Morphologic Effects of the Stress Response in Fish
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Morphologic Effects of the Stress Response in Fish
Claudia Harper and Jeffrey C. Wolf
Abstract 32). Subsequent researchers have proposed a variety of al-
ternate definitions, but, as commonly used, the word stress
Downloaded from https://academic.oup.com/ilarjournal/article-abstract/50/4/387/868673 by guest on 09 February 2020
Fish and other aquatic animals are subject to a broad variety still conveys a vague notion of unease, distress, discomfort,
of stressors because their homeostatic mechanisms are or disturbance. Ambiguity exists in part because the word
highly dependent on prevailing conditions in their immedi- can be used to indicate one of three different components of
ate surroundings. Yet few studies have addressed stress as a what is essentially a cause and effect relationship: (1) a
potential confounding factor for bioassays that use fish as physical or mental stimulus, (2) an individual’s physical or
test subjects. Common stressors encountered by captive fish mental awareness of that stimulus, or (3) the individual’s
include physical and mental trauma associated with capture, physical or behavioral response to the stimulus. For exam-
transport, handling, and crowding; malnutrition; variations ple, exposure to cold temperatures (stimulus) can make an
in water temperature, oxygen, and salinity; and peripheral individual feel cold (awareness) and evoke shivering (re-
effects of contaminant exposure or infectious disease. Some sponse); regarded independently, each of these elements
stress responses are detectable through gross or microscopic might be considered stress. As one frustrated scientist re-
examination of various organs or tissues; as reported in the portedly claimed, “Stress, in addition to being itself and the
literature, stress responses are most consistently observed in result of itself, is also the cause of itself” (Roberts 1950,
the gills, liver, skin, and components of the urogenital tract. 105). To avoid confusion, we use the term “stressors” to in-
In addition to presenting examples of various stressors and dicate stressful stimuli and “stress responses” to designate
corresponding morphologic effects, this review highlights the reactions to such stimuli.
certain challenges of evaluating stress in fish: (1) stress is an The purpose of this article is to review morphologic ef-
amorphous term that does not have a consistently applied fects of various stressors in fish as determined by gross or
definition; (2) procedures used to determine or measure histopathologic investigation. In addition, descriptions of
stress can be inherently stressful; (3) interactions between both tissue-specific and non-tissue-specific stress responses
stressors and stress responses are highly complex; and (4) are provided. For more general discussions of piscine stress,
morphologically, stress responses are often difficult to dis- several excellent reviews are available (Barton 2002; Gratzek
tinguish from tissue damage or compensatory adaptations and Reinert 1984; Iwama et al. 2004a; Pickering 1981).
induced specifically by the stressor. Further investigations
are necessary to more precisely define the role of stress in
the interpretation of fish research results.
Stressors and Stress Responses
Key Words: contaminant; crowding; fish; handling; histol- Throughout the animal kingdom, many types of stressors are
ogy; nutrition; temperature; salinity; stress universal simply because the basic needs of most animals are
similar. Examples of universal stressors include deviations
Introduction from optimal ranges for environmental parameters (e.g., ambi-
ent temperature, oxygen supply), insufficient food availability,
I
n 1936 a scientist named Hans Selye, upon observing inadequate refuge from sunlight or predators, and the demands
effects of noxious stimuli in laboratory animals, coined of social interactions such as territorial disputes. Other stress-
the term “stress” and defined it as “the non-specific re- ors are unique to certain animal groups or habitats.
sponse of the body to any demand for change” (Selye 1936, As compared to terrestrial inhabitants, fish and other
aquatic creatures are subject to a broader variety of stressors
because their homeostatic mechanisms are highly dependent
on prevailing conditions in their immediate surroundings.
Examples of additional stressors for fish include fluctuations
Claudia Harper, DVM, DACLAM, is Director of Preclinical at Amgen Inc. in water salinity, pH, hardness, alkalinity, dissolved solids,
Jeffrey C. Wolf, DVM, DACVP, is a toxicologic veterinary pathologist at
water level or current, and exposure to waterborne pathogens
Experimental Pathology Laboratories Inc. in Sterling, Virginia.
Address correspondence and reprint requests to Dr. Jeffrey C. Wolf, or toxicants. Fish reared in confinement systems often expe-
Experimental Pathology Laboratories, 45600 Terminal Drive, Sterling, rience further pressures of crowding, handling, suboptimal
VA 20166 or email jwolf@epl-inc.com. nutrition, and nitrogenous waste accumulation.
Volume 50, Number 4 2009 387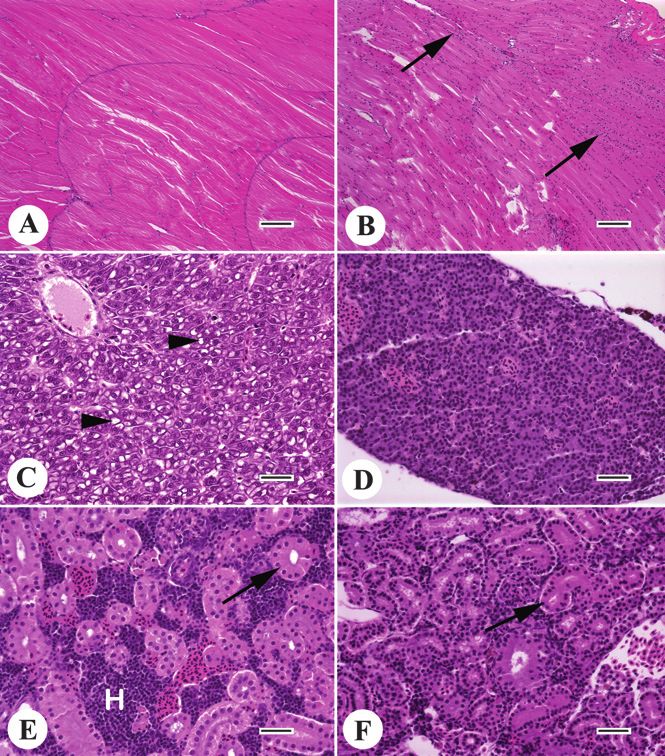
Evidence indicates that certain stress responses are well To date, methods for evaluating stress responses in fish
conserved evolutionarily. In terms of behavior, an obvious have involved a variety of endpoints:
example is the instinctive urge to fight or flee when faced • whole body or organ weight measurements (e.g., condi-
with an adverse stressor such as predation. Many physiologi- tion factor, hepatosomatic index, and gonadosomatic in-
cal responses to stressors are also remarkably comparable dex) (Dutta et al. 2005; Hosoya et al. 2007; Spencer et al.
among taxonomically diverse animals. For instance, com- 2008);
mon among all vertebrates is the stressor-induced secretion • biochemical assays (e.g., plasma cortisol, corticosterone,
of adrenergic and glucocorticoid hormones; the latter espe- glucose, tissue damage enzymes, and heat shock pro-
cially is considered a hallmark of the stress response (Nesse teins) (Acerete et al. 2004; Barton 2002; Dutta et al.
and Young 2000). Although fish lack adrenal glands per se, 2005; Hosoya et al. 2007; Iwama et al. 2004b; Olsen
analogous production and release of adrenal cortical and et al. 2008; Trenzado et al. 2008);
• immune function (Choi et al. 2007);
Downloaded from https://academic.oup.com/ilarjournal/article-abstract/50/4/387/868673 by guest on 09 February 2020
medullary hormones occur in the interrenal cells and chro-
maffin tissues, respectively, both of which are typically lo- • gene expression patterns (Basu et al. 2001, 2002;
cated in the piscine anterior kidney. Marques et al. 2008; van der Meer et al. 2005);
As in the case of so-called higher vertebrates, the secre- • measurement of fish steroids in water (Scott and Ellis
tion of stress-related hormones in fish can be a double-edged 2007); and
sword. The activities of these hormones are clearly benefi- • macroscopic and microscopic anatomy (numerous refer-
cial when acute action and its consequences take priority, as ences cited in the following text).
they elicit a heightened state of alertness, increase blood As a research tool, the histopathologic evaluation of whole
pressure and respiration, promote hepatic glycogen catabo- body sections from small fish species offers numerous advan-
lism to provide a source of energy via glucose, and limit ex- tages, including the ability to observe a wide variety of organ
cessive tissue damage from inflammatory reactions to trauma systems in relatively few tissue sections, the ability to iden-
or illness (Nesse and Young 2000). However, hormonal stress tify concurrent disease problems, the long-term stability of
responses that overcompensate or persist can also have nega- the raw data (because histologic sections are mounted on
tive effects, such as immune suppression, depletion of en- glass slides), and, perhaps most importantly, the ability to de-
ergy reserves, muscle breakdown, and, in fish, interference tect treatment-induced changes that might otherwise remain
with osmoregulation as a result of altered mineral metabo- undiscovered.
lism (Banerjee and Bhattacharya 1995). Although the potential for stress responses to confound
certain experimental results can be high, only a limited num-
ber of studies have specifically addressed the effects of such
Measurement of Stress Responses responses on tissue histomorphology. For example, despite
documented stressor-induced alterations of reproductive sys-
Despite the commonality of the stress response, for several tem endpoints (Cleary et al. 2002; Contreras-Sánchez et al.
reasons it is not always easy to measure its effects in an ex- 1998), there has been little effort to determine potential histo-
perimental setting. First, such responses are not “all or noth- pathologic effects of the stress response (e.g., as modeled by
ing” events. As exposure to a particular stressor increases in cortisol administration) on fish gonads or gonadal ducts; such
magnitude and duration, the outcome can progress from a effects might include increased germ cell degeneration in the
complete lack of clinical effects to relatively subtle manifes- ovary (oocyte atresia) and/or testis. Such findings would be
tations (e.g., decreased reproductive performance) to patent significant because those same types of changes are often re-
signs of disease (e.g., life-threatening microorganism infec- garded as prima facie evidence of endocrine disruption (Heiden
tions) (Benli et al. 2008). Further complicating this picture et al. 2006; Leino et al. 2005; Rasmussen et al. 2005). Sea-
are adaptive mechanisms that may compensate to varying sonal changes also are known to affect fish gonads and lead
degrees for chronic or low-level stress and thereby contrib- to morphological changes (Abe and Munehara 2007).
ute to inconsistency in stress responses among test subjects.
A second challenge is that, analogous to the “observer effect”
described in quantum physics, efforts to measure in vivo Fish-Specific Stressors
stress responses can be stressful in and of themselves; for
example, the capture process can affect levels of measured There are roughly 30,000 known species of fish, and both
cortisol levels in wild fish collected for stress management wild and captive fishes occupy a remarkably diverse array of
research (Cleary et al. 2002; Tsunoda et al. 1999). A third habitats. Accordingly, environmental conditions that might
challenge for scientists is that the effects of the stress response be optimal for one species are inherently stressful for an-
can be difficult to distinguish from effects of the stressor it- other. Given the number of potential stressors, and the fact
self (Selye 1955); this represents an important obstacle in that fish may be exposed to multiple stressors simultane-
endocrine disruption research, in which it is necessary to dif- ously, the range of potential stress-inducing situations is al-
ferentiate the particular effects of hormonally active sub- most limitless. This section provides brief descriptions of
stances from their concomitant ability to contribute to the commonly encountered stressors and the anatomic sites in
stress response (Norris 2000). which corresponding morphologic effects tend to occur.
388 ILAR Journal
Specific histopathologic findings are described in more de- ments, it is reasonable to surmise that certain durations of
tail in the following section, categorizing stress responses by confinement stress might therefore manifest in histopatho-
tissue type or organ system. logic findings such as decreased hepatocellular vacuolation
(especially in cultured fish) and muscle atrophy. Burgess and
Coss (1982) examined histologic specimens from adult jewel
Capture, Transport, and Handling fish (Hemichromis bimaculatus Gill) and determined that
moderate crowding stress was associated with morphologic
Capture, transport, and handling are obvious stressors for
changes in the brain.
captive fish, but wild fish may also experience these distur-
bances, for example through catch and release programs in
recreational fisheries. Procedures that can intensify the stress Hyper- or Hypothermia
response in aquacultured fish include sorting, grading, and
Downloaded from https://academic.oup.com/ilarjournal/article-abstract/50/4/387/868673 by guest on 09 February 2020
vaccine administration (Burgess and Coss 1982). Additional Fish are subject to stress from either rapid temperature fluc-
stressful sequelae include crowding, hypoxia, physical tuations that preclude acclimation or inappropriate water
trauma, aftereffects of anesthetics or sedatives, and baromet- temperature (beyond the high or low range of tolerance). A
ric disturbance in fish harvested at considerable depth. Evi- rapid temperature decrease limits a fish’s ability to produce
dence that these stimuli are intrinsically stressful is provided antibodies integral to an immediate immune response, and a
by experiments that have documented marked increases in delay in the immune response may enable pathogens to colo-
blood cortisol and/or glucose levels in fish following deliber- nize, reproduce, and establish an infection. Very cold tem-
ate handling and transport (Acerete et al. 2004; Barton 2002; peratures may inactivate defensive functions of nonspecific
Hosoya et al. 2007). leukocytes known as natural killer (NK) cells, although there
There may be some benefit to sedating fish before trans- is some evidence from studies in common carp (Cyprinus
port in order to mitigate shipping stress. In a study in which carpio) that NK cells may be able to accommodate tempera-
channel catfish (Ictalurus punctatus) were subjected to stress- ture changes over time (Kurata et al. 1995). Hyperthermia
ors such as confinement, high ammonia, and oxygen deple- has been used experimentally as a stressor in challenge stud-
tion, sedation resulted in lower cortisol elevations than those ies involving infectious agents, for example in rainbow trout
observed in control fish (Small 2004). But the magnitude of (Oncorhynchus mykiss) exposed to Saprolegnia parasitica
the stress response to netting, transport, and handling varies (Gieseker et al. 2006). This same stressor also contributed to
considerably among species and, typical of stress responses altered thyroid indices, including augmentation of thyroid
in fish, clinical effects often do not become apparent until epithelial cell height, in rainbow trout exposed to PCBs
several days after the stress-inducing event, when secondary (Buckman et al. 2007).
bacterial, viral, fungal, or parasitic infections manifest.
Notwithstanding the frequency at which fish experience
these stressors, there has been very little investigation of po- Hypoxia
tential histomorphologic consequences. For example, although
Anoxic conditions are commonly the result of plant, algae,
anecdotal observations suggest that fish may suffer micro-
or diatom overgrowth in either natural or captive environ-
scopically evident muscle degeneration (rhabdomyolysis) as a
ments, but hypoxia can also occur when fish are shipped in
consequence of collection (capture myopathy), experiments
insufficiently aerated containers, for example. The decrease
have not been conducted to confirm this causal relationship.
in oxygen availability to tissues can lead to necrotic or apop-
totic lesions in organs (Geng 2003; van der Meer et al. 2005).
Crowding In channel catfish, experimentally induced sublethal hypoxia
was responsible for histopathologically evident necrosis, hy-
For captive fish, appropriate stocking density varies greatly peremia (vascular congestion), edema, hemorrhage, hyper-
according to the species, housing system, and available re- plasia, and/or hypertrophy in a variety of anatomic sites
sources. Overcrowding may be accompanied by additional including the gills, liver, spleen, and anterior and posterior
stressors such as poor water quality, exposure to organic kidney (Scott and Rogers 1980). Although it could be rea-
wastes, and conspecific aggression and predation. Gilthead sonably argued that such lesions formed as a specific reac-
seabream (Sparus aurata L.) experienced significant rapid tion to acute localized oxygen deprivation rather than to
increases in blood cortisol and glucose following short-term stress per se, it is plausible that stress contributed to the re-
crowding (Ortuño et al. 2001), and similar results were ob- sponse on some level.
served in tilapia (Oreochromis mossambicus) (Vijayan et al. Some teleost fish, frogs, turtles, snakes, and insects have
1997), thus supporting the role of crowding as a stressor. In the capacity to tolerate or adapt to hypoxia (van der Meer
tilapia, glucose elevations after 2 hours of confinement were et al. 2005). For instance, zebrafish (Danio rerio) can survive
attributed to glycogenolysis, whereas in fish confined for 24 weeks of severe hypoxia through adaptive responses that
hours gluconeogenesis was considered the primary mecha- modulate their behavioral and physical phenotype: evidence
nism for glucose elevations (Vijayan et al. 1997). Although from cDNA microarray technology revealed changes in gene
morphologic changes were not the focus of these experi- expression in their gills as well as gene repression that affected
Volume 50, Number 4 2009 389protein biosynthesis and metabolic pathways (van der Meer not be provided because of cost or lack of availability, or
et al. 2005). nutrient degradation occurred during feed storage.
More typically, however, chronic hypoxia has been shown Because stress-reactive hormones such as glucocorticoids
to cause an assortment of phenotypic changes in a diverse have a constituent role in energy homeostasis, it is often dif-
range of organ systems and fish species, including the hearts ficult to separate stress responses from the direct effects of
of zebrafish and cichlids (Haplochromis piceatus) (Marques malnutrition in terms of morphologic consequences. For ex-
et al. 2008); the reproductive tracts of common carp (Wang ample, starvation may cause a histologically evident decrease
et al. 2008) and Atlantic croaker (Micropogonias undulatus) in liver glycogen stores not only as a result of increased energy
(Thomas et al. 2007); peripheral blood leukocytes of tilapia expenditure relative to intake but also because of stress-
(Choi et al. 2007); and the eyes of platyfish (Xiphophorus induced corticosteroid-mediated glycogenolysis (Barton and
maculatus) exposed to hypoxic conditions perinatally (Chan Schreck 1987; Vijayan et al. 1997). Furthermore, food depri-
Downloaded from https://academic.oup.com/ilarjournal/article-abstract/50/4/387/868673 by guest on 09 February 2020
et al. 2007). In the gills, hypoxia has been associated with an vation can lead to reduced stress resistance, as was the out-
adaptive increase in lamellar surface area in fishes such come when food-denied Atlantic cod (Gadus morhua L.) were
as certain African cichlids and Crucian carp (Carassius subjected to exhaustive exercise (Olsen et al. 2008). Another
carassius) (Chapman et al. 2000; Sollid et al. 2003; van der recent study further demonstrated that nutrient imbalances
Meer et al. 2005). can influence the stress response, as higher blood cortisol con-
centrations in rainbow trout were associated with dietary vari-
ations of vitamin E, vitamin C, and highly unsaturated fatty
Hyper- or Hyposalinity acids (Trenzado et al. 2008). In such cases it may be difficult
to discriminate the stressor from the stress response; for ex-
Freshwater fish are under continuous pressure to conserve
ample, interrenal ascorbic acid concentrations decreased in
salts, whereas the reverse is true for marine species, which
rainbow trout and coho salmon (Oncorhynchus kisutch) that
must conserve water (Greenwell et al. 2003). Among fishes
were subjected to nonspecific stress (Wedemeyer 1969).
in general, the ability to adapt to alterations in salinity varies
markedly and often is indirectly proportional to the pace of
the changes. In natural settings, salinity levels can fluctuate
with tides, season, or evaporation from surface waters. Contaminants
Few studies have investigated potential morphologic
Fish have been exposed, either intentionally or unintention-
effects of salinity as the sole stressor. An experiment to as-
ally, to a vast array of chemical and particulate contaminants,
sess optimal stocking densities for sea bass (Dicentrarchus
of both natural and man-made origin. Examples include
labrax) fingerlings applied hypersalinity as a stressor along
pharmaceuticals, agricultural chemicals, manufacturing by-
with temperature modifications (Via et al. 1998). But an ex-
products, animal and human waste materials, mining efflu-
periment that specifically evaluated the tolerance of hybrid
ents, and substances released as a consequence of natural
tilapia (Oreochromis mossambicus × O. urolepis hornorum)
disasters such as fires. Arguably, at sufficient concentration,
to hypersaline water found that the primary morphologic in-
almost any contaminant is capable of inducing a stress re-
dicators of hypersaline stress, and the most sensitive of sev-
sponse. In some exposures, the stressor is a mixture of known
eral endpoints tested, were ultrastructural changes in the
and unknown contaminants (Dutta et al. 2005; Teh et al.
gills (Sardella et al. 2004).
1997), in which case it is almost impossible to differentiate
In anadromous fish such as salmon, physiological changes
stress response effects from manifestations of toxicity. How-
associated with smoltification (the metamorphic transforma-
ever, such differentiation can be challenging even when the
tion that occurs in juveniles before their freshwater to marine
contaminant is a single compound.
migration) are consistently stressful, as suggested by changes
One of the most studied contaminants is ammonia, high
in plasma cortisol levels (Barton 2002).
levels of which result from agricultural or mining operation
runoff, excessive biological waste accumulation, insufficient
Malnutrition water aeration, or inadequate tank conditioning (Noga 1996;
Randall and Tsui 2002; Spencer et al. 2008). Ammonia is
Using a greatly simplified classification system, malnutrition toxic to all vertebrates, and the effects of both acute and
can be categorized as disorders that result from either (1) an chronic ammonia exposure have been investigated in a num-
insufficiency or overabundance of nutrients or (2) relative ber of fish species. Acute ammonia toxicity can cause an as-
nutrient imbalances. Factors that typically contribute to mal- sortment of clinical signs in fish, the most severe of which
nutrition in wild fish include depletion of species-appropriate include convulsions, coma, and death (Randall and Tsui
food sources or components (e.g., vitamins, minerals), 2002), as well as less severe impacts such as plasma cortisol
heightened competition for available food resources, and elevations and behavioral changes such as hyperexcitability
inappetence due to disease. Captive fish often endure the ad- and appetite suppression (Ortega et al. 2005). Its effects may
ditional challenge of suboptimal feed formulation, usually be exacerbated by increased pH or temperature, excessive
because the precise nutritional requirements for the fish spe- exercise, starvation, and stress (simulated by cortisol injec-
cies of interest have not been determined, a suitable diet can- tion) (Randall and Tsui 2002; Spencer et al. 2008).
390 ILAR JournalAmmonia exposure has been associated with morphologic
findings in a variety of fish tissues. The gills are one of the
most frequently reported targets (Benli et al. 2008; Frances
et al. 2000; Lease et al. 2003; Spencer et al. 2008), although in
one study involving chronic ammonia toxicity in rainbow
trout, gill changes were not observed histologically, even in
high-dose fish that had suffered from neurological dysfunc-
tion (Daoust and Ferguson 1984). In addition to the gills,
ammonia-related lesions have been reported in the liver, kid-
ney, intestine, and ovary of fish (Banerjee and Bhattacharya
1994, 1995; Benli et al. 2008; Dey and Bhattacharya 1989).
Downloaded from https://academic.oup.com/ilarjournal/article-abstract/50/4/387/868673 by guest on 09 February 2020
Stress Responses
Fish responses to stress can be divided into three phases:
primary, secondary, and tertiary (Barton 2002). The primary
phase refers to a generalized neuroendocrine response in
which catecholamines (epinephrine and norepinephrine) and
cortisol are released from chromaffin and interrenal cells,
respectively. Higher circulating levels of these hormones
trigger a secondary response that involves physiologic and
metabolic pathways; examples of the secondary response in-
clude hyperglycemia due to enhanced glycogenolysis and
gluconeogenesis, vasodilation of arteries in gill filaments,
increased cardiac stroke volume, and immune function de-
pression (Gratzek and Reinert 1984). The first two phases
are considered adaptive and enable fish to adjust to stressors
and maintain homeostasis. In contrast, tertiary responses in-
volve systemic changes in which animals may become inca-
pable of adapting to stressors, leading to adverse effects on
the animals’ overall health, including their performance,
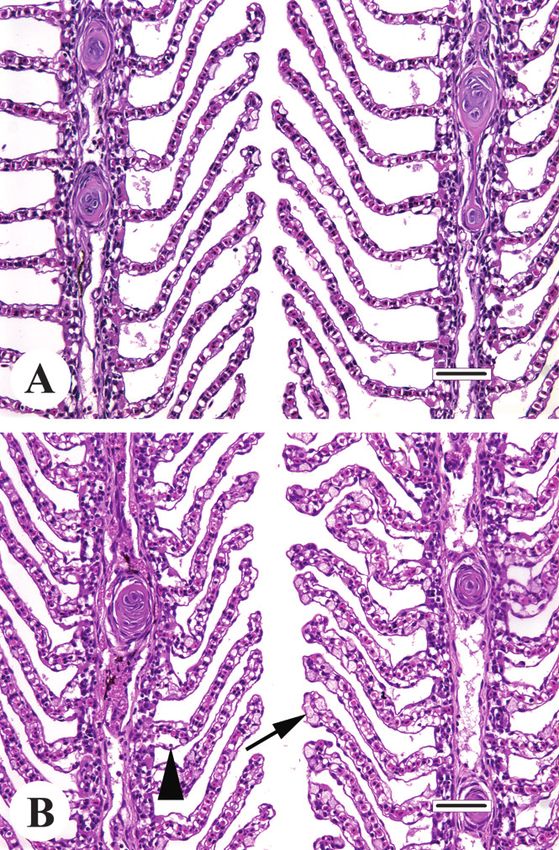
growth, reproduction, disease resistance, and behavior (Barton Figure 1 Nonspecific stress response in the gills of adult Atlantic
salmon (Salmo salar L.). (A) Normal gill (two adjacent filaments).
2002). The following sections provide examples of adaptive
(B) Findings associated with several types of stressors; the most
and postadaptive stress responses according to organ system. prominent changes are mucus cell hyperplasia (arrow) and epithe-
This is by no means an exhaustive record; undoubtedly, mor- lial lifting (arrowhead). Bar = 50 microns.
phologic indications of stress also exist in tissue types that
are less routinely examined.
sign, decreases the respiratory surface area. Under hypoxic
Gills conditions, this cell mass recedes due to the combined ef-
fects of increased apoptosis and diminished cell prolifera-
Given the relative fragility of the gills compared to other sur- tion, and as it shrinks it exposes the underlying lamellae,
face tissues, and the fact that they are continually exposed to thus increasing the overall surface area of the gills. This ad-
the fish’s external environment, it is remarkable that these aptation may have evolved to reduce water and ion flux
structures are able to survive and compensate for the chemi- under normoxic conditions and thus conserve energy for
cal and physical assaults to which they are invariably sub- osmoregulation. Similarly, in various African cichlid fish ex-
jected. It is therefore not surprising that, based on a survey of posure to long-term hypoxia resulted in elongation of bran-
the literature, the gills appear to be a frequent target for stress chial filaments and an increase in the size of secondary
responses (Figure 1). lamellae (Chapman et al. 2000).
Some fishes have developed intriguing adaptive stress Most species, however, are not capable of adapting so
response mechanisms. For example, the gills of Crucian carp effectively to hypoxic conditions. Channel catfish exposed
exhibit a reversible morphological reaction to decreased ox- to varying degrees of sublethal hypoxia exhibited a suite of
ygen availability (Sollid et al. 2003), thanks to a unique ana- nonspecific, histologically evident changes likely to inter-
tomic feature: under normal ambient oxygen concentrations, fere with respiratory gas exchange, such as gill epithelial
the gills lack protruding secondary lamellae (typically the hypertrophy and hyperplasia, goblet cell proliferation with
primary sites of gas exchange in other fishes); instead, the increased mucus secretion, hemorrhage, edema, and telangi-
secondary lamellae are embedded in a cell mass that, by de- ectasis (Scott and Rogers 1980).
Volume 50, Number 4 2009 391Hypersalinity results in a qualitatively different type of vere hepatic lipidosis, lymphoid cell depletion, vascular con-
negative response. Apoptosis of chloride cells (branchial cells gestion, and reticuloendothelial cell necrosis (in the spleen)
that facilitate ion transport and have an integral role in acid- were evident in the livers and spleens of wild freshwater fish
base regulation; Perry 1998) occurred in hybrid tilapia ex- exposed to mixed contaminants (Teh et al. 1997). The added
posed experimentally to various concentrations of hypersaline presence of a number of preneoplastic and neoplastic prolif-
water for a model of salinity tolerance (Sardella et al. 2004). erative lesions in those fish strongly suggests that factors
Ammonia-induced gill changes have been particularly other than glucocorticoid-mediated stress (e.g., chemical
well characterized, for species as diverse as Nile tilapia carcinogenesis, patent toxicity) may have contributed to at
(Oreochromis nilotica), slimy sculpin (Cottus cognatus), least some of the chronic changes.
and endangered Lost River suckers (Deltistes luxatus). They
include nonspecific responses such as lamellar thickening,
Integument
Downloaded from https://academic.oup.com/ilarjournal/article-abstract/50/4/387/868673 by guest on 09 February 2020
mucus cell hyperplasia and hypertrophy, epithelial cell lift-
ing, leukocyte infiltration, hyperemia, hemorrhage, chloride
The skin, with its scales and surface mucus, provides a protec-
cell proliferation, secondary lamellar fusion, and telangi-
tive physical barrier that is important in terms of both osmo-
ectasis (Benli et al. 2008; Lease et al. 2003; Spencer et al.
regulation and pathogen defense. But fish skin is susceptible
2008). After an investigation of the combined effects of am-
to damage from handling, fighting, physical trauma, preda-
monia and elevated pH in Lost River suckers, Lease and col-
tion, environmental irritants, and pathogens, and the damage
leagues (2003) concluded that structural gill changes were
can lead to opportunistic microbial infections. At that stage
more sensitive than other traditional assays for detecting am-
the stress response may further compromise the host’s de-
monia toxicity. An earlier study recorded similar types of
fenses, via corticosteroid-mediated immunosuppression or
morphologic findings in wild freshwater fish exposed to a
other stress-related immunosuppressive factors (Choi et al.
mixture of known and unknown contaminants (Teh et al.
2007; Harris et al. 2000; Kent and Hedrick 1987).
1997). Gill lesions in that study included hyperplastic mu-
Although fish skin has not been reported extensively as a
cous and chloride cells, deformed branchial cartilages, se-
stress response target, dermal ulceration was the chief finding
vere and diffuse lamellar aneurysms (telangiectasis), and
in a series of studies in which striped bass (Morone saxatilis)
edema at the bases of secondary lamellae.
and striped bass hybrids were exposed to acute confinement
At this point it may seem that any type of stressor might
stress (Noga et al. 1998; Udomkusonsri et al. 2004). Associ-
induce almost any type of gill lesion as part of a stress re-
ated histopathologic lesions, in addition to rapidly occurring
sponse. But apparently this is not necessarily the case, as one
epithelial erosions and ulcers that primarily affected the fins,
study has demonstrated that social stress did not lead to chlo-
included epithelial cell swelling, edema of the dermis and
ride cell proliferation in rainbow trout (Sloman et al. 2005).
hypodermis, melanophore aggregation, and stromal tissue
necrosis.
Liver
Unlike the gills, the liver is clearly protected from physical Genitourinary Tract
exposure to the external environment, at least under normal
circumstances. It is prone, however, to chemical assault, in Although there are reports of functional and/or hormonal
part due to an efficient enterohepatic cycling mechanism impairment of the fish reproductive system due to various
(Gingerich 1982). Stress responses may also be evident in stressors (capture, handling, crowding, hypoxia, tank drain-
the liver because of its prominent role in energy storage and ing, noise) (Cleary et al. 2002; Contreras-Sánchez et al.
metabolism. Often, quantitative alterations in hepatic energy 1998; Thomas et al 2007; Wang et al. 2008), there has been
storage are visible macroscopically as changes in liver size only limited investigation of the potential morphologic ef-
and coloration, and histologically as variations in hepatocel- fects of such stressors in the gonads or genital ducts. One
lular vacuolation and tinctorial staining characteristics (Wolf study found retarded oocyte maturation in common carp ex-
and Wolfe 2005). Decreased vacuolation can result from loss posed to chronic hypoxia (Wang et al. 2008). In another
of cytoplasmic glycogen and/or lipid caused by insufficient study conducted in Atlantic croaker, hypoxia was associated
energy intake relative to need and/or glucocorticoid-induced with decreased gonadosomatic index (gonadal weight/body
glycogenolysis. Conversely, increased hepatocellular vacu- weight) and impaired gametogenesis (determined via mor-
olation is more commonly associated with overnutrition or phometric counting of ovarian and testicular germ cells in
toxicity (Wolf and Wolfe 2005). As an example of the latter, histologic sections) in both male and female fish (Thomas
cloudy swelling and hydropic degeneration occurred in Nile et al. 2007).
tilapia exposed to sublethal concentrations of ammonia There are even fewer reports of stress responses that
(Benli et al. 2008). On the other hand, alterations in cyto- involve the fish urinary tract. Examples include hypoxia-
plasmic vacuolation were not features of hypoxia in channel induced hemorrhage, glomerular congestion, and edema in
catfish, which instead showed hepatic necrosis and hemor- the posterior kidneys of channel catfish (Scott and Rogers
rhage as well as splenic changes such as edema, hyperemia, 1980), and congestion in Nile tilapia exposed to sublethal
and necrosis. Chronic histopathologic changes such as se- concentrations of ammonia (Benli et al. 2008).
392 ILAR JournalNervous and Sensory Systems in many different fish tissues including blood vessels in hy-
poxia-tolerant fish (Cossins et al. 2009).
Routine diagnostic examinations or experimental investiga-
tions involving fish tissues tend to include sampling of the
brain and spinal cord less frequently than for other organs. Multiorgan and Systemic Stress Responses
Although inflammation and endoparasitism of the central
nervous system are often readily recognizable in standard Systemic stress responses include alterations (often de-
histologic sections, more subtle types of changes are not al- creases) in body condition and/or organ weights, with cor-
ways easily appreciated. For example, in a series of experi- responding histopathologic changes such as atrophy of
ments in which jewel fish were exposed to chronic crowding adipose tissue (fat), skeletal and cardiac muscle, and liver
stress, special histologic staining and morphometric tech- cells, among other tissue types (Figure 2).
One particular multiorgan stress response involves the for-
Downloaded from https://academic.oup.com/ilarjournal/article-abstract/50/4/387/868673 by guest on 09 February 2020
niques were required in order to determine that, compared to
controls, crowded fish had structural nerve cell alterations mation of histologically evident pigmented macrophage ag-
(both qualitative and quantitative) in the optic tectum, a ma- gregates (PMA; Figure 3). These melanomacrophage centers
jor area of the brain concerned with processing and integrat- are variably sized constituent nests of phagocytic cells that can
ing sensory information (Burgess and Coss 1982). Of course contain one or more intracytoplasmic pigments, such as ceroid,
it could be debated that the outcome was not truly a stress lipofuscin, melanin, and hemosiderin (Wolke 1992). Although
response but instead a developmental adaptation caused by the kidney and spleen tend to be common locations for these
long-term differences in patterns of sensory stimulation.
Comparable to the central nervous system, the detec-
tion of stress-related changes in the eyes may also require
detailed examination. For example, findings in perinatal
platyfish subjected to hypoxic conditions included central
corneal thinning, hyperplasia of corneal endothelial cells,
lens fiber derangement, and apoptotic cells in the retina
(Chan et al. 2007). Perhaps more obvious were the corneal
ulcerations induced by acute confinement stress in hybrid
striped bass (Morone saxatilis × M. chrysops) (Udomkusonsri
et al. 2004).
Cardiovascular System
Histologically evident changes in the hearts of adult ze-
brafish and Lake Victoria cichlids (Haplochromis piceatus)
subjected to chronic hypoxia included reduced ventricular
outflow tracts and reduced lacunae surrounding trabeculae
(Marques et al. 2008). Quantitation of myocyte nuclei in
both species also revealed that, relative to controls, hypoxic
fish had increased numbers of nuclei per unit area.
Occasionally, microscopic examinations of blood smears
can reveal morphologic evidence of stress that would be dif-
ficult to detect in tissue sections. For example, two classic
hematological manifestations of the stress response in mam- Figure 2 Histomorphologic effects of chronic starvation in adult
mals, neutrophilia and lymphopenia, were triggered in Nile female Japanese medaka (Oryzias latipes). Images (A), (C), and (E)
tilapia by acute hypoxia followed by reperfusion (Choi et al. are from a well-nourished fish; (B), (D), and (F) are from a fish that
2007). Although a description of hematological changes as- suffered a prolonged negative energy balance due to inanition and
sociated with stress is outside the scope of this article, many stress associated with egg retention. (A) Normal skeletal muscle.
publications clearly indicate that stressors such as handling, (B) Skeletal muscle atrophy; muscle cell nuclei (arrows) appear
crowding, capture, restraint, hypoxia, anesthesia, air expo- clumped as a result of the decrease in muscle fiber size. (C) Normal
liver; arrowheads indicate moderate hepatocyte vacuolation consis-
sure, and sampling technique can affect fish hematology
tent with glycogen storage. (D) Liver atrophy; the tissue is barely
and/or clinical chemistry values (Dror et al. 2006; Ellsaesser
recognizable as liver because hepatocytes are severely shrunken
and Clem 1987; Fast et al. 2007; Gbore et al. 2006; Greenwell and there is a loss of vacuolation due to glycogen depletion. (E)
et al. 2003; Groff and Zinkl 1999; Scott and Ellis 2007). In Normal kidney; epithelial cells of a renal tubule (arrow) have abun-
addition, evaluation of myoglobin seems to be relevant in the dant eosinophilic (pink) cytoplasm, and hematopoietic tissue (H) is
evaluation of hypoxic stress in fish; for example, recent evi- plentiful. (F) Kidney atrophy; arrow indicates a shrunken tubule.
dence indicates that unique types of myoglobin are present (A, B): bar = 100 microns; (C–F): bar = 250 microns.
Volume 50, Number 4 2009 393almost any type of adverse condition that a fish might en-
counter or any form of outcome. Purists may argue, some-
what justifiably, that at least some of the stress responses
discussed in this review are not actually the result of “stress”
per se because they are not necessarily mediated by stress
hormones. Thus, exposure to pollutants may indeed be stress-
ful, but the associated morphologic effects may actually re-
flect tissue damage due to toxic mechanisms or specialized
physiologic adaptations to an unfavorable environment.
Notwithstanding these reasons for lack of clarity, in live
animal research it is important to recognize the potential for
Downloaded from https://academic.oup.com/ilarjournal/article-abstract/50/4/387/868673 by guest on 09 February 2020
stress, however defined, to confound a study’s results. Fail-
ure to do so is likely to lead to erroneous conclusions that
may be perpetuated in the literature. Moreover, scientists
must determine the extent to which certain effects are attrib-
utable to a particular stressor under specified conditions.
Figure 3 Pigmented macrophage aggregates (PMA). In this photomi- Further challenge studies of fish may enhance understanding
crograph of the anterior kidney from a striped bass (Morone saxatilis), of stress and its effects in fish through the administration of
PMA are compared to an early stage granuloma (EG) and a late stage
glucocorticoid or adrenergic hormones, heat shock proteins,
granuloma (LG), both of which are an inflammatory response to a
or other types of mediators not yet identified.
mycobacterial infection. The inset illustrates a PMA at higher magni-
fication. Although it is likely that a few of the macrophages in this
PMA also contain mycobacteria, in most cases PMA formation oc-
curs secondary to noninfectious causes. Bar = 50 microns. Acknowledgments
Funding for this project was provided in part by Experimen-
aggregates, PMA may also be found in the liver, heart, gonads, tal Pathology Laboratories Inc., in Sterling, Virginia.
and many other anatomic sites. The predilection for PMA to
be present in certain tissues rather than others, and the pigment
constitution of PMA, both tend to be species dependent References
(Schwindt et al. 2006). Whenever possible, PMA should be
differentiated from foci of granulomatous inflammation, which Abe T, Munehara H. 2007. Histological structure of the male reproductive
are more typically a response to microbial infection, for ex- organs and spermatogenesis in a copulating sculpin, Radulinopsis
taranetzi (Scorpaeniformes: Cottidae). Ichth Res 54:137-144.
ample. Some of the many functions attributed to PMA include Acerete L, Balasch JC, Espinosa E, Josa A, Tort L. 2004. Physiological re-
sequestration of cell breakdown products, recycling and stor- sponses in Eurasian perch (Perca fluviatilis, L.) subjected to stress by
age of iron, antigen presentation, and detoxification of exoge- transport and handling. Aquaculture 237:167-178.
nous and endogenous substances (Agius and Roberts 1981; Agius C, Roberts RJ. 1981. Effects of starvation on the melano-macrophage
Ellis 1980; Herraez and Zapata 1986; Mori 1980). PMA tend centers of fish. J Fish Biol 19:161-169.
Banerjee S, Bhattacharya S. 1994. Histopathology of kidney of Channa
to increase in number and/or size as fish age, but reports indi- punctatus exposed to chronic nonlethal level of Elsan, mercury, and am-
cate that proliferation of these structures may also occur as a monia. Ecotoxicol Environ Saf 29:265-275.
nonspecific response to various stressors, such as heat (Blazer Banerjee S, Bhattacharya S. 1995. Histopathological changes induced by
et al. 1987), starvation (Agius and Roberts 1981; Herraez and chronic nonlethal levels of elsan, mercury, and ammonia in the small in-
Zapata 1986), and nutritional imbalance (Moccia et al. 1984). testine of Channa punctatus (Bloch). Ecotoxicol Environ Saf 31:62-68.
Barton BA. 2002. Stress in fishes: A diversity of responses with particular
The potential importance of PMA as a tool for monitoring reference to changes in circulating corticosteroids. Integ Comp Biol
stress is evident in recent efforts to quantify these aggregates 42:517-525.
morphometrically in histologic sections (Jordanova et al. Barton BA, Schreck CB. 1987. Influence of acclimation temperature on in-
2008; Russo et al. 2007; Schwindt et al. 2006). terrenal and carbohydrate stress responses in juvenile chinook salmon
(Oncorhynchus tshawytscha). Aquaculture 62:299-310.
Basu N, Nakano T, Grau EG, Iwama GK. 2001. The effects of cortisol on
Conclusions heat shock protein 70 levels in two fish species. Gen Comp Endocrinol
124:97-105.
Basu N, Todgham AE, Ackerman PA, Bibeau MR, Nakano K, Schulte PM,
For several reasons, scientists’ understanding of “stress” re- Iwama GK. 2002. Heat shock protein genes and their functional signifi-
mains nebulous. First, the interplay between stressors and cance in fish. Gene 295:173-183.
stress responses is highly complex, and some stress responses Benli ACK, Köksal G, Ozkul A. 2008. Sublethal ammonia exposure of Nile
may themselves function as stressors, and vice versa. Sec- tilapia (Oreochromis niloticus L.): Effects on gill, liver and kidney his-
tology. Chemosphere 72:1355-1358.
ond, there are few, if any, pathognomonic stress responses
Blazer VS, Wolke RE, Brown J, Powell CA. 1987. Piscine macrophage ag-
(the nonspecific nature of stress responses is in keeping with gregate parameters as health monitors: Effect of age, sex, relative weight,
Selye’s original definition). A third explanation concerns the season and site quantity in largemouth bass (Micropterus salmoides).
tendency of researchers to use the term “stress” to indicate Aquat Toxicol 10:199-215.
394 ILAR JournalBuckman AH, Fisk AT, Parrot JL, Solomon KR, Brown SB. 2007. PCBs Iwama GK, Afonso LOB, Todgham A, Ackerman P, Nakano K. 2004b.
can diminish the influence of temperature on thyroid indices in rainbow Commentary: Are HSPs suitable for indicating stressed states in fish? J
trout (Oncorhynchus mykiss). Aquat Toxicol 84:366-378. Exp Biol 207:15-19.
Burgess JW, Coss RG. 1982. Effects of chronic crowding stress on midbrain Jordanova M, Miteva N, Rocha E. 2008. A qualitative and quantitative study
development: Changes in dendritic spine density and morphology in of the hepatic pigmented macrophage aggregates during the breeding
jewel fish optic tectum. Dev Psychobiol 15:461-470. cycle of Ohrid trout, Salmo letnica Kar. (Teleostei, Salmonidae). Mi-
Chan CY, Lam WP, Wai MSM, Wang M, Foster EL, Yew DT-W. 2007. Peri- crosc Res Tech 71:822-830.
natal hypoxia induces anterior chamber changes in the eyes of offspring Kent ML, Hedrick RP. 1987. Effects of cortisol implants on the PKX myxo-
fish. J Reprod Dev 53:1159-1167. sporean causing proliferative kidney disease in rainbow trout, Salmo
Chapman LJ, Galis F, Shinn J. 2000. Phenotypic plasticity and the possible gairdneri. J Parasitol 73:455-461.
role of genetic assimilation: Hypoxia-induced trade-offs in the morpho- Kurata O, Okamoto N, Suzumura E, Sano N, Ikeda Y. 1995. Accommoda-
logical traits of an African cichlid. Ecol Lett 3:387-393. tion of carp natural killer-like cells to environmental temperatures.
Choi K, Lehmann DW, Harms CA, Law JM. 2007. Acute hypoxia-reperfusion Aquaculture129:421-424.
Downloaded from https://academic.oup.com/ilarjournal/article-abstract/50/4/387/868673 by guest on 09 February 2020
triggers immunocompromise in Nile tilapia. J Aquat Anim Health 19: Lease HM, Hansen JA, Bergman HL, Meyer JS. 2003. Structural changes in
128-140. gills of Lost River suckers exposed to elevated pH and ammonia concen-
Cleary JJ, Battaglene SC, Pankhurst NW. 2002. Capture and handling stress trations. Comp Biochem Physiol C Toxicol Pharmacol 134:491-500.
affects the endocrine and ovulatory response to exogenous hormone Leino RL, Jensen KM, Ankley GT. 2005. Gonadal histology and characteris-
treatment in snapper, Pagrus auratus (Bloch & Schneider). Aquacult tic histopathology associated with endocrine disruption in the adult fat-
Res 33:829-838. head minnow (Pimephales promelas). Env Toxicol Pharmacol 19:85-98.
Contreras-Sánchez WM, Schreck CB, Fitzpatrick MS, Pereira CB. 1998. Marques IJ, Leito JTD, Spaink HP, Testerink J, Jaspers RT, Witte F, van den
Effects of stress on the reproductive performance of rainbow trout (On- Berg S, Bagowski CP. 2008. Transcriptome analysis of the response to
corhynchus mykiss). Biol Reprod 58:439-447. chronic constant hypoxia in zebrafish hearts. J Comp Physiol B 178:77-92.
Cossins AR, Williams DR, Foulkes NS, Berenbrink M, Kipar A. 2009. Diverse Mori M. 1980. Studies on the phagocytic system in goldfish. I. Phagocyto-
cell-specific expression of myoglobin isoforms in brain, kidney, gill and sis of intraperitoneally injected carbon particles. Fish Pathol 15:25-30.
liver of the hypoxia-tolerant carp and zebrafish. J Exp Biol 212:627-638. Moccia RD, Hung SSO, Slinger SJ, Ferguson SW. 1984. Effect of oxidized
Daoust PY, Ferguson HW. 1983. Gill diseases of cultured salmonids in On- fish oil, vitamin E and ethoxyquin on histopathology and haematology
tario. Can J Comp Med 47:358-362. of rainbow trout, Salmo gairdneri Richardson. J Fish Dis 7:269-282.
Dey S, Bhattacharya S. 1989. Ovarian damage to Channa punctatus after Nesse RM, Young EA. 2000. Evolutionary origins and functions of the
chronic exposure to low concentrations of elsan, mercury, and ammo- stress response. In: Fink G, ed. Encyclopedia of Stress, vol 2. San Di-
nia. Ecotoxicol Env Saf 17:247-257. ego: Academic Press. p 79-84.
Dutta T, Acharya S, Das MK. 2005. Impact of water quality on the stress Noga EJ. 1996. Fish Disease: Diagnosis and Treatment. St. Louis: Mosby.
physiology of cultured Labeo rohita (Hamilton-Buchanan). J Environ Noga EJ, Botts S, Yang MS, Avtalion R. 1998. Acute stress causes skin ulcer-
Biol 26:585-592. ation in striped bass and hybrid bass (Morone). Vet Pathol 35:102-107.
Ellis AE. 1980. Antigen-trapping in the spleen and kidney of the plaice Norris DO. 2000. Endocrine disruptors of the stress axis in natural popula-
Pleuronectes platessa L. J Fish Dis 3:413-426. tions: How can we tell? Amer Zool 40:393-401.
Frances J, Nowak BF, Allan GL. 2000. Effects of ammonia on juvenile sil- Olsen RE, Sundell K, Ringø E, Mykelbust R, Hemre G-I, Hansen T, Karlsen
ver perch (Bidyanus bidyanus). Aquaculture 183:95-103. Ø. 2008. The acute stress response in fed and food-deprived Atlantic
Geng YJ. 2003. Molecular mechanisms for cardiovascular stem cell apopto- cod, Gadus morhua L. Aquaculture 280:232-241.
sis and growth in the hearts with atherosclerotic coronary disease and Ortega VA, Renner KJ, Bernier NJ. 2005. Appetite-suppressing effects of
ischemic heart failure. Ann N Y Acad Sci 1010:687-697. ammonia exposure in rainbow trout associated with regional and tempo-
Gieseker CM, Serfling SG, Reimschuessel R. 2006. Formalin treatment to ral activation of brain monoaminergic and CRF systems. J Exp Biol
reduce mortality associated with Saprolegnia parasitica in rainbow 208:1855-1866.
trout, Oncorhynchus mykiss. Aquaculture 253:120-129. Ortuño J, Esteban MA, Meseguer J. 2001 Effects of short-term crowding
Gingerich WH. 1982. Hepatic toxicology of fishes. In: Weber LJ, ed. stress on the gilthead seabream (Sparus aurata L.) innate immune re-
Aquatic Toxicology. New York: Raven Press. p 55-105. sponse. Fish Shell Immunol 11:187-197.
Gratzek JB, Reinert R. 1984. Physiological responses of experimental fish Pickering AD. 1981. Stress and Fish. London: Academic Press. p 367.
to stressful conditions. Natl Cancer Inst Monogr 65:187-193. Perry SF. 1998. Relationships between branchial chloride cells and gas
Greenwell MG, Sherrill J, Clayton LA. 2003. Osmoregulation in fish mech- transfer in freshwater fish. Comp Biochem Physiol A Mol Integr Physiol
anisms and clinical implication. Vet Clin Exot Anim 6:169-189. 119:9-16.
Harms CA, Sullivan CV, Hodson RG, Stoskopf MK. 1996. Clinical pathol- Randall DJ, Tsui TK. 2002. Ammonia toxicity in fish. Mar Pollut Bull
ogy and histopathology characteristics of net-stressed striped bass with 45:17-23.
red-tail. J Aquat Anim Health 8:82-86. Rasmussen TH, Teh SJ, Bjerregaard P, Korsgaard B. 2005. Anti-estrogen
Harris PD, Soleng A, Bakke TA. 2000. Increased susceptibility of salmo- prevents xenoestrogen-induced testicular pathology of eelpout (Zoarces
nids to the monogenean Gyrodactylus salaris following administration viviparus). Aquat Toxicol 72:177-194.
of hydrocortisone acetate. Parasitology 120:57-64. Roberts F. 1950. Correspondence regarding: “Stress and the General Adap-
Heiden TK, Carvan MJ III, Hutz RJ. 2006. Inhibition of follicular develop- tation Syndrome,” BMJ 2:104. Cited by: Rosch PJ. 1958. Growth and
ment, vitellogenesis, and serum 17 b-estradiol concentrations in zebrafish development of the stress concept and its significance in clinical
following chronic, sublethal dietary exposure to 2,3,7,8-tetrachlorodibenzo- medicine. In: Gardiner-Hill H, ed. Modern Trends in Endocrinology.
p-dioxin. Toxicol Sci 90:490-499. London: Butterworth & Co.
Herraez MP, Zapata AG. 1986. Structure and function of the melano- Russo R, Yanong RP, Terrell SP. 2007. Preliminary morphometrics of spleen
macrophage centres of the goldfish Carassius auratus. Vet Immunol and kidney macrophage aggregates in clinically normal blue gourami
Immunop 12:117-126. Trichogaster trichopterus and freshwater angelfish Pterophyllum scalare.
Hosoya S, Johnson SC, Iwama GK, Gamperl AK, Afonso LO. 2007. J Aquat Anim Hlth 19:60-67.
Changes in free and total plasma cortisol levels in juvenile haddock Sardella BA, Matey V, Cooper J, Gonzalez RJ, Brauner CJ. 2004. Physio-
(Melanogrammus aeglefinus) exposed to long-term handling stress. logical, biochemical and morphological indicators of osmoregulatory
Comp Biochem Physiol A Mol Integr Physiol 146:78-86. stress in ‘California’ Mozambique tilapia (Oreochromis mossambicus ×
Iwama GK, Afonso LOB, Vijayan MM. 2004a. Stress in fish. AquaNet Work- O. urolepis hornorum) exposed to hypersaline water. J Exp Biol
shop on Fish Welfare, Campbell River, BC, Canada. 27 September. 207:1399-1413.
Volume 50, Number 4 2009 395Schwindt AR, Truelove N, Schreck CB, Fournie JW, Landers DH, Kent ent levels of vitamins E and C and highly unsaturated fatty acids
ML. 2006. Quantitative evaluation of macrophage aggregates in brook (HUFA). Aquaculture 277:293-302.
trout Salvelinus fontinalis and rainbow trout Oncorhynchus mykiss. Dis Tsunoda A, Purbayanto A, Akiyama S, Arimoto T. 1999. Plasma cortisol
Aquat Organ 68:101-113. level for stress management of Japanese whiting Sillago japonica
Scott AL, Rogers WA. 1980. Histological effects of prolonged sublethal captured by sweeping trammel net. Nippon Suisan Gakkaishi
hypoxia on channel catfish Ictalurus punctatus (Rafinesque). J Fish Dis 65:457-463.
3:305-316. Udomkusonsri P, Noga EJ, Monteiro-Riviere NA. 2004. Pathogenesis of
Scott AP, Ellis T. 2007. Measurement of fish steroids in water. Gen Comp acute ulceration response (AUR) in hybrid striped bass. Dis Aquat Or-
Endocrinol 153:392-400. gan 61:199-213.
Selye H. 1936. A syndrome produced by diverse nocuous agents. Nature van der Meer DL, van den Thillart GE, Witte F, de Bakker MA, Besser J,
138:32-34. Richardson MK, Spaink HP, Leito JT, Bagowski CP. 2005. Gene ex-
Selye H. 1955. Stress and disease. Science 122:625-631. pression profiling of the long-term adaptive response to hypoxia in the
Sloman KA, Gilmour KM, Metcalfe NB, Taylor AC. 2005. Does socially gills of adult zebrafish. Am J Physiol Regul Integr Comp Physiol
induced stress in rainbow trout cause chloride cell proliferation? J Fish 289:R1512-R1519.
Downloaded from https://academic.oup.com/ilarjournal/article-abstract/50/4/387/868673 by guest on 09 February 2020
Biol 56:725-738. Via JD, Villani P, Gasteiger E, Niederstätter H. 1998. Oxygen consumption
Small BC. 2004. Effect of isoeugenol sedation on plasma cortisol, glucose, in sea bass fingerling Dicentrarchus labrax exposed to acute salinity
and lactate dynamics in channel catfish Ictalurus punctatus exposed to and temperature changes: Metabolic basis for maximum stocking den-
three stressors. Aquaculture 238:469-481. sity estimations. Aquaculture 169:303-313.
Sollid J, De Angelis P, Gundersen K, Nilsson GE. 2003. Hypoxia induces Vijayan MM, Pereira C, Grau EG, Iwama GK. 1997. Metabolic responses
adaptive and reversible gross morphological changes in crucian carp associated with confinement stress in tilapia: the role of cortisol. Comp
gills. J Exp Biol 206:3667-3673. Biochem Physiol Pharmacol Toxicol Endocrinol 116:89-95.
Spencer P, Pollock R, Dubé M. 2008. Effects of un-ionized ammonia on Wang S, Yuen SS, Randall DJ, Hung CY, Tsui TK, Poon WL, Lai JC,
histological, endocrine, and whole organism endpoints in slimy sculpin Zhang Y, Lin H. 2008. Hypoxia inhibits fish spawning via LH-depen-
(Cottus cognatus). Aquat Toxicol 90:300-309. dent final oocyte maturation. Comp Biochem Physiol Toxicol Pharma-
Teh SJ, Adams SM, Hinton DE. 1997. Histopathologic biomarkers in feral col 148:363-369.
freshwater fish populations exposed to different types of contaminant Wedemeyer GA. 1969. Stress-induced ascorbic acid depletion and corti-
stress. Aquat Toxicol 37:51-57. sol production in two salmonid fishes. Comp Biochem Physiol
Thomas P, Rahman MS, Khan IA, Kummer JA. 2007. Widespread endo- 29:1247-1251.
crine disruption and reproductive impairment in an estuarine fish popu- Wolf JC, Wolfe MJ. 2005. A brief overview of nonneoplastic hepatic toxic-
lation exposed to seasonal hypoxia. Proc Royal Soc B 274:2693-2701. ity in fish. Toxicol Pathol 33:75-85.
Trenzado CE, Morales AE, Higuera M. 2008. Physiological changes in Wolke RE. 1992. Piscine macrophage aggregates: A review. Annu Rev Fish
rainbow trout held under crowded conditions and fed diets with differ- Dis 2:91-108.
396 ILAR JournalYou can also read

















































