to the
unchanged upon RNase-free DNaseI treatment. This data thus ruled recipient cells. We found ∼2800 copies of miR-122 after 16 h of
out the possible presence of miR-122-encoding DNA fragment as a treatment of the recipient cells with miR-122-positive EVs (Fig. 1D,
contaminant in the EVs (Fig. S1F). Upon RT-qPCR threshold cycle right panel). We also quantified the levels of precursor miR-122 in
(Ct) value analysis, we detected a marginal level of pre-miR-122 in the recipient cells and found no change in the precursor levels
EVs that was resistant to DNaseI (Fig. S1G). Previous reports in both control and miR-122 EV-treated cells (Fig. 1E), which
suggest that EV-associated RNAs are usually localized inside the were 500-fold less compared to those in hepatic origin Huh7 cells
EVs (Del Pozo-Acebo et al., 2021; Fabbiano et al., 2020; Shurtleff (data not shown).
et al., 2017; Valadi et al., 2007). To confirm the miR-122 In order to investigate the functionality of the transferred miRNA,
localization within EVs, we treated the EVs with RNase A. We luciferase assays were performed using a Renilla luciferase (RL)-
found protection of the majority of miR-122 from RNase A unless based reporter plasmid containing a perfect binding site for miR-
the EVs were subjected to sonication before the RNase A treatment. 122 (RL-Perf-miR-122; Fig. 1F). This plasmid was transfected into
Sonication disrupts membranous structures (Alvarez-Erviti et al., recipient cells that were either treated or untreated with miR-122-
2011; Kojima et al., 2018), and we found decreased miR-122 containing EVs. The firefly luciferase-normalized expression levels
content in EVs that were sonicated and treated with RNase A (Fig. of RL-Perf-miR-122 were found to be low in the recipient cells
S1H,I). Thus, the majority of EV-associated miRNAs are present incubated with miR-122-positive EVs (Fig. 1G, left panel). Upon
inside the vesicles and protected from RNase A. transfer of miR-122 via EVs, we found decreased expression of
Ct value comparison revealed that the antisense strand of miR- miR-122 target mRNAs, including cationic amino acid transporter 1
122 (miR-122*) was present in a negligible amount, as reflected by (CAT1, also known as SLC7A1) and aldolase A (ALDOA), in EV-
the very high Ct value associated with it (Fig. S1J, left panel). This treated cells, which signifies functional transfer of EV-derived miR-
signifies strand-specific packing of mature miR-122 and not miR- 122 across donor and recipient cell boundaries (Fig. 1G, right
122* into EVs. Before proceeding further, we also quantified the panel). Similar to miR-122, ectopic expression of miR-146a in
copy number of mature miR-122 present in the HeLa EVs either HeLa cells also ensured its EV-mediated transfer to recipient HeLa
isolated from miR-122-transfected or untransfected cells and found cells that otherwise do not express miR-146a (Fig. 1H). This result
that EVs obtained from miR-122-transfected cells contain ∼2 signifies the universality of the heterogeneous miRNA transfer
copies (1.7 copies on average) of the miRNA (Fig. S1J, right panel). mechanism. We investigated whether the transferred miR-122 can
This is consistent with a previous report where a similar copy affect the levels and activity of endogenous miRNAs and found no
Journal of Cell Science
number of miRNAs in EVs has been reported (Alexander et al., change in endogenous let-7a and miR-16 levels upon miR-122
2015). Finally, a TUNEL assay showed that miR-122 transfection transfer. Let-7a repressive activity also remained unchanged upon
did not induce apoptosis in HeLa cells (Fig. S1K). miR-122 uptake for both the reporter mRNA and mRNA levels of
the endogenous let-7a target K-RAS (Fig. 1I–K).
EVs can transfer functional miRNAs between mammalian
cells The internalized miRNAs associate with Ago2 in recipient
Having established that the mature miR-122 is localized inside the cells in a Dicer1-independent manner
EVs, we wanted to see the transfer and functionality of the One possible mechanism that ensures miRNA homeostasis in animal
internalized miRNA in recipient cells devoid of miR-122. To cells is the EV-mediated transfer of miRNAs between neighbouring
visualize the transfer process kinetics, we transfected our donor cells cells. This may also happen between cells in contact co-culture (Basu
with a plasmid expressing GFP-tagged tetraspanin marker CD63. and Bhattacharyya, 2014). In order to revalidate the uptake of
2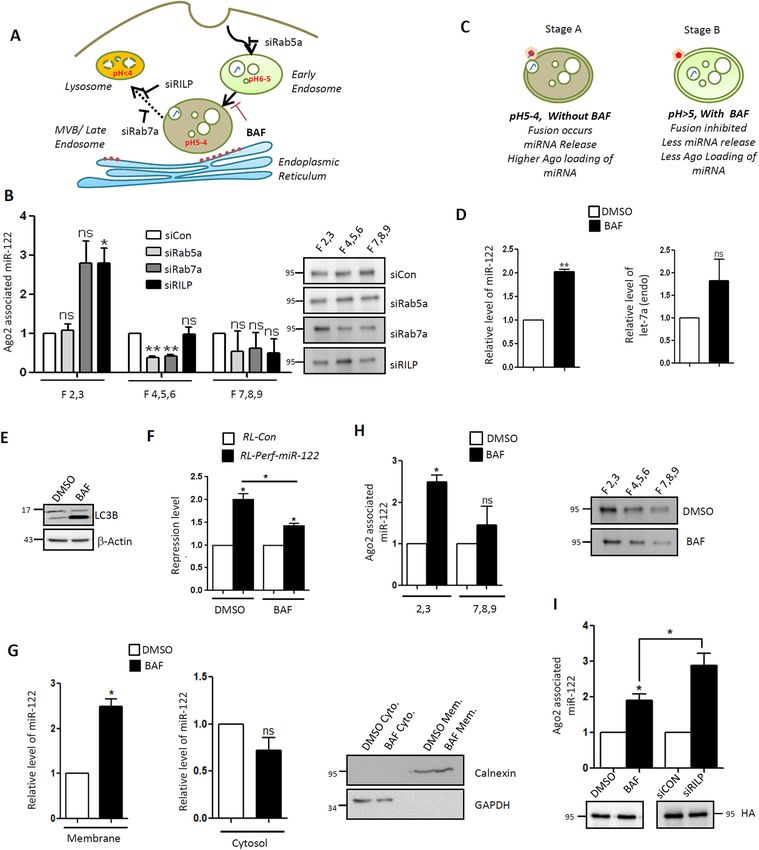
RESEARCH ARTICLE Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914
Journal of Cell Science
Fig. 1. See next page for legend.
3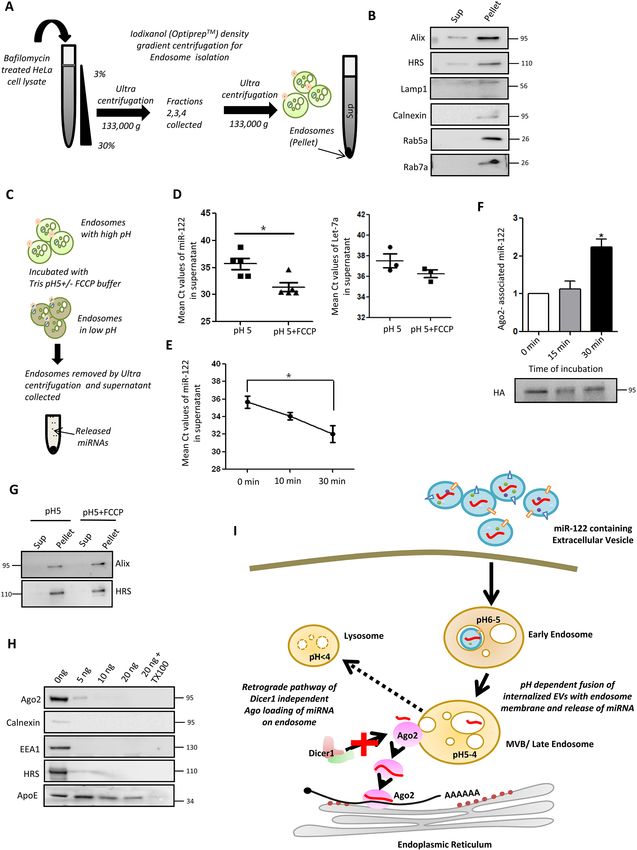
RESEARCH ARTICLE Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914
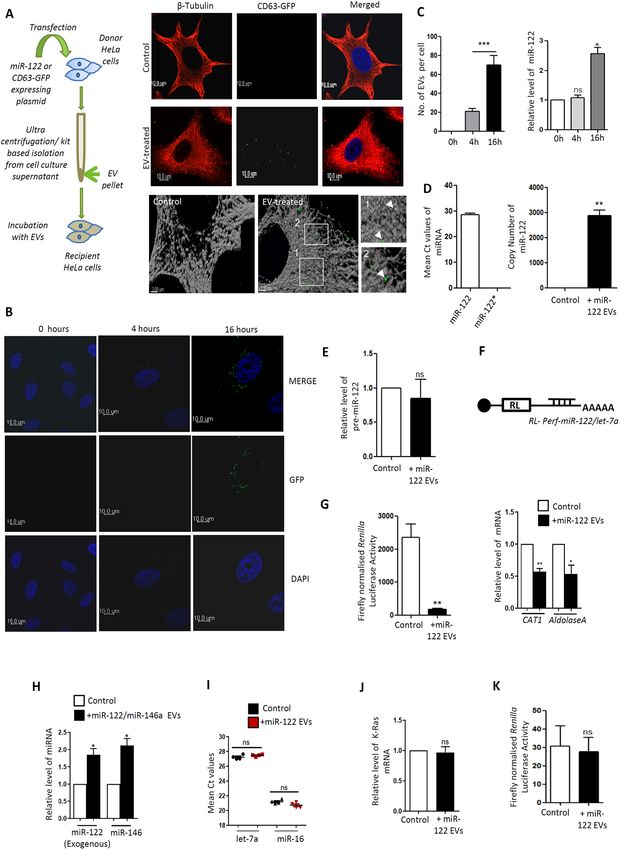
Fig. 1. EV-derived miRNAs are transferred across cell boundaries. (A) and the other with a miR-122 reporter, we tested the activity of the
Scheme of EV-mediated miR-122 transfer in non-hepatic HeLa cells (left). EVs transferred miRNA across cell boundaries. We noted transfer of miR-
isolated by ultracentrifugation from miR-122-expressing HeLa cells were
122 repressive activity to cells having no expression of miR-122, as
added to recipient HeLa cells. To visualize EV uptake, EVs isolated
commercially from CD63–GFP-expressing HeLa cells were added to HeLa expected, and this transfer had no effect on endogenous let-7a miRNA
recipient cells. Confocal images of one plane (top right panels) show activity (Fig. 2C,D). The functionality of the transferred miR-122 was
internalization of EVs in recipient cells following incubation with CD63–GFP also validated by a decrease in CAT1 levels in cells co-cultured with
EVs. The β-tubulin network (red) was detected simultaneously while imaging miR-122-expressing cells (Fig. 2E).
CD63–GFP (green). Nuclei were stained using DAPI (blue). Scale bars: We wanted to check the involvement of recipient cell Ago2 in the
10 µm. The bottom panel shows a zoomed 3D reconstituted image of the
uptake process. Using isolated miR-122-containing EVs as
recipient cells with or without CD63–GFP EVs. The β-tubulin is pseudo-
coloured grey and the CD63–GFP EVs are green. Boxes mark the positions of
described in the experimental scheme shown in Fig. 1A, we
inset images showing CD63–GFP EVs inside the recipient cells (white arrows). observed a similar time-dependent increase in the levels of Ago2-
Images are representative of three independent experiments. Scale bars: associated miR-122 upon EV treatment at 16 h (Fig. 2F). Transfer of
3 µm. (B) Confocal images of recipient cells after 0, 4 and 16 h of CD63–GFP- EV-derived miRNAs to argonaute proteins was not found to be
positive EV treatment. Detection of internalization of CD63–GFP EVs was exclusive to Ago2; Ago3, upon contact co-culture, was also found
monitored microscopically in one confocal plane. Nuclei were stained using to be loaded with internalized miRNAs, compared to negative
DAPI (blue). Images are representative of three independent experiments
comprising a total of 170 cells experiments. Scale bars: 10 µm for 4 h and 16 h,
control NHA–LacZ protein, when expressed in recipient HeLa cells
15 µm for 0 h. (C) Left: graphical representation of the number of EVs (Fig. 2G).
internalized at the abovementioned time points. n=170; unpaired t-test used for In the canonical pathway, Ago2 loading of miRNA is dependent
analysis. Right: relative levels of miR-122 in recipient cells at 0, 4, 16 h of on its interaction with Dicer1. After processing of pre-miRNA by
treatment with miR-122-containing EVs. U6 snRNA was used as a normalizing Dicer1 to generate the double-stranded miRNAs, the miRNA sense
control. n=3. (D) Left: comparison of the Ct values of miR-122 and miR-122* strand is then loaded onto argonaute proteins in a Dicer1-dependent
(antisense strand) in recipient HeLa cells treated with miR-122-containing EVs.
manner (Song and Rossi, 2017). To test whether the internalization
n=3. Right: the copy number of mature miR-122 in recipient cells untreated
(control) or treated with miR-122-positive EVs. The copy number was and argonaute-association of internalized single-stranded miRNA is
calculated based on a standard curve generated using known concentrations dependent on the prime components of the miRNA biogenesis
of synthetic miR-122. n=3. (E) No change in the amount of precursor miR-122 pathway in the recipient cell, Ago2 and Dicer1 proteins of the recipient
in recipient cells. The recipient cells, either treated or untreated with miR-122- cell were targeted using specific siRNAs. When the recipient cells
containing EVs, were subjected to quantification of precursor miR-122 levels were knocked down for Ago2, the internalization of EV-transferred
by RT-qPCR. 18 s rRNA was used as the normalizing control. n=3.
miR-122 decreased in the recipient cell, suggesting a requirement of
(F) Schematic representation of the Renilla luciferase mRNA reporter
comprising of one perfect binding site for either miR-122 or let-7a (either
recipient cell Ago2 for the internalization of the EV-transferred miR-
miRNA was used depending upon the experiment). This was used to study the 122 (Fig. 2H).
repression activity of transferred miR-122 in recipient cells. (G) Left: the When the recipient cells were knocked down for Dicer1 and
expression levels of RL-Perf-miR-122 in recipient HeLa cells as measured by incubated with miR-122-containing EVs, the levels of internalized
luciferase assay. The recipient cells transfected with RL-Perf-miR-122 plasmid miR-122 remained unchanged, as compared to levels in the control
were either treated or untreated with miR-122-containing EVs. Luciferase (Fig. 2I). Ago2-associated miR-122 levels in cells treated with
assay was performed to measure the expression of RL. Expression from a
firefly luciferase reporter co-transfected with RL reporter was used as
control non-targeting siRNA (siControl) or siRNA targeting Dicer1
transfection control. The graph shows the RL values normalized to the firefly (siDicer1) also remained unchanged in co-culture (Fig. 2J, left
luciferase control for both the groups of recipient cells. n=4; unpaired t-test panel). However, in HeLa cells co-transfected with pre-miR-122
used for analysis. Right: the relative levels of CAT1 and aldolase A mRNAs, expression plasmid and siDicer1, the levels of mature miR-122 were
known endogenous miR-122 targets, in recipient HeLa cells treated or reduced significantly along with a similar decrease, as expected, for
untreated with miR-122-containing EVs. n=3. 18 s rRNA was used as endogenous let-7a levels (Fig. 2J, right panel). Therefore the data on
normalizing control. (H) Detection of miRNA transfer by EVs to recipient cells.
internalization of EV-derived miRNA and its argonaute-loading
In addition to miR-122, miR-146a was studied to see whether the EV-mediated
transfer was restricted to miR-122 or occurred for other miRNAs as well. Level suggest that, independent of the presence of Dicer1 in recipient
of internalization of both miR-122 and miR-146a in recipient HeLa cells was cells, the transfer of miR-122 via EVs involves the single-stranded
observed. n=3. (I–K) No change in endogenous miRNA content or activity and mature form of the miRNA. If the precursor miRNA ( pre-miR-
upon treatment of HeLa cells with miR-122-containing EVs. (I) Real-time PCR 122) were transferred, then knockdown of Dicer1 would be
was used to show no difference between the Ct values of endogenous miRNAs expected to affect miRNA incorporation onto recipient cell Ago2,
let-7a and miR-16 in cells untreated or treated with miR-122 containing EVs
as Dicer1 is required for pre-miRNA processing. Dicer1-
(n=4); unpaired t-test was used for analysis. (J) Relative level of K-RAS mRNA,
an endogenous let-7a target, in recipient cells treated or untreated with miR-
independent non-canonical argonaute-loading of EV-derived
122-positive EVs was analysed. n=3. (K) Relative level of normalized Renilla- mature miRNA is possibly required to elicit a quick and effective
based let7a reporter with a perfect let-7a binding site (RL-Perf-let-7a), in response in the recipient cells.
recipient cells treated or untreated with miR-122-positive EVs. n=4; unpaired t-
Journal of Cell Science
test used for analysis. (I–K). Data are presented as mean±s.e.m. *P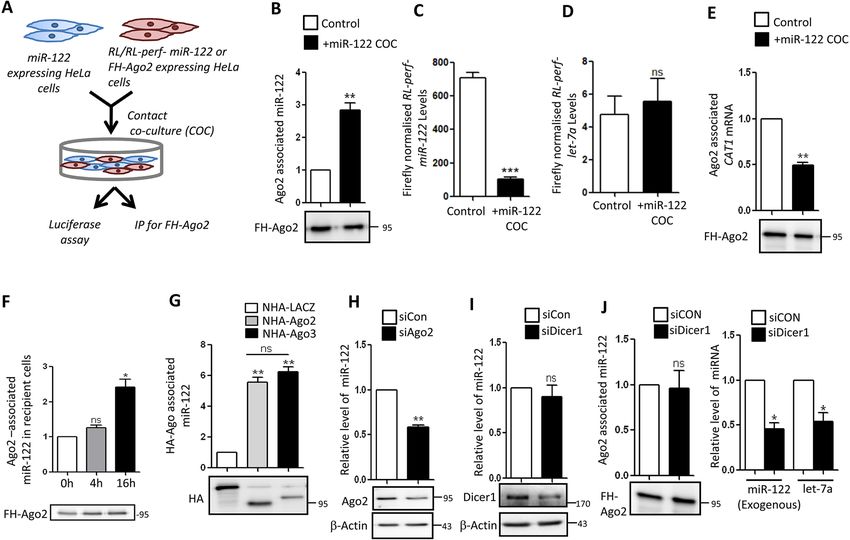
RESEARCH ARTICLE Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914 Fig. 2. Internalization and Ago2 loading of EV-derived miRNAs in recipient cells is independent of Dicer1. (A) Schematic representation of contact co- culture (COC) model of miR-122 transfer in HeLa cells. Donor HeLa cells were transfected with miR-122 expression plasmid and co-cultured with the recipient HeLa cells either expressing FLAG–HA–Ago2 (FH–Ago2) or luciferase reporters. (B) Transfer of miR-122 to recipient cell Ago2 protein. The FLAG–HA–Ago2- associated miR-122 levels in recipient cells co-cultured with or without miR-122-expressing HeLa cells (+miR-122 COC) were calculated using real-time PCR followed by normalization to the FLAG–HA–Ago2 protein levels in the recipient cells, as shown in the representative western blot. n=3. (C) Luciferase assay to show the repression of RL-Perf-miR-122 in recipient cells co-cultured with or without miR-122-expressing donor cells. The levels of firefly luciferase-normalized RL-Perf-miR-122 were compared to show a decrease in expression of the reporter in the presence of miR-122. n=4; unpaired Student’s t-test was performed. (D) Repression of let-7a reporter (RL-Perf-let-7a) in recipient cells co-cultured with HeLa cells expressing or not expressing miR-122. The firefly luciferase- normalized RL values were compared between the cells. n=4; unpaired Student’s t-test was performed. (E) Ago2-associated CAT1 mRNA levels in recipient Hela cells, co-cultured with donor HeLa cells with or without miR-122 expression, was quantified. The mRNA levels were normalized with the band intensity of immunoprecipitated FLAG–HA–Ago2, as shown in the representative western blot. n=3. (F) Time-dependent increase of Ago2-associated EV-derived miR- 122 in recipient cells. The Ago2-associated internalized miR-122 levels in recipient cells were quantified at 0, 4 and 16 h time points. The amount of immunoprecipitated Ago2, as shown in the representative western blot, was used for normalization of miR-122 content. n=3. (G) Association of transferred miR-122 with recipient cell Ago2 or Ago3 in contact co-culture model. Associated miRNA values were normalized to immunoprecipitated levels of NHA–Ago2 or –Ago3 levels (HA; lower panel). NHA–LacZ was used as a negative control. n=3. (H) Effect of depletion of Ago2 on relative level of miR-122 internalization in recipient cells. Levels of miR-122 were measured in HeLa cells transfected with control siRNA (siCon) or siRNA targeting Ago2 (siAgo2) and treated with miR-122-containing EVs. U6 snRNA was used for normalization. Western blotting was used to show knockdown of Ago2. β-actin was used as loading control. n=3. (I) Effect of depletion of Dicer1 on levels of EV-derived miR-122 in HeLa cells. Relative level of total miR-122 internalized after 16 h of treatment of siCon- or siDicer1-transfected recipient HeLa cells with EVs positive for miR-122 (n=4). Western blotting was used to show knockdown of Dicer1. β-actin was used as loading control. (J) Left: Ago2-associated miR-122 was measured in recipient cells transfected with siCon or siDicer1 in the contact co-culture model. Right: relative levels of endogenous let-7a or exogenously expressed miR-122 (expressed as pre-miR-122) in HeLa cells co-transfected with miR- 122-expressing plasmid and siCon or siDicer1. n=3. Data are presented as mean±s.e.m. Representative blots have molecular mass markers indicated in kDa. *P
RESEARCH ARTICLE Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914
Pagano, 2007). Thus, in our case, it is possible that the EV-associated required for early to late endosomal maturation), Rab7a (a late
miRNAs are either internalized by a clathrin-dependent pathway or by endosomal GTPase required for endosome maturation and lysosome
dynamin-dependent but clathrin-independent pathways (for instance interaction) and RILP (Rab-interacting lysosomal protein, which
caveolae-dependent pathways). facilitates the interaction of late endosomes with lysosomes) were
Because it has been reported that the ER serves as the nucleation targeted specifically to dissect which steps of endocytosis are
site for the miRNA machinery (Stalder et al., 2013), we wanted to required for functional miRNA transfer to ER attached polysomes
see what happened to the internalization of EV-derived miRNA (Fig. 3G). In a steady state, endosome numbers were not found to
upon ER stress, which is known to affect miRNA machineries significantly alter on knockdown of the endosomal proteins
(Mukherjee et al., 2016). Thapsigargin is an ER-stress inducer, and individually (Fig. S4A–C). Endosomes were visualized in cells
treatment of recipient cells with thapsigargin followed by incubation expressing an endosomal targeting variant of YFP protein (Endo-
with miR-122-containing EVs resulted in low internalization and YFP). Measuring the effect of knockdown of these factors, we
activity of EV-derived miRNA-122 in treated cells (Fig. S3E–G). found a stronger effect of RILP knockdown on internalized miRNA
Unlike the Dicer1-dependent canonical pathway, where availability content (Fig. 3H). However, the knockdown of RILP marginally
of the target messages has a positive influence on cognate miRNA altered the levels of endogenous let-7a and exogenously expressed
formation (Bose and Bhattacharyya, 2016), we did not detect an miR-122 in recipient HeLa cells (Fig. 3I). Interestingly, Rab5a
increase but rather a decrease of EV-delivered miR-122 content in depletion had a similar effect on both EV-internalized miR-122 and
the presence of its target message (RL-3×Bulge-miR-122). No endogenous miRNAs, and in each case an increase in overall
change in Ago2-associated miRNA level in co-cultured recipient miRNA content was detected. Surprisingly, the increase in miR-122
cells was detected in the presence of target message as well (Fig. content was not reflected in its repressive activity. A reduction in
S3H,I). This data further supports a non-canonical miRNA loading miRNA activity was noted after knockdown of Rab5a or Rab7a
pathway prevalent for EV-internalized miRNAs. and more significantly upon RILP depletion (Fig. 3J). Using the co-
culture model of miRNA transfer (Fig. 3K), we did observe an
EV-derived miRNAs enter recipient cells via the endosomal increase in Ago2 incorporation of internalized miRNAs upon
pathway depletion of the Rabs or RILP but, conversely, we also observed a
We wanted to assess the subcellular localization of the internalized decrease in repressive activity (Fig. 3L,M). We performed
miRNA in the recipient cells. We initially quantified the EV-derived microscopic analysis of the number of CD63–GFP-positive EVs
miR-122 levels in the cytosolic and membrane fractions of the internalized into recipient cells either knocked down for Rab5a,
recipient cells by digitonin fractionation (Fig. S3J). Digitonin Rab7a or RILP. Contrary to our finding that the levels of miRNA
treatment allows permeabilization of cells. This detergent makes the increase upon knockdown of RILP, we found an overall decrease
cell membrane porous by solubilizing high-cholesterol cell in the number of EVs internalized in the recipient cells knocked
membranes and thus separating the cytosolic part as soluble down for either Rab5a or RILP (Fig. S4D,E). As the endocytic
fraction and the detergent-insoluble membranes as the pellet pathway is hampered upon siRNA transfections, the uptake of EVs
fraction (Barman and Bhattacharyya, 2015). We observed that in is probably blocked but the internalized miRNA is stored elsewhere
cells that received EVs containing miR-122, the internalized away from its target mRNA. The mRNA–miRNA interaction,
miRNA was found in the membrane fractions (Fig. S3K,L). This which leads to degradation of the miRNA (Bitetti et al., 2018; de la
was similarly observed when miR-122 was delivered via liposomes Mata et al., 2015; Ghini et al., 2018), being absent causes
(Fig. S3K). To further dissect the subcellular localization of these accumulation of EV-derived miR-122 in Rab7a- or RILP-depleted
internalized miRNAs, cellular lysates of recipient cells post 16 h EV cells. However, this accumulation and Ago2 association appeared
treatment were collected in isotonic solution and fractionated using to be highest in the case of RILP depletion due to low degradation
Optiprep density gradient ultracentrifugation to separate the of EV-derived miR-122, which is consistent with a previous
organelles. Western blot analysis was performed using the fractions observation that RILP knockdown delays the trafficking of EGFR
to demarcate different subcellular compartments (Fig. 3A,B). The from early to late endosomes and prevents their degradation in
different subcellular fractions were collected and pooled together lysosomes (Progida et al., 2007). Thus, our data suggest that the
depending on the presence of marker proteins for different organelles newly formed miRNP is stable and does not reach its target, perhaps
(Fig. 3C), and RNA was isolated from the different pooled fractions. because of an altered localization when endosomal pathways are
Fractions 2 and 3 were enriched for early endosomes, while fractions hampered.
4–6 were enriched for lysosomes and fractions 7–9 had enrichment
for ER marker proteins. In a steady-state context, the majority of the Bafilomycin interferes with acidification of the endosomal
internalized EV-derived miRNAs (total and Ago2-associated) were compartments thus leading to hindered EV-derived miRNA
found to be associated with the ER (Fig. 3D) and possibly with ER- compartmentalization
attached polysomes. From the relative Ct value analysis, there was The endosomal pathway consists of different compartments that
Journal of Cell Science
also an increase in polysomal miR-122 content upon treatment of carry cargo within organelles. Each component of the endosomal
recipient cells with miR-122 containing EVs (Fig. 3E). This was pathway has a specific pH, culminating in the lysosomal
accompanied by a concomitant decrease in the target CAT1 mRNA compartment having a pH of less than 4 (Fig. 4A). From the
level associated with the polysomes (Fig. 3F). analysis described in the above experiments, it seems that majority
We explored the effect of depletion of specific endosomal of the internalized miRNAs are localized with ER-attached
maturation pathways proteins in mammalian cells on EV-mediated polysomes at steady state upon treatment with miR-122-loaded
miRNA entry and function. The endosomal pathway comprises EVs. With depletion of endogenous Rab7a protein and RILP
distinct membrane-bound compartments, which internalize individually, we observed an increase in miRNA–Ago2 association
molecules from the plasma membrane and recycle them back to specifically in the endosome fraction, which is possibly a result of
the surface or sort them for lysosomal degradation. Three proteins of the miRNA remaining blocked in the endosomes due to a faulty
the endosomal pathway, Rab5a (an early endosomal GTPase endosomal pathway (Fig. 4B). Thus, our data suggest that the
6RESEARCH ARTICLE Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914
Journal of Cell Science
Fig. 3. See next page for legend.
7RESEARCH ARTICLE Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914
Fig. 3. EV-derived miRNA reaches the ER of the recipient cell via the et al., 1998). Importantly, it has also been reported that low pH
endosomal pathway. (A) Schematic representation of the subcellular promotes the fusion of exosomal membrane with cell membranes
fractionation of EV-treated recipient cells to follow the localization of EV-
(Parolini et al., 2009). Thus, it can be hypothesized that upon
transferred miRNA. The recipient cells were incubated with miR-122-
containing EVs followed by lysis and 3–30% Optiprep density gradient lowering the pH of endocytic vesicles, fusion between the
ultracentrifugation to separate the different subcellular fractions. Individual endosomal membrane and internalized EV membrane should
fractions enriched for endosomal or ER marker proteins were pooled occur. Since the luminal pH lowers as endosomes mature,
separately, and RNA isolated from these fractions was quantified by RT-qPCR. endosomal maturation should be an effective mechanism to
EE, early endosome; LE, late endosome; lyso., lysosome; mito., mitochondria. trigger the release of EV-derived miRNAs in the proximity of
(B) Western blot analysis of different marker proteins after Optiprep density
endosomes, and its loading on host Ago2. To test this hypothesis,
gradient ultracentrifugation, to demarcate the different subcellular fractions
enriched for either ER (calnexin), early endosomes (EEA1, HRS and Rab5a)
we blocked endosomal acidification using bafilomycin. We
or lysosomes (Lamp1) from control cells (Con) or cells treated with miR-122- detected an increase in EV-derived miRNA content upon
containing EVs. Blots are representative of three experiments. (C) Western treatment of recipient cells with bafilomycin, concomitant with a
blotting of different pooled fractions from miR-122 EV-treated recipient cells, as decrease in the level of repression by internalized miRNAs
indicated, to show different subcellular regions. Blots are representative of two (Fig. 4C–F). Digitonin fractionation revealed an increase in
experiments. (D) Optiprep density gradient ultracentrifugation of lysates from membrane association of miR-122 in bafilomycin-treated cells
EV-treated recipient cells followed by quantitative real-time PCR estimation of
total miR-122 in pooled fractions. Immunoprecipitation of FLAG–HA–Ago2
compared to that in control cells, whereas the levels of miR-122
from pooled fractions was also performed to see the subcellular localization remained unchanged in the cytosolic fractions (Fig. 4G). We also
and Ago2 association of internalized EV-derived miR-122 (n=4 in both cases). revalidated the subcellular fractionation in recipient cells receiving
Ago2 levels in pooled fractions were estimated in accompanying western blots miR-122 EVs either treated with DMSO or bafilomycin and found
with anti-HA antibody (inset) and used for normalization of Ago2-associated a consistent accumulation of miR-122 in the endosomes compared
miRNA levels. (E) Polysome association of transferred miR-122 in recipient to the ER (Fig. 4H). This was accompanied by a relative reduction
cells. Ct values of transferred miR-122 associated with polysomes in recipient
in Ago2-bound internalized miR-122 levels compared to those in
HeLa cells are plotted. Values in untreated cells were used as control (n=4). (F)
The endogenous miR-122 target, CAT1 mRNA, was quantified in the RILP-knockdown cells, where RILP depletion also increased the
polysomes of recipient cells after miR-122-containing EV treatment to assess total internalized miRNA content with a concomitant increase in
the effect of transferred miRNA on the polysomal CAT1 levels. n=3. (G) Ago2 association (Fig. 4I). Thus, our data suggest that
Schematic representation of the endocytic pathway in human cells. The key bafilomycin, like knockdown of RILP, can block the EV-derived
proteins Rab5a, Rab7a and RILP, which are involved in different steps of miRNAs in the endosomal compartment. However, bafilomycin
endosome maturation pathway, were knocked down individually by treating
interferes with proper acidification of the endosomes, which
recipient HeLa cells with respective siRNAs (siRab5a, siRab7a and siRILP) to
check their effect on internalization of EV-derived miRNAs. MVB,
thereby hinders the endosomal Ago2 association of the
multivesicular body. (H) Relative level of internalized EV-derived miR-122 in internalized miRNAs.
recipient cells transfected with control siRNA (siCon), siRab5a, siRab7a or
siRILP, normalized to levels in siCon-transfected cells. n=3. (I) Relative level of Late endosomal maturation and low pH are required for
exogenously expressed miR-122 and endogenous let-7a levels in cells co- release and Ago2 loading of endocytosed EV-derived
transfected with miR-122-expressing plasmid and either with siCon, siRab5a, miRNAs
siRab7a or siRILP to show the difference in endogenous miRNA level (let-7a)
or exogenously expressed miR-122 level upon depletion of specific endosome
To prove that low pH favours release of miRNA and miRNP
maturation associated proteins. n=3. (J) Repression levels of RL-Perf-miR-122 formation with endosomal Ago2, we first isolated endosomes from
reporter in cells transfected with each of the abovementioned siRNAs against HeLa cells using ultracentrifugation. The cell lysates were first
the endocytic pathway components and treated with miR-122-containing EVs. subjected to subcellular fractionation followed by further
This panel shows fold repression levels of RL-Perf-miR-122 reporter relative to purification of endosome-positive fractions by ultracentrifugation
RL-con (without miR-122 site) in HeLa cells transfected with specific siRNAs (Fig. 5A). These endosomes were analysed by western blotting to
and miR-122-containing EVs, calculated as described in the Materials and
check for characteristic proteins and other subcellular organelle
Methods. n=3; unpaired Student’s t-test used for analysis. (K) Schematic
representation of the contact co-culture model used to see the effect of contaminations (Fig. 5B).
depletion of endocytic pathway proteins on internalization of transferred Bafilomycin prevents the lowering of endosomal pH, and thus
miRNA in recipient cells. (L,M) Revalidation of the effect of transfer of EV- endosomes isolated from bafilomycin-treated cells should have a
derived miR-122 upon knockdown of endocytic pathway proteins in contact co- lower capacity to release endocytosed EV-derived miRNAs, and
culture. (L) Ago2-associated miR-122 levels were quantified using real-time this capacity should be restored by lowering endosomal pH. We thus
PCR (n=3) and normalized for FLAG–HA–Ago2 levels under the different
purified endosomes after treatment of cells with bafilomycin and
knockdown conditions, as shown in the representative anti-HA western blot.
(M) The fold repression levels for miR-122 reporter RL-Perf-miR-122 were
miR-122-containing EVs, then incubated them in vitro in a buffer at
calculated using a luciferase assay (n=4; unpaired Student’s t-test used for pH 5 with or without FCCP, a protonophore that can equalize the pH
analysis). Data are presented as mean±s.e.m. Representative blots have across biological membranes. We then followed the release of EV-
molecular mass markers indicated in kDa. *PRESEARCH ARTICLE Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914
Journal of Cell Science
Fig. 4. See next page for legend.
damage or a change in size and number of the endosomes in a time-dependent miRNA loading of endosomal Ago2 (Fig. 5F).
(Fig. S5A–C). With isolated endosomal fractions from an Optiprep gradient, we
These data thus substantiate the idea of pH-dependent release of found that the Ago2 present on endosomes was sensitive to
miRNA content from endosomes for subsequent reloading to Proteinase K even in the absence of any detergent, such as Triton
membrane-attached Ago2. This was further supported by an in vitro X-100. ApoE is a lipoprotein that is endocytosed after binding with
loading assay performed using endosome-associated Ago2. low-density lipoprotein receptors, and hence should be present
Incubation of single-stranded miR-122 with endosomes resulted within endosomes (Dekroon and Armati, 2002). ApoE was found to
9RESEARCH ARTICLE Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914
Fig. 4. Bafilomycin prevents acidification of endosomal compartments et al., 2014; Thery et al., 2002; Valadi et al., 2007). However, a major
and thus inhibits miRNA release from endosomes. (A) Diagram of the limitation in the EV and miRNA field is a deficit in the understanding
endocytic pathway, showing the gradual change in pH reported to occur in each
of the mechanistic aspect of functional internalization of EV-carried
of the compartments during endosomal maturation and the effects of treatment
with siRNAs targeting Rab5a (siRab5a), Rab7a (siRab7a) or RILP (siRILP), or miRNAs into recipient cells. Our findings shed light on this
with bafilomycin (BAF). (B) Presence of Ago2-associated internalized miRNAs mechanism of transfer of miRNA between mammalian cells.
in the indicated subcellular fractions of recipient cells. After co-culture with cells In this study, we have elucidated how a functional miRNA can be
expressing miR-122, cell lysates were prepared from recipient cells co- transferred from donor to recipient cells via EVs (Fig. 5I). The
transfected with siCon (control non-targeting siRNA), siRab5a, siRab7a or transferred miRNA does not affect the pre-existing endogenous
siRILP, and FLAG–HA–Ago2, and Optiprep density gradient ultracentrifugation
miRNA levels or their activities. We have observed that the EV-
was used to separate organelles based on their densities. Fractions positive for
endosomal (F2,3), lysosomal (F4,5,6) and ER (F7,8,9) marker proteins (as
derived miRNA is transferred primarily in a Dicer1-independent
depicted in Fig. 3B,C) were pooled together for immunoprecipitation of FLAG– manner in its mature single-stranded form. EV sorting of Ago2 has
HA–Ago2. Left: miRNAs associated with Ago2 in these fractions were been reported to stabilize miRNAs in exosomes or microvesicles
measured. n=3. Right: western blot for HA–Ago2 from cells treated with the (Beltrami et al., 2015; Lv et al., 2014; McKenzie et al., 2016).
indicated siRNAs. (C) A schematic representation of the effect of bafilomycin, Despite a previous claim that Ago2 is present in EVs, a recent
an inhibitor of V-type ATPase, on EV-derived miRNA loading on recipient cell publication has questioned the presence of protein components of
Ago2. Bafilomycin prevents acidification of endosomal compartments essential
for fusion of miRNA-containing internalized EVs with the endosomal membrane
miRNP machinery in mammalian cell-derived EVs (Jeppesen et al.,
and subsequent loading of internalized miRNAs (blue) onto Ago2 (red) located 2019). Therefore, it is most likely that single-stranded miRNAs may
on endosomal membrane. (D) Left: relative level of EV-derived miR-122 in travel from one cell to another via EVs in an Ago2-unbound form.
recipient HeLa cells treated with 25 nM bafilomycin. DMSO was added to We have observed that the EV-derived miRNA gets bound to the
control cells. n=3. Right: level of endogenous let-7a in recipient HeLa cells recipient cell Ago2 that drives the sustained function of the
treated or untreated with bafilomycin. n=3. (E) Western blot of LC3B (also transferred miRNA in mammalian cells.
known as MAP1LC3B) to show the increased cleavage of LC3B in cells treated
We have demonstrated that the uptake of the EV-containing
with bafilomycin compared to DMSO-treated control. β-actin is shown as a
loading control. Blot is representative of three experiments. (F) Repression miRNA is dependent on dynamin2, a GTP binding protein involved
levels of RL-Perf-miR-122 reporter relative to the control RL-Con, in cells treated in the endocytosis pathway. The involvement of the endocytic
with miR-122-containing EVs and either DMSO or bafilomycin. n=3; unpaired pathway in the uptake of EVs has been discussed in previous studies
Student’s t-test was used for analysis. (G) Relative levels of EV-derived miR- (Bonsergent and Lavieu, 2019; Joshi et al., 2020). We also conclude
122 in recipient cell membranes (left) and cytosol (middle) either treated with that miRNAs utilize the endosomal pathway to reach the ER of the
DMSO or bafilomycin. n=3. A representative western blot for cellular (GAPDH)
recipient cell. The compartmentalization of these miRNAs to the ER
and membrane (calnexin) markers in DMSO- and bafilomycin-treated cells is
shown (right). (H) Effect of bafilomycin on localization of EV-derived miR-122 in
suggests adaptability of the EV-derived miRNAs to elicit a
recipient cells. The recipient cells treated with bafilomycin and miR-122- repressive activity in the recipient cell. We have found that
containing EVs were subjected to Optiprep density gradient ultracentrifugation knockdown of the endosomal pathway effector protein RILP
followed by immunoprecipitation of Ago2 from the endosomal (F2,3) and ER leads to accumulation of the internalized miRNA in the recipient
(F7,8,9) fractions. Left: the Ago2-associated EV-derived miR-122 in the cell and defects in repression of target genes. Subcellular
endosomal and ER fractions in DMSO- and bafilomycin-treated recipient cells localization assays revealed a differential localization and Ago2
was quantified and normalized to the immunoprecipitated Ago2 levels. n=3.
Right: representative western blot showing HA–Ago2 in the different pooled
association of the transferred miRNAs upon knockdown of RILP.
fractions. (I) Relative levels of Ago2-associated EV-derived miR-122 in recipient These results explain why the internalized miRNA cannot be
cells. Top: miRNA associated with Ago2 immunoprecipitated from DMSO- or functional; it fails to reach the ER for its target mRNA interaction
bafilomycin-treated recipient cells was measured and compared against the and repressive activity in endosome maturation-defective cells. The
amount of miRNA found to be present with Ago2 from siRILP-treated cells. A increase in miRNP content could be due to lowered target-
relative decrease of Ago2 association of internalized miRNA was detected dependent miRNA turnover (Bitetti et al., 2018; de la Mata et al.,
when cells were treated with bafilomycin. n=3;unpaired Student’s t-test was
2015; Ghini et al., 2018) in RILP-depleted cells as a result of
used for analysis. Bottom: representative western blot for HA–Ago2 for both the
bafilomycin treatment and siRNA transfection sets. Data are presented as possible reduced miRNP–target RNA interaction.
mean±s.e.m. Representative blots have molecular mass markers indicated in The pH of the endosomal vesicles plays a role in release of cargo
kDa. *PRESEARCH ARTICLE Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914
Journal of Cell Science
Fig. 5. See next page for legend.
11RESEARCH ARTICLE Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914
Fig. 5. Release of miRNAs from EVs is dependent on the pH change in genistein (a clathrin-independent pathway inhibitor; Sigma); or 10 µM
endosomal compartments. (A) Schematic illustration of the procedure for chlorpromazine (a clathrin-dependent pathway inhibitor; Sigma) for 30 min
isolation of endosomes from lysates of HeLa cells treated with bafilomycin prior to EV treatment for 16 h.
(Sup, supernatant). (B) Western blot analysis of the indicated proteins in the
endosomal pellet and residual supernatant after ultracentrifugation and
siRNA and plasmid transfections
purification of the endosomes obtained from fractions 2–4 of an Optiprep
siRNAs against different endocytic pathway proteins (dynamin2, Rab5a,
gradient, as described in Fig. 3A,B, to confirm low levels of ER (calnexin) or
Rab7a and RILP), Dicer1 and Ago2, as well as control non-targeting
lysosomal (Lamp1) contamination in these endosomes. Blots are
representative of three experiments. (C) Schematic representation of the in
siRNA, were purchased from Dharmacon. siRNA transfection was
vitro assay utilized to understand the role of pH change in the release of performed using RNAi Max (Invitrogen) according to the manufacturer’s
internalized EV-entrapped miRNA. The endosomes were incubated with buffer instructions. HeLa cells were transfected with 15 pmoles of siRNA per well
of pH 5 in the presence or absence of FCCP, a protonophore. After the of a 24-well plate. The siRNA-transfected cells were split 48–72 h post
reaction, the endosomes were separated by ultracentrifugation as a pellet to transfection for proper knockdown. Details of the siRNAs used are shown in
separate any released components from the membranes. (D) Left: Table S5. For EV isolation, donor HeLa cells were transfected with 1 μg
quantification by RT-qPCR of EV-derived miRNA-122 released by endosomes pmiR-122 per well of a 6-well plate. pmiR122 plasmid, a plasmid encoding
after incubation with or without FCCP in pH 5 buffer. Ct values of miR122 in the the precursor miR-122, was described previously (Ghosh et al., 2013)
endosomal supernatant after incubation are shown (n=5; unpaired Student’s
t-test was used for analysis). Right: Ct values of endogenous let-7a in the
supernatant of the endosomes after incubation with or without FCCP in pH 5 EV isolation and characterization
buffer (n=3). (E) Time kinetics of release of miR-122 into the supernatant from HeLa cells transfected with pmiR-122 were split 24 h post transfection into
endosomes incubated with FCCP in pH 5 buffer, as determined using real-time 90 mm plates and incubated for 24 h at 70–80% confluency. These cell
PCR (n=4; unpaired Student’s t-test was used for analysis). (F) Time- culture supernatants were subjected to EV isolation as described previously,
dependent increase in miR-122 incorporation with endosome-associated with minor modifications (Thery et al., 2006). For EV isolation, cells were
Ago2 in FLAG–HA–Ago2-expressing HEK293 cells. Isolated endosomes, as grown in EV-depleted medium to prevent any background from exosomes
described in panel A, were incubated with single-stranded miRNA and lysed or EVs present normally in FBS. Briefly, the cell supernatant (EV-depleted
followed by immunoprecipitation of Ago2 to estimate the amount of associated growth medium that was used to culture the donor miR-122-expressing
miRNAs. Levels of Ago2 in immunoprecipitated materials, as shown in the HeLa cells) was centrifuged at 2000 g for 15 min to remove cell debris.
representative anti-HA western blot (bottom), were used for normalization. n=3. Next, the cell supernatant was collected and centrifuged at 10,000 g for
(G) After the in vitro pH-change assay was performed, western blotting of the 30 min. The supernatant that was obtained was passed through a 0.22μm
supernatant and pellet of the FCCP untreated and treated endosomes was filter unit to further clear it. This was followed by ultracentrifugation of the
performed for the endosomal proteins Alix and HRS. This was done to show supernatant at 100,000 g for 90 min. After ultracentrifugation, the pellet was
the absence of endosomal membrane contamination in the supernatant resuspended in EV-depleted growth medium and added to recipient cells.
recovered for miRNA estimation and to minimize the chance of membrane For characterization of EVs by AFM, the EVs were isolated on a 30%
contaminants accounting for the increase in miR-122 content in the
sucrose cushion, and the layer of EVs was further washed and pelleted with
supernatant. Blots are representative of two experiments. (H) Effect of
1× phosphate-buffered saline (PBS) by ultracentrifugation at 100,000 g for
Proteinase K treatment on endosome-associated Ago2. Western blotting for
90 min and resuspended in 1 ml PBS. Then, 5 µl of the EV suspension was
the indicated proteins shows the Proteinase K sensitivity of Ago2 protein
compared to endosomal luminal proteins like ApoE. Increasing concentration
placed onto a mica sheet (ASTM V1 Grade Ruby Mica from MICAFAB,
of Proteinase K was used alone or in the presence of Triton X-100 (TX100). Chennai, India) and dried for 15 min. The sample slides were then gently
Blots are representative of three experiments. (I) Diagram showing the washed with autoclaved MilliQ water to remove molecules that were not
mechanism of internalization of EVs and associated miRNAs into recipient firmly attached to the mica before being dried again. AAC mode AFM was
cells. The EVs utilize the endosomal pathway of the recipient cells for entry. performed using a Pico plus 5500 ILM AFM (Agilent Technologies, USA)
The gradual change in pH of the endosomal compartments from the early to with a piezoscanner maximum range of 9 µm. Microfabricated silicon
the late endosomes allow the fusion of internalized EVs with the endosomal cantilevers of 225 µm in length were used (Nano sensors, USA). Images
membranes thus releasing the miRNAs. The miRNAs are loaded onto the were processed by flatten using Pico view1.1 version software (Agilent
Ago2 proteins on endosomes in a Dicer1-independent manner before being Technologies, USA).
delivered to the ER to elicit a response in the recipient cell (MVB, multivesicular For NTA, EVs were resuspended in 1 ml PBS. EVs were diluted 10-fold,
body). Data are presented as mean±s.e.m. Representative blots have and 1 ml of diluted EVs was injected into the sample chamber of a
molecular mass markers indicated in kDa. *PRESEARCH ARTICLE Journal of Cell Science (2021) 134, jcs253914. doi:10.1242/jcs.253914
Copy number calculation from the top. The fractions were pooled accordingly for subsequent analysis
In order to calculate the copy number of miR-122 in the EVs isolated from of proteins and RNA. For immunoprecipitation of FLAG–HA–Ago2
miR-122 untransfected or transfected HeLa cells, we made a standard curve from the fractions, the pooled fractions were lysed with lysis buffer (20 mM
with known concentrations of synthetic miR-122. Following this, the Tris-HCl, pH 7.5, 150 mM KCl, 5 mM MgCl2, 1 mM DTT, 0.5% Triton X-
concentration of miR-122 in the EVs was quantified. We used the formula 100, 0.5% sodium deoxycholate and 1× PMSF) for 20 min at 4°C. The
reported previously (Simmonds, 2019): lysate was clarified by centrifugation at 16,000 g for 5 min and incubated
with anti-FLAG-M2 agarose beads overnight followed by RNA and protein
½fmols of RNA ðtotal average RNA yield in ng=200ngÞ analysis of the Ago2 associated with the beads.
6:02 108
copies per cell ¼ : Microscopic analysis of EV entry into recipient cells
number of cells
To detect the uptake of EVs by microscopy, the donor HeLa cells were
The yield of RNA was divided by the amount of RNA used for the PCR transfected with CD63–GFP expression plasmid encoding a GFP-tagged
reaction (200 ng). The copy number of miR-122 in recipient HeLa cells was tetraspanin. The supernatant of these cells was used for isolation of EVs
also calculated in the above manner using the same formula. from the cell culture medium by Exosome Isolation Reagent (Thermo
Scientific). The supernatant was cleared of cell debris by centrifugation at
Luciferase assay 2000 g for 30 min, followed by addition of the reagent, which was incubated
The plasmids pRL-Con (encoding RL control; RL-con) and pRL-Perf-miR- overnight at 4°C. On the following day, the supernatant was centrifuged at
122 (encoding the RL-Perf-miR-122 reporter) were a kind gift from 10,000 g for 60 min to pellet the EVs. These were added to the recipient cells
Dr Witold Fillipowicz (Friedrich Meisher Institute Basel, Switzerland). For seeded on coverslips in different conditions to observe under the
miRNA repression assays, 30 ng of Renilla luciferase (RL) reporter microscope. Cells were fixed using 4% paraformaldehyde for 20 min. For
plasmids (both pRL-Con and pRL-Perf-miR-122) with 300 ng of firefly detection of proteins, including β-tubulin, blocking and permeabilization
luciferase (FL) plasmid ( pGL3-FF) were co-transfected per well of a 12- was performed using 1% BSA, 10% goat serum and 0.1% Triton X-100 for
well plate. RL and FL activities were measured using a Dual-Luciferase 30 mins. The anti-β-tubulin (mouse) was used at 1:1000 dilution. Alexa
Assay Kit (Promega, Madison, WI) following the supplier’s protocol on a Fluor 568-conjugated anti-mouse IgG secondary antibody (Molecular
VICTOR X3 Plate Reader (PerkinElmer, Waltham, MA). The RL Probes) was used at 1:500 dilution. All the microscopic detection was done
expression levels for reporter and control were normalized against FL using a Zeiss LSM800 microscope followed by analysis using Imaris
levels. These normalized values were then used to calculate fold repression software.
as the ratio of normalized control RL values to normalized reporter RL
values. In vitro miRNA release assay
To see the effect of pH on release of miRNA from endocytic vesicles, an
Immunoprecipitation assay in vitro assay was used. For this, the recipient cells were incubated with
For IP of Ago2, HeLa cells were transfected with FLAG–HA-tagged Ago2 25 nM bafilomycin ( pre-treated for 1 h) along with miR-122-positive EVs
plasmid. Briefly, Protein G agarose beads (Invitrogen) or anti-FLAG-M2 for 16 h. The cell lysates were subjected to Optiprep density gradient
agarose beads (Sigma) were used for FLAG-tagged Ago2 IP. For HA, beads ultracentrifugation, and the endosomal marker-positive fractions 2, 3 and 4
were blocked with 5% BSA in lysis buffer [20 mM Tris-HCl, pH 7.5, were collected and diluted with buffer containing 78 mM KCl, 4 mM
150 mM KCl, 5 mM MgCl2, 1 mM dithiothreitol (DTT), 0.5% Triton MgCl2, 8.4 mM CaCl2, 10 mM EGTA and 50 mM HEPES ( pH 7.0). This
X-100, 0.5% sodium deoxycholate and 1× PMSF (Sigma)] for 1 h followed was followed by further purification of the endosomes by ultracentrifugation
by antibody incubation (1:50 dilution) for 4 h at 4°C. For IP reactions, HeLa at 133,000 g for 2.5 h. The endosome pellet was then resuspended in 100 µl
cells were lysed in lysis buffer for 20 min at 4°C. The lysates, clarified by buffer (20 mM Tris-HCl, pH 5, and 1 mM ATP) with or without 5 µM
centrifugation at 3000 g for 10 min, were incubated with HA antibody pre- FCCP. The endosomal suspension was then incubated for 30 min at 37°C.
bound Protein G Agarose beads or pre-blocked anti-FLAG M2 beads and The reaction was stopped by transferring the mixture to 4°C. This was
rotated overnight at 4°C. Subsequently, the beads were washed thrice with further diluted with a buffer containing 78 mM KCl, 4 mM MgCl2, 8.4 mM
1× IP buffer (20 mM Tris-HCl, pH 7.5, 150 mM KCl, 5 mM MgCl2 and CaCl2, 10 mM EGTA and 50 mM HEPES ( pH 7.0). Then, the mixture was
1 mM DTT), and separated into two halves for RNA and protein analysis ultracentrifuged at 133,000 g for 2.5 h to pellet the endosomes, and then the
from the bound Ago2 on the beads. All antibody information is in Table S4. supernatant was used to isolate RNA to see the amount of miRNA released
from the vesicles. The remaining supernatant and pellet were used for
Digitonin fractionation western blot analysis of marker proteins to rule out the contamination of
For digitonin fractionation, the recipient cells were pelleted and lysed using endosomes in the supernatant.
digitonin lysis buffer [10 mM Tris-HCl, pH 7.5, 25 mM KCl, 5 mM MgCl2,
1 mM CaCl2, RNase inhibitor (Applied Biosystems), 1 mM DTT, PMSF Proteinase K protection assay
(Sigma) and 50 µg/ml digitonin (Calbiochem)] for 10 min on ice. This was In order to detect whether the Ago2 protein is present on the outer or inner
followed by centrifugation at 2500 g for 5 min, and the supernatant (soluble surface of the endosomal membranes, a Proteinase K assay was used.
cytosolic fraction) was collected. The membrane pellet was washed again in Briefly, HEK293 cells were lysed, then loaded on an Optiprep density
the same lysis buffer without digitonin. The fractions were divided for gradient and ultracentrifuged as mentioned above. Then, the endosome-
protein analysis and RNA isolation using TriZol LS (Invitrogen). enriched fractions 2, 3 and 4 were collected. The endosomal fractions were
incubated with 0, 5, 10 or 20 ng/µl Proteinase K without Triton X-100, or
Journal of Cell Science
Optiprep density gradient ultracentrifugation with 20 ng/µl Proteinase K with 1% Triton X-100, for 30 min at 37°C. After
Optiprep (Sigma-Aldrich, USA) was used to prepare a 3–30% continuous the reaction, the solutions were subjected to methanol-chloroform protein
gradient in a buffer containing 78 mM KCl, 4 mM MgCl2, 8.4 mM CaCl2, precipitation. Then, western blotting for Ago2 was performed to see whether
10 mM EGTA and 50 mM HEPES ( pH 7.0) for separation of subcellular the Ago2 was present on the outside or inside of the endosomes.
organelles. HeLa cells were washed with PBS and homogenized with a
Dounce homogenizer in a buffer containing 0.25 M sucrose, 78 mM KCl, Polysome isolation
4 mM MgCl2, 8.4 mM CaCl2, 10 mM EGTA and 50 mM HEPES ( pH 7.0) For total polysome isolation, HeLa cells were lysed in a buffer containing
in addition to 100 μg/ml cycloheximide, 5 mM vanadyl ribonucleoside 10 mM HEPES ( pH 8.0), 25 mM KCl, 5 mM MgCl2, 1 mM DTT, 5 mM
complex (VRC; Sigma-Aldrich), 0.5 mM DTT and 1× PMSF. The lysate VRC, 1% Triton X-100, 1% sodium deoxycholate and 1× PMSF (Sigma)
was clarified by centrifugation at 1000 g for 5 min twice, then layered on top along with cycloheximide (100 μg/ml; Calbiochem). The lysate was cleared
of the prepared gradient. The tubes were centrifuged at 133,000 g for 5 h for at 3000 g for 10 min followed by another round of pre-clearing at 20,000 g
separation of the gradient, and ten fractions were collected by aspiration for 10 min at 4°C. The clarified lysate was loaded on a 30% sucrose cushion
13You can also read

















































