Leonurine Regulates Hippocampal Nerve Regeneration in Rats with Chronic and Unpredictable Mild Stress by Activating SHH/GLI Signaling Pathway and ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Hindawi Neural Plasticity Volume 2023, Article ID 1455634, 12 pages https://doi.org/10.1155/2023/1455634 Research Article Leonurine Regulates Hippocampal Nerve Regeneration in Rats with Chronic and Unpredictable Mild Stress by Activating SHH/GLI Signaling Pathway and Restoring Gut Microbiota and Microbial Metabolic Homeostasis Pan Meng,1,2 Xi Zhang,1,3 Dandan Li,1 Hui Yang ,2,4 Xiaoyuan Lin,2,4 Hongqing Zhao,1,2 Ping Li,1 Yuhong Wang,1,2 Xiaoye Wang ,3 and Jinwen Ge 5 1 Hunan University of Chinese Medicine, Changsha, Hunan, China 2 Hunan Key Laboratory of Traditional Chinese Medicine Prevention & Treatment of Depressive Diseases, Changsha, Hunan, China 3 The Second People's Hospital of Hunan Province, Changsha, Hunan, China 4 The First Hospital of Hunan University of Chinese Medicine, Changsha, Hunan, China 5 Hunan Academy of Chinese Medicine, Changsha, Hunan, China Correspondence should be addressed to Xiaoye Wang; 004181@hnucm.edu.cn and Jinwen Ge; 001267@hnucm.edu.cn Received 17 September 2022; Revised 17 November 2022; Accepted 13 December 2022; Published 7 January 2023 Academic Editor: Michele Fornaro Copyright © 2023 Pan Meng et al. This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Depression is a highly prevalent and heterogeneous disorder that requires new strategies to overcome depression. In this study, we aimed to investigate whether leonurine modulated hippocampal nerve regeneration in chronic and unpredictable mild stress (CUMS) rats through the SHH/GLI signaling pathway and restoring gut microbiota and microbial metabolic homeostasis. The CUMS rat model was constructed and treated with leonurine. The body weight of rats was recorded, and a series of tests were performed. Western blot was utilized to measure the expression of BDNF and 5-HT in the hippocampus. Then the expression of SHH, GLI, PTCH, and SMO were measured by qRT-PCR and western blot. The colocalization of BrdU+DCX and BrdU +NeuN was evaluated by IF. 16S rDNA high-throughput sequencing was applied to detect the composition and distribution of gut microbiota. The differential metabolites were analyzed by untargeted metabolomics. The correlation between gut microbiota and microbial metabolites was analyzed by Pearson correlation coefficient. After CUMS modeling, the body weight of rats was decreased, and the expression of BDNF and 5-HT were decreased, while the body weight was recovered, and the expression of BDNF and 5-HT were increased after leonurine treatment. Leonurine reversed the reduction in the colocalization of BrdU+DCX and BrdU+NeuN and the reduction in the levels of SHH, GLI, PTCH, and SMO induced by CUMS modeling. Leonurine also restored gut microbiota and microbial metabolites homeostasis in CUMS rats. Furthermore, Prevotellaceae_ Ga6A1_group was negatively correlated with 3-Oxocholic acid, nutriacholic acid, and cholic acid. Collectively, leonurine regulated hippocampal nerve regeneration in CUMS rats by activating the SHH/GLI signaling pathway and restoring gut microbiota and microbial metabolic homeostasis. 1. Introduction is a major risk factor for psychiatric disorders, including major depressive disorder, and can induce inflammation, Depression is a highly common and heterogeneous disorder which is known to be dysregulated in depression [4]. [1]. It is a very common psychiatric disorder, often related to Chronic stress induces neuronal death and impairs hippo- gender, genetic, environmental, and/or psychological causes campal neurogenesis, thus leading to cognitive deficits and [2], that affects many people worldwide, hinders every aspect depressive-like behaviors [5]. Depression and inflammation of their lives, and leads to many suicides each year [3]. Stress promote each other. For some patients with depression,

2 Neural Plasticity inflammation plays a key role in the pathogenesis of depres- behavior and altered hippocampal neurotransmitter levels, sion; depression also triggers a greater cytokine response to and there was a significant correlation between metabolites stressors [6]. Therefore, new strategies are needed to over- and gut microbiota [19]. However, whether leonurine come depression and inflammation. attenuates depressive symptoms and hippocampal nerve Leonurine is an active ingredient of the traditional regeneration in CUMS rats by modulating gut microbiota Chinese medicine (TCM) Herba Leonuri [7]. Leonurine and microbial metabolic homeostasis needs to be further has microcirculation-improving, antioxidant, antiapoptotic, explored. free radical scavenging, and anti-inflammatory effects, and Based on the above background, we reasonably and effectively treats cardiovascular disease in various ways. boldly speculate that leonurine may regulate hippocampal Moreover, the biological activity of leonurine is expected to nerve regeneration in CUMS rats by activating the SHH/ enable its clinical application [8]. Leonurine could promote GLI signaling pathway and restoring gut microbiota and neurite outgrowth and neurotrophic activity in nerve cells microbial metabolic homeostasis. This study enriches the [9]. Leonurine has been reported to improve cognitive dys- mechanism research of CUMS and provides a new reference function by antagonizing excitotoxic glutamate and inhibit- for treating CUMS. ing autophagy [10]. Leonurine also reduced depression-like behavior in chronic mild stress mice, which may be medi- 2. Materials and Methods ated at least in part by improving monoamine neurotrans- mitters and inhibiting neuroinflammation [11]. However, 2.1. Construction of CUMS Rat Model. Male Sprague Dawley the underlying molecular mechanism of leonurine alleviat- (SD) rats of 6-8 weeks old and weighing 250 ± 20 g were pur- ing depressive symptoms and hippocampal nerve regenera- chased from Hunan SJA Laboratory Animal Co., Ltd and tion in chronic and unpredictable mild stress (CUMS) rats raised in the Animal Experiment Center of the Hunan Uni- is still unclear, and further research is needed. versity of Chinese Medicine, maintaining an ambient room Sonic hedgehog (SHH) plays an important role in the temperature of 22 ± 1°C, a humidity of 40-60%, alternates development of the vertebrate animal central nervous system day and night for 12 h/12 h. The rats were fed with standard (CNS), and GLI is its downstream signal molecule [12]. food and available tap water. SHH/GLI signaling pathway has been recognized as a key Thirty SD rats were randomly divided into Control, mediator of many fundamental processes in embryonic CUMS and Leonurine groups, with 10 rats in each group. development, including CNS growth, patterning, and mor- CUMS procedure was as follows: Animals were individually phogenesis [13]. Previous studies have shown that maternal housed in acrylic cages for 5 weeks to induce social isolation, prenatal chronic unpredictable mild stress led to the upreg- while five cycles of stress regimens were administered ulation of GSK-3β, which in turn inhibited SHH, β-catenin, weekly, including the following stressors: 45° cage tilt Notch, and brain-derived neurotrophic factor (BDNF) (24 h), food deprivation (48 h), cold swimming (5 min at expression, thereby affecting neonatal hippocampal neuro- 4°C), tail clipping (1 min, 1 cm from distal end of tail), body development [14]. Tayyab et al. reported the antidepressant restraint (2 h), damp bedding (8 h), and strobe lighting at and neuroprotective effects of naringenin via SHH/GLI1 sig- night [20]. The rats in the CUMS group were administered naling pathway in a CUMS rat model [15]. These studies stressors in random order every day for five consecutive suggest that SHH/GLI signaling pathway may be closely weeks, without the same stressor for two consecutive days. related to the neural development of CUMS rats, but The rats in the Control group were placed in a separate whether leonurine alleviates depressive symptoms and room to avoid contact with the stressed rats, fed normally, hippocampal nerve regeneration in CUMS rats through and unaffected by any of these stressors throughout the SHH/GLI signaling pathway needs further experiments experiment. The rats in the Leonurine group were given to determine. 60 mg/kg of leonurine by gavage every day from 7 days It is well known that antidepressants can modulate the before the administration of stressors until the end of the CNS, and the gut microbiota can play a role in depression behavior test [21]. through the microbiota-gut-brain axis [16]. An imbalance Behavioral tests were performed after 4 weeks of CUMS in the microbiota-gut-brain axis reflects the constant bidi- exposure to assess the induction of depression. The body rectional communication between the CNS and the gastroin- weight of rats was recorded 1 week before modeling and testinal tract, which has been used as a hypothesis to explain 0, 1, 2, 3, and 4 weeks after modeling. The tumor tissue the pathogenesis of depression [17]. However, the effects of and stool were obtained and fixed with 4% neutral formal- antidepressants on gut microbiota function and composition dehyde or stored at -80°C. Rats were sacrificed with remain poorly understood. Studies have shown that fecal 150 mg/kg sodium pentobarbital. This study was approved microbiota transplantation (FMT) affected gut microbiota by the Laboratory Animal Ethics Committee of the Hunan in CUMS rats and induced higher levels of depressive behav- University of Chinese Medicine, and carried out according ior and neuroinflammation. Gut microbiota exacerbated to the guidelines for the care and use of laboratory animals anxiety and depression-like phenotypes by modulating pro- and the principles of laboratory animal care and protection inflammatory cytokines in the hippocampus through a dys- (LL2020111001). functional microbiota-gut-brain axis [18]. In addition, changes in fecal metabolite and plasma metabolite abun- 2.2. Sucrose Preference Test (SPT). SPT was performed dances in CUMS rats were associated with antidepressant according to previous reports in the literature [22, 23]. The
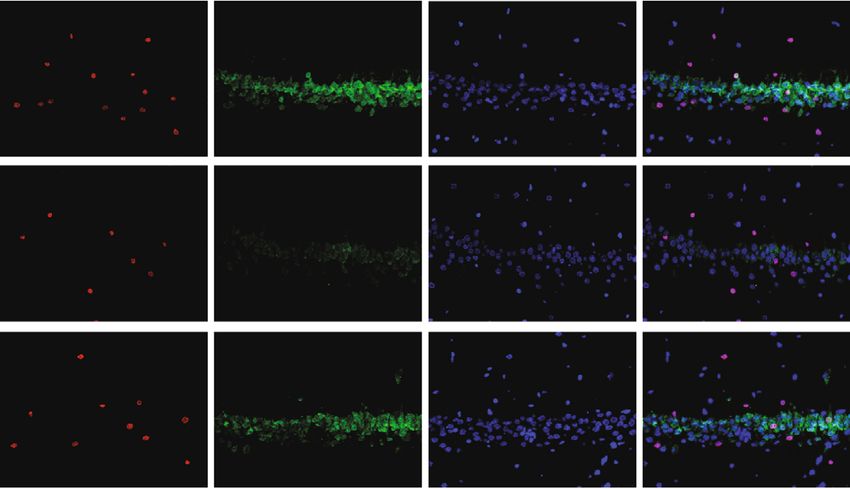
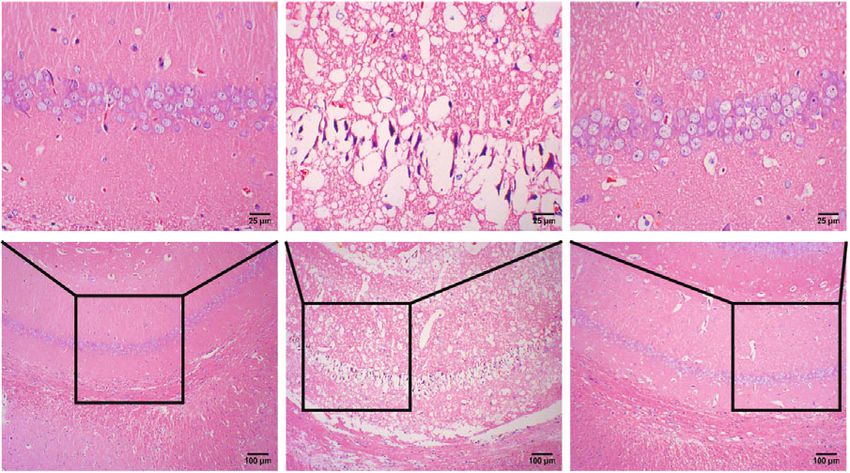
Neural Plasticity 3 100 50 ⁎ 80 40 Sucrose preference Immobility time (s) 600 # ⁎ 60 30 Body weight (g) # 400 ⁎ 40 20 200 20 10 0 0 0 –1 0 1 2 3 4 CUMS Leonurine Control CUMS Leonurine Control Weeks post model Control CUMS Leonurine (a) (b) (c) 400 20 30 ⁎ Number of climbing Number of crossing 300 15 Immobility time (s) # 20 # 200 10 # ⁎ 10 100 5 ⁎ 0 0 0 CUMS Leonurine Control CUMS Leonurine Control CUMS Leonurine Control (d) Control CUMS Leonurine 0.5 BDNF Protein/ -actin 0.4 5-HT # # 0.3 -actin ⁎ 0.2 ⁎ CUMS Leonurine Control 0.1 0.0 BDNF 5-HT Control CUMS Leonurine (e) (f) BrdU DCX DAPI Merge 800 Control BrdU+DCX+ colocalization 600 # 400 ⁎ CUMS 200 0 CUMS Control Leonurine Leonurine (g) Figure 1: Continued.
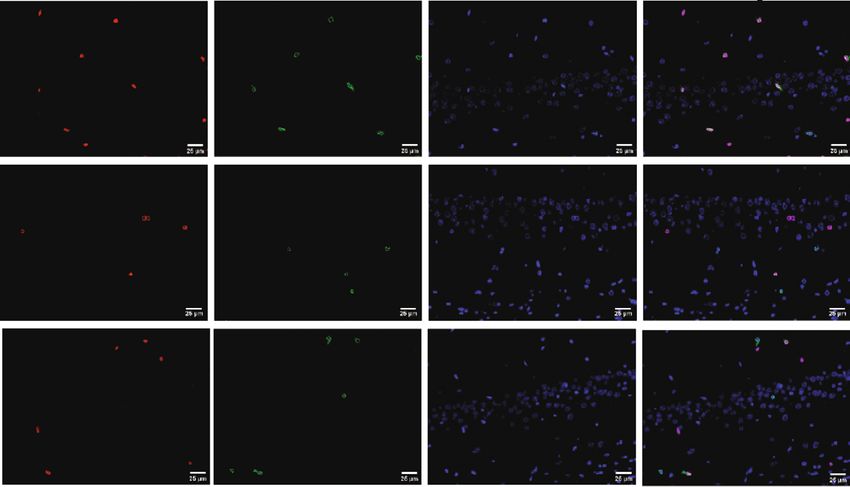
4 Neural Plasticity BrdU NeuN DAPI Merge 25000 Control BrdU+NeuN+ colocalization 20000 # 15000 ⁎ CUMS 10000 5000 0 Leonurine CUMS Leonurine Control (h) Figure 1: The effects of leonurine on behavioral indexes, inflammation, and nerve regeneration in CUMS rats. (a) Rats body weight assessment. (b) SPT. (c) OFT. (d) FST. (e) HE detection of brain tissue morphology. (f) Western blot detection of BDNF and 5-HT expressions in the hippocampus. (g, h) IF detection of the colocalization of BrdU+DCX and BrdU+NeuN in the hippocampus. ∗ P < 0:05 vs. Control. #P < 0:05 vs. CUMS. SPT included four stages of adaptation, baseline determina- hol (95-100%) was applied for dehydration. After removal, tion, testing and data analysis, which took 8 days. Sugar the slices were placed in xylene for 10 min, twice, sealed with water preference percentage = sugar water consumption/ neutral gum, and observed under microscope. ðsugar water consumption + distilled water consumptionÞ. 2.6. Quantitative Real-Time PCR (qRT-PCR). Briefly, total 2.3. Open Field Test (OFT). Animals were placed in a 60 RNA was extracted by Trizol method, RNA was reverse cm × 45 cm × 60 cm box with a bottom consisting of 12 transcribed into cDNAs using cDNA reverse transcription equal 15 cm × 15 cm squares. After the animals were placed kit (#CW2569, Beijing ComWin Biotech, China), and the in the center grid, the measurement was started, each mea- UltraSYBR Mixture (#CW2601, Beijing ComWin Biotech, surement was 5 min, and each rat only had one behavior China) was applied in the ABI 7900 system. Using β-actin measurement. After the measurement was completed, the as an internal reference gene, the relative level of the gene feces were cleaned, the bottom of the box was wiped with was calculated by the 2-ΔΔCt method. The primer sequences 75% ethanol, and the next measurement was performed. were as follows: SMO-F: ATGCGTGTTTCTTTGTGGGC, Manual records are used. The measurement indicators were SMO-R: ACACAGGATAGGGTCTCGCT; PTCH-F: CGCC immobility time, the numbers of climbing (two front paws GGGGTTTTTACACTTTC, PTCH-R: GGTCTCTTTGT leave the bottom surface), and the number of crossing (the CTGCCCTGTC; SHH-F: ATCCAAAGCTCGCATCCACT, number of 4 paws passing through the grid) within 5 min. SHH-R: CGCGTCTCGATCACGTAGAA; GLI-F: GGACTT TCTGGTCTGCCCTTT, GLI-R: GGGTGAGGTACGGATT 2.4. Forced Swimming Test (FST). In this experiment, rats ACGG; β-actin-F: ACATCCGTAAAGACCTCTATGCC, β- were placed in an open cylindrical white plastic container actin-R: TACTCCTGCTTGCTGATCCAC. (40 cm in diameter, 80 cm in height), 40 cm of water was injected into the container, the temperature was 22-25°C, 2.7. Western Blot. Western blot was used to detect the and the total recording time was 3 min. Immobility time expression of BDNF, 5-hydroxytryptamine (5-HT), SHH, (the time when the rat floated on the water surface with only GLI, patched (PTCH), and smoothened (SMO) in the hip- slight activity, or when the body was perpendicular to the pocampus. Total protein was extracted from tissues using water surface and only the nose was out of the water) was RIPA lysis buffer. BCA protein assay kits were used for pro- manually recorded. tein quantification. SDS-PAGE loading buffer was mixed, the protein was adsorbed on PVDF membrane by gel elec- 2.5. Hematoxylin-Eosin (HE) Staining. HE staining was per- trophoresis, sealed with 5% skim milk, and incubated with formed to detect the morphological changes of brain tissue. primary antibodies BDNF (28205-1-AP, 1: 1000, protein- The slices were baked at 60°C for 12 h, dewaxed to water, tech), 5-HT (ab85615, 1 μg/mL, Abcam), SHH (20697-1- hematoxylin stained for 1-10 min, washed with distilled AP, 1: 1500, proteintech), GLI (66905-1-Ig, 1: 3000, protein- water, and PBS turned blue. The slices were dyed with eosin tech), PTCH (#2468, 1: 1000, proteintech), SMO (66851-1- for 1-5 min and washed with distilled water. Gradient alco- Ig, 1: 4000, proteintech), and β-actin (66009-1-Ig, 1: 5000,
Neural Plasticity 5 1.5 1.5 1.5 1.5 PTCH mRNA expression SMO mRNA expression SHH mRNA expression GLI mRNA expression 1.0 1.0 1.0 1.0 # # # 0.5 0.5 0.5 0.5 # ⁎ ⁎ ⁎ ⁎ 0.0 Control 0.0 0.0 0.0 CUMS Leonurine Control CUMS Leonurine Control CUMS Leonurine Control CUMS Leonurine (a) (b) (c) (d) 0.5 0.6 0.6 0.6 PTCH protein/ -actin SMO protein/ -actin SHH protein/ -actin GLI protein/ -actin 0.4 # SHH 0.4 0.4 # 0.4 # 0.3 # GLI PTCH 0.2 ⁎ ⁎ ⁎ ∗ 0.2 0.2 0.2 SMO 0.1 -Actin 0.0 0.0 0.0 0.0 Control CUMS Leonurine Control CUMS Leonurine Control CUMS Leonurine Control CUMS Leonurine CUMS Leonurine Control (e) (f) (g) (h) (i) Figure 2: The effects of leonurine on the SHH/GLI signaling pathway. (a–i) SHH, GLI, PTCH, and SMO expressions in the hippocampus were measured by qRT-PCR and Western blot. ∗ P < 0:05 vs. Control. #P < 0:05 vs. CUMS. proteintech). Afterward, HRP secondary antibody was incu- libraries. Then, the composition and distribution of gut bated. ECL chemiluminescence solution (K-12045-D50, microbiota were detected. Advansta, USA) was used for chromogenic exposure with β-actin as an internal reference to detect expression levels. 2.10. Metabolomics Analysis. Fecal samples were collected from each group, and differential metabolites were analyzed 2.8. Immunofluorescence (If). The colocalization of BrdU by nontargeted metabolomics based on the ultrahigh- +DCX and BrdU+NeuN in hippocampus was detected by performance liquid chromatography coupled with mass IF. The slices were baked at 60°C for 12 h, dewaxed to water, spectrometry (UPLC-MS). UPLC system (Agilent Technolo- and the antigen was repaired by heat. The slices were placed gies, China) and Q Exactive Orbitrap (Thermo Fisher Scien- in sodium borohydride solution at room temperature for tific, USA) were used for analysis. Substance identification 30 min, and rinsed with water for 5 min. The slices were was carried out using Compound Discover (version 2.0, placed in Sudan black dye solution at room temperature Thermo, USA) and OSI-SMMS software (version 1.0, Dalian for 5 min, rinsed with water for 3 min, and sealed with 5% ChemDataSolution Information Technology Co. Ltd) in con- BSA for 60 min. The slices were incubated with BrdU junction with the McCloud database and self-built database. (ab8152, 1 : 50, Abcam), DCX (ab18723, 1 : 50, Abcam), Finally, the normalized data matrix was used for multivariate and NeuN (26975-1-AP, 1 : 50, PTG) primary antibodies statistical analysis and screening of differential metabolites. overnight at 4°C. Fluorescence secondary antibody was added and incubated at 37°C for 90 min, and DAPI was 2.11. Statistical Analysis. Graphpad Prism8.0 software was stained at 37°C for 10 min. The tablets were sealed and used for statistical analysis. The measurements were observed under the fluorescence microscope. expressed as mean ± standard deviation. Normality and homogeneity of variance tests were performed first for con- 2.9. 16S rDNA High-Throughput Sequencing. Fecal samples formity to normal distribution and homogeneity of variance. were collected from each group, and fecal DNA was One-way analysis of variance (ANOVA) was used for com- extracted. The extracted total DNA was quantified using parison between multiple groups, followed by Tukey’s post Nanodrop (ND-1000, USA), and 1-1.5% agarose gel was hoc test. Pearson correlation coefficient was applied to ana- used for quality control, such as DNA integrity. Subsequent lyze the correlation between gut microbiota and microbial PCR amplification was performed to obtain variable regions metabolites. P < 0:05 indicated that the difference was statis- of prokaryotic 16S rDNA to construct high-throughput tically significant.
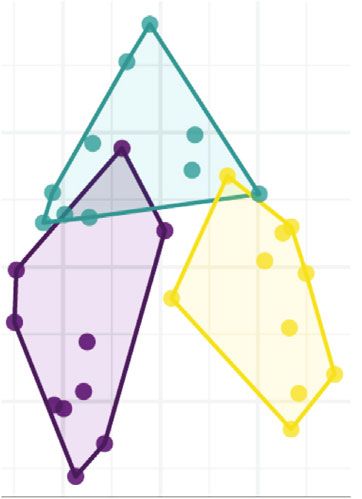
6 Neural Plasticity Control 305 CUMS 412 199 636 155 77 Leonurine 300 (a) Observe Chao1 ACE Shannon Simpson J 1000 1000 1.05 0.52 0.58 1000 0.53 0.58 0.39 0.39 0.029 0.029 0.029 0.53 0.48 0.043 0.054 0.052 0.052 0.74 1.00 0.97 0.53 5 0.8 800 800 800 0.95 0.7 0.90 600 600 600 4 0.85 0.6 400 400 400 0.80 3 CUMS Leonurine CUMS Leonurine CUMS Leonurine CUMS Leonurine CUMS Leonurine CUMS Leonurine Control Control Control Control Control Control Grouping_1 Control CUMS Leonurine (b) Figure 3: Continued.
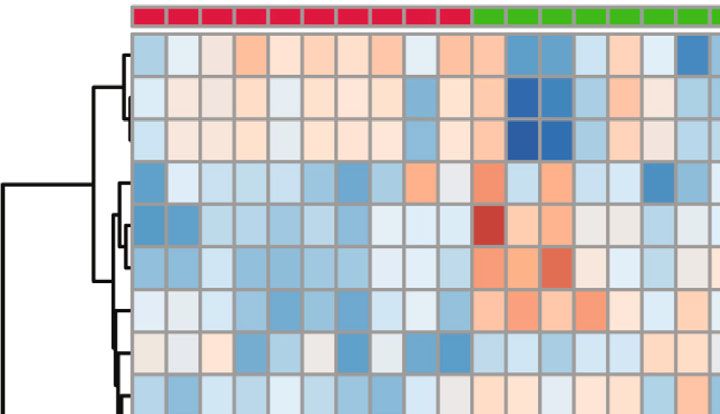
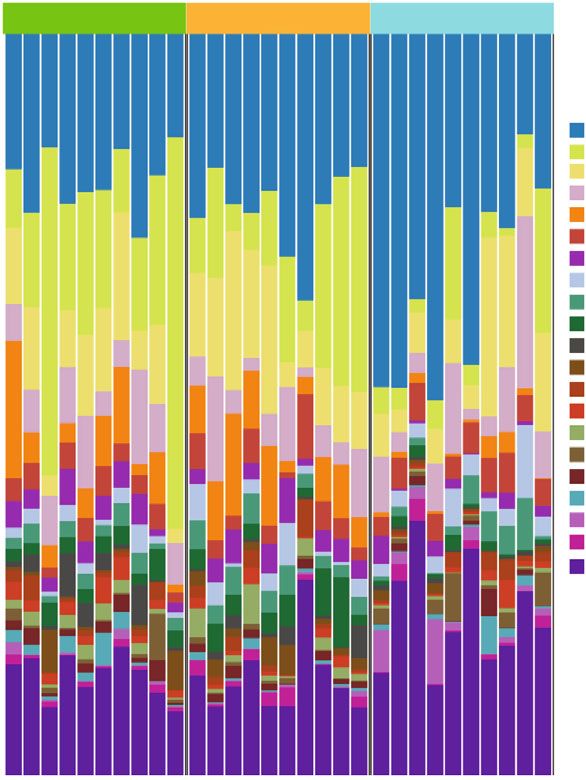
Neural Plasticity 7 PCo2 (13.9%) 0.2 0.0 –0.2 –0.2 0.0 0.2 PCo1 (20.3%) Grouping_1 Control CUMS Leonurine (c) Figure 3: The effects of leonurine on gut microbiota in CUMS rats. (a) Veen diagram: microbiota comparison of Control, CUMS, and Leonurine groups. (b) Alpha diversity analysis. (c) PCoA analysis. 3. Results 3.2. The Effects of Leonurine on SHH/GLI Signaling Pathway. Previous studies have shown that SHH/GLI signaling path- 3.1. The Effects of Leonurine on Behavioral Indexes, way may be closely related to neural development in CUMS Inflammation, and Nerve Regeneration in CUMS Rats. To rats [14, 15]. Therefore, we wanted to further explore the explore the role of leonurine, we first constructed CUMS rats effect of leonurine on the SHH/GLI signaling pathway. We model and conducted a series of tests. As shown in found that the mRNA and protein levels of SHH, GLI, Figure 1(a), compared with the Control group, the body PTCH, and SMO were decreased after CUMS modeling weight of rats was decreased after CUMS modeling, while compared to the Control group. After leonurine treatment, the body weight was increased after leonurine treatment. the mRNA and protein levels of SHH, GLI, PTCH, and Sucrose preference was reduced in the CUMS group com- SMO were increased (Figures 2(a)–2(i)). These results sug- pared to the Control group. However, sucrose preference gested that leonurine attenuated depressive symptoms and was increased after leonurine treatment (Figure 1(b)). OFT hippocampal nerve regeneration in CUMS rats via SHH/ and FST results showed that the immobility time of rats GLI signaling pathway. was increased after CUMS modeling, and the numbers of crossing and climbing were decreased compared with the 3.3. The Effects of Leonurine on Gut Microbiota in CUMS Control group. The immobility time was decreased and the Rats. It was reported that gut microbiota could play a role numbers of crossing and climbing were increased after leo- in depression through microbiota-gut-brain axis [16]. nurine treatment (Figures 1(c) and 1(d)). HE staining results Therefore, we wanted to explore the effect of leonurine on showed that hippocampal neurons in the Control group had the gut microbiota of CUMS rats, and detected the composi- complete structure, regular cell morphology, normal inter- tion and distribution of gut microbiota by 16S rDNA high- cellular space, neat arrangement, and clear nucleoli. In the throughput sequencing technology. Venn plots showed the CUMS group, neurons were atrophic and loosely arranged, common and unique amplicon sequence variants (ASVs) intercellular space was enlarged, some cells were vacuolated, among the groups. Among them, there were 412 unique and the nuclei were pyknotic. The number of hippocampal ASVs in the Control group, 199 unique ASVs in the CUMS neurons in the Leonurine group was increased, the intercel- group, 300 unique ASVs in the Leonurine group, and 636 lular space was narrowed, the arrangement was more common ASVs in all three groups (Figure 3(a)). Alpha orderly, and the histopathological changes were improved diversity was the analysis of species diversity within a single (Figure 1(e)). In addition, the expressions of BDNF and 5- sample. Compared with the Control group, the Observe, HT were decreased after CUMS modeling, and the expres- Chao1, ACE, and Shannon indexes of the CUMS group were sions of BDNF and 5-HT were increased after leonurine decreased and further decreased in the leonurine group. treatment (Figure 1(f)). Afterwards, we used IF to detect There was no significant difference between the three groups the colocalization of BrdU+DCX and BrdU+NeuN in the in Simpson index. However, J index was increased in CUMS hippocampus. The colocalization of BrdU+DCX and BrdU group compared to the Control group, and J index was +NeuN were decreased after CUMS modeling, and the colo- decreased after leonurine treatment (Figure 3(b)). The β- calization of BrdU+DCX and BrdU+NeuN were increased diversity was calculated and visualized by PCoA, and as after leonurine treatment (Figures 1(g) and 1(h)). These shown in Figure 3(c), there were differences in species abun- results revealed that leonurine alleviated depressive symp- dance among the three groups of samples. The relative abun- toms and hippocampal nerve regeneration in CUMS rats. dance histogram of top 20 ASVs at the genus level further
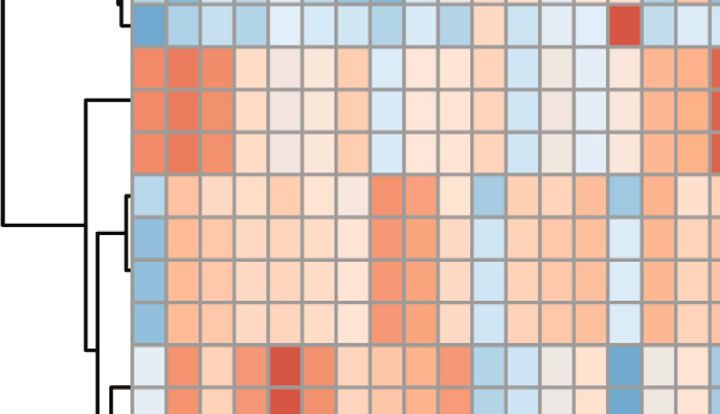
8 Neural Plasticity 100 Control CUMS Leonurine Bacteroidota; Muribaculaceae Bacteroidota; Prevotella Firmicutes; Lactobacillus g_g_Muribaculaceae g_g_Prevotella Firmicutes;Lachnospiraceae_NK4A136_group g_un_f_f_Lachnospiraceae Firmicutes; Ruminococcus g_g_Lactobacillus 75 g_g_Lachnospiraceae_NK4A136_group Firmicutes; Clostridia_UCG-014 g_un_f_f_Oscillospiraceae g_g_Ruminococcus Bacteroidota; Prevotellaceae_Ga6A1_group Relative abundance (%) g_un_c_c_Bacilli g_g_Clostridia_UCG-D14 Firmicutes; c_Bacilli_ASV_5 g_g_Prevotellaceae_Ga6A1_group Firmicutes; Quinella g_g_UCG–005 g_un_f_f_Prevotellaceae Firmicutes; Roseburia g_g_Helicobacter 50 g_g_Roseburia Firmicutes; f_Oscillospiraceae_ASV_20 g_un_f_f_Ruminococcaceae g_g_Prevotellaceae_UCG-001 Campilobaterota; Helicobacter g_g_Quinella Firmicutes; uncultured g_g_[Eubacterium]_siraeum_group g_g_Blautia Bacteroidota; Alloprevotella g_g_Alloprevotella Others Firmicutes; [Eubacterium]_coprostanoligenes_group 25 Firmicutes; Monoglobus Desulfobacterota; uncultured Firmicutes; Oscillibacter Bacteroidota; Prevotellaceae_UCG–001 Cyanobacteria; Gastranaerophilales 0 A1 A2 A3 A4 A5 A6 A7 A8 A9 A10 B1 B2 B3 B4 B5 B6 B7 B8 B9 B10 C1 C2 C3 C4 C5 C6 C7 C8 C9 C10 0 10 20 30 40 50 Read abundance (%) Group Leonurine CUMS Control (a) (b) Abundance of Prevotellaceae_Ga6A1_group 15 15 5 8 Abundance of Clostridia_UCG-014 Lachnospiraceae_NK4A136_group Abundance of Lactobacillus 4 6 10 Abundance of 10 3 4 2 5 5 2 1 0 0 0 0 Control CUMS Leonurine Leonurine Leonurine Leonurine CUMS CUMS CUMS Control Control Control (c) (d) (e) (f) Figure 4: The effects of leonurine on gut microbiota in CUMS rats. (a) Top 20 dominant gut microbiota with relative abundance at the genus level. (b) Relative abundance of microbiota at the genus level. (c–f) Relative abundance comparison of Lactobacillus, Lachnospiraceae_NK4A136_group, Clostridia_UCG-014, and Prevotellaceae_Ga6A1_group in the Control, CUMS, and Leonurine groups. showed that the abundance and composition of the micro- of this technique and identify relevant biomarkers. Among bial community in each group were different. (Figure 4(a)). them, the index of the differential factor variable was Figure 4(b) further showed the relative abundance of the 27.4%, which indicated that the three groups of samples top 20 dominant gut microbiota at the genus level. We were basically separated (Figure 5(a)). Heat map showed found that the abundance of Lactobacillus was decreased that the top 25 potential metabolites in the Control, but the abundances of Lachnospiraceae_NK4A136_group, CUMS, and Leonurine groups. These potential metabolites Clostridia_UCG-014, and Prevotellaceae_Ga6A1_group were included 3-Oxocholic acid, nutriacholic acid, and cholic increased after CUMS modeling compared to the Control acid (Figure 5(b)). Collectively, leonurine restored micro- group. Leonurine treatment reversed the abundances of bial metabolite changes in CUMS rats. these gut microbiotas (Figures 4(c) and 4(f)). These results indicated that leonurine restored gut microbiota homeosta- sis in CUMS rats. 3.5. Correlation Analysis of Key Gut Microbiota and Microbial Metabolites. In order to investigate whether key gut microbiota and microbial metabolites were correlated, 3.4. The Effects of Leonurine on Microbial Metabolites in we performed Pearson correlation analysis. Figure 6 showed CUMS Rats. At the same time, we performed untargeted that Prevotellaceae_Ga6A1_group was negatively correlated metabolomics to analyze the differential metabolites. Partial with 3-Oxocholic acid (R = −0:38, P < 0:05), nutriacholic least sums discriminate analysis (PLS-DA) was applied to acid (R = −0:45, P < 0:05), and cholic acid (R = −0:44, P < analyze metabolomic profile changes in the Control, CUMS, 0:05). While Lactobacillus, Lachnospiraceae_NK4A136_ and Leonurine groups to eliminate any non-specific effects group and Clostridia_UCG-014 were not correlated with 3-
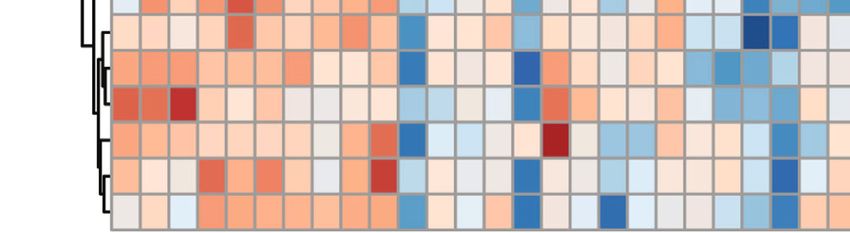
Neural Plasticity 9 Scores Plot 20 10 Component 2 (11.3 %) 5 0 –5 –10 –15 –20 –10 0 10 20 Component 1 (27.4 %) C M L (a) class 3 3–Oxocholic acid Nutriacholic acid Cholic acid Octadecanedioic ac 2 Pentadecanoic acid Tridecanoic acid Methorimeprazine METHYL ACETOACETAT 1 CORTISOL Pregnenolone sulfa D–Erythronolactone METHYLMALONATE 0 SUCCINATE PHENOL SALICYLATE 3–Hydroxybenzoic a 4–HYDROXYBENZOATE –1 TRIGONELLINE Methyl Nicotinate 3–Indolepropionic EICOSAPENTAENOATE –2 (3,4–Dimethoxyphen Daidzein Nonadecanoic acid STACHYOSE –3 A1 A2 A3 A4 A5 A6 A7 A8 A9 A10 B1 B2 B3 B4 B5 B6 B7 B8 B9 B10 C1 C2 C3 C4 C5 C6 C7 C8 C9 C10 Class Control CUMS Leonurine (b) Figure 5: The effects of leonurine on microbial metabolites in CUMS rats. (a) PLS-DA analysis of metabolic profile changes. (b) Heat map showed the metabolic profiles of rats in Control, CUMS, and Leonurine groups.
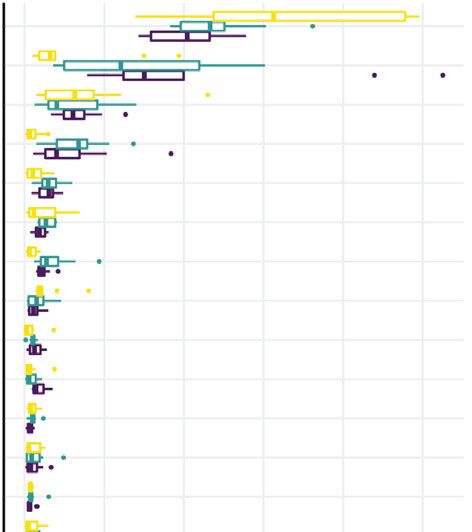
10 Neural Plasticity leonurine alleviated depressive symptoms and hippocampal 3–Oxocholic acid ∗ 0.2 nerve regeneration in CUMS rats. SHH signaling pathway plays a vital role in the develop- 0 ment of many tissues and organs [29]. SHH signaling path- Nutriacholic acid ∗ way consists of three main components: PTCH, SMO, and –0.2 GLI [30]. Abnormal signaling pathways of SHH lead to Cholic acid ∗ many neurological diseases, including depression [12]. –0.4 SHH may be involved in the pathophysiology of depression [31]. However, the mechanism of SHH signaling pathway is Clostridia_UCG-014 Lactobacillus Lachnospiraceae_NK4A136_group Prevotellaceae_Ga6A1_group complex and still not fully understood. Studies have shown that SHH could regulate adult hippocampal neurogenesis [32]. Mahino et al. reported that SHH expression downreg- ulated in the CUMS induced prenatal stress in the hippo- campus [14]. In this study, we treated CUMS rats with leonurine and found that leonurine reversed the reduction of SHH, GLI, PTCH, and SMO levels caused by CUMS. Our study confirmed that the reduction of depressive symptoms and hippocampal nerve regeneration in CUMS rats by leonurine was mediated through SHH/GLI signal- Figure 6: Correlation analysis of key gut microbiota and microbial ing pathway. metabolites. Pearson correlation coefficient analyzed the correlation TCM may regulate gut microbiota and microbial meta- between gut microbiota and microbial metabolites. ∗ P < 0:05. bolic homeostasis. Chi et al. demonstrated that oral admin- istration of inulin-type fructooligosaccharides extracted from Morinda officinalis alleviated intestinal epithelial dam- Oxocholic acid, nutriacholic acid, and cholic acid. These age, restores fecal microbial homeostasis, and relieves results suggested that Prevotellaceae_Ga6A1_group might depressive symptoms in CUMS rats [33]. Total glycosides play an important role in CUMS rats. isolated from the dried succulent stems of Cistanche deserti- cola alleviated low-grade inflammation of colonic and intes- 4. Discussion tinal barrier disruption, restored disturbed gut microbiota, and then depression-like behavior was reduced [34]. In addi- Depression and related mood disorders pose a huge burden tion, TCM restored gut microbiota and brain metabolic on health, quality of life, and the global economy [24]. microbial homeostasis, alleviating depressive behaviors Stress-induced failure of the brain’s ability to restore plastic- induced by CUMS, and there were strong correlation ity may contribute to the onset and relapse of depression between brain metabolites and disturbed gut microbiota [25]. CUMS is currently the most commonly used, reliable, [35]. In the present study, we also found that leonurine and valid rodent model of depression [26]. In this study, attenuated depressive symptoms and hippocampal nerve we performed CUMS rats model to explore the effect and regeneration in CUMS rats by regulating gut microbiota mechanism of leonurine. Our study revealed that leonurine and microbial metabolic homeostasis; that was, leonurine regulated hippocampal nerve regeneration in CUMS rats restored gut microbiota and microbial metabolite homeo- by activating SHH/GLI signaling pathway and restoring stasis in CUMS rats. Moreover, Zhang et al. found the gut microbiota and microbial metabolic homeostasis. This enrichment of Prevotellaceae-Ga6A1-group in the gastric is the first time we report the effect of leonurine on CUMS microbiota of patients with gastric intraepithelial neoplasia rats through the SHH/GLI signaling pathway. [36]. We also found that Prevotellaceae_Ga6A1_group was Leonurine, the main bioactive component of Herba increased after CUMS modeling and decreased after leonur- Leonuri, has shown therapeutic potential in depression ine treatment. Furthermore, the enrichment of 3-Oxocholic [27]. Meng et al. reported that leonurine promotd neurite acid in noncachectic patients was correlated with gut outgrowth and neurotrophic activity in cultured PC12 cells, microbiota Lactobacillus gasseri [37]. Alam et al. identified and the mechanism may involve GR/SGK1 signaling path- nutriacholic acid, a novel triterpenoid, in DIBEt for the first way [9]. Studies have shown that depression-like behavior time. DIBEt has the potential to defend against oxidative could be effectively reversed by increasing the expression stress stimulation [38]. In addition, cholic acid was level of neurotrophic factors in the rat brain [28]. Jia et al. increased markedly in plasma of the depression rats [39]. revealed that administration of leonurine for 4 weeks signif- Through correlation analysis, we found that Prevotella- icantly reduced depressive-like behaviors in chronic mildly ceae_Ga6A1_group was associated with 3-Oxocholic acid, stressed mice, including increased sucrose preference and nutriacholic acid and cholic acid, which suggested that Pre- decreased immobility time in FST. At the same time, leonur- votellaceae_Ga6A1_group might play acrucial role in CUMS ine could effectively restore the level of 5-HT in the hippo- rats. Although the current study was conducted in rats, campus and prefrontal cortex of chronic mildly stressed CUMS model we used was widely accepted as an ideal mice, and improve the damage of hippocampal neurons model for studying the antidepressant effects of various poten- [11]. Consistent with their results, our study confirmed that tial therapeutic agents. However, the effect of leonurine on
Neural Plasticity 11 SHH/GLI signaling pathway in humans needs to be further [2] S. Sonali, B. Ray, H. Ahmed Tousif et al., “Mechanistic insights investigated. into the link between gut dysbiosis and major Depression: An Extensive Review,” Cells, vol. 11, no. 8, p. 1362, 2022. 5. Conclusion [3] M. A. Tayab, M. N. Islam, K. A. A. Chowdhury, and F. M. Tas- nim, “Targeting neuroinflammation by polyphenols: a promis- In this study, we performed CUMS modeling and explored ing therapeutic approach against inflammation-associated the role and mechanism of leonurine in CUMS rat model depression,” Biomedicine & Pharmacotherapy, vol. 147, article by in vivo experiments, 16S rDNA high-throughput 112668, 2022. sequencing and metabolomics analysis. Our study revealed [4] T. C. Franklin, C. Xu, and R. S. Duman, “Depression and ster- for the first time that leonurine modulated hippocampal ile inflammation: essential role of danger associated molecular nerve regeneration in CUMS rats via SHH/GLI signaling path- patterns,” Brain, Behavior, and Immunity, vol. 72, pp. 2–13, way and restoring gut microbiota and microbial metabolic 2018. homeostasis. This study provides a reference for exploring [5] Y. Ge, Y. Chen, C. Guo et al., “Pyroptosis and intervertebral the mechanism of action of leonurine in depression. disc degeneration: mechanistic insights and therapeutic impli- cations,” Journal of Inflammation Research, vol. Volume 15, pp. 5857–5871, 2022. Data Availability [6] J. K. Kiecolt-Glaser, H. M. Derry, and C. P. Fagundes, “Inflam- The datasets used and analysed during the current study are mation: depression fans the flames and feasts on the heat,” The American Journal of Psychiatry, vol. 172, no. 11, pp. 1075– available from the corresponding author on reasonable 1091, 2015. request. [7] T. Xu, X. Li, T. Leng et al., “CYP2A13 acts as the main meta- bolic CYP450s enzyme for activating leonurine in human Ethical Approval bronchial epithelial cells,” Medical Science Monitor, vol. 26, 2020. This study was approved by the Laboratory Animal Ethics Committee of the Hunan University of Chinese Medicine [8] Y. Z. Zhu, W. Wu, Q. Zhu, and X. Liu, “Discovery of leonuri and therapeutical applications: from bench to bedside,” Phar- and carried out according to the guidelines for the care macology & Therapeutics, vol. 188, pp. 26–35, 2018. and use of laboratory animals and the principles of labora- [9] P. Meng, Q. Zhu, H. Yang et al., “Leonurine promotes neurite tory animal care and protection (LL2020111001). outgrowth and neurotrophic activity by modulating the GR/ SGK1 signaling pathway in cultured PC12 cells,” Neuroreport, Conflicts of Interest vol. 30, no. 4, pp. 247–254, 2019. [10] C. Liu, H. Yin, J. Gao, X. Xu, T. Zhang, and Z. Yang, “Leonurine The authors declare that they have no conflict of interest. ameliorates cognitive dysfunction via antagonizing excitotoxic glutamate insults and inhibiting autophagy,” Phytomedicine, Authors’ Contributions vol. 23, no. 13, pp. 1638–1646, 2016. [11] M. Jia, C. Li, Y. Zheng et al., “Leonurine exerts antidepressant- Pan Meng, Xi Zhang, Dandan Li, Xiaoye Wang, and Jinwen like effects in the chronic mild stress-induced depression Ge contributed to the conception and design of the study. model in mice by inhibiting neuroinflammation,” The Interna- Pan Meng, Hui Yang, and Xiaoyuan Lin collected the infor- tional Journal of Neuropsychopharmacology, vol. 20, no. 11, mation. Pan Meng wrote the first draft of the manuscript. pp. 886–895, 2017. Hongqing Zhao, Ping Li, and Yuhong Wang wrote sections [12] X. Li, Y. Li, S. Li, H. Li, C. Yang, and J. Lin, “The role of Shh of the manuscript. All authors have revised, read, and signalling pathway in central nervous system development approved the final version of the manuscript. and related diseases,” Cell Biochemistry and Function, vol. 39, no. 2, pp. 180–189, 2021. Acknowledgments [13] P. W. Ingham and A. P. McMahon, “Hedgehog signaling in animal development: paradigms and principles,” Genes & This work was funded by grants from the National Natural Development, vol. 15, no. 23, pp. 3059–3087, 2001. Science Foundation of China (No.81403387), the Outstand- [14] M. Fatima, S. Srivastav, M. H. Ahmad, and A. C. Mondal, ing Youth Project of Natural Science Foundation of Hunan “Effects of chronic unpredictable mild stress induced prenatal Province (No.2020JJ3027), Excellent Postdoctoral Innova- stress on neurodevelopment of neonates: role of GSK-3β,” Sci- tive Talents of Hunan Province (No.2020RC2060), the entific Reports, vol. 9, no. 1, p. 1305, 2019. Training Plan of Outstanding Innovative Youth of Changsha [15] M. Tayyab, S. Farheen, P. M. Mubeena Mariyath, N. Khanam, (No.kq2009018), Natural Science Foundation of Hunan M. M. Hossain, and M. H. Shahi, “Antidepressant and neuropro- Province (2019JJ50432). tective effects of naringenin via sonic hedgehog-GLI1 cell signal- ing pathway in a rat model of chronic unpredictable mild stress,” References Neuromolecular Medicine, vol. 21, no. 3, pp. 250–261, 2019. [16] W. Zhang, W. Qu, H. Wang, and H. Yan, “Antidepressants flu- [1] I. Badini, J. R. I. Coleman, S. P. Hagenaars et al., “Depression oxetine and amitriptyline induce alterations in intestinal with atypical neurovegetative symptoms shares genetic predis- microbiota and gut microbiome function in rats exposed to position with immuno-metabolic traits and alcohol consump- chronic unpredictable mild stress,” Translational Psychiatry, tion,” Psychological Medicine, vol. 52, no. 4, pp. 726–736, 2022. vol. 11, no. 1, p. 131, 2021.
12 Neural Plasticity [17] Y. Du, X.-R. Gao, L. Peng, and J.-F. Ge, “Crosstalk between the [33] L. Chi, I. Khan, Z. Lin et al., “Fructo-oligosaccharides from microbiota-gut-brain axis and depression,” Heliyon, vol. 6, _Morinda officinalis_ remodeled gut microbiota and alleviated no. 6, article e04097, 2020. depression features in a stress rat model,” Phytomedicine, [18] N. Li, Q. Wang, Y. Wang et al., “Fecal microbiota transplanta- vol. 67, article 153157, 2020. tion from chronic unpredictable mild stress mice donors [34] L. Fan, Y. Peng, J. Wang, P. Ma, L. Zhao, and X. Li, “Total gly- affects anxiety-like and depression-like behavior in recipient cosides from stems of _Cistanche tubulosa_ alleviate mice via the gut microbiota-inflammation-brain axis,” Stress, depression-like behaviors: bidirectional interaction of the phy- vol. 22, no. 5, pp. 592–602, 2019. tochemicals and gut microbiota,” Phytomedicine, vol. 83, arti- [19] L. Jianguo, J. Xueyang, W. Cui, W. Changxin, and Q. Xuemei, cle 153471, 2021. “Altered gut metabolome contributes to depression-like [35] W. Qu, S. Liu, W. Zhang et al., “Impact of traditional Chinese behaviors in rats exposed to chronic unpredictable mild medicine treatment on chronic unpredictable mild stress- stress,” Translational Psychiatry, vol. 9, no. 1, p. 40, 2019. induced depression-like behaviors: intestinal microbiota and [20] L. Wei, Y. Li, W. Tang et al., “Chronic unpredictable mild gut microbiome function,” Food & Function, vol. 10, no. 9, stress in rats induces colonic inflammation,” Frontiers in Phys- pp. 5886–5897, 2019. iology, vol. 10, p. 1228, 2019. [36] X. Zhang, C. Li, W. Cao, and Z. Zhang, “Alterations of gastric [21] Z. Y. Hong, X. R. Shi, K. Zhu, T. T. Wu, and Y. Z. Zhu, “SCM- microbiota in gastric cancer and precancerous stages,” Fron- 198 inhibits microglial overactivation and attenuates Aβ(1- tiers in Cellular and Infection Microbiology, vol. 11, article 40)-induced cognitive impairments in rats via JNK and NF- 559148, 2021. кB pathways,” Journal of Neuroinflammation, vol. 11, no. 1, [37] Y. Ni, Z. Lohinai, Y. Heshiki et al., “Distinct composition and p. 147, 2014. metabolic functions of human gut microbiota are associated [22] M. Y. Liu, C. Y. Yin, L. J. Zhu et al., “Sucrose preference test for with cachexia in lung cancer patients,” The ISME Journal, measurement of stress-induced anhedonia in mice,” Nature vol. 15, no. 11, pp. 3207–3220, 2021. Protocols, vol. 13, no. 7, pp. 1686–1698, 2018. [38] M. B. Alam, A. Ahmed, S. Islam et al., “Phytochemical charac- [23] M. A. Schalla, S. G. Kühne, T. Friedrich et al., “Sucrose prefer- terization of Dillenia indica L. bark by paper spray ionization- ence and novelty-induced Hypophagia tests in rats using an mass spectrometry and evaluation of its antioxidant potential automated food intake monitoring system,” Journal of Visual- against t-BHP-induced oxidative stress in RAW 264.7 cells,” ized Experiments, no. 159, 2020. Antioxidants (Basel), vol. 9, no. 11, 2020. [24] E. S. Williams, M. Mazei-Robison, and A. J. Robison, “Sex Dif- [39] F. Zhang, Z. Jia, P. Gao et al., “Metabonomics study of urine ferences in Major Depressive Disorder (MDD) and Preclinical and plasma in depression and excess fatigue rats by ultra fast Animal Models for the Study of Depression,” Cold Spring Har- liquid chromatography coupled with ion trap-time of flight bor Perspectives in Biology, vol. 14, no. 3, 2022. mass spectrometry,” Molecular BioSystems, vol. 6, no. 5, [25] W. Liu, X. Xue, J. Xia, J. Liu, and Z. Qi, “Swimming exercise pp. 852–861, 2010. reverses CUMS-induced changes in depression-like behaviors and hippocampal plasticity-related proteins,” Journal of Affec- tive Disorders, vol. 227, pp. 126–135, 2018. [26] S. Antoniuk, M. Bijata, E. Ponimaskin, and J. Wlodarczyk, “Chronic unpredictable mild stress for modeling depression in rodents: Meta- analysis of model reliability,” Neuroscience and Biobehavioral Reviews, vol. 99, pp. 101–116, 2019. [27] X. Liu, W. Cao, J. Qi et al., “Leonurine ameliorates adriamycin- induced podocyte injury via suppression of oxidative stress,” Free Radical Research, vol. 52, no. 9, pp. 952–960, 2018. [28] J. Fu, Y. Zhang, R. Wu et al., “Shuganjieyu capsule increases neurotrophic factor expression in a rat model of depression,” Neural Regeneration Research, vol. 9, no. 5, pp. 489–497, 2014. [29] A. R. I. Altaba, V. Nguyên, and V. Palma, “The emergent design of the neural tube: prepattern, SHH morphogen and GLI code,” Current Opinion in Genetics & Development, vol. 13, no. 5, pp. 513–521, 2003. [30] M. Frank-Kamenetsky, X. M. Zhang, S. Bottega et al., “Small- molecule modulators of hedgehog signaling: identification and characterization of smoothened agonists and antagonists,” Journal of Biology, vol. 1, no. 2, p. 10, 2002. [31] M. Tayyab, M. H. Shahi, S. Farheen et al., “Sonic hedgehog, Wnt, and brain-derived neurotrophic factor cell signaling pathway crosstalk: potential therapy for depression,” Journal of Neuroscience Research, vol. 96, no. 1, pp. 53–62, 2018. [32] P. J. Yao, R. S. Petralia, and M. P. Mattson, “Sonic hedgehog signaling and hippocampal neuroplasticity,” Trends in Neuro- sciences, vol. 39, no. 12, pp. 840–850, 2016.
You can also read

















































