JMJD2C-mediated long non-coding RNA MALAT1/microRNA- 503-5p/SEPT2 axis worsens non-small cell lung cancer
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
www.nature.com/cddis
ARTICLE OPEN
JMJD2C-mediated long non-coding RNA MALAT1/microRNA-
503-5p/SEPT2 axis worsens non-small cell lung cancer
1✉
Jun Zhang1, Mingliang Wang1, Jiashun Wang1 and Wendong Wang
© The Author(s) 2022
Jumonji domain containing protein 2C (JMJD2C) could epigenetically regulate cancer cells. We specifically explored the
downstream mechanism of JMJD2C in non-small cell lung cancer (NSCLC) from the long non-coding RNA metastasis associated
with lung adenocarcinoma transcript 1/microRNA-503-5p/septin 2 (MALAT1/miR-503-5p/SEPT2) axis. NSCLC clinical tissues were
utilized to assess JMJD2C, MALAT1, miR-503-5p and SEPT2 levels. NSCLC cell lines (A549 and H1299) were applied for loss-of-
function and gain-of-function tests to identify the functional roles of JMJD2C, MALAT1, miR-503-5p, and SEPT2. The interactions
among JMJD2C, MALAT1, miR-503-5p, and SEPT2 were assessed. Augmented JMJD2C, MALAT1, and SEPT2 and reduced miR-503-5p
levels were found in NSCLC. Depleting JMJD2C or MALAT1, or restoring miR-503-5p exerted anti-tumor effects on NSCLC cells
in vitro and in vivo. JMJD2C is bound to the promoter of MALAT1. MALAT1 bound to miR-503-5p and miR-503-5p targeted SEPT2.
Knocking down MALAT1 or SEPT2, or elevating miR-503-5p mitigated the pro-tumor effects of upregulated JMJD2C on NSCLC. It is
1234567890();,:
evident that the JMJD2C-mediated MALAT1/miR-503-5p/SEPT2 axis takes part in the process of NSCLC and even worsens NSCLC.
Cell Death and Disease (2022)13:65 ; https://doi.org/10.1038/s41419-022-04513-5
INTRODUCTION individualized therapy for NSCLC [15]. As a member of miRNA,
Lung cancer is composed of subpopulations of cells or clones with miR-503 has the capacity to retard NSCLC [16] and mediate
different molecular characteristics, resulting in intratumoral cisplatin resistance [17]. miR-503-5p is an inhibitory effector in
heterogeneity [1]. Non-small cell lung cancer (NSCLC) takes the ovarian cancer that could be sponged by MALAT1 [18]. Based on
majority of lung cancer cases, of which about a quarter are that, we would like to know whether MALAT1/miR-503-5p works in
classified as locally advanced disease, otherwise known as stage III NSCLC cell progression. Septins (SEPTs) are connected to the
disease [2]. Late diagnosis of lung cancer primarily blames on cytosolic actin cytoskeleton in matrix-attached cancer cells and act
asymptom in early-stage, and misdiagnosis with early symptoms critically in metastatic cancer cells [19]. Through bioinformatics
[3]. Stage-based treatments vary in patients with NSCLC which are software, we found that SEPT2 was a predicted target of miR-503-
comprised of complete surgical resection, conventional, or 5p and studies have partially explored SEPT2-related mechanisms
stereotactic radiotherapy, and percutaneous thermal ablation [4]. in cancers from its feedback with lncRNA-miRNA [20, 21].
Having a thorough insight into the mechanism of NSCLC could Thus, we conducted the research with the hypothesis that
facilitate to managing the disease. JMJD2C-mediated histone methylation of MALAT1 stimulates the
JMJD2 histone demethylases are thought to be epigenetic progression of NSCLC by interacting with miR-503-5p and SEPT2.
mediators in cancers [5] and JMJD2 overexpression has oncogenic
potential [6]. In addition, JMJD2s have been reported to
dramatically correlate with cisplatin resistance in NSCLC [7]. METHODS AND MATERIALS
Concerning JMJD2C, some research has pointed out that it drives Ethics statement
cancer migration and invasion in lung cancer [8] and mediates All patients signed an informed consent form. Experimental approval was
tumorigenesis in osteosarcoma [9]. Given that JMJD2C promotes obtained from the ethics committee of Union Hospital, Tongji Medical
colorectal cancer (CRC) metastasis via histone methylation of College, Huazhong University of Science and Technology. Animal research
metastasis-related lung adenocarcinoma transcript 1 (MALAT1) [10], has been approved by the animal care and use Committee of Union
Hospital, Tongji Medical College, Huazhong University of Science and
it is unclear whether and how JMJD2C-mediated downstream Technology.
involving MALAT1 functions in NSCLC. MALAT1 has been identified
as an onco-lncRNA involved in the promotion of NSCLC [11]. It is
noted that MALAT1 confers acquired malignant phenotypes of lung Subjects
cancer cells [12, 13], as well as enhanced drug resistance for NSCLC Tissue specimens (cancer tissues and adjacent tissues) were acquired from
cells by cooperation with microRNAs [14]. Altered miRNA expres- 116 patients with NSCLC in Union Hospital, Tongji Medical College, Huazhong
sion profiles have diagnostic value and can even program University of Science and Technology. The patients including 69 males and 47
1
Department of Thoracic Surgery, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, 430022 Wuhan, China.
✉email: wendong027@163.com
Edited by Professor Roberto Mantovan
Received: 24 March 2021 Revised: 14 December 2021 Accepted: 10 January 2022
Official journal of CDDpress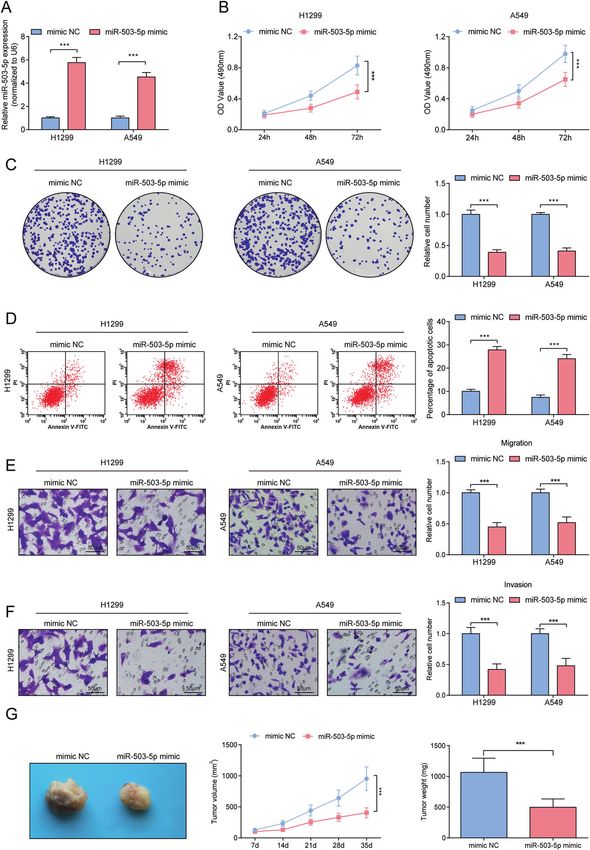
J. Zhang et al.
2
females were aged at 46–72 years old. All patients were diagnosed as NSCLC Western blot assay
by histopathology and received no pre-operation treatments. After lysis by radio-immunoprecipitation assay lysis solution (Beyotime),
the tissues and cells were centrifuged and the supernatant was collected.
The protein concentration was measured by using the BCA kit (Beyotime)
Cell culture The proteins (30 μg) were added with β-mercaptoethano, boiled for
Normal lung bronchial epithelial cell line (BEAS-2B) and lung cancer cell 10 min, and separated by sodium dodecyl sulphate polyacrylamide gel
line (A549) were cultured in Dulbecco’s modified Eagle medium (DMEM; electrophoresis. Then, the protein transferred to a nitrocellulose mem-
Gibco, CA, USA) while another lung cancer cell line (H1299) in Roswell Park brane (Bio-Rad, CA, USA) was combined with the primary antibodies, anti-
Memorial Institute-1640 medium (Gibco). The above medium was SEPT2 (1:500, Proteintech, USA), anti-JMJD2C (1:500), and anti-GAPDH
supplemented with 10% fetal bovine serum (FBS; Gibco), penicillin (500 (1:1000, both from Abcam, MA, USA), as well as the secondary antibody for
units/mL) and streptomycin (200 µg/mL). The cell lines were from BeNa
reaction. The membrane was treated with enhanced chemiluminescence
culture collection (Beijing, China).
reagent (SuperSignal Western Pico Chemiluminescent Substrate; Pierce,
USA)). Image J software (National Institutes of Health) was applied to
Cell transfection protein quantitative analysis. The gray value of the band was normalized
Transfection was conducted with Lipofectamine 3000 (Invitrogen, CA, USA) to GAPDH.
according to the manufacturer’s instructions. A549 and H1299 cells were
transfected with sh-NC, sh-JMJD2C, sh-CTR(control shRNA with scrambled Dual luciferase reporter gene assay
sequence), sh-MALAT1, mimic NC, miR-503-5p mimic, JMJD2C over-
To evaluate the function of JMJD2C on MALAT1 promoter activity, A549
expression plasmids, JMJD2C overexpression plasmids + sh-MALAT1,
cells were transfected with sh-NC or sh-JMJD2C with a luciferase reporter
JMJD2C overexpression plasmids + miR-503-5p mimic, or JMJD2C plasmid containing MALAT1 promoter through Lipofectamine 3000
overexpression plasmids + sh-SEPT2. Shortly, cells were cultured in a (Invitrogen). After 48 h, cell lysates were subjected to analysis of relative
6-well plate at 4 × 105 cells/mL and transfected at 80% cell confluence. The luciferase activity on the dual luciferase reporter gene detection system
medium was replaced 6 h after transfection, and the cells were harvested (Promega, WI, USA) [10].
48 h later. Genepharma (Shanghai, China) was responsible for synthesis of The interaction between MALAT1 and miR-503-5p and the targeting
various constructs.
relationship between miR-503-5p and SEPT2 were conformed. MALAT1
wild-type (WT), MALAT1 mutant (MUT), SEPT2 WT and SEPT2 MUT
3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium potentially binding to miR-503-5p were inserted into pGL4 luciferase
bromide (MTT) assay reporter and co-transfected with miR-503-5p mimic or mimic-NC into A549
Cells after transfection were seeded into 96-well plates at 3000 cells/plate cells. After 48 h, the relative luciferase activity was measured using a dual
and cultured for 24, 48, and 72 h, respectively. Then, each well was added luciferase detection system (Promega).
with 20 µL MTT (5 mg/mL) for 4 h, and the purple formazan precipitate was
dissolved in 200 µL dimethyl sulfoxide (Sigma-Aldrich). Optical densi- Chromatin immunoprecipitation (ChIP) assay
ty490nm was measured by a spectrophotometer and cell growth curve was ChIP assay kit (Millipore, MA, USA) was utilized. The chromatin extracted
drawn [22]. from cells were immunoprecipitated with anti-JMJD2C antibody (Abcam).
A control immunoglobulin G (Merck, Germany) was applied. Primers of
Colony formation assay MALAT1 promoter: forward: 5′-GGTCAGCCTGAGACCACTTC-3′, reverse: 5′-
Cells were counted after transfection, seeded into 6-well plates at 3000 cells/ CTGTGCCTGTTCTGGGGAAT-3′ [10, 26].
plate and cultured for 9–14 days until visible colonies appeared, during
which the medium was refreshed every 48 h. Then, cells fixed in 75% RNA immunoprecipitation (RIP) assay
ethanol were subjected to staining by 0.1% crystal violet. At least 50 cells With Magna RIP™ RIP kit (Millipore), RIP analysis was carried out. Cells that
were contained in each colony and viewed under ten fields of view [23]. were lysed in complete radio-immunoprecipitation assay (protease
inhibitor cocktail and RNase inhibitor) were combined with RIP buffer
Flow cytometry containing magnetic beads coupled with human anti-Ago2 antibody
After 72 h of transfecion, cells were rinsed with PBS twice, immersed in (Millipore) or immunoglobulin G(IgG). The sample was added with
binding buffer and combined with annexin V-fluorescein isothiocyanate proteinase K and RNase inhibitors, and isolation of immunoprecipitated
(BD Biosciences, NJ, USA) and propidium iodide (BD Biosciences) in the RNA was performed. The purified RNA was analyzed by RT-qPCR [23].
darkness. Cell apoptosis was measured on a flow cytometer at 488 nm
(Agilent, Hangzhou, China) [24]. Tumor formation assay
BALB/c nude mice (5 weeks old) were employed for animal studies. A549
Transwell assay cells with stably JMJD2C-inhibition, MALAT1-inhibition, miR-503-5p-
Migration test: Cells after transfection were seeded in the upper chamber overexpression were established and injected subcutaneously into the
of the transwell containing 24-well plate in serum-free medium at 1 × 105 left side of the posterior flank of nude mouse. Tumor growth was
cells/well. The lower chamber was filled with complete medium. Cells were monitored by measuring tumor maximum length and minimum length
incubated for 48 h, and those migrated under the membrane were fixed every 7 days, and tumor volume was calculated by the formula: volume
with 4% paraformaldehyde, and stained with 1% crystal violet for 20 min. (mm3) = (width2 × length/2). On the 28th day, tumors were excised from
The cells were sealed with neutral resin and observed in five random fields euthanized mice [27].
of view.
Invasion test: Matrigel was added to the membrane of the upper Statistical analysis
chamber, the rest steps were similar to the migration test [25]. Data were analyzed with GraphPad Prism 6 (La Jolla, CA, USA). Fisher’s
exact test, Kaplan–Meier survival analysis, Pearson correlation test
Reverse transcription quantitative polymerase chain reaction t-test, and analysis of variance (ANOVA) were used to analyze data.
(RT-qPCR) P < 0.05 was considered statistically significant (*P < 0.05, **P < 0.01,
After being extracted from tissues and cells using Trizol kit (Invitrogen; ***P < 0.001).
Thermo Fisher Scientific, MA, USA), and quantified using NanoDrop ND-1000
(NanoDrop), total RNA was transformed into cDNA through Mir-XTM miRNA
First Strand Synthesis Kit (for miRNA) and Primpscript®RT reagent kit (for RESULTS
mRNA and lncRNA) (both Takara, Japan). PCR was amplified on the ABI JMJD2C-related MALAT1 is highly expressed in NSCLC tissues;
PRISM 7700 (Applied Biosystems; Thermo Fisher Scientific) using SYBR JMJD2C is related to NSCLC clinicopathological factors
Premix Ex Taq™ II PCR kit (Takara). Supplementary Table 1 showed primer The differences in levels of JMJD2C, MALAT1 in NSCLC tumor
sequences (U6 and glyceraldehyde-3-phosphate dehydrogenase [GAPDH] tissues and para-tumor tissues were assessed by RT-qPCR. As
as internal controls). The 2−ΔΔCt method was applied to quantitative
analysis. indicated by the outcome, JMJD2C and MALAT1 were highly
Cell Death and Disease (2022)13:65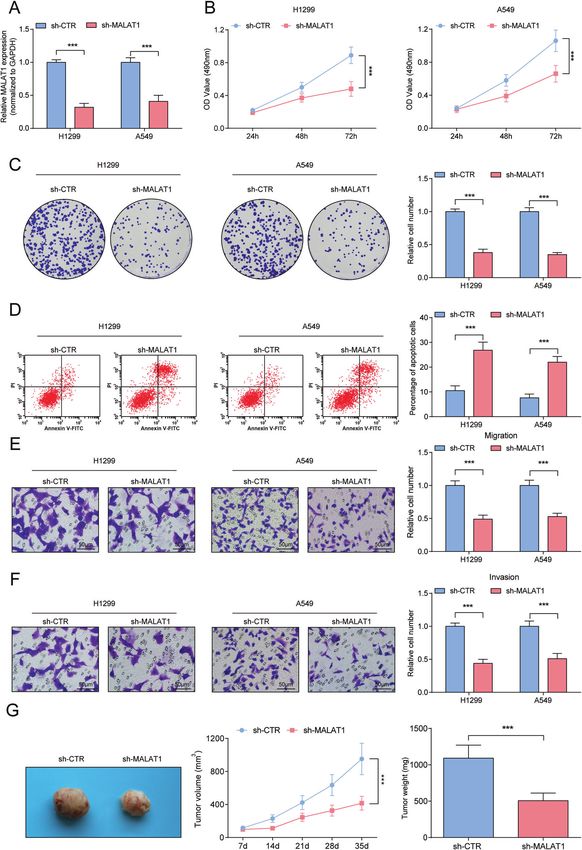
J. Zhang et al.
3
Fig. 1 JMJD2C-related MALAT1 is highly expressed in NSCLC tissues. A RT-qPCR analysis of JMJD2C in NSCLC tissue; B RT-qPCR analysis of
MALAT1 in NSCLC tissue; C Correlation between JMJD2C and MALAT1; D Relation between JMJD2C expression and survival of patients with
NSCLC. ***P < 0.001; Data statistics was by t-test, Pearson’s correlation analysis, Kaplan–Meier analysis.
expressed in NSCLC tissues compared with para-tumor tissues
(Fig. 1A, B). Subsequently analyzed by Pearson test, we found the Table 1. The relationship between JMJD2C expression and
positive correlations between levels of JMJD2C and MALAT1 in clinicopathological characteristics in patients with NSCLC.
tumor tissues (Fig. 1C). Parameter Case JMJD2C P value
For further analysis of the clinical significance of JMJD2C in
NSCLC, we divided NSCLC patients into JMJD2C high expression Low High
and JMJD2C low expression groups (Table 1) and found that expression expression
JMJD2C expression was associated with tumor size and tumor (n = 50) (n = 66)
node metastasis. Also, after Kaplan–Meier analysis, we determined Age (years) 0.254
that patients with high JMJD2C expression had shorter overall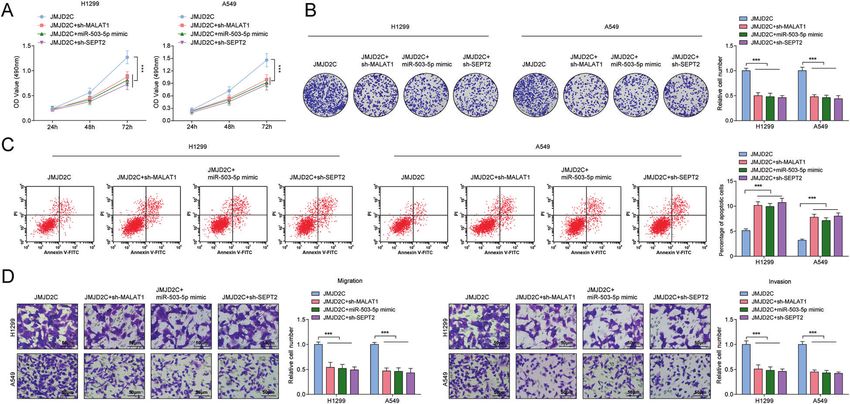
J. Zhang et al.
4
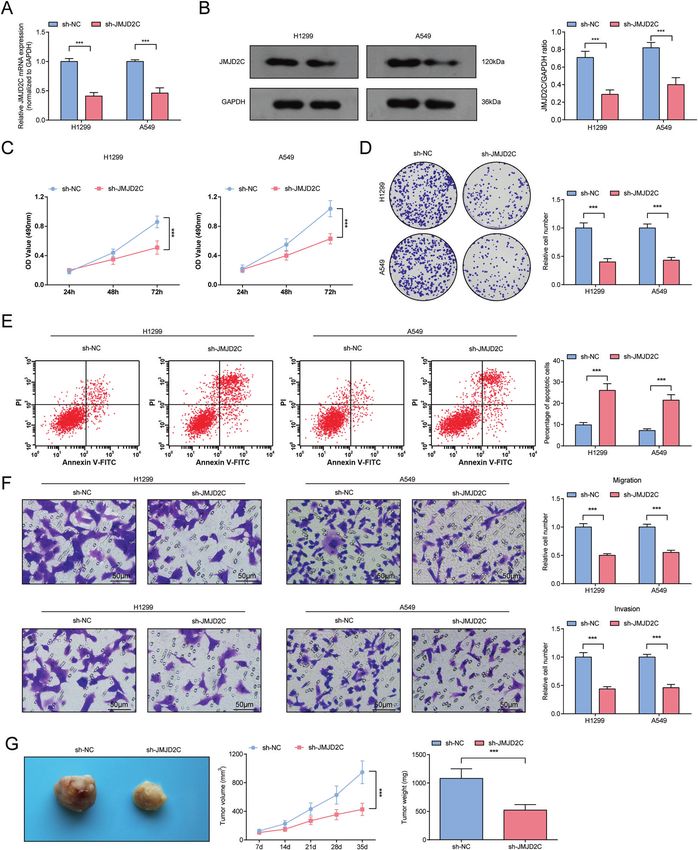
Fig. 2 Preventive effects of silenced JMJD2C on NSCLC. A, B RT-qPCR and Western blot analysis of JMJD2C level after downregulating
JMJD2C; C MTT measured the growth curve of A549 and H1299 cells after downregulating JMJD2C; D Colony formation assay measured
colony formation ability of A549 and H1299 cells after downregulating JMJD2C; E Flow cytometry measured A549 and H1299 cell apoptosis
after downregulating JMJD2C; F Transwell measured cell migration and invasion ability after downregulating JMJD2C; G Tumor formation
assay evaluated tumor volume and weight after downregulating JMJD2C. ***P < 0.001; Data statistics was by t-test.
Cell Death and Disease (2022)13:65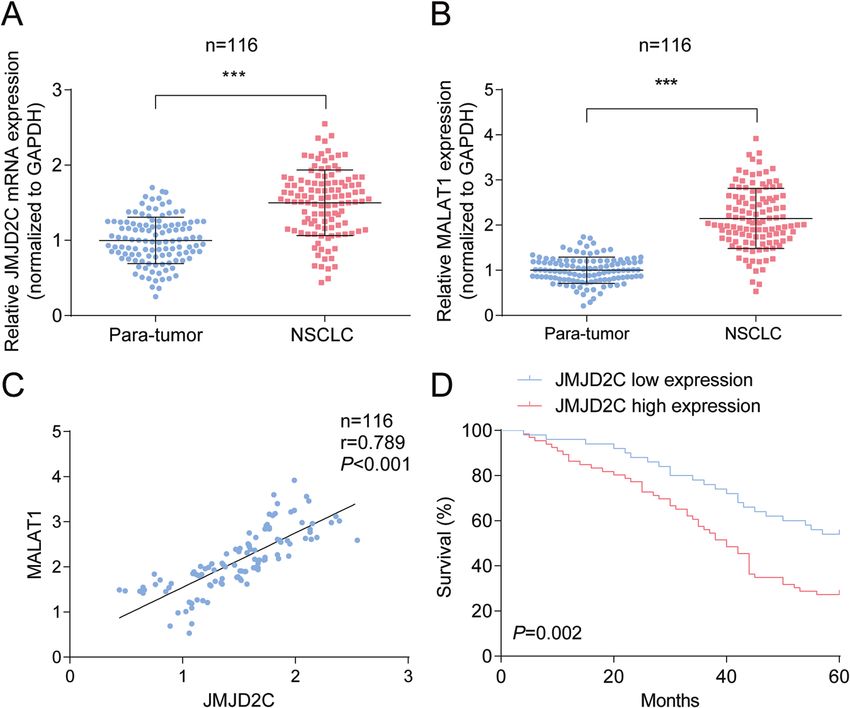
J. Zhang et al.
5
Next, the carcinogenic potential of JMJD2C in vivo was uncovered DISCUSSION
through injecting A549 cells with stable and low expression of NSCLC is caused by multi-step carcinogenesis induced by
JMJD2C. The outcomes manifested that downregulating JMJD2C chemical and physical mutagens [30]. The molecular mechanism
suppressed the growth of xenograft tumors (Fig. 2G). of NSCLC was partly explored from JMJD2C-regulated MALAT1/
Evidently, silencing JMJD2C suppressed cellular and tumor miR-503-5p/SEPT2 axis in our research. A summary was obtained
growth of NSCLC in vitro and in vivo. that JMJD2C regulated the histone methylation of MALAT1, thus
to upregulate MALAT1 level and deteriorate the malignant
Inhibitory effects of downregulated MALAT1 on NSCLC cells phenotypic of NSCLC cells via suppressing miR-503-5p and
A549 and H1299 cells were successfully transfected with sh- promoting SEPT2 (Fig. 6).
MALAT1 to knock down MALAT1 expression (Fig. 3A). Subse- From the experimental data, we found that JMJD2C was
quently, in A549 and H1299 cells with lowered MALAT1, the upregulated in NSCLC that was connected with shorter survival
cellular malignant phenotypes were destructed (Fig. 3B–F). time, and silencing JMJD2C reduced tumorigenicity of NSCLC. As a
Moreover, in the xenograft nude mouse model, sh-MALAT1 matter of fact, JMJD2C has been identified as the oncogene in
reduced the growth of xenograft tumors (Fig. 3G). pancreatic cancer, and deficiency of JMJD2C is capable to weaken
cell proliferation and invasion [31]. The aberrant increment is seen in
MALAT1 binds to miR-503-5p JMJD2C expression in osteosarcoma tissues that is suggested to
miR-503-3p inhibits lung cancer cell viability and induces cell associate with cell metastasis, and JMJD2C could drive the growth of
apoptosis [29]. Using RT-qPCR, we uncovered that miR-503-5p was osteosarcoma cells through inducing fibroblast growth factor 2 [9].
downregulated in NSCLC cancer tissues and NSCLC cell lines In the context of lung cancer, the elevated level of JMJD2C indicates
(Supplementary Fig. 2A, B). Then we found that MALAT1 an association with metastasis, and suppression of JMJD2C partly
expression was negatively correlated with miR-503-5p level in attributes to the impairments in migration and invasion, as well as
tumor tissue (Supplementary Fig. 2C). tumor hepatic metastasis and epithelial–mesenchymal transition [8].
On the Starbase website, the binding sites between MALAT1 Apart from that, the augmented level of JMJD2C has been implied
and miR-503-5p were found (Supplementary Fig. 2D). Then, in to supply the niche for colon cancer cell growth, but knockout of
dual luciferase reporter gene detection, miR-503-5p mimic could JMJD2C oppresses cell growth and colony forming ability [28], as
reduce the luciferase activity of MALAT1-WT in A549 cells well as retards lung metastasis in breast cancer [32]. Showing the
(Supplementary Fig. 2E). Also, RIP experiment tested the consistence with these reports, JMJD2C adversely performed
enriched MALAT1 and miR-503-5p levels in Ago2 immunopreci- in NSCLC.
pitation (Supplementary Fig. 2F). Besides, RT-qPCR revealed an MALAT1 has been ensured to be positively regulated by
increment in miR-503-5p level in A549 and H1299 cells after JMJD2C through modification of histone methylation, and works
downregulating MALAT1 (Supplementary Fig. 2G). Collectively, to enhance metastasis of colorectal cancer cells [10]. Consistently,
MALAT1 can specifically bind miR-503-5p, thereby regulating we discovered the mechanism that JMJD2C upregulated MALAT1
miR-503-5p level. through mediating the methylation modification of H3K9 and
H3K36 on the promoter of MALAT1. Previously proved, MALAT1
Repressive effects of miR-503-5p on tumorigenicity of NSCLC acts crucially in regulating the metastatic phenotype of lung
We transfected miR-503-5p mimic into A549 and H1299 cells, and cancer cells, and lung cancer cells deficient of MALAT1 are
verified the success of transfection by RT-qPCR (Fig. 4A). Notably, impaired in terms of their migration and metastasis abilities in
suppressed cellular progression was seen in A549 and H1299 cells xenografted tumors [12]. In NSCLC, the raised level of MALAT1 is
up-regulating miR-503-5p (Fig. 4B–F). Also, in the xenograft nude detectable in NSCLC [33], and knocking out MALAT1 weakens the
mouse model, overexpression of miR-503-5p suppressed the biological functions and tumorigenic ability of NSCLC cells
growth of xenograft tumors (Fig. 4G). through binding to miR-185-5p [25]. MALAT1 expression shows
a reduction in NSCLC clinical samples, and elevated level of
miR-503-5p negatively mediates SEPT2 MALAT1 fuels the proliferation, migration and invasion activities of
SEPT2 level measured by RT-qPCR and Western blot was NSCLC cells [34].
upregulated in NSCLC tissues and A549 and H1299 cells Our discovery revealed that MALAT1 drove the development in
(Supplementary Fig. 3A, B). We found that SEPT2 expression was NSCLC through interacting with the specific miRNA, miR-503-5p.
negatively correlated with miR-503-5p level in tumor tissue Supported by a late report, MALAT1 targets miR-503-5p, thus to
(Supplementary Fig. 3C). The targeting relationship between induce proliferation and suppress apoptosis of ovarian cancer cells
miR-503-5p and SEPT2 was predicted through the Starbase [18]. Extensive reports have discussed that miR-503-5p represses
website (Supplementary Fig. 3D). The dual luciferase reporter tumors, complying our findings that elevating miR-503-5p
gene test further examined that the luciferase activity of cells hampered NSCLC cell growth in vitro and in vivo. In colon cancer,
transfected with miR-503-5p mimic and SEPT2-WT was reduced the success in suppressing tumorigenesis and lymphangiogenesis
(Supplementary Fig. 3E). could be partly attributed to introduction of elevated miR-503-5p
The outcome of Western blot also clarified that upregulating [35]. In addition, as to hepatocellular carcinoma cells introduced
miR-503-5p in A549 and H1299 cells depressed SEPT2 level with upregulated miR-503-5p, their mobility and metastasis are
(Supplementary Fig. 3F), proving miR-503-5p targeting SEPT2. impaired [36]. Mechanistically, miR-503-5p demonstrates a
reduced level in osteosarcoma, and if suppressing miR-503-5p in
The pro-tumor effects of elevated JMJD2C on NSCLC cells are osteosarcoma cells, the tumorigenesis progression could be push
reversed by silencing MALAT1 or SEPT2, or restoring miR-503-5p forward [37]. Overall, targeted restoration of miR-503-5p could
The mechanism of JMJD2C, MALAT1, miR-503-5p, and SEPT2 in restrain the malignant progression in tumors.
NSCLC was further assessed. We recommended sh-MALAT1, SEPT2 was a target of miR-503-5p and downregulating SEPT2
miR-503-5p mimic, or sh-SEPT2 into A549 and H1299 cells that was proved to delay tumorigenesis in NSCLC. SEPT2 has been
had been transfected with JMJD2C overexpression plasmid. The implicated to aggrandize the aggressiveness of various tumors,
pro-tumor effects of elevated JMJD2C on NSCLC cells were and silencing of SEPT2 is inhibitory for tumor growth. Evidently,
reversed by silencing MALAT1 or SEPT2, or restoring miR-503-5p downregulating SEPT2 restrains the invasive capability of glio-
(Fig. 5A–D). blastoma cells [38] and the proliferative ability of epithelial ovarian
In summary, JMJD2C mediated NSCLC by regulating MALAT1/ cancer cells [39]. Similar to our findings, SEPT2 has been revealed
miR-503-5p/SEPT2 axis. to exert as a number of lncRNA-miRNA-mRNA network, thus to
Cell Death and Disease (2022)13:65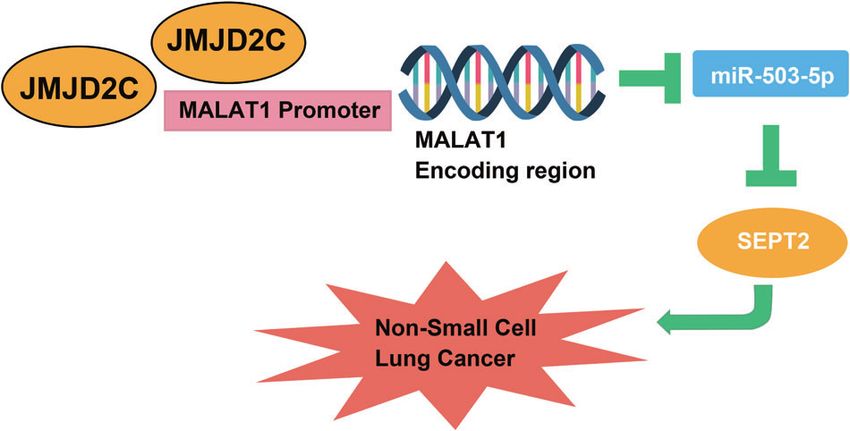
J. Zhang et al.
6
Fig. 3 Inhibitory effects of down-regulated MALAT1 on NSCLC. A RT-qPCR analysis of MALAT1 level after down-regulating MALAT1; B MTT
measured the growth curve of A549 and H1299 cells after downregulating MALAT1; C Colony formation assay measured colony formation
ability of A549 and H1299 cells after downregulating MALAT1; D Flow cytometry measured A549 and H1299 cell apoptosis after
downregulating MALAT1. E, F Transwell measured cell migration and invasion ability after downregulating MALAT1. G Tumor formation assay
evaluated tumor volume and weight after downregulating MALAT1. ***P < 0.001; Data statistics was by t-test.
Cell Death and Disease (2022)13:65J. Zhang et al.
7
Fig. 4 Repressive effects of miR-503-5p on tumorigenicity of NSCLC. A RT-qPCR analysis of miR-503-5p level after upregulating miR-503-5p.
B MTT measured the growth curve of A549 and H1299 cells after upregulating miR-503-5p. C Colony formation assay measured colony
formation ability of A549 and H1299 cells after upregulating miR-503-5p. D Flow cytometry measured A549 and H1299 cell apoptosis after
upregulating miR-503-5p. E, F Transwell measured cell migration and invasion ability after upregulating miR-503-5p. G Tumor formation assay
evaluated tumor volume and weight after upregulating miR-503-5p. ***P < 0.001; Data statistics was by t-test.
Cell Death and Disease (2022)13:65J. Zhang et al.
8
Fig. 5 The promoting effects of elevated JMJD2C on NSCLC are reversed by silencing MALAT1 or SEPT2, or restoring miR-503-5p. A MTT
measured the growth curve of A549 and H1299 cells. B Colony formation assay measured colony formation ability of A549 and H1299 cells.
C Flow cytometry measured A549 and H1299 cell apoptosis. D Transwell measured cell migration and invasion ability; Data statistics was by
t-test, ANOVA, Pearson’s correlation analysis.
3. Goebel C, Louden CL, McKenna R Jr, Onugha O, Wachtel A, Long T. Diagnosis of
non-small cell lung cancer for early stage asymptomatic patients. Cancer Genom.
Proteom. 2019;16:229–44.
4. Duma N, Santana-Davila R, Molina JR. Non-small cell lung cancer: epidemiology,
screening, diagnosis, and treatment. Mayo Clin Proc. 2019;94:1623–40.
5. Berry WL, Janknecht R. KDM4/JMJD2 histone demethylases: epigenetic regulators
in cancer cells. Cancer Res. 2013;73:2936–42.
6. Young LC, Hendzel MJ. The oncogenic potential of Jumonji D2 (JMJD2/KDM4)
histone demethylase overexpression. Biochem Cell Biol. 2013;91:369–77.
7. Duan L, Perez RE, Chastain PD 2nd, Mathew MT, Bijukumar DR, Maki CG. JMJD2
promotes acquired cisplatin resistance in non-small cell lung carcinoma cells.
Oncogene. 2019;38:5643–57.
8. Li N, Jiang D. Jumonji domain containing 2C promotes cell migration and inva-
sion through modulating CUL4A expression in lung cancer. Biomed Pharmac-
Fig. 6 The network of JMJD2C. A schematic model of JMJD2C- other. 2017;89:305–15.
mediated MALAT1/miR-503-5p/SEPT2 regulatory network in NSCLC. 9. Li X, Dong S. Histone demethylase JMJD2B and JMJD2C induce fibroblast growth
factor 2: mediated tumorigenesis of osteosarcoma. Med Oncol. 2015;32:53.
encourage growth of tumors. For instance, enhanced cellular 10. Wu X, Li R, Song Q, Zhang C, Jia R, Han Z, et al. JMJD2C promotes colorectal
proliferation in prostate cancer is triggered by LINC00473/miR- cancer metastasis via regulating histone methylation of MALAT1 promoter
and enhancing beta-catenin signaling pathway. J Exp Clin Cancer Res.
195-5p-regulated SEPT2 upregulation [40].
2019;38:435.
The present research for the first time discusses the mechanism 11. Chen J, Wang R, Zhang K, Chen LB. Long non-coding RNAs in non-small cell
of JMJD2C-mediated MALAT1/miR-503-5p/SEPT2 axis in NSCLC. To lung cancer as biomarkers and therapeutic targets. J Cell Mol Med.
conclude briefly, the investigation explained that JMJD2C drives 2014;18:2425–36.
the progression of NSCLC through modifying histone methylation 12. Gutschner T, Hammerle M, Eissmann M, Hsu J, Kim Y, Hung G, et al. The non-
of MALAT1 to upregulate miR-503-5p-targeted SEPT2, supplying a coding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung
possible chance to control NSCLC. Due to laboratory conditions, cancer cells. Cancer Res. 2013;73:1180–9.
animal models of tumor metastasis were not used to detect the 13. Wei S, Wang K, Huang X, Zhao Z, Zhao Z. LncRNA MALAT1 contributes to non-
effect of the JMJD2C/MALAT1/miR-503-5p/SEPT2 signal axis on small cell lung cancer progression via modulating miR-200a-3p/programmed
death-ligand 1 axis. Int J Immunopathol Pharm. 2019;33:2058738419859699.
NSCLC in vivo. Also, the expression and survival analysis of
14. Feng C, Zhao Y, Li Y, Zhang T, Ma Y, Liu Y. LncRNA MALAT1 promotes lung cancer
JMJD2C, MALAT1, miR-503-5p, and SEPT2 was not provided to the proliferation and gefitinib resistance by acting as a miR-200a sponge. Arch
public database due to resource constraints. Bronconeumol. 2019;55:627–33.
15. Florczuk M, Szpechcinski A, Chorostowska-Wynimko J. miRNAs as biomarkers and
therapeutic targets in non-small cell lung cancer: current perspectives. Target
DATA AVAILABILITY Oncol. 2017;12:179–200.
Not applicable. 16. Wei Y, Liao Y, Deng Y, Zu Y, Zhao B, Li F. MicroRNA-503 inhibits non-small cell
lung cancer progression by targeting PDK1/PI3K/AKT pathway. OncoTargets Ther.
2019;12:9005–16.
REFERENCES 17. Qiu T, Zhou L, Wang T, Xu J, Wang J, Chen W, et al. miR-503 regulates the
1. Herbst RS, Morgensztern D, Boshoff C. The biology and management of non- resistance of non-small cell lung cancer cells to cisplatin by targeting Bcl-2. Int J
small cell lung cancer. Nature. 2018;553:446–54. Mol Med. 2013;32:593–8.
2. Tabchi S, Kassouf E, Rassy EE, Kourie HR, Martin J, Campeau MP, et al. Manage- 18. Sun Q, Li Q, Xie F. LncRNA-MALAT1 regulates proliferation and apoptosis of ovarian
ment of stage III non-small cell lung cancer. Semin Oncol. 2017;44:163–77. cancer cells by targeting miR-503-5p. OncoTargets Ther. 2019;12:6297–307.
Cell Death and Disease (2022)13:65J. Zhang et al.
9
19. Ostevold K, Melendez AV, Lehmann F, Schmidt G, Aktories K, Schwan C. Septin 37. Li J, Zhang F, Li H, Peng F, Wang Z, Peng H, et al. Circ_0010220-mediated miR-
remodeling is essential for the formation of cell membrane protrusions (micro- 503-5p/CDCA4 axis contributes to osteosarcoma progression tumorigenesis.
tentacles) in detached tumor cells. Oncotarget. 2017;8:76686–98. Gene. 2020;763:145068.
20. Tian Q, Yan X, Yang L, Liu Z, Yuan Z, Shen Z, et al. lncRNA NORAD promotes 38. Xu D, Liu A, Wang X, Chen Y, Shen Y, Tan Z, et al. Repression of Septin9 and
hepatocellular carcinoma progression via regulating miR-144-3p/SEPT2. Am J Septin2 suppresses tumor growth of human glioblastoma cells. Cell Death Dis.
Transl Res. 2020;12:2257–66. 2018;9:514.
21. Cai, B, Wang X, Bu Q, Li P, Xue Q, Zhang J, et al. LncRNA AFAP1-AS1 knockdown 39. James NE, Cantillo E, Yano N, Chichester CO, DiSilvestro PA, Hovanesian V, et al.
represses cell proliferation, migration, and induced apoptosis in breast cancer by Septin-2 is overexpressed in epithelial ovarian cancer and mediates pro-
downregulating SEPT2 via sponging miR-497-5p. Cancer Biother Radiopharm. liferation via regulation of cellular metabolic proteins. Oncotarget. 2019;10:
2020; https://doi.org/10.1089/cbr.2020.3688. 2959–72.
22. Liu C, Tao T, Xu B, Lu K, Zhang L, Jiang L, et al. BTG1 potentiates apoptosis and 40. Xing Z, Li S, Liu Z, Zhang C, Meng M, Bai Z. The long non-coding RNA LINC00473
suppresses proliferation in renal cell carcinoma by interacting with PRMT1. Oncol contributes to cell proliferation via JAK-STAT3 signaling pathway by regulating
Lett. 2015;10:619–24. miR-195-5p/SEPT2 axis in prostate cancer. Biosci Rep. 2020;40:BSR20191850.
23. Zhang Z, Lin W, Lin Y, Kang M, Zhu J, Tong Z, et al. Long intergenic non-coding
RNA Linc00485 promotes lung cancer progression by modulating miR-298/c-Myc
axis. J Cell Mol Med. 2021;25:309–22. AUTHOR CONTRIBUTIONS
24. Wang Y, Wang X, Han L, Hu D. LncRNA MALAT1 regulates the progression and Wendong Wang contributed to manuscript editing; Jun Zhang, Mingliang Wang and
cisplatin resistance of ovarian cancer cells via modulating miR-1271-5p/E2F5 axis. Jiashun Wang contributed to manuscript data analysis; Mingliang Wang contributed
Cancer Manag Res. 2020;12:9999–10010. to experimental studies; Jiashun Wang and Wendong Wang contributed to study
25. Wang D, Zhang S, Zhao M, Chen F. LncRNA MALAT1 accelerates non-small cell design. All authors read and approved the final manuscript.
lung cancer progression via regulating miR-185-5p/MDM4 axis. Cancer Med.
2020;9:9138–49.
26. Watson AH, Burrows M. Immunocytochemical and pharmacological evidence for
GABAergic spiking local interneurons in the locust. J Neurosci. 1987;7:1741–51. COMPETING INTERESTS
27. Chen S, Wang G, Tao K, Cai K, Wu K, Ye L, et al. Long noncoding RNA metastasis- The authors declare no competing interests.
associated lung adenocarcinoma transcript 1 cooperates with enhancer of zeste
homolog 2 to promote hepatocellular carcinoma development by modulating
the microRNA-22/Snail family transcriptional repressor 1 axis. Cancer Sci. ADDITIONAL INFORMATION
2020;111:1582–95. Supplementary information The online version contains supplementary material
28. Kim TD, Fuchs JR, Schwartz E, Abdelhamid D, Etter J, Berry WL, et al. Pro-growth available at https://doi.org/10.1038/s41419-022-04513-5.
role of the JMJD2C histone demethylase in HCT-116 colon cancer cells and
identification of curcuminoids as JMJD2 inhibitors. Am J Transl Res. 2014;6:236–47. Correspondence and requests for materials should be addressed to Wendong Wang.
29. Sun Y, Li L, Xing S, Pan Y, Shi Y, Zhang L, et al. miR-503-3p induces apoptosis of
lung cancer cells by regulating p21 and CDK4 expression. Cancer Biomark. Reprints and permission information is available at http://www.nature.com/
2017;20:597–608. reprints
30. Nagano T, Tachihara M, Nishimura Y. Molecular mechanisms and targeted
therapies including immunotherapy for non-small cell lung cancer. Curr Cancer Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims
Drug Targets. 2019;19:595–630. in published maps and institutional affiliations.
31. An Y, Cai H, Zhang Y, Liu S, Duan Y, Sun D, et al. circZMYM2 competed endo-
genously with miR-335-5p to regulate JMJD2C in pancreatic cancer. Cell Physiol
Biochem. 2018;51:2224–36.
32. Luo W, Chang R, Zhong J, Pandey A, Semenza GL. Histone demethylase JMJD2C is
a coactivator for hypoxia-inducible factor 1 that is required for breast cancer
progression. Proc Natl Acad Sci USA. 2012;109:E3367–76. Open Access This article is licensed under a Creative Commons
33. Chen W, Zhao W, Chen S, Zhang L, Guo Z, Wang L, et al. Expression and corre- Attribution 4.0 International License, which permits use, sharing,
lation of MALAT1 and SOX9 in non-small cell lung cancer. Clin Respir J. adaptation, distribution and reproduction in any medium or format, as long as you give
2018;12:2284–91. appropriate credit to the original author(s) and the source, provide a link to the Creative
34. Liu C, Li H, Jia J, Ruan X, Liu Y, Zhang X. High metastasis-associated lung ade- Commons license, and indicate if changes were made. The images or other third party
nocarcinoma transcript 1 (MALAT1) expression promotes proliferation, migration, material in this article are included in the article’s Creative Commons license, unless
and invasion of non-small cell lung cancer via ERK/mitogen-activated protein indicated otherwise in a credit line to the material. If material is not included in the
kinase (MAPK) signaling pathway. Med Sci Monit. 2019;25:5143–9. article’s Creative Commons license and your intended use is not permitted by statutory
35. Wei L, Sun C, Zhang Y, Han N, Sun S. miR-503-5p inhibits colon cancer tumor- regulation or exceeds the permitted use, you will need to obtain permission directly
igenesis, angiogenesis, and lymphangiogenesis by directly downregulating from the copyright holder. To view a copy of this license, visit http://creativecommons.
VEGF-A. Gene Ther. 2020. https://doi.org/10.1038/s41434-020-0167-3. org/licenses/by/4.0/.
36. Jiang SP, Li ZR. MiR-503-5p regulates cell epithelial-to-mesenchymal transition,
metastasis and prognosis of hepatocellular carcinoma through inhibiting WEE1.
Eur Rev Med Pharm Sci. 2019;23:2028–37. © The Author(s) 2022
Cell Death and Disease (2022)13:65You can also read

















































