MAGE-A1, GAGE and NY-ESO-1 cancer/testis antigen expression during human gonadal development
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Human Reproduction Vol.22, No.4 pp. 953–960, 2007 doi:10.1093/humrep/del494
Advance Access publication January 5, 2007
MAGE-A1, GAGE and NY-ESO-1 cancer/testis antigen
expression during human gonadal development
Morten F.Gjerstorff1, Kirsten Kock2, Ole Nielsen2 and Henrik J.Ditzel1,3,4
1
Medical Biotechnology Center, Institute of Medical Biology, University of Southern Denmark, Odense C, Denmark, 2Departments of
Clinical Pathology and 3Oncology, Odense University Hospital, Odense C, Denmark
4
To whom correspondence should be addressed at: Medical Biotechnology Center, Institute of Medical Biology, University of Southern
Downloaded from https://academic.oup.com/humrep/article/22/4/953/698623 by guest on 06 February 2022
Denmark, Winsloewparken 25, 3., DK-5000 Odense C, Denmark. E-mail: hditzel@health.sdu.dk
BACKGROUND: Cancer/testis antigens (CTAs) are expressed in several cancers and during normal adult male
germ cell differentiation. Little is known about their role in fetal development of human germ cells. METHODS:
We examined expression of the CTAs MAGE-A1, GAGE and NY-ESO-1 in fetal gonads by single and double immu-
nohistochemical staining. RESULTS: We found that GAGE was expressed in the primordial germ cells of the gonadal
primordium, whereas MAGE-A1 and NY-ESO-1 were first detected in germ cells of both testis and ovary after sexual
differentiation was initiated. The number of positive germ cells and the staining intensity of all three CTAs peaked
during the second trimester and gradually decreased towards birth in both male and female germ cells. In oocytes,
MAGE-A1 expression terminated around birth, whereas NY-ESO-1 expression persisted through the neonatal
stage and GAGE expression was maintained until adulthood. The population of GAGE-expressing male and
female germ cells partially overlapped the population of OCT4-positive cells, whereas MAGE-A1 and NY-ESO-1
were clearly expressed only by OCT4-negative cells. CONCLUSIONS: Our results suggest that MAGE-A1 and
NY-ESO-1 are associated with highly proliferating germ cells, whereas GAGE proteins have a more general function
in germ cells unrelated to any specific developmental stage. The recognition of differential cellular expression of
GAGE, MAGE-A1, NY-ESO-1 and OCT4 may help define biologically distinct germ cell subpopulations.
Key words: fetal germ cells/germ cell subpopulations/immunohistochemistry/OCT4/primordial germ cells
Introduction have emerged indicating that several members of the MAGE
Cancer/testis antigens (CTAs) are comprised of at least 44 family (MAGE-A1 and -A4) interact with the nuclear proteins
distinct families that are traditionally considered to be SKIP and histone deacetylase 1, thereby inhibiting transcrip-
expressed only in normal adult testis germ cells and in different tional activation mediated by Notch-IC (Laduron et al.,
types of cancer. Most CTA genes are located in clusters at 2004). Moreover, MAGE-A2 was observed to down-regulate
chromosome X, comprising multiple genes of high identity p53 transactivation by recruiting histone deacetylase 3
(e.g. MAGE-A, GAGE and SSX), whereas others exist as auto- (Monte et al., 2006). GAGE proteins have been shown to loca-
somally encoded single genes (Simpson et al., 2005). lize to the nucleus of a subset of spermatogonia and cancer
Spermatogonia are testicular stem cells that can proliferate cells, indicating that GAGE may also be a regulator of gene
throughout life to maintain the stem cell pool or undergo differ- expression (Gjerstorff et al., 2006).
entiation to produce spermatozoa. This process includes two The primordial germ cells (PGCs) are the progenitors of both
meiotic divisions from tetraploid primary spermatocytes to male and female germ cells, and they migrate from the dorsal
diploid secondary spermatocytes to haploid spermatids that, yolk sac via the dorsal mesentery to arrive at the indifferent
in turn, develop into spermatozoa (Sadler, 1985). Many gonad during the fifth week of gestation (Burger and de
CTAs, including most of the chromosome X-encoded CTAs, Kretser, 1981). In the male gonad, the PGCs become enclosed
such as MAGE-A, GAGE and NY-ESO-1, are found in sper- by Sertoli cells in the seminiferous cords, where they acquire a
matogonia and primary spermatocytes, whereas a smaller different morphology and become gonocytes. The gonocytes
number of CTAs have been identified in the more differentiated proliferate and turn into prospermatogonia, which then arrest
stages of spermatogenesis (Simpson et al., 2005). in the G0/G1 phase until spermatogenesis is initiated at
The functional biology of germ cells and tumour cells puberty (Burger and de Kretser, 1981). In the female gonad,
remains to be determined for most CTAs, although clues the PGCs are the progenitors of oogonia, which increase in
# The Author 2007. Published by Oxford University Press on behalf of the European Society of Human Reproduction and Embryology. 953
All rights reserved. For Permissions, please email: journals.permissions@oxfordjournals.org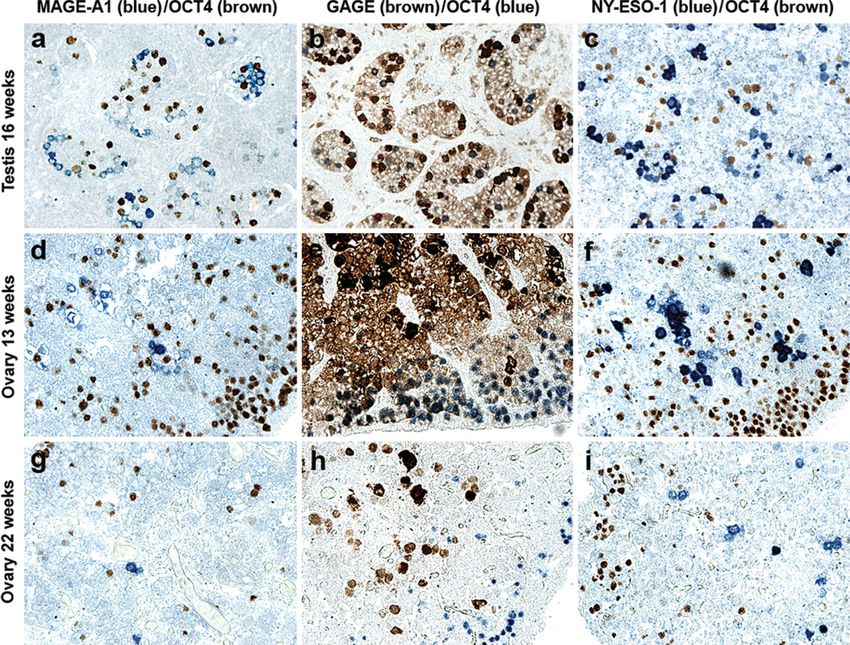
M.F.Gjerstorff et al.
number during the first and second trimesters by mitotic cell microwave boiling for 15 min in (i) T-EG buffer (10 mM Tris, 0.5 mM
divisions. This proliferation coincides with a major wave of EGTA, pH 9.0), (ii) 10 mM citrate buffer, pH 6.0 or (iii) Dako Target
cell degeneration by apoptosis (McGee et al., 1998; McGee, retrieval solution (Dako S1699), or proteolytic treatment using (iv)
2000). At the end of the first trimester, the epithelial precursors 0.05% protease type XIV (pronase E, Sigma; cat. no. P5147) in
TBS, pH 7.4 for 15 min at 378C or (v) 0.4% pepsin (Sigma; cat. no.
of human granulosa cells begin to encircle the oogonia, and at
P7012) in 0.01 M HCl for 20 min at 378C. Microwave boiling in
birth all female germ cells are enclosed in primordial follicles.
T-EG buffer for 15 min proved to be the optimal antigen retrieval
During the second trimester, the oogonia develop into oocytes method for anti-GAGE, anti-MAGE-A1 and anti-NY-ESO-1 mAbs
when they initiate meiosis to arrest in the prophase of the first and was used in the successive experiments. Sections were sub-
meiotic division. These cells constitute the resting pool of adult sequently incubated with anti-GAGE mAb M3 [1/100; produced
female germ cells and will complete the meiotic division just in-house (Gjerstorff et al., 2006)], anti-GAGE-7 mAb (1/2000;
prior to ovulation (Sadler, 1985). Clone 26, BD Biosciences), anti-MAGE-A1 mAb (1/200; Clone
Little is known about the complex processes of human sex MA454, Lab Vision Corporation, Newmarket Suffolk, UK),
determination and gonadal differentiation, although it has anti-NY-ESO-1 mAb (1/25; Clone E978, Zymed Laboratories Inc.,
been shown that human fetal germ cells are heterogeneous in San Francisco, CA, USA), anti-p450scc pAb (1/8.000; Corgen Inc.,
Downloaded from https://academic.oup.com/humrep/article/22/4/953/698623 by guest on 06 February 2022
terms of morphology and marker expression (Wartenberg Taipei, Taiwan), anti-SF-1 pAb (1/500; a gift from Professor
Ken-Ichirou Morohashi, National Institute of Natural Sciences,
et al., 1998; Gaskell et al., 2004). To enhance our understand-
Aichi, Japan) or anti-OCT3/4 mAb (1/400; Clone C10, Santa Cruz
ing of the maturation process, the identification of further
Biotechnology, Heidelberg, Germany) diluted in antibody diluent
markers that can distinguish between germ cells with distinct (S2022, DAKO, Glostrup, Denmark) for 1 h at room temperature.
biological properties is needed. Observations of MAGE-A4 Sections were washed with TNT buffer and incubated with horse-
and NY-ESO-1 expression in fetal gonocytes (Satie et al., radish peroxidase (HRP)-conjugated ‘Ready-to-use’ EnVisionTM þ
2002; Rajpert-De Meyts et al., 2003; Gaskell et al., 2004; polymer K4001 (mouse mAb) or polymer K4003 (rabbit pAb)
Pauls et al., 2006; Rajpert-De Meyts, 2006) have suggested (DAKO) for 30 min, followed by another wash with TNT buffer.
that at least some CTAs play a role in the development and The final reaction product was visualized by incubating with
differentiation of fetal germ cells, but a systematic evaluation 3,30 -diaminobenzidine (DAB) þ substrate –chromogen (DAKO) for
of CTAs in fetal gonads has never been conducted. 10 min, followed by washing with H2O and counterstaining of sec-
In this study, we characterized the protein expression pat- tions with Mayers hematoxylin before mounting in AquaTex (Merck
Inc., Whitehouse Station, NJ, USA). A significant portion of the speci-
terns of three different types of CTAs, e.g. MAGE-A1,
mens were examined twice with similar results. For each experiment,
GAGE and NY-ESO-1, in gonads of first and second trimester
samples of either an isotype-matched antibody or no primary antibody
fetuses. Using immunohistochemical double staining, the cel- were included as controls. For the immunohistochemical double stain-
lular expression of these proteins was further correlated to ing, antigen retrieval and primary antibody incubations were carried
the expression of the pluripotency marker OCT4 and to the out as above. The first layer primary antibody incubation was followed
gonadal parenchymal cell markers p450scc and steroidogenic by incubations with alkaline phosphatase conjugated PowerVision
factor 1 (SF-1). polymer (ImmunoVision Technologies, Brisbane, CA, USA) for 30
min and Vector Blue substrate-chromogen (Vector Laboratories,
Burlingame, CA, USA) for 20 min, whereas the second layer
Materials and methods primary antibody was followed by incubations with HRP-conjugated
Specimens EnVision polymerTM + K4001 (mouse mAb) or K4003 (rabbit
Tissues were obtained from abortion specimens collected for diagnos- pAb) (DAKO) for 30 min and DAB + substrate-chromogen
tic purposes and deposited in the tissue bank of Odense University (DAKO) for 10 min. A 15 min microwave boiling in T-EG buffer
Hospital. The tissues included were indifferent human embryonic was carried out between the two antibody-staining sequences to
gonads (weeks 6 and 8), fetal testis [weeks 9 (n ¼ 1), 12 (n ¼ 1), 13 block antibody cross-reactivity. Stained sections were mounted in
(n ¼ 1), 14 (n ¼ 3), 15 (n ¼ 1), 16 (n ¼ 2), 17 (n ¼ 2), 18 (n ¼ 2), AquaTex (Merck Inc.).
19 (n ¼ 2), 20 (n ¼ 2), 21 (n ¼ 1), 22 (n ¼ 2), 23 (n ¼ 2) and 27
(n ¼ 1)] and fetal and neonatal ovaries [weeks 13 (n ¼ 1), 17 Morphometric quantification
(n ¼ 1), 18 (n ¼ 1), 19 (n ¼ 3), 22 (n ¼ 2), 28 (n ¼ 1), 29 (n ¼ 1), The frequencies of GAGE-, MAGE-A1- or NY-ESO-1-positive
34 (n ¼ 1), 38 (n ¼ 1), 39 (n ¼ 1) and 41 (n ¼ 1)]. The study was female germ cells in third trimester and neonatal specimens were
approved by the ethical committee of Funen and Vejle County calculated as the mean of the frequencies of positive cells (number
(VF20050069). of positive cells/number of germ cells) in three to five randomly
True fetal age was deduced from the gestational age (e.g. the time chosen microscope frames using the 20 microscope lens.
after the first day of the last period), supplemented with physiological
data (e.g. fetal weight, fetal length, organ weight, etc.). Results
Immunohistochemistry Protein expression of MAGE-A1, NY-ESO-1 and GAGE was
Tissue biopsies were fixed in 4% formaldehyde (pH 7.4) for 24 h and evaluated by immunohistochemistry on indifferent human
paraffin-embedded. Sections of tissues were cut, deparaffinized, embryonic gonads (6 – 8 weeks), fetal testis (9 – 27 weeks)
treated with 1.5% H2O2 in Tris-buffered saline (TBS; pH 7.5) for and fetal and neonatal ovaries (13– 41 weeks).
10 min to block endogenous peroxidase activity, rinsed in distilled
H2O, demasked, processed for antigen retrieval and washed in TNT GAGE expression in PGCs of the indifferent gonad
buffer (0.1 M Tris, 0.15 M NaCl, 0.05% Tween-20, pH 7.5). A In the gonadal primordium of a 6-week-old embryo, PGCs
panel of antigen retrieval protocols was initially evaluated, including derived from the dorsal yolk sac exhibited GAGE expression
954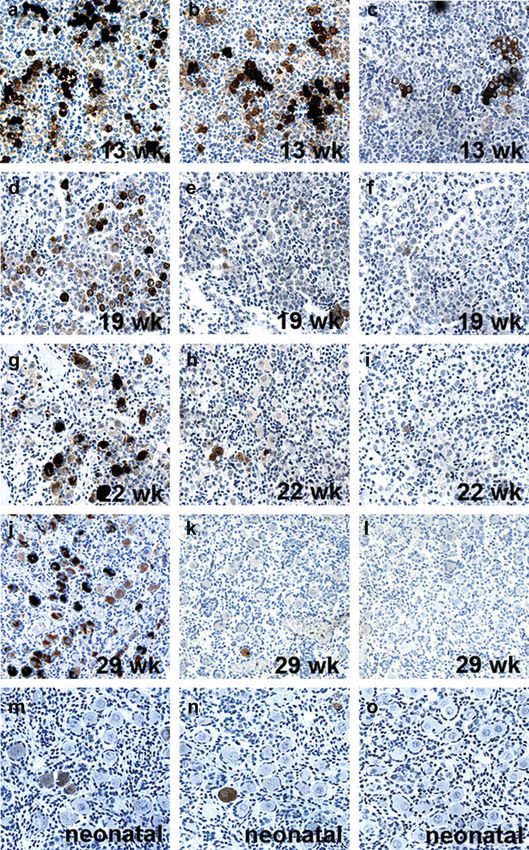
Cancer testis antigen expression in fetal gonads
the cells of the connective tissue were negative for all three
CTAs in all sections. The identity of the GAGE-positive
Leydig and Sertoli cells was confirmed by immunohistochem-
ical double staining, showing that the GAGE-expressing cells
also expressed p450scc (Figure 2m) and SF-1 (Figure 2n).
Ontogenic expression of MAGE-A1, GAGE and
NY-ESO-1 in human ovary
Since male and female germ cells are derived from the
same progenitor cells and undergo similar prenatal prolifer-
ation and differentiation processes, we examined whether
CTAs were also expressed in early female germ cells.
Second trimester ovary sections (13, 17, 18, 19 and 22
weeks) were found to contain MAGE-A1-, GAGE- and
Downloaded from https://academic.oup.com/humrep/article/22/4/953/698623 by guest on 06 February 2022
NY-ESO-1-positive germ cells (Figure 3). Strong expression
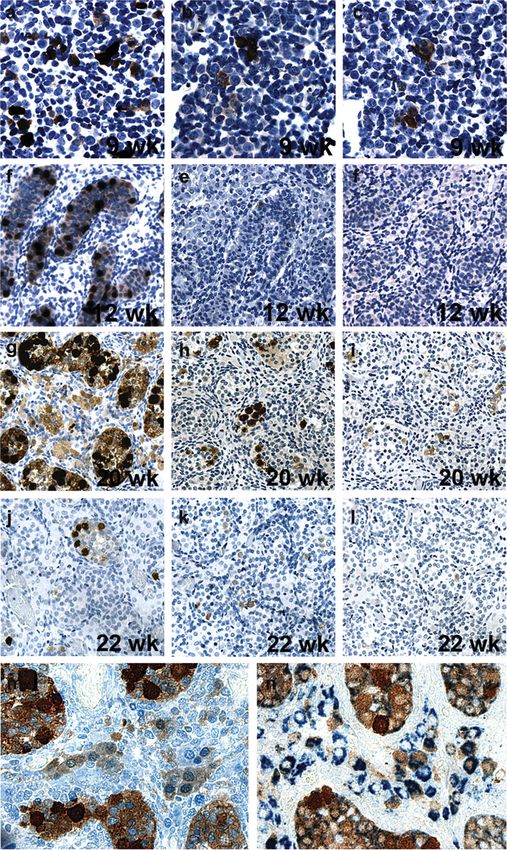
Figure 1. Expression of GAGE in primordial germ cells (PGCs). of all CTAs was observed in most cells at week 13
GAGE was expressed in the PGCs of the gonadal primordium of (Figure 3a –c), whereas both the number of cells positive for
6-(a) and 8-week-old (b) embryos. MAGE-A1 (c) and NY-ESO-1 MAGE-A1 and NY-ESO-1 and the staining intensity of the
(d) were not detected in the embryonal gonad. No staining was
observed using the negative control antibody (Magnification: 40). positive germ cells were clearly reduced at weeks 17, 18, 19
and 22 compared with week 13 (Figure 3e, f, h and i). In con-
trast, GAGE expression remained unchanged in the number of
(Figure 1a), whereas no MAGE-A1 and NY-ESO-1 expression positive cells and the staining intensity (Figure 3d and g).
was observed. At 8 weeks, the PGCs, now associated with the Similar to male germ cells, GAGE staining of female germ
primitive sex cords, remained GAGE-positive (Figure 1b) and cells was localized to the cytoplasm and the nucleus, whereas
MAGE-A1- and NY-ESO-1-negative (Figure 1c and d). No MAGE-A1 and NY-ESO-1 staining was detected only in the
MAGE-A1, GAGE and NY-ESO-1 staining was observed in cytoplasm (Figure 3). Female germ cells, exhibiting strong
the gonadal mesenchymal cells. At 6 and 8 weeks, only a expression of CTAs, were often located in clusters
subset of the PGCs was GAGE-positive and varied in staining (Figure 3a – c).
intensity. In the third trimester specimens (weeks 28 and 29), most
germ cells (,95%) remained GAGE-positive, whereas 3%
Ontogenic expression of MAGE-A1, GAGE and were NY-ESO-1-positive. A few MAGE-A1-positive oocytes
NY-ESO-1 in human testis (,1%) were detected at week 28, whereas the 29-week-old
Germ cell expression of MAGE-A1, GAGE and NY-ESO-1 specimen was negative. Moreover, among the resting
was further examined in first, second and third trimester fetal primordial follicles of neonatal ovaries (weeks 34, 38, 39 and
testis (Figure 2). At week 9, germ cells were strongly positive 41), the number of GAGE-positive cells was reduced to
for all three CTAs (Figure 2a – c), but while GAGE expression 11%, while still 3% of oocytes were NY-ESO-1-positive
was continuously strong throughout the fetal period, (Figure 3j –o). In neonatal ovaries, MAGE-A1 was detected
MAGE-A1 and NY-ESO-1 expression was absent (weeks 12, at 34 and 41 weeks (,1%), but not at 38 and 39 weeks.
14 and 15) or weak (weeks 13 and 14) in the beginning of
the second trimester (Figure 2d – f). In the middle and end Co-expression studies of OCT4 and MAGE-A1, NY-ESO-1
of the second trimester, a peak in staining intensity for or GAGE in the normal fetal gonad
MAGE-A1 and NY-ESO-1 (weeks 16, 17, 18 and 20) was The pluripotency marker OCT4 has been shown to be
followed by a reduction in the number of positive cells and expressed in migrating PGCs, and a large number of germ
in the intensity of cellular staining (weeks 21, 22 and 23) cells in both the testis and ovary remain OCT4-positive until
(Figure 2g– l). A third trimester specimen (27 weeks) exhibited the beginning of the second trimester, where the number of
GAGE and MAGE-A1 but not NY-ESO-1 expression. The positive cells and the staining intensity decline (Rajpert-De
subcellular localization of MAGE-A1 and NY-ESO-1 staining Meyts et al., 2004). We compared the OCT4 immunolocaliza-
in fetal male germ cells was primarily cytoplasmic, tion to that of MAGE-A1, NY-ESO-1 and GAGE to identify
whereas GAGE was localized to both the cytoplasm and the germ cell subpopulations (Figure 4). As expected, a subset of
nucleus. germ cells of both second trimester male and female gonads
Interestingly, we found that most Sertoli cells and a subset of was OCT4-positive (weeks 13– 22), and the frequency of posi-
the interstitial Leydig cells, in addition to the prospermatogo- tive cells declined gradually with age. Double immunohisto-
nia, were GAGE-positive at weeks 14– 20 (Figure 2g), chemical staining revealed that a subset of the OCT4-positive
whereas these cells were GAGE-negative at all other ages. germ cells in both male and female gonads also expressed
The GAGE staining intensity in Sertoli and Leydig cells was GAGE (Figure 4b, e and h). Interestingly, not all GAGE-
less pronounced than that of the germ cells and was confined positive cells were OCT4-positive, demonstrating the existence
to the cytoplasm. No expression of MAGE-A1 and of three different populations of cells: OCT4pos/GAGEpos,
NY-ESO-1 was ever observed in Sertoli and Leydig cells, and OCT4neg/GAGEpos and OCT4pos/GAGEneg. Double-staining
955
M.F.Gjerstorff et al.
Downloaded from https://academic.oup.com/humrep/article/22/4/953/698623 by guest on 06 February 2022
Figure 2. Expression of GAGE, MAGE-A1 and NY-ESO-1 in fetal testis. GAGE (a, d, g and j), MAGE-A1 (b, e, h and k) and NY-ESO-1 (c, f, i
and l) were detected in a subset of testicular germ cells of 9– 22-week-old fetuses. A peak in expression was observed at weeks 16 –20, whereas
MAGE-A1 and NY-ESO-1 expression was significantly reduced by week 22. Double immunohistochemical staining combining GAGE (DAB/
brown stain) and p450scc (Vector Blue/blue stain) (m) or Steroidogenic factor 1 (Vector Blue/blue stain) (n) showed that GAGE was also
expressed in some Sertoli and Leydig cells during weeks 16 –20. No staining was observed using the negative control antibody [Magnification:
40 (a –c, m and n), 10 (d–l)].
combining OCT4 and MAGE-A1 or NY-ESO-1 staining germ cells positive for any of the three CTAs was observed.
clearly showed that these proteins were never expressed in The OCT4-positive germ cells tended to be located in the per-
the same germ cells (Figure 4a, c, d, f, g and i). ipheral part of the ovary, whereas the CTA-positive germ cells
In ovaries, a clear difference in the macroscopic tissue were located primarily in the central part of the ovary
distribution of OCT4-positive germ cells compared with (Figure 4d– i).
956Cancer testis antigen expression in fetal gonads
Downloaded from https://academic.oup.com/humrep/article/22/4/953/698623 by guest on 06 February 2022
Figure 3. Expression of GAGE, MAGE-A1 and NY-ESO-1 in the developing fetal ovary. GAGE (a, d, g and j), NY-ESO-1 (b, e, h and k) and
MAGE-A1 (c, f, i and l) were detected in a subset of germ cells in the ovary of 13–29-week-old fetuses. Expression of all three CTAs was stron-
gest at 13 weeks, and while GAGE expression was detected in most germ cells up to week 29, MAGE-A1 and NY-ESO-1 expression was sig-
nificantly reduced by the middle of second trimester. GAGE (m) and NY-ESO-1 (n) expression persisted in 11% and 3% of oocytes of
neonatal primordial follicles, respectively, whereas no or very few MAGE-A1-positive oocytes (,1%) were detected either at this stage (o)
or in the third trimester specimens (l). No staining was observed using the negative control antibody (Magnification: 10).
of CTAs, we evaluated their expression in human embryonic
Discussion and fetal gonads.
To enhance our understanding of the function of MAGE-A1, In the gonadal primordium of the first trimester embryos (6
NY-ESO-1 and GAGE, members of three different subfamilies and 8 weeks), a subset of PGCs was strongly positive for
957M.F.Gjerstorff et al.
Downloaded from https://academic.oup.com/humrep/article/22/4/953/698623 by guest on 06 February 2022
Figure 4. Identification of subpopulations of fetal germ cells by double immunostaining of OCT4 and MAGE-A1, GAGE or NY-ESO-1. The
GAGE-positive cell population partially overlapped the population of OCT4-positive cells, allowing the identification of three subpopulations
of germ cells: OCT4pos/GAGEpos, OCT4neg/GAGEpos and OCT4pos/GAGEneg (b, e and h). In contrast, dual staining combining OCT4- and
MAGE-A1- or NY-ESO-1-reactivity clearly showed that OCT4 and MAGE-A1 or NY-ESO-1 were not co-expressed (a, c, d, f, g and i). No stain-
ing was observed using the negative control antibody (Magnification: 10).
GAGE, but not MAGE-A1 or NY-ESO-1. The PGCs migrate cells and is not associated with a specific developmental step in
from the dorsal yolk sac and arrive at the indifferent gonad at germ cell differentiation.
the fifth week of gestation, where they reside in the sex cords In addition to germ cells, GAGE was also detected in both
(Sadler, 1985). Although not conclusive, the finding of Leydig and Sertoli cells in 14– 20-week-old testis. This was
GAGE-negative PGCs in the gonadal primordium together surprising, since the ontogeny of these cells differs from that
with the lack of GAGE-positive migrating PGCs suggests of PGCs, although the exact origin of these three cell types
that PGCs do not express GAGE until embedded in the in humans is controversial. The PGCs are known to be
gonadal mesenchyme. derived from cells of the yolk sac (Burger and de Kretser,
High GAGE expression was seen in a subset of both male 1981), whereas the coelomic epithelium or the mesonephos
and female germ cells throughout the second trimester. have been suggested as the source of Leydig cells (O’Shaugh-
GAGE proteins were localized to both cytoplasm and nessy et al., 2006). The coelomic epithelium also seems to give
nucleus, similar to the subcellular localization in spermatogo- rise to the Sertoli cells (Karl and Capel, 1998), and the coordi-
nia of adult testis (Gjerstorff et al., 2006). GAGE expression nated expression of GAGE in fetal Leydig and Sertoli cells may
was found to persist in 11% of primordial follicles of the neo- indicate a common origin. It is also possible that local factors
natal ovary, and we previously observed GAGE expression in of the gonadal parenchyma contribute to the coordinated
30% of adult oocytes (Gjerstorff et al., 2006). The prolonged expression of GAGE in germ, Leydig and Sertoli cells.
expression of GAGE in germ cells, e.g. PGCs to neonatal germ In contrast to GAGE, MAGE-A1 and NY-ESO-1 were first
cells, indicates that this protein has a general function in germ detected in testis at 9 weeks and in ovary at 13 weeks.
958Cancer testis antigen expression in fetal gonads
However, in the fetal ovary, the lack of fetuses of age 8 – 13 both pluripotent germ cells and more differentiated germ
weeks prohibited us from establishing the initial induction of cells. This observation is consistent with our finding that
MAGE-A1 and NY-ESO-1 expression. The number and stain- GAGE proteins were expressed in the PGCs of indifferent
ing intensity of MAGE-A1- and NY-ESO-1-positive cells was gonads, which are known to express high levels of OCT4
significantly reduced at the end of second trimester in both (Rajpert-De Meyts et al., 2004). MAGE-A1 and NY-ESO-1
testis and ovary. The strong expression of these two proteins expression was initiated later than GAGE expression and
in the beginning of the second trimester correlates with the was clearly distinct from OCT4 expression, demonstrating
period of high proliferation of oogonia and gonocytes and the that these two CTAs are only expressed in more differen-
beginning of differentiation of these cells, and the subsequent tiated stages of fetal germ cells. Similarly, an earlier report
reduction in expression indicates that MAGE-A1 and observed that MAGE-A4 was also expressed in fetal testis
NY-ESO-1 expression is reduced as the oogonia and gonocytes by a population of cells distinct from OCT4-positive cells.
differentiate and enter cell cycle arrest. Strongly stained female MAGE-A4 was identified in a small number of cells as
germ cells were often located in clusters, indicating that high early as at weeks 7 – 9, and the number of positive cells mark-
expression of MAGE-A1, GAGE or NY-ESO-1 is a synchro- edly increased from week 17 (Aubry et al., 2001; Gaskell
Downloaded from https://academic.oup.com/humrep/article/22/4/953/698623 by guest on 06 February 2022
nized process in interconnected oogonia. et al., 2004). Thus, GAGE, MAGE-A1, NY-ESO-1 and
Analysis of third trimester and neonatal ovaries demon- MAGE-A4 seem to be sequentially expressed in fetal germ
strated a strong expression of GAGE in 95% and 11% of cells (Figure 5). In the adult testis, they have all been loca-
primary follicle oocytes, respectively. During this period, lized to the spermagonial cells, but whether they are also
NY-ESO-1 was consistently expressed in 3% of primary fol- differentially expressed in the different known stages of
licle oocytes, while MAGE-A1 was detected in not more than a spermatogonia remains to be determined.
few cells per specimen (,1%) and not at all in some speci- In conclusion, we present for the first time a detailed analysis
mens. These data are in accordance with previous results of MAGE-A1, GAGE and NY-ESO-1 expression in fetal testis,
showing that the adult ovary also contained oocytes positive and we demonstrate that CTAs per se are also expressed in fetal
for GAGE, whereas no adult oocytes were MAGE-A1- or ovary germ cells. Based on the time course of their expression,
NY-ESO-1-positive (Gjerstorff et al., 2006). Our results indi- we found indications of the function of these proteins and
cate that, at least in oocytes, MAGE-A1 is the first of these provided markers that may help differentiate germ cell
three CTAs to be turned off, while NY-ESO-1 expression per- populations.
sists through the neonatal stage and GAGE expression remains
through adulthood (Figure 5). GAGE and NY-ESO-1
expression in oocytes of primordial follicles did not seem to Acknowledgements
correlate with any morphologically distinct subtype of We would like to thank Lisbet Mortensen for her excellent technical
follicles, and the characteristics of the GAGE/NY-ESO- assistance with the immunohistochemical staining, and M.K.
1-positive oocytes remains to be elucidated. Occhipinti-Bender for editorial assistance.
Double immunohistochemical staining revealed that the
GAGE-expressing population of germ cells in the second tri-
mester gonads partially overlapped the population of OCT4- References
positive germ cells, showing that GAGE is expressed in Aubry F, Satie AP, Rioux-Leclercq N, Rajpert-De Meyts E, Spagnoli GC,
Chomez P, De Backer O, Jegou B and Samson M (2001) MAGE-A4, a
germ cell specific marker, is expressed differentially in testicular tumors.
Cancer 92,2778–2785.
Burger H and de Kretser DM (1981) The Testis, 2nd edn. Raven Press,
New York.
Gaskell TL, Esnal A, Robinson LL, Anderson RA and Saunders PT (2004)
Immunohistochemical profiling of germ cells within the human fetal testis:
identification of three subpopulations. Biol Reprod 71,2012 –2021.
Gjerstorff MF, Johansen LE, Nielsen O, Kock K and Ditzel HJ (2006)
Restriction of GAGE protein expression to subpopulations of cancer cells
is independent of genotype and may limit the use of GAGE proteins as
targets for cancer immunotherapy. Br J Cancer 94,1864– 1873.
Karl J and Capel B (1998) Sertoli cells of the mouse testis originate from the
coelomic epithelium. Dev Biol 203,323– 333.
Laduron S, Deplus R, Zhou S, Kholmanskikh O, Godelaine D, De Smet C,
Hayward SD, Fuks F, Boon T and De Plaen E (2004) MAGE-A1
interacts with adaptor SKIP and the deacetylase HDAC1 to repress
transcription. Nucl Acids Res 32,4340–4350.
Figure 5. Sequential expression of CTAs during fetal development of McGee EA (2000) The regulation of apoptosis in preantral ovarian follicles.
Biol Signals Recept 9,81–86.
human germ cells. Expression of GAGE, MAGE-A1, NY-ESO-1 and
McGee EA, Hsu SY, Kaipia A and Hsueh AJ (1998) Cell death and survival
MAGE-A4 is initiated and terminated at different time points during during ovarian follicle development. Mol Cell Endocrinol 140,15– 18.
development of fetal human germ cells. Black lines indicate CTA Monte M, Simonatto M, Peche LY, Bublik DR, Gobessi S, Pierotti MA,
expression, a reduction in the thickness of the line indicates a Rodolfo M and Schneider C (2006) MAGE-A tumor antigens target p53
reduction in CTA expression, dashed lines indicate infrequent detec- transactivation function through histone deacetylase recruitment and
tion of the antigen and white lines indicate no expression. *Data confer resistance to chemotherapeutic agents. Proc Natl Acad Sci USA
reported in Aubry et al. (2001). 103,11160 –11165.
959M.F.Gjerstorff et al.
O’Shaughnessy PJ, Baker PJ and Johnston H (2006) The foetal Leydig cell – (OCT-3/4) in normal and dysgenetic human gonads. Hum Reprod
differentiation, function and regulation. Int J Androl 29,90–95; discussion 19,1338–1344.
105– 108. Sadler TW (1985) Langman’s Medical Embryology, 5th edn. Williams &
Pauls K, Schorle H, Jeske W, Brehm R, Steger K, Wernert N, Buttner R and Wilkins, Baltimore.
Zhou H (2006) Spatial expression of germ cell markers during maturation of Satie AP, Rajpert-De Meyts E, Spagnoli GC, Henno S, Olivo L, Jacobsen GK,
human fetal male gonads: an immunohistochemical study. Hum Reprod Rioux-Leclercq N, Jegou B and Samson M (2002) The cancer–testis gene,
21,397– 404. NY-ESO-1, is expressed in normal fetal and adult testes and in spermatocytic
Rajpert-De Meyts E (2006) Developmental model for the pathogenesis of seminomas and testicular carcinoma in situ. Lab Invest 82,775–780.
testicular carcinoma in situ: genetic and environmental aspects. Hum Simpson AJ, Caballero OL, Jungbluth A, Chen YT and Old LJ (2005) Cancer/
Reprod Update 12,303–323. testis antigens, gametogenesis and cancer. Nat Rev Cancer 5,615– 625.
Rajpert-De Meyts E, Jacobsen GK, Bartkova J, Aubry F, Samson M, Bartek J Wartenberg H, Hilscher B and Hilscher W (1998) Germ cell kinetics during
and Skakkebaek NE (2003) The immunohistochemical expression pattern of early ovarian differentiation: an analysis of the oogonial cell cycle and the
Chk2, p53, p19INK4d, MAGE-A4 and other selected antigens provides subsequent changes in oocyte development during the onset of meiosis in
new evidence for the premeiotic origin of spermatocytic seminoma. the rat. Microsc Res Tech 40,377– 397.
Histopathology 42,217– 226.
Rajpert-De Meyts E, Hanstein R, Jorgensen N, Graem N, Vogt PH and Submitted on August 23, 2006; resubmitted on October 24, 2006; accepted on
Skakkebaek NE (2004) Developmental expression of POU5F1 December 1, 2006
Downloaded from https://academic.oup.com/humrep/article/22/4/953/698623 by guest on 06 February 2022
960You can also read

















































