Long non coding RNA NEAT1 promotes the regulating the microRNA 204 5p/SEMA4B axis
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
ONCOLOGY LETTERS 22: 802, 2021
Long non‑coding RNA NEAT1 promotes the
malignancy of laryngeal squamous cell carcinoma by
regulating the microRNA‑204‑5p/SEMA4B axis
LING HAN1*, CHAOPAN ZHENG1* and SHIHAI WU2
Departments of 1Otorhinolaryngology and 2Radiation Oncology, Shenzhen People's Hospital,
The Second Clinical Medical College of Jinan University, The First Affiliated Hospital of
Southern University of Science and Technology, Shenzhen, Guangdong 518020, P.R. China
Received September 6, 2020; Accepted June 11, 2021
DOI: 10.3892/ol.2021.13063
Abstract. Laryngeal squamous cell carcinoma (LSCC) is a prediction and a dual luciferase reporter assay were used to
highly invasive malignant tumor in the head and neck area. analyze the target relationship. The RT‑qPCR results showed
As an oncogene, long non‑coding RNA (lncRNA) nuclear that NEAT1 was highly expressed and miR‑204‑5p had
enriched abundant transcript 1 (NEAT1) promotes cell decreased expression in LSCC tissues and cells compared with
proliferation, migration and invasion several types of cancer. that in the normal tissue and the 16HBE‑14o cell line, respec‑
The present study aimed to reveal the effects of NEAT1 on tively. Knockdown of NEAT1 using small interfering (si) RNA
the progression of LSCC. Reverse transcription‑quantitative and overexpressed miR‑204‑5p both effectively inhibited the
PCR (RT‑qPCR) was used to detect relative mRNA expression proliferation, migration and invasion of LSCC cells. Besides,
levels of NEAT1, microRNA (miR)‑204‑5p and semaphorin further experiments revealed that miR‑204‑5p was a target
(SEMA) 4B. Kaplan‑Meier analysis was used to analyze overall of NEAT1. At the same time, silenced miR‑204‑5p reversed
survival times. RNA in‑situ hybridization (ISH) exhibited the the anti‑tumor effects of si‑NEAT1. In addition, SEMA4B
distribution of NEAT1 and miR‑204‑5p in tissues. RNA fluo‑ was targeted by miR‑204‑5p in LSCC cells and upregulated
rescence ISH was used to analyze the distribution of NEAT1 SEMA4B weakened the antitumor effects of miR‑204‑5p in
and miR‑204‑5p in the cells. Western blot analysis was used to LSCC cells. NEAT1 regulated the expression of SEMA4B
detect the expression level of target proteins. Cell viability was by targeting miR‑204‑5p in LSCC cells. Overall, NEAT1
analyzed using a MTT assay, while flow cytometry was used promoted the proliferation and invasion of LSCC cells by
to determine cell apoptosis. Wound healing and Transwell regulating the miR‑204‑5p/SEMA4B axis.
invasion assays were used to value cell migration and invasion,
respectively. RNA immunoprecipitation assay, bioinformatics Introduction
Great progress has been made in clinical surgeries, radiotherapy
and chemotherapy; however, there are still >80,000 laryn‑
Correspondence to: Professor Shihai Wu, Department of Radiation geal cancer‑related deaths in the world annually (1). Among
Oncology, Shenzhen People's Hospital, The Second Clinical Medical which, laryngeal squamous cell carcinoma (LSCC) is the
College of Jinan University, The First Affiliated Hospital of Southern most common type (2). A retrospective review, including
University of Science and Technology, 1017 Yard, Dongmen North 477 patients with laryngeal cancer between 2001 and 2014 in
Road, Luohu, Shenzhen, Guangdong 518020, P.R. China Switzerland showed that most patients with laryngeal cancer
E‑mail: shihaiwumsdoc@163.com were diagnosed at an advanced stage, and the 5‑year survival
rate was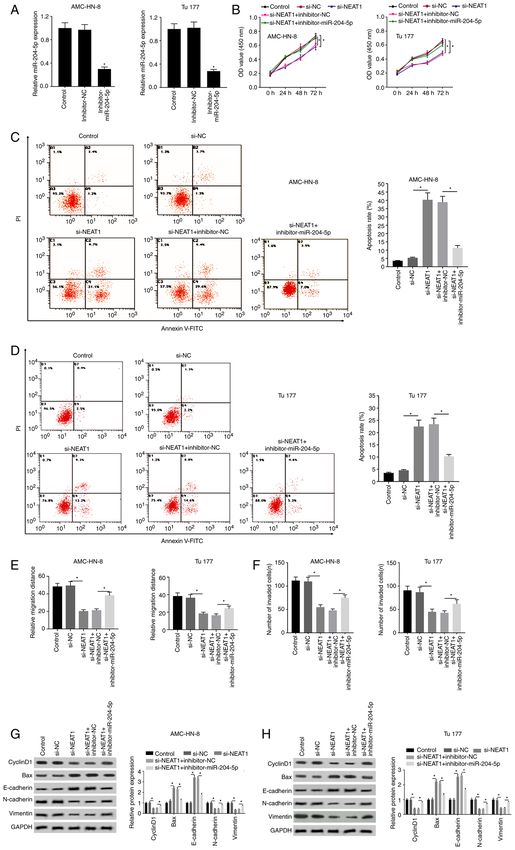
2 HAN et al: ANTITUMOR EFFECT OF NEAT1/miR-204-5p/SEMA4B AXIS IN LSCC
acted as an important oncogene in several types of cancer, Medical College of Jinan University (Shenzhen, China). The
such as colon (9), pancreatic (10) and breast cancers (11). In following inclusion criteria was used: patients with LSCC were
addition, NEAT1 was highly expressed in LSCC tissues and confirmed by pathological biopsy and they provided written
NEAT1‑knockdown using specific siRNA significantly inhib‑ informed consent. The following exclusion criteria was used:
ited the proliferation and induced apoptosis in the LSCC cells i) Patients with other malignant tumors; ii) patients with other
via the miR‑107/CDK6 pathway (12). However, to the best digestive system diseases; iii) patients with severe infection;
of our knowledge, the association between NEAT1 and cell iv) patients with mental illness or family history of mental
mobility in LSCC has never been investigated, and the related illness; v) patients with Hamilton Anxiety and Hamilton
downstream targets of NEAT1 in LSCC also requires further Depression scale score ≤5 before enrollment; vi) patients
investigation. with severe anemia; and vii) patients with abnormal liver and
Researchers have made progress in the field of oncology kidney function.
by investigating the role of miRNA in tumor occurrence,
diagnosis and treatment (13). As a novel tumor suppressor, Prognosis and follow‑up. The follow‑up period started
decreased expression of miR‑204‑5p was detected in the tissues with the discharge date and the average length of follow‑up
and cell lines of several cancer types, including hepatocellular was 45 months. The longest follow‑up lasted for 72 months.
cancer (14) and head and neck squamous cell carcinoma (15). The follow‑up was short if the patient died. Follow‑up was
miR‑204‑5p was associated with cell proliferation, clonoge‑ conducted via a telephone interview or door to door review.
nicity and aggressiveness in tumors and it also functioned as The survival time was recorded in the unit of month.
a prospective therapeutic target for clinical intervention. For
example, Gao et al (16) also reported that miR‑204‑5p inhib‑ Cell culture. LSCC cell lines AMC‑HN‑3, HN‑10 and Tu177
ited the invasion and metastasis of LSCC by suppressing the and the human bronchial epithelioid cells 16HBE‑14o were
expression level of forkhead box C1. In addition, miR‑204‑5p purchased from ScienCell Research Laboratories, Inc. and
was found to be the downstream target of lncRNAs in LSCC, were maintained in Dulbecco's modified Eagle's medium
such as lncRNA MIR100HG (17). However, the association (DMEM) containing 10% fetal bovine serum (Invitrogen;
between miR‑204‑5p and NEAT1 has never been investi‑ Thermo Fisher Scientific, Inc.) in a humidified incubator
gatedin LSCC, to the best of our knowledge. at 37˚C with 5% CO2. The AMC‑HN‑3 and Tu177 cells were
Semaphorins (SEMAs) were first identified as axon used for most of the experiments, due to their representative
guidance molecules during the process of neuronal devel‑ expression.
opment (18). Subsequent research revealed that SEMAs
regulate cell migration, angiogenesis and the adaptive Reverse transcription‑quantitative PCR (RT‑qPCR). Total
immune response (19). SEMA4B is a class 4 SEMA, one of RNA from the aforementioned cells and tissues was extracted
the seven classes of the SEMA family of proteins. In addi‑ using TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
tion, SEMA4B was reported to be related with the growth and Inc.) according to the manufacturer's instructions. cDNA was
metastasis of non‑small cell lung cancer (20,21). However, synthesized using the One‑step PrimeScript TM RT Reagent
in‑depth study on the regulatory mechanism of SEMA4B kit according to the manufacturer's instructions (Takara
in cancer has not yet been conducted. The present study Biotechnology Co., Ltd.). Following standard operation, the
explored the association among SEMA4B, lncRNA NEAT1 RT‑qPCR mixture system containing the cDNA templates,
and miR‑204‑5p in LSCC. primers and SYBR Green qPCR Master mix (Takara
The aim of the present study was to investigate the effects Biotechnology Co., Ltd.) were subjected to RT‑qPCR using
of the NEAT1/miR‑204‑5p/SEMA4B axis in LSCC cell an ABI Prism 7500 Sequence Detection system (Applied
progression. It was revealed, for the first time, that SEMA4B Biosystems; Thermo Fisher Scientific, Inc.). The following
was a target of miR‑204‑5p in LSCC cells and upregulated thermocycling conditions were used: Initial denaturation
SEMA4B weakened the antitumor effects of miR‑204‑5p at 97˚C for 5 min, denaturation at 70˚C for 35 sec, annealing
mimic in LSCC. In addition, miR‑204‑5p mediated the regula‑ at 64˚C for 40 sec, extension at 72˚C for 2 min, for 35 cycles and
tion of NEAT1 on the expression SEMA4B protein. In general, final extension at 72˚C for 5 min. β‑actin and U6 small nuclear
the present study investigated the effects of NEAT1 on LSCC RNA (snRNA) were used as the internal controls for mRNA
and provided novel targets for gene targeting therapy in LSCC. or miRNA, respectively. Relative gene expression was quanti‑
fied using the 2‑ΔΔCq method (22). Related primers (Vazyme
Materials and methods Biotech Co., Ltd.) are listed as following: NEAT1 forward,
5'‑TTGGGACAGTFFACGTGTGG‑3' and reverse, 5'‑TCA
Tissue collection. In total, 20 pairs of LSCC tissues and AGTCCAGCAGAGCA‑3', miR‑204‑5p forward, 5'‑CGAAGT
adjacent normal tissues (5 cm away from the cancer tissue) TCCCTTTGTCATCCT‑3' and reverse 5'‑GTGCAGGGTCCG
were obtained from 20 patients (median age 68 years; range, AGGTATTC‑3'; U6 forward, 5'‑GCTTCGGCAGCACATATA
40‑80 years old) with LSCC from resection surgery. The CT‑3' and reverse, 5'‑GTGCAGGGTCCGAGGTATTC‑3' and
patients were admitted to The Second Clinical Medical β‑actin forward, 5'‑CTCCATCCTGGCCTCGT‑3' and reverse
College of Jinan University (Guangdong, China) between 5'‑GCTGTCACCTTCACCGTTCC‑3'.
December 2017 and December 2019. The study was conducted
in accordance with the Declaration of Helsinki. All patients Kaplan‑Meier analysis. The related information on
signed written informed consent, and the present study was NEAT1, miR‑204‑5p and SEMA4B were firstly down‑
approved by The Ethics Committee of The Second Clinical loaded from The Cancer Genome Atlas database
ONCOLOGY LETTERS 22: 802, 2021 3 (https://www.cancer.gov/about‑nci/organization/ccg/research/ overnight, in the dark. Then, the slides were incubated with structural‑genomics/tcga/using‑tcga/types). All LSCC cases biotin‑conjugated anti‑digoxin antibody (cat. no. ab30512; were grouped into the low expression group and the high 1:500; Abcam) at 37˚C for 3 h and StreptAvidin Biotin expression group. Subsequently, Kaplan‑Meier survival plots Complex‑FITC in turn. After the nucleus was stained with were generated using SPSS v13.0 software (SPSS, Inc.). The DAPI at room temperature for 10 min, cell images were hazard ratio (HR) with 95% confidence interval (CI) and captured using confocal microscopy (Leica TCS SP8; Leica log‑rank P‑value were then calculated automatically. The HR Microsystems, Inc.) at x630 magnification. values with 95% CI of the three genes are listed as follows: NEAT1: HR, 1.73; CI, 81.80‑88.90; miR‑204‑5p: HR, 0.725; CI, Western blot analysis. Western blot analysis was conducted 53.50‑91.50; SEMA4B: HR, 1.64; CI: 67.30‑87.50. Log‑rank following standard procedures. The total protein from the P
4 HAN et al: ANTITUMOR EFFECT OF NEAT1/miR-204-5p/SEMA4B AXIS IN LSCC
was performed at room temperature and the transfected cells vector. AMC‑HN‑3 and Tu177 cells were cultured in 96‑well
were kept at 37˚C for 24 h before further experimentation. plates (1x104 cells/well) and were co‑transfected with 400 ng
of either 3'‑UTR‑WT or 3'‑UTR‑MUT (Promega Corporation)
MTT assay. Cell viability was assessed using an MTT and 50 nmol/l miR‑204‑5p mimic using Lipofectamine® 3000
assay. Cells were seeded into 96‑well plates in triplicates according to the manufacturer's protocols. Luciferase activity
(8x103 cells/well). Then, 10 µl of MTT reagent (5 mg/ml; Sangon assays were conducted using the Dual‑Luciferase Reporter
Biotech Co., Ltd.) was added into each well and was incubated Assay system (Promega Corporation) 24 h later. The results
for 1.5 h. Then, the formazan in the well was dissolved with were normalized to luciferase activity (firefly luciferase/Renilla
dimethyl sulfoxide and the OD value was measured with a luciferase).
spectrophotometer (Bio‑Rad Laboratories) at 540 nm at 0,
24, 48, 72 h, respectively. Each experiment was performed in RNA immunoprecipitation (RIP) assay. The Magna RIP
triplicate. RNA‑Binding Protein Immunoprecipitation kit (cat. no. 17‑700;
Sigma‑Aldrich; Merck KGaA) was used for the RIP assay
Flow cytometry. Apoptosis was examined using BD according to the instructions. Briefly, AMC‑HN‑3 and Tu177
FACSCalibur (BD Biosciences). In short, AMC‑HN‑3 and cells were incubated with Argonaute2 (anti‑Ago2; Abcam) or
Tu177 cells were labeled with annexin V‑FITC and prop‑ a negative IgG (anti‑IgG; Abcam) at 4˚C for 2 h. Then, the
idium iodide (PI), and then were examined with an apoptosis Ago2 antibody was recovered with the protein A/G beads
detecting kit (cat. no. V13242; Invitrogen; Thermo Fisher (cat. no. LSKMAGAG02; Sigma‑Aldrich; Merck KGaA).
Scientific, Inc.). Samples were examined by flow cytometry The enrichment of NEAT1, miR‑204‑5p and SEMA4B was
and apoptosis rates were then analyzed by CellQuest software assessed using RT‑qPCR.
v6.0 (BD Biosciences) following the manufacturer's protocols.
Statistical analysis. All the experiments in the study were
Wound healing assay. The Tu177 and AMC‑HN‑3 cells were conducted in triplicate. SPSS 13.0 software (SPSS Inc.) and
seeded in the 6‑well cell culture plate (1x106/well) and were GraphPad Prism 7 (GraphPad Software, Inc.) were used
cultured overnight in DMEM containing 10% FBS (Invitrogen; to examine statistical analysis. Data were presented as the
Thermo Fisher Scientific, Inc.). After the cells reached 100% mean ± SD. The overall survival rate was analyzed through
confluence, a wound was made using a sterile pipette tip. The Kaplan‑Meier analysis. The Kaplan‑Meier analysis was
cells were then washed gently with sterile PBS, 3 times, to assessed using a log‑rank test. Spearman correlation analysis
remove non‑adherent cells. Then, the medium in the culture between NEAT1 or SEMA4B expression and miR‑204‑5p
plate was replaced with fresh serum‑free medium. The width expression was also conducted. The difference between two
of the wound was measured 24 h later, using an inverted groups was analyzed using an unpaired two‑tailed Student's
phase microscope with DP v2 controller software (Olympus t‑test. One‑way ANOVA followed by Dunnett's post hoc
Corporation). test was used to analyze differences among more than two
groups. The small sample sizes in Tables I‑III were compared
Transwell invasion assay. Matrigel was melted overnight using Fisher's exact test. P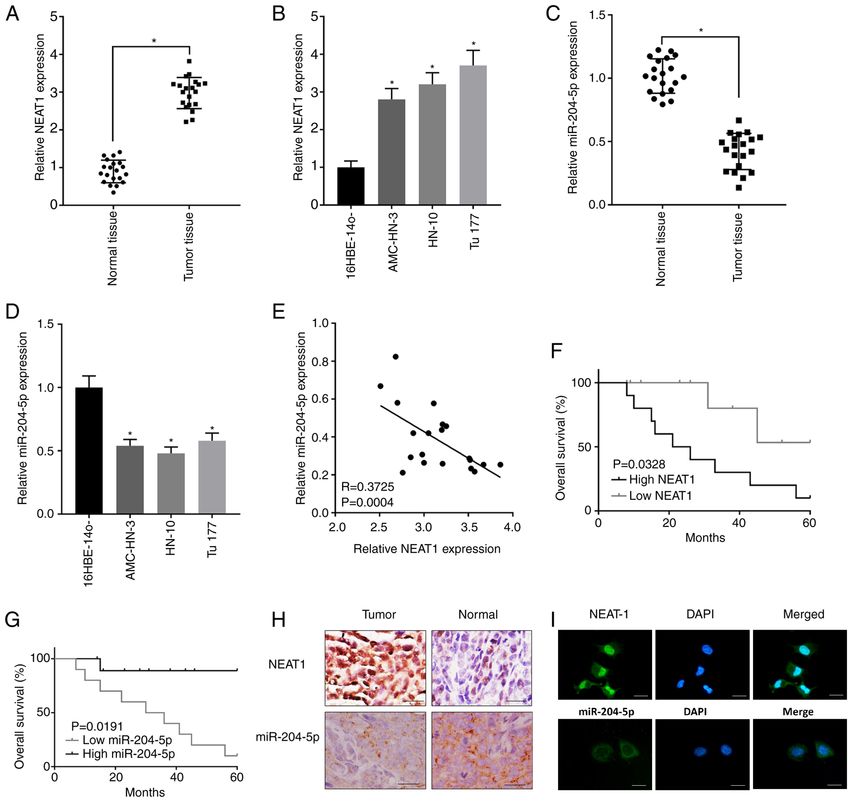
ONCOLOGY LETTERS 22: 802, 2021 5
Table I. Association between NEAT1 expression and the clinicopathological features of patients with laryngeal squamous cell
carcinoma (n=20).
NEAT1 expression
‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑
Characteristics Number Low (n=10) High (n=10) P‑value
Age, years 0.657
≤60 6 2 4
>60 14 8 6
Sex 0.489
Female 8 3 5
Male 12 7 5
Lymph node metastasis 0.002a
No 9 8 1
Yes 11 2 9
Clinical stage 0.034a
I‑II 9 7 2
III‑IV 11 3 8
P60 14 8 6
Sex 0.568
Female 8 5 3
Male 12 5 7
Lymph node metastasis 0.043a
No 9 2 7
Yes 11 8 3
Clinical stage 0.003a
I‑II 9 1 8
III‑IV 11 9 2
a
P
6 HAN et al: ANTITUMOR EFFECT OF NEAT1/miR-204-5p/SEMA4B AXIS IN LSCC
Table III. Association between SEMA4B expression and the clinicopathological features of patients with laryngeal squamous
cell carcinoma (n=20).
SEMA4B expression
‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑
Characteristics Number Low (n=10) High (n=10) P‑value
Age, years 0.452
≤60 6 4 2
>60 14 6 8
Sex 0.568
Female 8 3 5
Male 12 7 5
Lymph node metastasis 0.042a
No 9 7 2
Yes 11 3 8
Clinical stage 0.028a
I‑II 9 7 2
III‑IV 11 3 8
P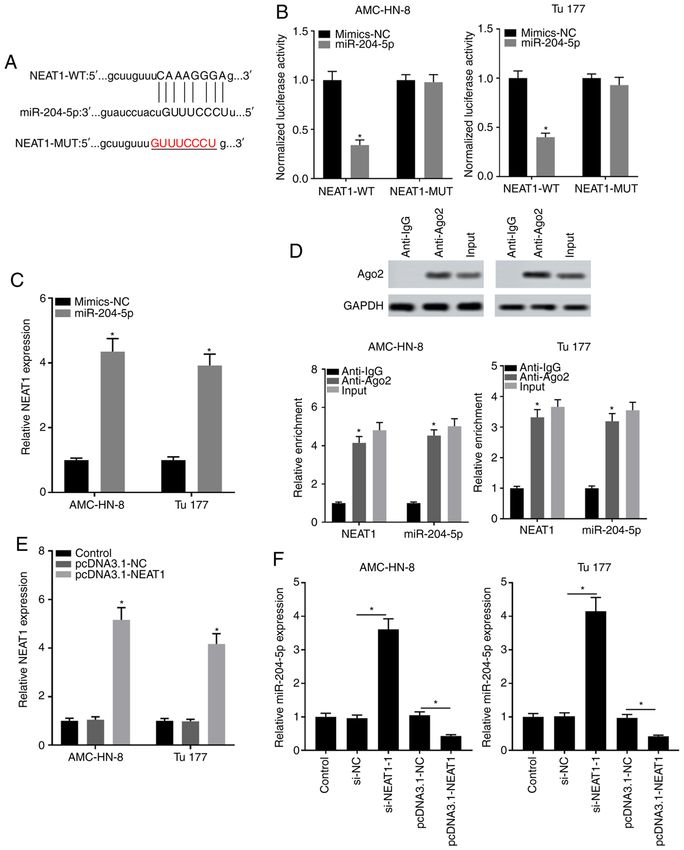
ONCOLOGY LETTERS 22: 802, 2021 7 Figure 1. NEAT1 is highly expressed and miR‑204‑5p is lowly expressed in LSCC tissues and cell lines. (A) Relative expression levels of NEAT1 in LSCC and normal tissues were detected using RT‑qPCR. (B) Relative expression levels of NEAT1 in LCSS cell lines (AMC‑HN‑3, HN‑10 and Tu177) and human bronchial epithelioid cells (16HBE‑14o) were detected usingRT‑qPCR. (C) Relative expression levels of miR‑204‑5p in LSCC and normal tissues were detected using RT‑qPCR. (D) Relative expression levels of miR‑204‑5p in LCSS cell lines (AMC‑HN‑3, HN‑10 and Tu177) and human bronchial epithelioid cells (16HBE‑14o) were detected using RT‑qPCR. (E) Correlation analysis between NEAT1 and miR‑204‑5p expression in LSCC tissues. Association between overall survival rate and (F) NEAT1 and (G) miR‑204‑5p expression level in LSCC tissues. (H) Representative images of in situ hybridization showing NEAT1 and miR‑204‑5p expression within LSCC and normal tissues. Scale bar, 50 µm. (I) Representative fluorescence in situ hybridization images indicating subcellular location of NEAT1 and miR‑204‑5p in AMC‑HN‑3 cells (green). Nuclei were stained by DAPI (blue). *P
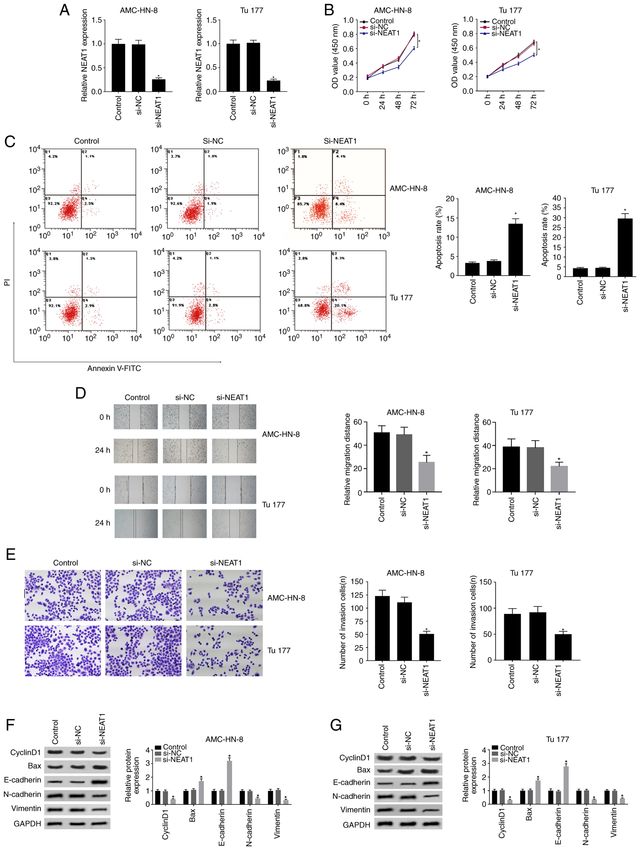
8 HAN et al: ANTITUMOR EFFECT OF NEAT1/miR-204-5p/SEMA4B AXIS IN LSCC Figure 2. Knockdown of NEAT1 inhibits cell proliferation and mobility, and promotes apoptosis. AMC‑HN‑3 and Tu177 cells were used for these experiments. (A) Silencing effect of si‑NEAT1 was firstly verified in AMC‑HN‑3 and Tu177 cells through reverse transcription‑quantitative PCR. (B) Effect of si‑NEAT1 on cell proliferation was evaluated through MTT assay. (C) Effect of si‑NEAT1 on apoptosis was assessed through flow cytometry. (D) Effect of si‑NEAT1 on cell migration was analyzed using wound healing assay. Magnification, x100. (E) Transwell invasion assay was used to evaluate cell invasion. Western blotting results of the protein expression levels of cyclinD1, N‑cadherin, vimentin, E‑cadherin and Bax in (F) AMC‑HN‑3 and (G) Tu177 cells. *P
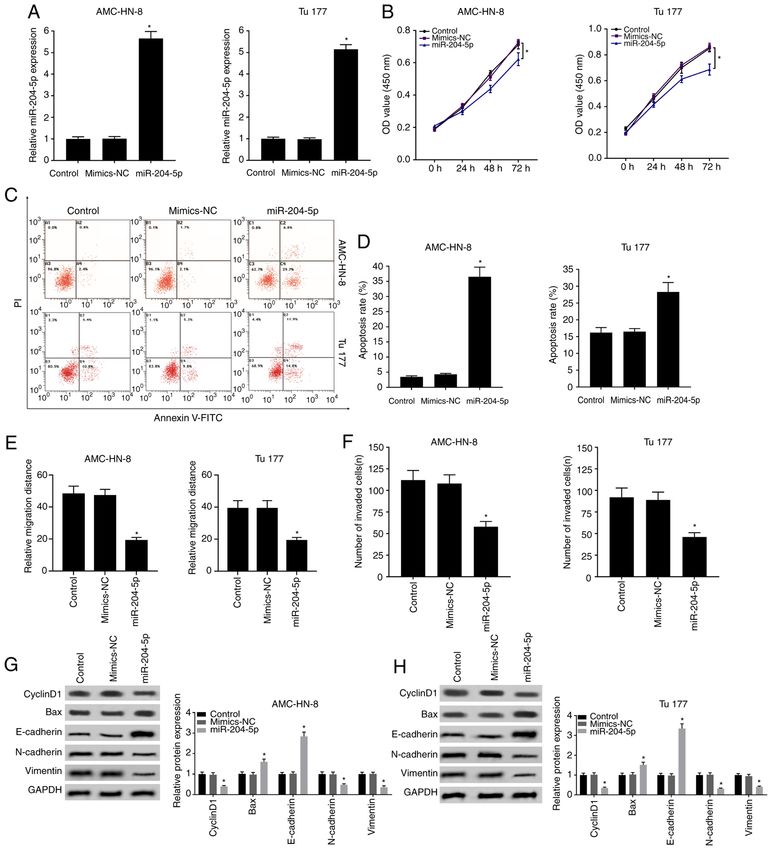
ONCOLOGY LETTERS 22: 802, 2021 9 Figure 3. Overexpression of miR‑204‑5p inhibits cell mobility and proliferation, and promotes apoptosis. (A) Overexpression of miR‑204‑5p was confirmed using reverse transcription‑quantitative PCR. (B) Effect of overexpressed miR‑204‑5p on cell proliferation was evaluated using a MTT assay. (C and D) Effect of overexpressed miR‑204‑5p on apoptosis was assessed using flow cytometry. Statistical results for (E) cell migration and (F) cell invasion. Western blotting results for the protein expression levels of cyclinD1, N‑cadherin, vimentin, E‑cadherin and Bax in (G) AMC‑HN‑3 and (H) Tu177 cells. *P
10 HAN et al: ANTITUMOR EFFECT OF NEAT1/miR-204-5p/SEMA4B AXIS IN LSCC Figure 4. miR‑204‑5p acts as a target of NEAT1. (A) Target complementary sequence between miR‑204‑5p and NEAT1 was analyzed using bioinformatics analysis. (B) Targeting association between NEAT1 and miR‑204‑5p was verified using dual luciferase reporter assay. (C) Effect of overexpressed miR‑204‑5p on NEAT1 expression was evaluated using RT‑qPCR. *P
ONCOLOGY LETTERS 22: 802, 2021 11 Figure 5. Inhibitory effects of si‑NEAT1 on the progression of laryngeal squamous cell carcinoma are reversed by knockdown of miR‑204‑5p. (A) Transfection efficiency of inhibitor‑miR‑204‑5p was evaluated using reverse transcription‑quantitative PCR. (B) Cell proliferation was measured through MTT assay. Effect of miR‑204‑5p and NEAT1 on apoptosis was assessed through flow cytometry in (C) AMC‑HN‑3 and (D) Tu177 cells. (E) Statistical results of the effect of miR‑204‑5p and NEAT1 on cell migration. (F) Statistical results of the effect of miR‑204‑5p and NEAT1 on cell invasion. Effect of miR‑204‑5p and NEAT1 on the protein expression levels of cyclinD1, N‑cadherin, vimentin, E‑cadherin and Bax in (G) AMC‑HN‑3 and (H) Tu177 cells was analyzed through western blotting. *P
12 HAN et al: ANTITUMOR EFFECT OF NEAT1/miR-204-5p/SEMA4B AXIS IN LSCC Figure 6. SEMA4B is targeted by miR‑204‑5p in LSCC. (A) Target complementary sequence between miR‑204‑5p and SEMA4B was analyzed using bioin‑ formatics analysis. (B) Targeting association between SEMA4B and miR‑204‑5p was verified using dual luciferase reporter assay. (C) Effect of overexpressed miR‑204‑5p on SEMA4B expression was analyzed using RT‑qPCR. *P
ONCOLOGY LETTERS 22: 802, 2021 13 Figure 7. SEMA4B overexpression weakens the antitumor effect of miR‑204‑5p mimic in laryngeal squamous cell carcinoma. (A) Transfection efficiency of pcDNA3.1‑SEMA4B was confirmed using reverse transcription‑quantitative PCR. *P
14 HAN et al: ANTITUMOR EFFECT OF NEAT1/miR-204-5p/SEMA4B AXIS IN LSCC Figure 8. NEAT1 regulates SEMA4B by targeting miR‑204‑5p. (A) Effect of inhibitor miR‑204‑5p and si‑NEAT1 on SEMA4B expression was evaluated using RT‑qPCR. (B) Effect of inhibitor miR‑204‑5p and si‑NEAT1 on SEMA4B protein expression was measured using western blotting. (C) Effect of miR‑204‑5p mimic and pcDNA3.1‑NEAT1 on SEMA4B expression was evaluated using RT‑qPCR. (D) Effect of miR‑204‑5p mimic and pcDNA3.1‑NEAT1 on SEMA4B protein expression was analyzed through western blotting. (E) Correlation analysis between SEMA4B and NEAT1 expression. *P
ONCOLOGY LETTERS 22: 802, 2021 15
NEAT1 is the core component of paraspeckles and was constraints. Thus, corresponding in vivo experiments should
associated with the nucleo‑cytoplasmic transport of mRNA be performed in future experiments, and further therapeutic
by paraspeckles (29,30). It has been shown that some mature targets with clinical application value should be gradually
miRNAs are enriched in the nucleus, thus nuclear NEAT1 acts explored.
as a competing endogenous RNA to elevate the expression
levels of targeting miRNAs through dis‑inhibition, in this way Acknowledgements
promoting tumor progression (31,32). However, the targeting
miRNAs of NEAT1 in LSCC have not been widely studied. Not applicable.
The present study revealed that miR‑204‑5p expression was
low in LSCC tissues and cell lines. Additionally, miR‑204‑5p Funding
overexpression using a miR‑204‑5p mimic inhibited cell
mobility and proliferation, and promoted apoptosis. In addi‑ The present study was funded by the Natural Science
tion, miR‑204‑5p was found to be a target of NEAT1 and the Foundation of China (grant no. N31801160) and Shenzhen
antitumor effects of si‑NEAT1 on LSCC were reversed by Natural Science Foundation (grant nos. JCYJ2019072922262
silencing miR‑204‑5p. Similarly, Wang et al (33) reported 9201 and JCYJ20190807150811212).
that lncRNA OIP5‑AS1 served as a competing endogenous
RNA of miR‑204‑5p in LSCC cells, and the restoration of Availability of data and materials
miR‑204‑5p counteracted the OIP5‑AS1‑mediated oncogenic
effects. Additionally, Jiang et al (34) previously reported that The datasets used and/or analyzed during the current study are
lncRNA NEAT1 enhanced docetaxel resistance in prostate available from the corresponding author on reasonable request.
cancer by regulating ACSL4 via sponging miR‑34a‑5p and
miR‑204‑5p. Thus, the targeting association between NEAT1 Authors' contributions
and miR‑204‑5p may also be applicable in LSCC cells, and
NEAT1 and miR‑204‑5p may be jointly involved in the LH conducted the experiments and arranged the figures and
progression of LSCC. the manuscript. CZ assisted with sample collection, checked
The corresponding downstream target genes of miR‑204‑5p the information of the patient, assisted with some of the
were further explored in the present study. Tang et al (35) experiments, analyzed the data and revised the manuscript
reported that miR‑204‑5p regulated cell proliferation, inva‑ critically. SW initiated and supervised the project, analyzed
sion and apoptosis by targeting IL‑11 in esophageal squamous the data, formed the conclusion and edited the manuscript.
cell carcinoma. PIK3CB, a major regulator of the PI3K/Akt LH and SW confirm the authenticity of the data. All authors
signaling pathway, is also a direct target of miR‑204‑5p, have read and approved the final manuscript.
and miR‑204‑5p regulates the growth, metastasis and
immune microenvironment remodeling in breast cancer by Ethics approval and consent to participate
targeting PIK3CB (36). In the present study, bioinformatics
analysis revealed a target complementary sequence between All patients signed written informed consent forms, and the
miR‑204‑5p and SEMA4B. SEMA4B is a class IV semaphorin present study was approved by The Ethics Committee of
involved in the regulation of cell motility (37). Results of The Second Clinical Medical College of Jinan University
multiple experiments in the present study, including dual lucif‑ (Shenzhen, China).
erase reporter assay, RIP assay and RT‑qPCR, further verified
the targeting association between miR‑204‑5p and SEMA4B. Patient consent for publication
Furthermore, Li et al (38) revealed that SEMA4B could be
used as a biomarker for potential clinical evaluation of gastric Not applicable.
cancer. SEMA4B also acts as a target of miR‑34a and as a
potential therapeutic target of colorectal cancer (39). However, Competing interests
SEMA4B has mainly been studied in lung cancer in previous
studies (40‑42) and has not been evaluated in LSCC. The The authors declare that they have no competing interests.
current study revealed that SEMA4B overexpression weak‑
ened the antitumor effects of miR‑204‑5p mimic in LSCC. References
Additionally, SEMA4B expression was positively correlated
with NEAT1 expression and negatively correlated with 1. Fusconi M, Campo F, Gallo A, Zambetti G, Martellucci S,
Seccia A and de Vincentiis M: Laryngeal cancer, HPV DNA
miR‑204‑5p expression. Overall, the regulation of NEAT1 on vs E6/E7 mRNA test: A systematic review. J Voice 31: 248.
SEMA4B may be mediated by miR‑204‑5P. e241‑248.e245, 2017.
In conclusion, the current results revealed that NEAT1 2. Steuer CE, El‑Deiry M, Parks JR, Higgins KA and Saba NF: An
update on larynx cancer. CA Cancer J Clin 67: 31‑50, 2017.
contributed to cell proliferation and mobility, and suppressed 3. Anschuetz L, Shelan M, Dematte M, Schubert AD, Giger R and
apoptosis by regulating the miR‑204‑5p/SEMA4B axis in Elicin O: Long‑term functional outcome after laryngeal cancer
LSCC. The present study presented new potential biomarkers treatment. Radiat Oncol 14: 101, 2019.
4. Fang YW and Fullwood MJ: Roles, functions, and mechanisms
for targeted therapy of LSCC. However, other downstream of long non‑coding RNAs in cancer. Genomics Proteomics
targets of NEAT1 and miR‑204‑5p should be further explored Bioinformatics 14: 42‑54, 2016.
in future studies. There are also some limitations in the 5. Gupta SC and Tripathi YN: Potential of long non‑coding RNAs
in cancer patients: From biomarkers to therapeutic targets. Int
present study, such as a lack of in vivo experiments due to time J Cancer 140: 1955‑1967, 2017.16 HAN et al: ANTITUMOR EFFECT OF NEAT1/miR-204-5p/SEMA4B AXIS IN LSCC
6. Hung T and Chang HY: Long noncoding RNA in genome regula‑ 26. Bhan A, Soleimani M and Mandal SS: Long noncoding RNA and
tion: Prospects and mechanisms. RNA Biol 7: 582‑585, 2010. cancer: A new paradigm. Cancer Res 77: 3965‑3981, 2017.
7. Fang XY, Pan HF, Leng RX and Ye DQ: Long noncoding RNAs: 27. Yang G, Lu X and Yuan L: LncRNA: A link between RNA and
Novel insights into gastric cancer. Cancer Lett 356: 357‑366, 2015. cancer. Biochim Biophys Acta 1839: 1097‑1109, 2014.
8. Zhang Y and Tang L: The application of lncRNAs in cancer 28. Wang W, Ge L, Xu XJ, Yang T, Yuan Y, Ma XL and Zhang XH:
treatment and diagnosis. Recent Pat Anticancer Drug Discov 13: LncRNA NEAT1 promotes endometrial cancer cell prolifera‑
292‑301, 2018. tion, migration and invasion by regulating the miR‑144‑3p/EZH2
9. Zhuang ST, Cai YJ, Liu HP, Qin Y and Wen JF: LncRNA axis. Radiol Oncol 53: 434‑442, 2019.
NEAT1/miR‑185‑5p/IGF2 axis regulates the invasion and migra‑ 29. Yamazaki T and Hirose T: The building process of the functional
tion of colon cancer. Mol Genet Genomic Med 8: e1125, 2020. paraspeckle with long non‑coding RNAs. Front Biosci (Elite
10. Feng Y, Gao L, Cui G and Cao Y: LncRNA NEAT1 facilitates Ed) 7: 1‑41, 2015.
pancreatic cancer growth and metastasis through stabilizing 30. Kawaguchi T, Tanigawa A, Naganuma T, Ohkawa Y, Souquere S,
ELF3 mRNA. Am J Cancer Res 10: 237‑248, 2020. Pierron G and Hirose T: SWI/SNF chromatin‑remodeling
11. Knutsen E, Lellahi SM, Aure MR, Nord S, Fismen S, Larsen KB, complexes function in noncoding RNA‑dependent assembly of
Gabriel MT, Hedberg A, Bjørklund SS; Oslo Breast Cancer nuclear bodies. Proc Natl Acad Sci USA 112: 4304‑4309, 2015.
Research Consortium (OSBREAC), et al: The expression of the 31. Sun C, Li S, Zhang F, Xi Y, Wang L, Bi Y and Li D: Long
long NEAT1_2 isoform is associated with human epidermal growth non‑coding RNA NEAT1 promotes non‑small cell lung cancer
factor receptor 2‑positive breast cancers. Sci Rep 10: 1277, 2020. progression through regulation of miR‑377‑3p‑E2F3 pathway.
12. Wang P, Wu T, Zhou H, Jin Q, He G, Yu H, Xuan L, Oncotarget 7: 51784‑51814, 2016.
Wang X, Tian L, Sun Y, et al: Long noncoding RNA NEAT1 32. Liu H, Li A, Sun Z, Zhang J and Xu H: Long non‑coding RNA
promotes laryngeal squamous cell cancer through regulating NEAT1 promotes colorectal cancer progression by regulating
miR‑107/CDK6 pathway. J Exp Clin Cancer Res 35: 22, 2016. miR‑205‑5p/VEGFA axis. Human Cell 33: 386‑396, 2020.
13. Pichler M and Calin GA: MicroRNAs in cancer: From develop‑ 33. Wang H, Qian J, Xia X and Ye B: Long non‑coding RNA
mental genes in worms to their clinical application in patients. OIP5‑AS1 serves as an oncogene in laryngeal squamous cell
Br J Cancer 113: 569‑573, 2015. carcinoma by regulating miR‑204‑5p/ZEB1 axis. Naunyn
14. Chu Y, Jiang M, Du F, Chen D, Ye T, Xu B, Li X, Wang W, Schmiedebergs Arch Pharmacol 393: 2177‑2184, 2020.
Qiu Z, Liu H, et al: miR‑204‑5p suppresses hepatocellular cancer 34. Jiang X, Guo S, Zhang Y, Zhao Y, Li X, Jia Y, Xu Y and Ma B:
proliferation by regulating homeoprotein SIX1 expression. FEBS LncRNA NEAT1 promotes docetaxel resistance in prostate
Open Bio 8: 189‑200, 2018. cancer by regulating ACSL4 via sponging miR‑34a‑5p and
15. Zhuang Z, Yu P, Xie N, Wu Y, Liu H, Zhang M, Tao Y, Wang W, miR‑204‑5p. Cell Signal 65: 109422, 2020.
Yin H, Zou B, et al: MicroRNA‑204‑5p is a tumor suppressor 35. Tang J, Li Z, Zhu Q, Wen W, Wang J, Xu J, Wu W, Zhu Y, Xu H
and potential therapeutic target in head and neck squamous cell and Chen L: miR‑204‑5p regulates cell proliferation, invasion,
carcinoma. Theranostics 10: 1433‑1453, 2020. and apoptosis by targeting IL‑11 in esophageal squamous cell
16. Gao W, Wu Y, He X, Zhang C, Zhu M, Chen B, Liu Q, Qu X, carcinoma. J Cell Physiol 235: 3043‑3055, 2020.
Li W, Wen S and Wang B: MicroRNA‑204‑5p inhibits inva‑ 36. Hong BS, Ryu HS, Kim N, Kim J, Lee E, Moon H, Kim KH,
sion and metastasis of laryngeal squamous cell carcinoma by Jin MS, Kwon NH, Kim S, et al: Tumor suppressor miRNA‑204‑5p
suppressing forkhead box C1. J Cancer 8: 2356‑2368, 2017. regulates growth, metastasis, and immune microenvironment
17. Huang Y, Zhang C and Zhou Y: LncRNA MIR100HG promotes remodeling in breast cancer. Cancer Res 79: 1520‑1534, 2019.
cancer cell proliferation, migration and invasion in laryngeal 37. Nagai H, Sugito N, Matsubara H, Tatematsu Y, Hida T, Sekido Y,
squamous cell carcinoma through the downregulation of Nagino M, Nimura Y, Takahashi T and Osada H: CLCP1 inter‑
miR‑204‑5p. Onco Targets Ther 12: 2967‑2973, 2019. acts with semaphorin 4B and regulates motility of lung cancer
18. Kolodkin AL, Matthes DJ and Goodman CS: The semaphorin cells. Oncogene 26: 4025‑4031, 2007.
genes encode a family of transmembrane and secreted growth 38. Li F, Yoshizawa JM, Kim KM, Kanjanapangka J, Grogan TR,
cone guidance molecules. Cell 75: 1389‑1399, 1993. Wang X, Elashoff DE, Ishikawa S, Chia D, Liao W, et al:
19. Ahmed A and Eickholt BJ: Intracellular kinases in semaphorin Discovery and validation of salivary extracellular RNA
signaling. Adv Exp Med Biol 600: 24‑37, 2007. biomarkers for noninvasive detection of gastric cancer. Clin
20. Jian H, Zhao Y, Liu B and Lu S: SEMA4B inhibits growth of Chem 64: 1513‑1521, 2018.
non‑small cell lung cancer in vitro and in vivo. Cell Signal 27: 39. Wang T, Xu H, Liu X, Chen S, Zhou Y and Zhang X: Identification
1208‑1213, 2015. of key genes in colorectal cancer regulated by miR‑34a. Med Sci
21. Jian H, Zhao Y, Liu B and Lu S: SEMA4b inhibits MMP9 to Monit 23: 5735‑5743, 2017.
prevent metastasis of non‑small cell lung cancer. Tumour Biol 35: 40. Hong J, Yi Z, Bin L and Shun L: SEMA4b inhibits MMP9 to
11051‑11056, 2014. prevent metastasis of non‑small cell lung cancer. Tumour Biol 35:
22. Livak KJ and Schmittgen TD: Analysis of relative gene expres‑ 11051‑11056, 2014.
sion data using real‑time quantitative PCR and the 2(‑Delta Delta 41. Jian H, Liu B and Zhang J: Hypoxia and hypoxia‑inducible factor
C(T)) method. Methods 25: 402‑408, 2001. 1 repress SEMA4B expression to promote non‑small cell lung
23. Chen Z, Zhang Z, Zhao D, Feng W, Meng F, Han S, Lin B and cancer invasion. Tumour Biol 35: 4949‑4955, 2014.
Shi X: Long noncoding RNA (lncRNA) FOXD2‑AS1 promotes 42. Nagai H, Sugito N, Matsubara H, Tatematsu Y, Hida T, Sekido Y,
cell proliferation and metastasis in hepatocellular carcinoma by Nagino M, Nimura Y, Takahashi T and Osada H: CLCP1 inter‑
regulating MiR‑185/AKT axis. Med Sci Monitor 25: 9618‑9629, acts with semaphorin 4B and regulates motility of lung cancer
2019. cells. Oncogene 26: 4025‑4031, 2007.
24. Chaffer CL and Weinberg RA: A perspective on cancer cell
metastasis. Science 331: 1559‑1564, 2011. This work is licensed under a Creative Commons
25. Wang B, Lv K, Chen W, Zhao J, Luo J, Wu J, Li Z, Qin H, Attribution-NonCommercial-NoDerivatives 4.0
Wong TS, Yang W, et al: miR‑375 and miR‑205 regulate the International (CC BY-NC-ND 4.0) License.
invasion and migration of laryngeal squamous cell carcinoma
synergistically via AKT‑Mediated EMT. Biomed Res Int 2016:
9652789, 2016.You can also read

















































