Extracellular vesicles as distinct biomarker reservoirs for mild traumatic brain injury diagnosis
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

doi:10.1093/braincomms/fcab151 BRAIN COMMUNICATIONS 2021: Page 1 of 14 | 1
Extracellular vesicles as distinct biomarker
reservoirs for mild traumatic brain injury
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
diagnosis
Kryshawna Beard,1,* Zijian Yang,2,* Margalit Haber,3 Miranda Flamholz,3
Ramon Diaz-Arrastia,3 Danielle Sandsmark,3 David F. Meaney4,5 and David Issadore4,6
* These authors equally contributed to this work.
Mild traumatic brain injury does not currently have a clear molecular diagnostic panel to either confirm the injury or to guide its
treatment. Current biomarkers for traumatic brain injury rely mainly on detecting circulating proteins in blood that are associated
with degenerating neurons, which are less common in mild traumatic brain injury, or with broad inflammatory cascades which are
produced in multiple tissues and are thus not brain specific. To address this issue, we conducted an observational cohort study
designed to measure a protein panel in two compartments—plasma and brain-derived extracellular vesicles—with the following
hypotheses: (i) each compartment provides independent diagnostic information and (ii) algorithmically combining these compart-
ments accurately classifies clinical mild traumatic brain injury. We evaluated this hypothesis using plasma samples from mild
(Glasgow coma scale scores 13–15) traumatic brain injury patients (n ¼ 47) and healthy and orthopaedic control subjects (n ¼ 46)
to evaluate biomarkers in brain-derived extracellular vesicles and plasma. We used our Track Etched Magnetic Nanopore technol-
ogy to isolate brain-derived extracellular vesicles from plasma based on their expression of GluR2, combined with the ultrasensitive
digital enzyme-linked immunosorbent assay technique, Single-Molecule Array. We quantified extracellular vesicle-packaged and
plasma levels of biomarkers associated with two categories of traumatic brain injury pathology: neurodegeneration and neuronal/
glial damage (ubiquitin C-terminal hydrolase L1, glial fibrillary acid protein, neurofilament light and Tau) and inflammation (inter-
leukin-6, interleukin-10 and tumour necrosis factor alpha). We found that GluR2þ extracellular vesicles have distinct biomarker
distributions than those present in the plasma. As a proof of concept, we showed that using a panel of biomarkers comprised of
both plasma and GluR2þ extracellular vesicles, injured patients could be accurately classified versus non-injured patients.
1 Department of Pharmacology, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104, USA
2 Department of Mechanical Engineering and Applied Mechanics, University of Pennsylvania, Philadelphia, PA 19104, USA
3 Department of Neurology, University of Pennsylvania, Philadelphia, PA 19104, USA
4 Department of Bioengineering, University of Pennsylvania, Philadelphia, PA 19104, USA
5 Department of Neurosurgery, University of Pennsylvania Perelman School of Medicine, Philadelphia, PA 19104, USA
6 Department of Electrical and Systems Engineering, University of Pennsylvania, Philadelphia, PA 19104, USA
Correspondence to: David F. Meaney, PhD
Department of Bioengineering, University of Pennsylvania
210 33rd Street, Philadelphia, PA 19104, USA
E-mail: dmeaney@seas.upenn.edu
Correspondence may also be addressed to: David Issadore, PhD.
E-mail: issadore@seas.upenn.edu
Received December 18, 2020. Revised May 14, 2021. Accepted May 20, 2021. Advance Access publication July 8, 2021
C The Author(s) (2021). Published by Oxford University Press on behalf of the Guarantors of Brain.
V
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted reuse,
distribution, and reproduction in any medium, provided the original work is properly cited.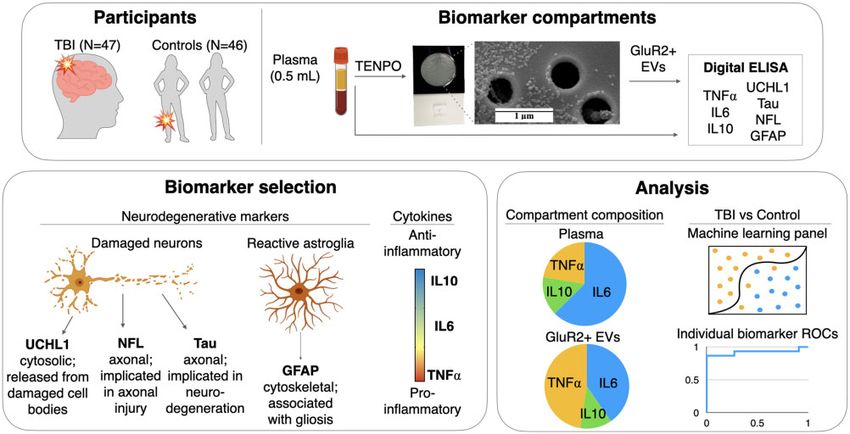
2 | BRAIN COMMUNICATIONS 2021: Page 2 of 14 K. Beard et al.
Keywords: traumatic brain injury; biomarkers; extracellular vesicles; machine learning
Abbreviations: AUC ¼area under the receiver operating characteristic curve; EV ¼extracellular vesicles; GFAP ¼glial fibrillary
acid protein; GluR2 ¼glutamate ionotropic receptor AMPA type subunit 2; IL6 ¼interleukin-6; IL10 ¼interleukin-10; mTBI ¼mild
traumatic brain injury; NFL ¼neurofilament light; ROC ¼receiver operating characteristic; SIMOA ¼single-molecule array;
TENPO ¼track etched magnetic nanopore; TNFa ¼tumour necrosis factor alpha; UCHL1 ¼ubiquitin C-terminal hydrolase L1
Graphical Abstract
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
Introduction individual patient outcomes and developing new treat-
ments has generated great interest in recent years.
Although most individuals who experience a mild trau- However, identifying sufficiently sensitive and specific
matic brain injury (mTBI) recover within weeks after the biomarkers has been confounded by the particularly dy-
injury, a significant number of patients suffer from per- namic and heterogeneous nature of TBI. Each TBI results
sistent symptoms that include headaches, cognitive in a unique combination of initial tissue damage and sec-
changes and mood disturbances for months afterward.1–3 ondary pathology including vascular dysfunction,6,7 axon-
Conventional approaches to evaluate mTBI have focussed al injury8,9 and inflammation10 that evolve following the
on currently known hallmarks of moderate-to-severe injury.11–13 These distinct aspects of an individual’s
brain damage, such as clinical assessment using the TBI—each of which maps to multiple potential bio-
Glasgow Coma Scale, macroscale lesions visualized with markers in the blood—is considered key to a patient’s
CT imaging and circulating neurodegenerative markers.4 possible recovery or progression to behavioural and cog-
Unfortunately, these methods lack the sensitivity and spe- nitive deficits.14,15 Moreover, profiles of biomarkers in
cificity needed to clinically characterize milder injuries, to the blood are dynamic, originating from both the initial
identify patients who are likely to have persistent symp- tissue damage and the multiple secondary pathologies
toms in the time following mTBI, and to guide each pa- that develop over time after the injury.14,16,17 The com-
tient to a personalized, effective therapy.5 Because plexity of biomarker expression following an injury
adequate biomarkers are lacking, the identification of results in diagnostics measurements that can be challeng-
mTBI patients in need of intervention remains mainly ing to interpret.
limited to monitoring for and treating post-concussive To gain a more comprehensive assessment of mTBI,
symptoms as they arise rather than treating the underly- many researchers have shifted their attention away from
ing pathology much earlier in the course of the disease, measurements of single biomarkers to measurements of
when therapies are more likely to be effective. biomarker panels, where each constituent biomarker can
Biomarker discovery to accurately diagnose and classify be chosen to assess a different aspect of the patient’s
an individual’s TBI into categories for improving TBI.18 For example, combined analysis of circulating glial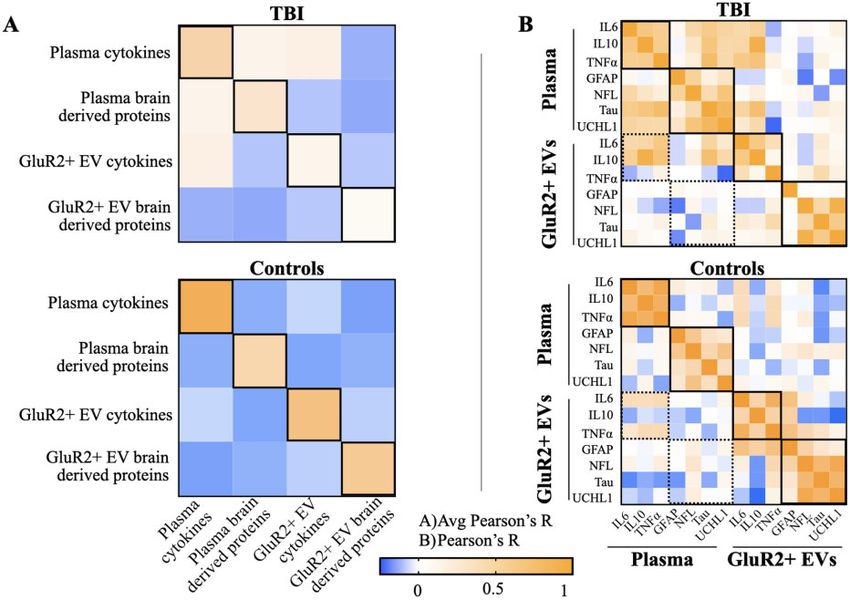
Distinct biomarkers for brain injury diagnosis BRAIN COMMUNICATIONS 2021: Page 3 of 14 | 3
fibrillary acidic protein (GFAP), an astrocyte-derived surface marker, from plasma using a nanomagnetic chip,
intermediate filament protein, and ubiquitin C-terminal and analysing RNA cargo we could identify RNA signa-
hydrolase L1 (UCHL1), a neuronal cytosolic protein, ac- tures that accurately classified the injury, including its
curately identifies injury severity and CT scan lesions in presence, severity, history of previous injuries and tim-
clinical TBI. While this assay—the Banyan Brain trauma ing.37–39 However, this work was limited to the RNA
indicator test—has demonstrated promise as a TBI diag- cargo of EVs, and did not incorporate known biomarkers
nostic for more severe injuries, such biomarkers have not of neuronal and glial cell damage or inflammation pack-
yet been identified that reliably classify underlying TBI aged in EVs.35
endophenotypes or predict patient outcomes after In this study, we combined conventional assessment of
mTBI.19 Other proposed TBI biomarkers have potential TBI-associated biomarkers in plasma with our approach
to directly assess specific underlying TBI pathologies. of acquiring molecular information from brain-derived
These include neurofilaments,20,21 a major cytoskeletal
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
EVs. Our main purpose was 2-fold: to determine if EVs
component of neuronal axons, and Tau, a cytoskeletal and plasma biomarker proteins represented independent
protein whose phosphorylation and aggregation are hall- information for mTBI diagnostic use, and to evaluate the
marks of neurodegenerative conditions.22,23 Dysregulated relative effectiveness of applying this approach on a set
central and peripheral immune cell function following of mTBI patients. We used our TENPO technology to en-
TBI results in the release of cytokines, chemokines and rich for brain-derived EVs and leveraged an existing
complement components that may provide an assessment ultrasensitive digital enzyme-linked immunosorbent assay
of inflammation, a key driver of neurologic deficit post- (ELISA) technique, Single-Molecule Array (SIMOA), to
TBI.24,25 accurately determine common protein biomarker levels in
Extracellular vesicles (EVs) have generated particular these two compartments. We demonstrate that the inde-
interest for multiplexed TBI diagnostics. EVs are nano- pendence of molecular information stored in the GluR2þ
scale vesicles ranging from 100 to 1000 nm26 formed EVs and in plasma allows for the development of a mul-
through a variety of mechanisms including plasma mem- tianalyte approach to mTBI diagnosis. In this work, we
brane budding or the fusion of multivesicular bodies use a machine learning algorithm developed using bio-
(MVBs) to the cellular membrane for release into the markers from both compartments as a proof-of-concept
extracellular space.27 EVs possess surface proteins derived of this approach, and illustrate its ability to successfully
from the parent cell, and cargo (proteins, mRNA, discriminate mTBI patients from control subjects.
miRNAs) within the vesicle lumen that reflect the status
of their cells of origin, and that, when transferred to re-
cipient cells, can act as agents of cell–cell communica- Materials and methods
tion.28–30 EVs are emerging as a promising complement
to plasma-derived biomarkers, as they contain cargo that Study design, participants and
may play direct roles in TBI pathology, and contain sur-
face proteins that allow brain-derived EVs to be isolated
sample collection
from the blood. EVs and their cargo also provide a Owing to the subtle nature of the physical injury in
work-around to the impracticality of brain tissue biopsy mTBI, we hypothesized that algorithmically combining
by crossing the blood–brain barrier into CSF,31 peripheral biomarkers from both brain-derived EVs and plasma
circulation,32 and other bodily fluids making them easily could result in more sensitive and specific discrimination
accessible CNS biomarkers for monitoring TBI progres- of mTBI patients from controls than that of any individ-
sion.33,34 Moreover, EVs are shed by both healthy and ual biomarker, or any one biomarker compartment. To
degenerating cells, providing a broader view into the mo- test this hypothesis, we obtained human plasma samples
lecular processes that occur within a tissue or organ. The from TBI subjects admitted to an urban, academic Level
combination of information extracted from the EVs of 1 trauma centre (University of Pennsylvania’s Penn
injured, but not necessarily degenerating, neurons with Presbyterian Medical Center) following a head impact—
neuronal biomarkers and inflammatory mediators could representing the diversity of injury types encountered in
lead to accurate classifications and prognoses of patients the clinic, including assault, road traffic incidents, and
with mTBI. falls—as well as healthy control and orthopedically
On their own, the diagnostic potential of EVs has been injured participants yielding a study size consistent with
shown in military personnel, where circulating exosome- previous experiments (Fig. 1A).38 Our blood-based assess-
packaged Tau and IL10 levels are elevated with mTBI ment of mTBI included a panel of neuronal and glial cell
and correlate with post-concussive and post-traumatic damage biomarkers [UCHL1, neurofilament light (NFL),
stress disorder symptoms.35 Other studies have found Tau and GFAP] and key drivers of inflammation [inter-
variations in EV microRNA concentration after TBI.36 In leukin-6 (IL6), interleukin-10 (IL10) and tumour necrosis
previous work, we showed that by enriching brain associ- factor alpha (TNFa)] quantified in both plasma and with-
ated EVs from plasma, which expressed the glutamate in brain-derived EVs expressing GluR2 (Fig. 1B). We
ionotropic receptor AMPA type subunit 2 (GluR2) then used these data to investigate protein distribution
4 | BRAIN COMMUNICATIONS 2021: Page 4 of 14 K. Beard et al.
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
Figure 1 Project workflow. (A) Samples were obtained from subjects sustaining TBIs through a variety of mechanisms and from a
combination of orthopaedic injured and healthy controls. One 500 ml aliquot of plasma from each subject was used to isolate brain-derived
EVs based on their expression of GluR2 using our nanofluidic platform, TENPO. Lysate from GluR2þ EVs and a second 500 ml aliquot of
plasma were subjected to digital ELISA assessment. (B) Biomarkers were selected based on known or emerging role in neuronal (UCHL1,
NFL, Tau) or astrocyte (GFAP) pathology, or on their roles in the spectrum of inflammatory function (TNFa, IL6, IL10). (C) Analyses served
two purposes: comparison of biomarker distribution in plasma and in GluR2þ EVs, and the discrimination of TBI and control subjects. A
machine learning approach was used to combine the multiplexed data into biomarker panels for comparison with the performance of
individual biomarker ROC curves and panels of biomarkers from each compartment alone.
across the two compartments and to evaluate mTBI-asso- were incarcerated. For this study, we analysed plasma
ciated changes in biomarker concentration and signatures samples collected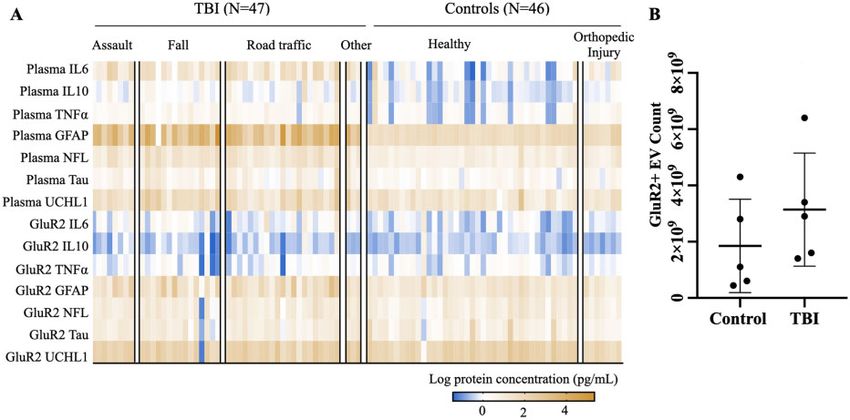
Distinct biomarkers for brain injury diagnosis BRAIN COMMUNICATIONS 2021: Page 5 of 14 | 5
process samples, and results in higher EV recovery from run using a 1:4 dilution. GluR2þ EV lysates were diluted
each sample than other isolation methods.37 EVs isolated in the Quanterix-provided sample diluent at a 1:10 ratio.
with the same GluR2þ TENPO assay we use in this For Neurology 4Plex analysis, 35 ml of EV lysate was
work were visualized with scanning electron microscopy combined with 315 ml of Neurology 4Plex sample diluent.
(repeated for this study in Supplementary Fig. 1), probed For cytokine analysis, 25 ml of EV lysate was combined
for canonical EV markers, and validated with isotype with 225 ml of Cytokine 3Plex sample diluent. 110–120 ml
control antibody in accordance with standards devised by of plasma and quality control samples were loaded into
the International Society of Extracellular Vesicles40 during sample plates and run at 4 dilution. The full volume
the assay’s initial development.37–39 (315 or 225 ml) of diluted the EV lysate was loaded into
In previous work, we successfully used TENPO to iso- sample plates and run neat. The results were corrected
late brain-derived EVs based on their expression of using the 10 or 4 dilution factor after analysis. To
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
GluR2 from plasma obtained from mouse models of TBI avoid measurement biases, GluR2þ EV samples and
and human clinical TBI patients.38 The device consists of plasma were run on the same plate with their respective
track-etched polycarbonate membranes with 600 nm SIMOA run settings.
pores (Whatman, Millipore Sigma) coated with a 200 nm
layer of permalloy (Ni80Fe20) and 30 nm layer of gold Statistical analysis of single
(Kurt Lesker PVD-75; Singh Nanofabrication Facility,
University of Pennsylvania) embedded in layers of mois- biomarkers
ture-resistant polyester film (McMaster-Carr, 0.00400 We began our analysis by comparing biomarker levels
thick, Elmhurst, Illinois) and solvent-resistant tape between mTBI and control subjects, and assessing their
(McMaster-Carr, Elmhurst, Illinois). The reservoir for ability to discriminate the two groups. We used one-way
inputting the sample was laser cut from a cast acrylic ANOVA (95% confidence intervals) to assess the effects
sheet (McMaster-Carr), and TENPO’s polydimethylsilox- of specific injury/control groups on biomarker expression,
ane output was connected to a negative pressure supply and evaluated the correlation between biomarker levels
(Programmable Syringe Pump; Braintree Scientific, and age. To visualize the spread of the data across indi-
Braintree, Massachusetts). To isolate brain-derived EVs viduals, we used the log-transformed values of biomarker
from plasma, we incubated 500 ml of subject plasma, first measurements to plot heatmaps (Fig. 2) and scatter plots
with biotin anti-human, mouse GluR1/GluR2 antibody (Fig. 3). For scatter plots, P-values were calculated using
(Bioss, Woburn, Massachusetts) and then with anti-biotin Student’s t-test with log-transformed data. Correlations
ultrapure microbeads (Miltenyi Biotec, Germany) at room between biomarkers were calculated using Pearson correl-
temperature with shaking, each for 20 min. After incuba- ation. To assess each biomarker’s ability to discriminate
tion, we loaded each sample into the reservoir of a mTBI from control groups, we calculated receiver operat-
TENPO device and ran it through the chip using a pro- ing characteristic (ROC) curves and reported the area
grammable syringe pump (Braintree). Captured EVs were under the curve (AUC) for each. To statistically compare
lysed using 500 ml 1% SDS in PBS as this completely sol- the performance of each biomarker, we quantified the
ubilizes even membrane-bound proteins,41 inactivates standard deviation for each biomarker’s AUC calculation
most cellular proteases,42 and is more concentrated than using bootstrapping, analysing a random subset (90% of
the 0.025% minimum shown to achieve vesicle lysis.43 the total sample size) 10 times with replacement. We
We also found this lysis condition yielded higher protein compared the AUCs of each individual biomarker with
levels than lysis with RIPA buffer containing 1% SDS our machine learning model’s AUC via Student’s t-test
(Supplementary Fig. 2). We diluted the lysate 1:10 to re- function while taking into account the correlation be-
duce the SDS concentration to 0.1% before running the tween the AUCs that is induced by the paired nature of
assay. Lysates were stored at 80 C until dilution and data.44 To account for variability in biomarker yield
further analysis. To avoid measurement biases, a combin- from TENPO-isolated EVs and centrifuged plasma, we
ation TBI and control subject plasma EVs were obtained reported biomarker expression as the relative abundance
during each run. of each biomarker in its given category (cytokine or
brain-derived protein). To this end, we summed the pa-
Single-Molecule Array digital ELISA tient or control average pg/ml levels of plasma IL6, IL10
measurement and TNFa and then divided the pg/ml level of each indi-
vidual cytokine by this total. We repeated this process
EV lysates and plasma samples were analysed using the with EV-packaged biomarkers, and for the brain-derived
Neurology 4Plex (A) and Cytokine 3Plex (A) Single- proteins.
Molecule Array kits (SIMOA; Quanterix, Billerica,
Massachusetts). Five hundred microlitre of plasma sam-
ples and quality controls provided by the kit were
Machine learning analysis
thawed on ice and spun at 10 000 RCF for 5 min. We used a machine learning-based approach to algorith-
Supernatant from this spin was used for plasma analysis mically determine a biomarker signature that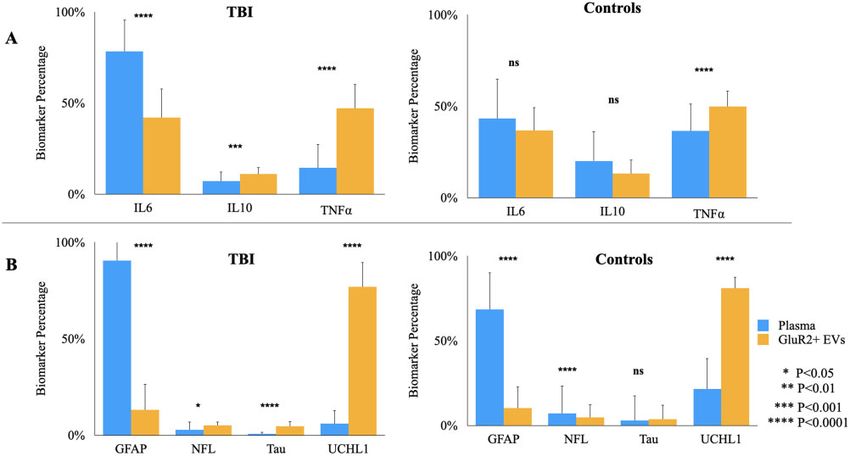
6 | BRAIN COMMUNICATIONS 2021: Page 6 of 14 K. Beard et al.
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
Figure 2 Expression of brain-derived proteins and cytokines is heterogeneous across TBI and controls in both plasma and
GluR21 EV compartments. (A) Log-transformed biomarker levels plotted in heat map. Columns represent subjects, each arranged
within respective TBI or control types by increasing age. (B) Number of GluR2þ EVs isolated from 0.5 ml plasma from N ¼ 5 TBI and N ¼ 5
control subjects.
Figure 3 Mild TBI is associated with elevations in both brain-derived proteins and cytokines in plasma and GluR21 EVs.
Scatter plots of mean log biomarker values and standard deviation as error bars. Calculation of P-values using student’s t-test were done using
log-transformed data. AUCs were generated using raw values.Distinct biomarkers for brain injury diagnosis BRAIN COMMUNICATIONS 2021: Page 7 of 14 | 7
discriminates mTBI from control subjects, using protein Pennsylvania and particle count results were corrected
concentrations from both plasma and GluR2þ EVs. Our with the dilution factor. Student’s t-test was used to as-
machine learning approach consisted of two stages. In sess differences in particle count across the groups.
the first, we used one set of patient (n ¼ 30) and control
(n ¼ 31) samples for feature selection and model training. Data availability
To perform feature selection and train our model, we
first performed Least Absolute Shrinkage and Selection Data were generated at the School of Engineering and
Operator (LASSO) to attain a biomarker panel. To miti- Applied Science at the University of Pennsylvania and
gate the effect of overfitting, we averaged the predictive Penn Presbyterian Medical Center. Data supporting the
values of an ensemble of five classifier algorithms—K- findings of this study are available from the correspond-
ing author upon request.
Nearest-Neighbors, SVM, Linear Discriminate Analysis,
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
Logistic Regression and Naive Bayes—instead of relying
on a single classifier.41 Additionally, we applied a boot-
strapping method that averages the predictions of mul- Results
tiple subgroups of the training set to diminish the effects
of outlier data. We performed initial evaluation of the
Participant demographics
performance of the machine learning panels using 5-fold To test our hypothesis that algorithmic combination of
cross validation. In the second stage of our machine biomarker data yields a more accurate mTBI diagnostic,
learning analysis, we validated the model developed in we collected plasma and EV samples from GCS mild
training phase with a user-blind, independent set of pa- (13–15) clinical TBI patients (n ¼ 47; Fig. 1). The study
tient (n ¼ 15) and control (n ¼ 11) subjects. Testing was design also included a control group (n ¼ 46) consisting
done only once to avoid the possibility of overfitting our of both healthy age-matched and orthopaedic injured
model to the test set data. The classifier model was controls to assess the specificity of blood and EV-pack-
implemented in Python and LASSO was carried out using aged biomarkers to mTBI. Although controls and mTBI
standard machine learning packages in Matlab 2017a. Of subjects were of similar ages (mean ¼ 36 years 6 16
the 93 total subjects assessed, 61 were used during ma- TBI, 614 controls), there were 20% more males in the
chine learning training. Of the 32 subjects later obtained mTBI than in the control group (Table 1).
during user-blind assessment of our machine learning al-
gorithm, 6 were excluded for incomplete biomarker data. Plasma- and brain-derived GluR21
To assess the performance of our multi-analyte ap-
proach, we also generated biomarker panels obtained
vesicles display variable biomarker
from plasma brain-derived proteins, plasma cytokines, distribution across individuals
GluR2þ EV brain-derived proteins and GluR2þ EV cyto- To visualize the spread of the data across individuals, we
kines. For statistical analysis, we randomly selected 90% first plotted log values of each biomarker across TBI and
of the total test set to evaluate the variances of our pre- control subjects (Fig. 2A). For the mTBI group, coeffi-
diction, and repeated this approach 10 times to obtain an cient of variance (CV) values for plasma biomarker levels
average AUC and standard deviation across panels. We ranged from 31% for GFAP, to 120% for TNFa. In the
compared the average AUC from each panel with our control subjects, CV values for plasma biomarkers ranged
original machine learning panel using t-tests while taking from 27% for IL10, and 140% for GFAP. Levels of
into account the correlation between AUCs that is plasma biomarkers for mTBI subjects were not signifi-
induced by the paired nature of data.44 cantly affected by injury type (Supplementary Table 1;
ANOVA; P > 0.65 across all biomarkers), nor did they
Scanning electron microscopy correlate with age (Supplementary Table 5). We thus
We imaged captured GluR2þ EVs by fixing them to the
TENPO membrane using 2.5% glutaraldehyde, 2.0% Table 1 Descriptive characteristics of traumatic brain
injury patient and control subjects; mean 6 SD or N
paraformaldehyde in 0.1 sodium cacodylate buffer, pH
(%).
7.4. Images were taken at the Cell and Developmental
Biology Microscopy Core at University of Pennsylvania. Characteristics TBI Control TBI
Control
Nanotracking analysis Machine learning group Training Training Test Test
N 30 31 17 15
500 ml of plasma from 5 TBI and 5 control subjects was Demographics
used to isolate GluR2þ vesicles with TENPO, and eluted Age, mean 6 SD (years) 35 6 14 36 6 16 44 6 18 28 6 8
in 500 ml of PBS. Vesicles were diluted 1:1000 in molecu- Male gender, N (%) 83 53 61 60
lar grade water. Nanotracking analysis was performed at GCS mean 14.4 N/A 14.5 N/A
Positive CT, N (%) 57 N/A 72 N/A
the Extracellular Vesicle Core facility at University of8 | BRAIN COMMUNICATIONS 2021: Page 8 of 14 K. Beard et al.
collapsed all injury types into a single mTBI group for significantly altered in the GluR2þ EV compartment fol-
our subsequent analysis of plasma biomarker measures. lowing mTBI. Additionally, biomarkers significantly ele-
In the control group, orthopaedic controls exhibited sig- vated in mTBI plasma also displayed the highest AUCs
nificantly higher levels of plasma GFAP compared to the for discriminating mTBI and control subjects (plasma
control mean (Supplementary Table 2; ANOVA; GFAP ¼ 0.91; GluR2þ EV GFAP ¼ 0.70; plasma
P < 0.05; Dunett correction for multiple comparisons), NFL ¼ 0.86; plasma IL6 ¼ 0.86). AUCs for all other bio-
but there were no additional effects of control type or markers were likely affected by the heterogeneous distri-
age on plasma biomarker levels. We thus consolidated bution of the dataset.
control subjects into a single group for plasma biomarker
analyses. Brain-derived EVs and plasma
Our analysis and visualization of the data also
possess distinct protein
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
revealed, similar to plasma biomarkers, that proteins
packaged in GluR2þ EVs are also expressed heteroge- compositions
neously across individual subjects (Fig. 2A). For the
Until this point, we analysed the performance of single
mTBI group, CV for GluR2þ EV-packaged biomarkers
proteins, regardless of its originating in plasma or
ranged from 45% for IL6 to 140% for Tau. We found
GluR2þ EVs, to best discriminate between mTBI and
neither injury/control type nor age has an effect on the
control samples. However, simply combining the best in-
expression of EV-packaged biomarkers (Supplementary
dividually high performing biomarkers would potentially
Tables 3–4; P > 0.38 (injury type, across all biomarkers);
overlook combinations of biomarkers that would better
P > 0.24 (control type, across all biomarkers)). Nor were
predict the presence/absence of mTBI. To evaluate distinc-
there any significant correlations between age and
tions in biomarker information across plasma and
GluR2þ EV biomarker levels for either group
GluR2þ EV compartments, we first assessed the distribu-
(Supplementary Table 6; P > 0.05 across all biomarkers
tion of biomarkers in each group. In the mTBI group,
for both groups). Therefore, we consolidated all injury
plasma and GluR2þ EVs displayed significantly different
types into a single injury group, and the two control
proportions of all measured cytokines (P < 0.001 across
types into a second group for analyses of these bio-
markers. Furthermore, we demonstrated that there is no all cytokines; Fig. 4A). Specifically, the relative abundance
significant difference in the number of EVs across TBI of IL6 was significantly elevated in plasma compared to
and controls (P > 0.05), eliminating the need to normalize GluR2þ EVs. Conversely, GluR2þ EVs contain signifi-
across EV count (Fig. 2B). cantly higher proportions of both IL10 and TNFa than
Once we consolidated our dataset, we hypothesized— plasma. The distribution of cytokines is also more bal-
based on other studies of mTBI biomarkers, and on the anced in GluR2þ EVs, with IL10 and TNFa making up
rate of CT scan abnormality of our mTBI subjects (Table similar proportions, while in plasma, cytokine distribution
1)—that mTBI subjects would exhibit significant eleva- is skewed with the relative abundance of IL6 dwarfing
tions in conventionally-studied plasma biomarkers relative that of IL10 and TNFa by 11- and 5-fold respectively. In
to controls.19 To test this hypothesis, and to investigate contrast to the mTBI group, the control group displayed
whether mTBI was also associated with significant no significant differences in proportions of IL6 or IL10
changes to biomarker levels in the GluR2þ EV compart- across the two compartments (P > 0.05). In this group,
ment, we compared mean levels of individual biomarkers only TNFa abundance differed in plasma and GluR2þ
in both compartments (Fig. 3). EVs, showing a significant increase in the latter
Of the seven biomarkers measured, GFAP and IL6 (P < 0.0001). Lastly, in this group, distribution of the
were each significantly elevated in both plasma (GFAP three cytokines exhibits balance in both compartments,
P < 0.0001, 147-fold change compared to controls; IL6 each having similar proportions of IL6 and TNFa.
P < 0.01, 7-fold change relative to controls) and GluR2þ Like the cytokines, our analysis also revealed differen-
EVs (GFAP P < 0.001, 6-fold change compared to con- ces between plasma and GluR2þ EV distributions of the
trols; IL6 P < 0.01, 3-fold change relative to controls) in four brain-derived proteins (Fig 4B). For both mTBI and
mTBI. Of the remaining brain-derived proteins, plasma control subjects, abundance of three of the four (GFAP,
levels of NFL, Tau and UCHL1 were all significantly ele- NFL and UCHL1) display significant differences in
vated in the mTBI group. (NFL P < 0.0001, 3-fold plasma compared to GluR2þ EVs, and Tau abundance is
change relative to controls; Tau P < 0.01, 4-fold change significantly elevated in GluR2þ EVs in the mTBI group
relative to controls; UCHL1 P < 0.0001, 3-fold change (P < 0.05 across all brain-derived proteins; Fig. 4B). In
relative to controls). GluR2þ EV levels of NFL, Tau and both groups, the distributions of these proteins are
UCHL1 were unchanged following mTBI. Of the two uniquely skewed in each compartment; in plasma, GFAP
remaining cytokines, both were significantly elevated in abounds (fold increases of 31, 136 and 15 relative to
plasma in the mTBI group (IL10 P < 0.001, 2-fold change NFL, Tau and UCHL1 respectively for mTBI group; fold
relative to controls; TNFa P < 0.001, 1-fold change rela- increases of 10, 24 and 3 relative to NFL, Tau and
tive to controls). Neither of these cytokines were UCHL1 respectively for controls) while GluR2þ EVs areDistinct biomarkers for brain injury diagnosis BRAIN COMMUNICATIONS 2021: Page 9 of 14 | 9
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
Figure 4 Plasma- and brain-derived EVs possess distinct protein composition that are each altered by TBI. Mean levels of each
biomarker were totalled across individuals to determine the relative percentage of each (A) cytokine and (B) brain-derived protein in plasma
and GluR2þ EVs. Error bars represent SD calculated by propagation of uncertainty. T-tests were performed to assess statistically significant
differences in biomarker levels across compartments.
dominated by UCHL1 (fold increases of 6, 15 and 17 within plasma (avg Pearson’s R ¼ 0.50) and within
compared to GFAP, NFL and Tau respectively for mTBI GluR2þ EVs (avg Pearson’s R ¼ 0.35) than across plasma
group; fold increases of 8, 16 and 22 relative to GFAP, and GluR2þ EV compartments (avg Pearson’s
NFL and Tau respectively for controls). R ¼ 0.0065). As with the mTBI group, cytokine levels are
most correlated within plasma (avg Pearson’s R ¼ 0.80)
Biomarker levels correlate more in the control group. In contrast to the mTBI group,
cytokine levels are also more correlated within EVs (avg.
within than across plasma- and Pearson’s R ¼ 0.57) than across plasma and EVs (avg.
brain-derived EV compartments Pearson’s R¼0.20). Like the mTBI group, pools of brain-
derived biomarkers are more distinct: levels of these pro-
To develop a combinatorial method for discriminating
teins are most correlated in EVs and within the plasma
TBI from control subjects, each biomarker should hold
compartment, and least correlated across compartments
the potential to contribute unique information about each
(avg. Pearson’s R ¼ 0.55, 0.42, 0.0065, respectively). Our
patient’s TBI. To assess this, we calculated three sets of
results showed that different groups of biomarkers had
correlation values for each biomarker category (cytokines
very little correlation across compartments though levels
or brain-derived proteins): correlations within plasma, correlated within each group.
within the GluR2þ EVs, and across these two compart- Given the independence of information collected from
ments (Fig. 5A). plasma and plasma-derived EVs, we next assessed
We found levels of cytokines (IL6, IL10 and TNFa) whether we could develop a machine learning-based clas-
and brain-derived proteins (GFAP, NFL, Tau and sifier of TBI using the complimentary biomarker informa-
UCHL1) correlated more with each other within the tion contained within each. By first separating out a
same compartment than between the plasma and EVs group of samples to train our model and using a separate
compartments (Fig. 5B). For mTBI subjects, cytokine lev- group to test its accuracy, we found the combination of
els are most correlated within the plasma (avg. Pearson’s information from plasma-derived EVs and plasma showed
R ¼ 0.61). In comparison, cytokine correlations within the high accuracy (AUC ¼ 0.913, Accuracy ¼ 0.825).
EV compartment (avg Pearson’s R ¼ 0.38) and across However, this combination of measures was only margin-
plasma and EVs (avg Pearson’s R ¼ 0.41) were similar. ally better than a single measure of plasma GFAP or IL-6
Levels of brain-derived biomarkers are more correlated (Supplementary Fig. 3) and did slightly better in10 | BRAIN COMMUNICATIONS 2021: Page 10 of 14 K. Beard et al.
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
Figure 5 Biomarker levels are uncorrelated across plasma and brain-derived EV compartments. Pearson’s correlation
coefficients calculated for all possible combination of biomarkers. (A) Average R for each biomarker type (cytokines or brain-derived
markers) and for each compartment (plasma or GluR2þ EVs) were plotted in a heat map matrix for TBI patients and controls. Solid boxes
indicate average R for each biomarker type-compartment combination. (B) R for each biomarker comparison was plotted into heat map
matrices for both TBI patients and controls. Solid boxes indicate R for individual biomarkers of the same type (cytokines or brain-derived
proteins) within each compartment. Dashed boxes indicate R values for biomarkers of the same type, but of different compartments.
prediction of injury when compared to combination of of accurately detecting levels of biomarkers that often cir-
measures from plasma. However, removing a measure of culate at levels too low to detect with conventional tech-
Tau in the EVs significantly worsened the predictive nologies. We then demonstrate that algorithmically
power of the panel, implying the importance of including combining information from each biomarker compart-
an EV-based measure in a combined diagnostic ment accurately classifies mTBI (AUC ¼ 0.92,
(Supplementary Fig. 3). While the sample size in this Accuracy ¼ 0.885). The combined use of biomarkers of
study is not sufficient to fully develop and evaluate a ma- specific TBI pathologies analysed in the context of dis-
chine learning classifier, these results suggest its promise tinct biofluid environments from separate cellular pools is
in future clinical studies. a promising approach for developing a more comprehen-
sive assessment of the state of the injured and recovering
brain.
Discussion We began our analysis with measuring circulating levels
of brain-derived proteins GFAP, NFL, Tau and UCHL1,
Our study demonstrates that circulating brain-derived which demonstrated predictive power as individual bio-
EVs and plasma represent two distinct reservoirs of mo- markers similar to past studies of these biomarkers in
lecular information, the composition of each differentially mTBI.19,45 Since a sizable proportion of mTBI subjects in
altered by mTBI. We combined two technologies— this study (57%) sustained brain pathology observable
TENPO that can specifically enrich for GluR2þ EVs through CT scan, the significant elevations in plasma lev-
from plasma, and digital ELISA that can measure mul- els of UCHL1 and NFL (P < 0.0001 for each) were
tiple protein biomarkers with 100–1000 better sensitiv- expected.19 As mTBI results in few degenerating neu-
ity than conventional ELISA—to address the challenges rons,46 it is not surprising that we did not detectDistinct biomarkers for brain injury diagnosis BRAIN COMMUNICATIONS 2021: Page 11 of 14 | 11
significant elevations in Tau after mTBI. In contrast, re- following the initial mild injury, and can be further com-
active gliosis can be observed throughout the brain even plicated by repeated, periodic opening of the blood–brain
after mild injury,47 and as expected we found plasma barrier.55–58 These primed or activated neurons and glia
GFAP the most robust single biomarker to discriminate undergo subtler forms of cellular damage or distress as
mTBI subjects from controls (AUC ¼ 0.89). However, we they constitutively secrete exosomes, potentially as a
observed plasma GFAP levels were significantly elevated mechanism for clearing cellular debris as they recover.
in orthopaedic-injured controls and controls over 51 years We observed that GluR2þ EVs contain the same inflam-
(ANOVA; P < 0.046 and P < 0.0001, respectively) com- matory cytokines and markers of cell damage expressed
pared to the total control plasma GFAP mean. This find- by lesioned cells.59 Interestingly, we found UCHL1—a
ing, combined with observations that GFAP is released deubiquitinating enzyme—dominated the EV pool of
from other cell types of the body,48 may complicate brain-derived proteins (Fig. 4). As UCHL1 plays a neuro-
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
GFAP’s specificity to brain injury, especially if it is used protective role in brain injury by degrading reactive lipids
in isolation in polytrauma cases. We also observed sig- and misfolded proteins,60,61 the high relative abundance
nificant elevations in plasma levels of IL6, IL10 and of this protein in GluR2þ EVs in relation to the other
TNFa (P < 0.001 across all cytokines), and indeed, measured brain-derived proteins suggests GluR2þ EVs
plasma IL6 followed directly behind plasma GFAP in dis- may serve as a protein clearing system for the damaged
criminating mTBI (AUC ¼ 0.86). But the broad role that or distressed cells of the brain, a role for EVs already
cytokines play in mediating systemic trauma,49 immune demonstrated for other EV populations in other contexts
challenges50,51 and other neurological disorders52 may and cell types.62 Thus, our machine learning approach
limit the specificity of these biomarkers in plasma. combines molecular information from three different cate-
In our assessment of mTBI, we also included brain- gories: markers from a small number of severely dam-
derived EVs expressing GluR2, an appealing alternative aged, degenerating cells (plasma brain-derived markers),
to co-opting the circulating neurodegenerative markers markers of broad-scale inflammation, and markers origi-
typically associated with moderate-to-severe TBI for nating from a potentially less-damaged population of
mTBI diagnostics. On their own, the proteins in brain- brain cells (brain-derived EVs). By investigating what
derived EVs performed no better than those in plasma as downstream molecular targets GluR2þ EV-packaged
individual biomarkers, despite the IL6 and GFAP eleva- UCHL1 interacts with, such as other components of the
tions observed in this compartment relative to controls ubiquitin ligase system and potential degradation targets,
(P < 0.01 and P < 0.001, respectively). However, it was we can broaden our understanding of the role the
intriguing to see that protein concentration of the same GluR2þ EV population plays in TBI pathology, and our
biomarkers across the two compartments did not correl- potential pool of EV-associated biomarkers.
ate with each other. One potential explanation for this Though we demonstrate the orthogonality of molecular
result is that plasma levels of some biomarkers appeared information in plasma and brain-derived EVs, the ma-
from active degeneration processes in a small population chine panel we devised by algorithmically combining mo-
of cells, while the exosome derived measurements origin- lecular information across the two compartments did not
ate from a large population of largely intact neurons and significantly benefit mTBI diagnosis when compared to
glia responding to the mild mechanical trauma. Other the performance of plasma neurodegenerative markers
studies of EV-based biomarkers of TBI have similarly alone. Our analysis of brain-derived EVs was limited to
observed differences in EV-contained and plasma molecu- those expressing GluR2þ, and by expanding our ap-
lar cargo. In a study measuring time-dependent changes proach to mTBI biomarker development—from broaden-
in protein biomarkers within the total circulating EV ing the EV subtypes and cargo that we isolate to surveil
population and plasma, investigators found no correlation a more comprehensive set of cells affected by TBI, to
between the two compartments out to 5 days after advancing the technologies with which we measure and
injury.53 analyse this complex information—we can improve our
Although studies on EVs and their contents is only ability to diagnose mTBI, and to extend this approach to
emerging, the broader sampling of EV signatures from monitor mTBI outcome and identify accurate treatment
cells that do not later degenerate provides a new oppor- strategies. Since this work we have extensively optimized
tunity for understanding the consequences and recovery the TENPO protocol, incorporating sequential wash steps
processes of mild trauma to the brain. With the broad following vesicle capture which greatly reduce back-
disruption in blood–brain barrier integrity that occurs ground relative to relying on the chip’s small dead vol-
after mTBI,54 it is possible that plasma activates patho- ume to promote removal of unbound material.39
logic cascades in neurons and glia that do not later de- Additionally, while lysis with 0.1% SDS using RIPA buf-
generate, resulting in a cellular population that largely fer resulted in lower protein concentration than lysing
outnumbers actively dying or degenerating ones in mTBI with 1% SDS (Supplementary Fig. 2), it is possible that
that are not assessed with traditional plasma biomarkers. this resulted in some protein denaturing. Since this work,
In experimental models of concussion and in clinical we have improved lysis conditions to maximize protein
studies, degenerative changes can occur days to months yield and efficacy of biomarker detection. We now use12 | BRAIN COMMUNICATIONS 2021: Page 12 of 14 K. Beard et al.
phosphatase/protease inhibitor cocktail and are investigat- NS086090, K23 NS104239, and Department of Defense
ing non-denaturing reagents, both of which have been W81XWH-14-2-0176.
used in other recently published SIMOA studies of exo-
some protein expression.35,53
We are also exploring capturing circulating EVs derived Conflict of interest
from multiple brain cell types (neurons, astrocytes, micro-
glia, etc.), which, when combined with advancements in David Issadore is a founder and holds equity in Chip
downstream EV cargo analysis, maximizes the molecular Diagnostics, a start-up company developed out of his lab.
information at our disposal for these goals. Multianalyte No competing financial interests exist for the other authors.
approaches to disease diagnosis have already shown
promise in other fields, resulting in higher accuracy in
early detection and staging of cancer.63–65 For mTBI, as References
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
technology for isolating EVs from different populations 1. Rimel RW, Giordani B, Barth JT, Boll TJ, Jane JA. Disability
of distressed-but-not-dying brain cells evolves, we may caused by minor head injury. Neurosurgery. 1981;9(3):221–228.
improve our ability to identify pathologies like gliosis 2. Iverson G. Outcome from mild traumatic brain injury. Curr Opin
and brain endothelial cell dysfunction as they occur Pyschiatry. 2005;18(3):301–317.
3. Nelson LD, Temkin NR, Dikmen S, et al.; and the TRACK-TBI
across individual patients to better ‘grade’ the TBI.
Investigators. Recovery after mild traumatic brain injury in
Though we used digital ELISA in this study, a platform patients presenting to US level I trauma centers: A transforming re-
incompatible with point-of-care diagnostics, widespread search and clinical knowledge in traumatic brain injury (TRACK-
efforts to scale down these assays into portable platforms TBI) study. JAMA Neurol. 2019;76(9):1049–1059.
makes accessibility and clinical use of molecular diagnos- 4. Dadas A, Washington J, Diaz-Arrastia R, Janigro D. Biomarkers
in traumatic brain injury (TBI): A review. Neuropsychiatr Dis
tic more achievable.66 Our previous work also points to
Treat. 2018;14:2989–3000.
the promise of miRNA cargo for a more open-ended TBI 5. Levin HS, Diaz-Arrastia R. Diagnosis, prognosis, and clinical man-
assessment than is provided by known protein bio- agement of mild traumatic brain injury. Lancet Neurol. 2015;
markers, but combining EV protein cargo information 14(5):506–517.
with amplifiable EV-associated miRNA results in a 6. DeWitt DS, Prough DS. Traumatic cerebral vascular injury: The
wealth of potential opportunities to develop more accur- effects of concussive brain injury on the cerebral vasculature. J
Neurotrauma. 2003;20(9):795–825.
ate mTBI characterization. Advanced approaches to data
7. Kenney K, Amyot F, Haber M, et al. Cerebral vascular injury in
analysis such as machine learning coupled with improved traumatic brain injury. Exp Neurol. 2016;275(3):353–366.
understanding of the pathologic roles brain-derived EVs 8. Johnson VE, Stewart W, Smith DH. Axonal pathology in traumat-
play in mTBI progression broadens our potential to com- ic brain injury. Exp Neurol. 2013;246:35–43.
bine this wealth of information into meaningful molecular 9. Povlishock JT. Traumatically induced axonal injury: Pathogenesis
and pathobiological implications. Brain Pathol. 1992;2(1):1–12.
signatures to monitor and intervene in this insidious
10. Xiong Y, Mahmood A, Chopp M. Current understanding of neu-
neurologic condition. roinflammation after traumatic brain injury and cell-based thera-
peutic opportunities. Chin J Traumatol. 2018;21(3):137–151.
11. Kaur P, Sharma S. Recent advances in pathophysiology of trau-
Supplementary material matic brain injury. Curr Neuropharmacol. 2018;16(8):1224–1238.
12. Prins M, Greco T, Alexander D, Giza CC. The pathophysiology of
Supplementary material is available at Brain traumatic brain injury at a glance. Dis Model Mech. 2013;6(6):
1307–1315.
Communications online.
13. Kochanek PM, Berger RP, Bayir H, Wagner AK, Jenkins LW,
Clark RSB. Biomarkers of primary and evolving damage in trau-
matic and ischemic brain injury: Diagnosis, prognosis, probing
Acknowledgements mechanisms, and therapeutic decision making. Curr Opin Crit
Care. 2008;14(2):135–141.
We thank Erin Anderson (University of Pennsylvania, 14. McAllister TW. Neurobiological consequences of traumatic brain
Philadelphia, PA, USA) and Stephanie Yang (University of injury. Dialogues Clin Neurosci. 2011;13(3):287–300.
Pennsylvania, Philadelphia, PA, USA) for editorial feedback. 15. Zetterberg H, Blennow K. Fluid biomarkers for mild traumatic
brain injury and related conditions. Nat Rev Neurol. 2016;12(10):
563–574.
16. Gan ZS, Stein SC, Swanson R, et al. Blood biomarkers for trau-
Funding matic brain injury: A quantitative assessment of diagnostic and
prognostic accuracy. Front Neurol. 2019;10:446.
This work was supported by a grant from the Paul G Allen 17. Thelin E, Nimer FA, Frostell A, et al. A serum protein biomarker
Frontiers group (grant #12347), the New Jersey Commission panel improves outcome prediction in human traumatic brain in-
for Brain Injury Research, and grant W81XWH1920002 jury. J Neurotrauma. 2019;36(20):2850–2862.
18. Beard K, Meaney D, Issadore D. Clinical applications of extracel-
from the Department of Defense. Work in the authors’ la-
lular vesicles in the diagnosis and treatment of traumatic brain in-
boratory was also supported by the Pennsylvania jury. J Neurotrauma. 2020;37(19):2045–2056.
Department of Health. This work was also funded by 19. Diaz-Arrastia R, Wang KKW, Papa L, et al. TRACK-TBI
National Institute of Neurological Disorders and Stroke U01 Investigators. Acute biomarkers of traumatic brain injury:Distinct biomarkers for brain injury diagnosis BRAIN COMMUNICATIONS 2021: Page 13 of 14 | 13
Relationship between plasma levels of ubiquitin C-terminal hydro- functions: A position statement from the International Society for
lase-L1 and glial fibrillary acidic protein. J Neurotrauma. 2014; Extracellular Vesicles. J Extracell Vesicles. 2014;3:26913.
31(1):19–25. 41. Davies KD, Goebel-Goody SM, Coultrap SJ, et al. Long term syn-
20. Shaw G, Yang C, Ellis R, et al. Hyperphosphorylated NF-H is a aptic depression that is associated with GluR1 phosphorylation
serum biomarker of axonal injury. Biochem Biophys Res but not alpha-amino-3-hydroxy-4-isoxazolepropionic acid
Commun. 2005;336(4):1268–1277. (AMPA) receptor internalization. J Biol Chem. 2008;283(48):
21. Shahim P, Politis A, van der Merwe A, et al. Neurofilament light 33138–33146.
as a biomarker of traumatic brain injury. Neurology. 2020;95(6): 42. Lantz MS, Ciborowski P. Zymographic techniques for detection
e610–e622. and characterization of microbial proteases. Methods Enzymol.
22. Edwards G, Zhao J, Dash PK, Soto C, Moreno-Gonzalez I. 1994;235:563–594.
Traumatic brain injury induces Tau aggregation and spreading. J 43. Osteikoetxea X, Sodar B, Nemeth A, et al. Differential detergent
Neurotrauma. 2020;37(1):80–92. sensitivity of extracellular vesicle subpopulations. Org Biomol
23. Castellani RJ, Perry G. Tau biology, tauopathy, traumatic brain Chem. 2015;13(38):9775–0782.
injury, and diagnostic challenges. J Alzheimers Dis. 2019;67(2): 44. Hanley JA, McNeil BA. method of comparing the areas under re-
Downloaded from https://academic.oup.com/braincomms/article/3/3/fcab151/6317828 by guest on 03 December 2021
447–467. ceiver operating characteristic curves derived from the same cases.
24. Di Battista AP, Rhind SG, Hutchison MG, et al. Inflammatory Radiology. 1983;148(3):839–483.
cytokine and chemokine profiles are associated with patient out- 45. Bazarian JJ, Biberthaler P, Welch RD, et al. Serum GFAP and
come and the hyperadrenergic state following acute brain injury. J UCH-L1 for prediction of absence of intracranial injuries on head
Neuroinflammation. 2016;13(13):40. CT (ALERT-TBI): A multicentre observational study. Lancet
25. Ritzel RM, Doran SJ, Barrett JP, et al. Chronic alterations in sys- Neurol. 2018;17(9):782–789.
temic immune function after traumatic brain injury. J 46. Shultz SR, MacFabe DF, Foley KA, Taylor R, Cain DP. Sub-con-
Neurotrauma. 2018;35(13):1419–1436. cussive brain injury in the Long-Evans rat induces acute neuroin-
26. Kalluri R, LeBleu VS. The biology, function, and biomedical appli- flammation in the absence of behavioral impairments. Behav Brain
cations of exosomes. Science. 2020;367(6478):eaau6977. Res. 2012;229(1):145–152.
27. Tkach M, Théry C. Communication by extracellular vesicles: 47. Ren Z, Iliff J, Yang L, et al. ‘Hit & Run’ model of closed-skull
Where we are and where we need to go. Cell. 2016;164(6): traumatic brain injury (TBI) reveals complex patterns of post-trau-
1226–1232. matic AQP4 dysregulation. J Cereb Blood Flow Metab. 2013;
28. Colombo M, Raposo G, Théry C. Biogenesis, secretion, and inter- 33(6):834–845.
cellular interactions of exosomes and other extracellular vesicles. 48. Buniatian GH, Hartmann H, Traub P, et al. Glial fibrillary acidic
Annu Rev Cell Dev Biol. 2014;30:255–289. protein-positive cells of the kidney are capable of raising a protect-
29. Mathivanan S, Ji H, Simpson RJ. Exosomes: Extracellular organ- ive biochemical barrier similar to astrocytes: Expression of metal-
elles important in intercellular communication. J Proteomics. lothionein in podocytes. Anat Rec. 2002;267(4):296–306.
2010;73(10):1907–1920. 49. Binkowska AM, Michalak G, Pilip S, Kopacz M, Slotwinski R.
30. Simons M, Raposo G. Exosomes – vesicular carriers for intercellu- The diagnostic value of early cytokine response in patients after
lar communication. Curr Opin Cell Biol. 2009;21(4):575–581. major trauma – Preliminary report. Cent Eur J Immunol. 2018;
31. Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJA. 43(1):33–41.
Delivery of siRNA to the mouse brain by systemic injection of tar- 50. Tisoncik JR, Korth MJ, Simmons CP, Farrar J, Martin TR, Katze
geted exosomes. Nat Biotechnol. 2011;29(4):341–345. MG. Into the eye of the cytokine storm. Microbiol Mol Biol Rev.
32. Caby MP, Lankar D, Vincendeau-Scherrer C, Raposo G, Bonnerot 2012;76(1):16–32.
C. Exosomal-like vesicles are present in human blood. Int 51. Spurlock M. Regulation of metabolism and growth during im-
Immunol. 2005;17(7):879–887. mune challenge: an overview of cytokine function. J Anim Sci.
33. Cheng Y, Pereira M, Raukar N, et al. Potential biomarkers to de- 1997;75(7):1773–1783.
tect traumatic brain injury by the profiling of salivary extracellular 52. Dursun E, Gezen-Ak D, Hanagasi H, et al. The interleukin 1alpha,
vesicles. J Cell Physiol. 2019;234(8):14377–14388. interleukin 1 beta, interleukin 6 and alpha-2-macroglobulin serum
34. Pisitkun T, Shen R, Knepper MA. Identification and proteomic levels in patients with early or late onset Alzheimer’s disease, mild
profiling of exosomes in human urine. Proc Natl Acad Sci USA. cognitive impairment or Parkinson’s disease. J Neuroimmunol.
2004;101(36):13368–13373. 2015;283:50–57.
35. Gill J, Mustapic M, Diaz-Arrastia R, et al. Higher exosomal tau, 53. Mondello S, Guedes VA, Lai C, et al. Circulating brain injury exo-
amyloid-beta 42 and IL-10 are associated with mild TBIs and somal proteins following moderate-to-severe traumatic brain in-
chronic symptoms in military personnel. Brain Inj. 2018;32(10): jury: Temporal profile, outcome prediction and therapy
1277–1284. implications. Cells. 2020;9(4):977.
36. Harrison EB, Hochfelder CG, Lamberty BG, et al. Traumatic brain 54. Prakash R, Carmichael ST. Blood-brain barrier breakdown and
injury increases levels of miR-21 in extracellular vesicles: neovascularization processes after stroke and traumatic brain in-
Implications for neuroinflammation. FEBS Open Bio. 2016;6(8): jury. Curr Opin Neurol. 2015;28(6):556–564.
835–846. 55. Hirad AA, Bazarian JJ, Merchant-Borna K, et al. A common neur-
37. Ko J, Hemphill M, Yang Z, et al. Diagnosis of traumatic brain in- al signature of brain injury in concussion and subconcussion. Sci
jury using miRNA signatures in nanomagnetically isolated brain- Adv. 2019;5(8):eaau3460.
derived extracellular vesicles. Lab Chip. 2018;18(23):3617–3630. 56. Marchi N, Bazarian JJ, Puvenna V, et al. Consequences of
38. Ko J, Hemphill M, Yang Z, et al. Multi-dimensional mapping of repeated blood-brain barrier disruption in football players. PLoS
brain-derived extracellular vesicle microRNA biomarker for trau- One. 2013;8(3):e56805.
matic brain injury diagnostics. J Neurotrauma. 2020;37(22): 57. Tagge CA, Fisher AM, Minaeva OV, et al. Concussion, micro-
2424–2434. vascular injury, and early tauopathy in young athletes after impact
39. Ko J, Yee SS, Ortiz N, et al. Combining machine learning and head injury and an impact concussion mouse model. Brain. 2018;
nanofluidic technology to diagnose pancreatic cancer using exo- 141(2):422–458.
somes. ACS Nano. 2017;11(11):11182–11193. 58. Johnson VE, Weber MT, Xiao R, et al. Mechanical disruption of
40. Lotval H, Hill AF, Hochberg F, et al. Minimal experimental the blood-brain barrier following experimental concussion. Acta
requirements for definition of extracellular vesicles and their Neuropathol. 2018;135(5):711–726.You can also read

















































