Intestinal immunity: to be, or not to be, induced? That is the question - Oxford ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
International Immunology, Vol. 33, No. 12, pp. 755–759 © The Japanese Society for Immunology. 2021. All rights reserved.
https://doi.org/10.1093/intimm/dxab051 For permissions, please e-mail: journals.permissions@oup.com
Advance Access publication 10 August 2021
Intestinal immunity: to be, or not to be, induced? That
is the question
Daisuke Takahashi1,*, Shunsuke Kimura1,* and Koji Hase1,2,
1
Division of Biochemistry, Faculty of Pharmacy and Graduate School of Pharmaceutical Sciences, Keio University, Minato-ku,
Tokyo 105-8512, Japan
2
International Research and Development Center for Mucosal Vaccines, The Institute of Medical Science, The University of
Tokyo (IMSUT), Minato-ku, Tokyo 108-8639, Japan
Downloaded from https://academic.oup.com/intimm/article/33/12/755/6347559 by guest on 17 December 2021
REVIEW
*These authors contributed equally to this work.
Correspondence to: K. Hase; E-mail: hase-kj@pha.keio.ac.jp
Received 22 July 2021, editorial decision 5 August 2021; accepted 9 August 2021
Abstract
The intestinal immune system maintains intestinal homeostasis in collaboration with diverse
immune cell subsets residing at the epithelial layer, lamina propria and gut-associated lymphoid
tissue (GALT). Bacterial components and their metabolites are essential for the establishment
of the gut immune system. In addition, nutritional signals contribute to maintaining the mucosal
immune response. Specialized epithelial microfold (M) cells in GALT facilitate immune surveillance
on the mucosal surface by actively taking up external antigens to transport them into the lymphoid
follicles. Because hyperplasia of M cells causes an excessive immune response in GALT, there is a
self-regulatory mechanism to control the development of M cells appropriately. In this review, we will
discuss the molecular mechanisms of mucosal immune regulation and their biological importance.
Keywords: commensal microbiota, fasting, IgA, intestinal immune system, M cell
Introduction
The intestinal tract is responsible for digestion and absorption by infectious agents, the extraction of energy from food res-
and is constantly exposed to various antigens such as food- idues and the metabolism of drugs. In addition, multiple lines
borne pathogens and commensal microbes. Therefore, the in- of evidence have proved that microbiome-derived proteins
testinal mucosa must conduct the two contradictory functions: and metabolites regulate the immune system and barriers, and
nutrient absorption and exclusion of foreign microbes. The thus the microbiome is now considered a critical factor in the
intestinal tract contains the secondary lymphoid tissue called regulation of inflammatory diseases.
gut-associated lymphoid tissue (GALT). The follicle-associated Nutritional factors are also crucial for the maintenance of
epithelium (FAE) covering GALT contains specialized epithelial the intestinal immune system. For instance, retinoic acid,
microfold (M) cells, which actively take up luminal antigens to an active metabolite of vitamin A, propels the gut homing of
facilitate antigen-specific immunoglobulin A (IgA) production lymphocytes and germinal center (GC) formation, enhancing
(1). Meanwhile, a substantial number of pathogens and toxins the IgA response (4, 5). Furthermore, food intake is essential
exploit M cells to gain entry into the body. Therefore, the fre- to retain lymphocytes in Peyer’s patches (PPs). Together, mul-
quency of M cells must be tightly regulated. tiple factors regulate the gut immune systems. In particular,
Recent studies have revealed that the gut microbiota and food antigens and nutritional factors have an enormous im-
nutritional factors greatly influence the establishment and pact on the immune system in the upper intestine, whereas
maintenance of the intestinal immune system. There are ap- commensal microbes affect it mainly in the lower intestine.
proximately 40 trillion commensal bacteria in the human colon
(2); they actively digest food residues to synthesize short-chain
M-cell-dependent antigen uptake
fatty acids (SCFAs), which are utilized by host cells as an en-
ergy source. The gut microbiota also provides B-vitamins and In 1973, Max Cooper et al. identified an atypical epithelial cell
vitamin K to the hosts. On the basis of these facts, Joshua type, which is specialized for luminal antigen uptake, in the
Lederberg proposed that the host and its parasites behave bursa of Fabricius of chickens (6). Robert Owen also iden-
like ‘superorganisms’, with their genomes linked together (3). tified a similar cell type in the FAE of human PPs (7). The
The microbiome is sometimes referred to as the ‘hidden organ’ authors named this cell type ‘M cell’ based on its morpho-
because of its pleiotropic effects: the resistance to colonization logical feature; M cells harbor irregular and sparse microfolds756 Immune regulation in the intestine
rather than regular and dense microvilli on their apical sur- animals via the gastrointestinal tract to cause brucellosis.
face. Meanwhile, the basolateral membrane of M cells is Thus, B. abortus establishes systemic infection mainly by
deeply invaginated into the cytoplasm to form M-cell pockets, utilizing PrPC on the M cells as an invasive receptor. These
to which engulfed luminal antigens are released and subse- studies at least partially uncovered the molecular machinery
quently transferred to antigen-presenting cells. Because of by which M cells take up luminal micro-organisms.
their characteristic morphology and potential importance in The mechanism of M-cell differentiation has also been de-
mucosal immunity, M cells have drawn attention; however, the fined over the past decade or so. Williams et al. identified
technical difficulty in isolating M cells because of the lack of RANKL as an inducer of M cell in PPs (14). Kanaya et al.
a molecular marker had hampered attempts to clarify the mo- reported that the TRAF6–RelB-dependent non-canonical
lecular basis of their functions and differentiation. NF-κB pathway, downstream of RANKL–RANK signaling, is
In the early 2000s, we and others extensively attempted cardinal to induce M-cell differentiation, whereas the canon-
to identify M-cell-specific molecules. We established a tech- ical NF-κB pathway also indirectly contributes to the differen-
nique to isolate FAE in PPs to perform transcriptome analysis tiation by enhancing RelB expression (15). Furthermore, we
Downloaded from https://academic.oup.com/intimm/article/33/12/755/6347559 by guest on 17 December 2021
of M-cell-enriched FAE, which enabled us to identify several identified two master transcription factors of M cells, namely
M-cell-specific molecules, including glycoprotein 2 (GP2), Spi-B and Sox8 (16, 17), both of which are transactivated
TNF-α-induced protein 2 (Tnfaip2)/M-sec and cellular prion by RelB. Newly generated immature M cells express Spi-B
protein (PrPC) (8–11). Among them, GP2, a GPI-anchored and Sox8 but lack GP2 (18) and, in turn, undergo terminal
protein, is expressed on the apical surface of M cells to serve differentiation into functionally mature Spi-B+Sox8+GP2high M
as an antigen uptake receptor. GP2 selectively binds a subset cells. Together, the RANKL–RANK-RelB–Spi-B/Sox8 axis is
of commensal and pathogenic enterobacteria, including responsible for the differentiation and functional maturation
Escherichia coli and Salmonella enterica serovar Typhimurium of M cells (Fig. 1).
(S. Typhimurium), by recognizing FimH, a component of type M-cell-dependent antigen uptake can be a double-edged
1 pili (9). GP2 deficiency attenuates T-cell responses against sword; it induces antigen-specific mucosal responses and
orally introduced recombinant S. Typhimurium expressing a serves as a vulnerable gateway for entry of pathogenic agents.
tetanus toxoid, resulting in diminished production of antigen- Therefore, it is not surprising that there is machinery to keep
specific secretory IgA. This work proved that M-cell-dependent M-cell differentiation in check. In fact, we recently defined
antigen transcytosis elicits the mucosal immune response. osteoprotegerin (OPG) as a negative regulator of M cells (19).
Interestingly, GP2 also binds botulinum toxin A to facilitate OPG is highly expressed by mature M cells and competitively
its translocation into the body via M cells (12). Depleting M inhibits the interaction between RANKL and RANK in the ad-
cells by an anti-RANKL [receptor activator of nuclear factor jacent cells, eventually suppressing the further development
κB (NF-κB) ligand] antibody or genetic deletion of GP2 pro- of M cells (Fig. 1). Notably, OPG deficiency caused expan-
tects host animals against lethal neurotoxicity after oral ad- sion of M cells in GALT and enhanced commensal bacterium-
ministration of botulinum toxin A. PrPC, another GPI-anchored specific IgA and IgG production. Accordingly, the ablation of
protein on the apical surface of M cells, binds to Hsp60 on OPG ameliorated disease symptoms in mice with experimental
Brucella abortus (13). This zoonotic bacterium infects host colitis. However, OPG-deficient mice were highly susceptible
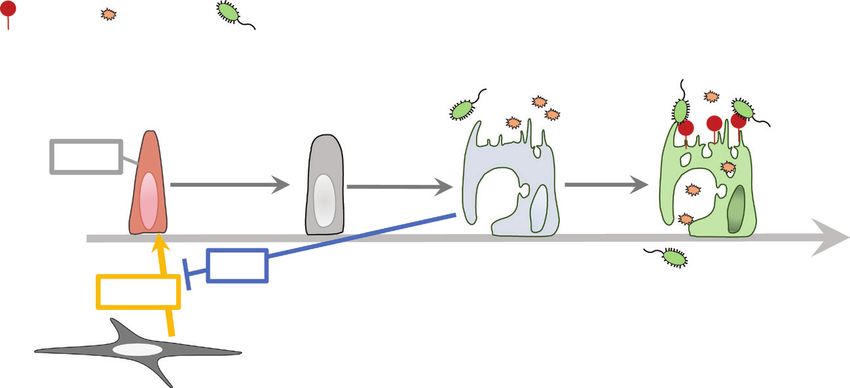
Fig. 1. Schematic diagram of differentiation of M cells in PPs. RANKL expressed on M-cell inducer cells initiates the M-cell differentiation by
activating canonical NF-κB (p50/RelA) through TRAF6. Canonical NF-κB signaling is required to activate non-canonical NF-κB (p52/RelB),
which induces the transcription factors, Spi-B and Sox8 in parallel. Spi-B and Sox8 are master transcription factors that direct progenitor cells to
M cells. Spi-B+Sox8+GP2low/− cells are immature M cells with insufficient uptake activity. Sox8 directly induces the expression of GP2 to facilitate
the development of mature M cells with high uptake activity. Other markers of immature/mature M cells are shown.Immune regulation in the intestine 757
to oral infection with S. Typhimurium, with enhanced systemic Regulation by microbial metabolites
translocation of the bacteria via M cells. Thus, the self-regula-
The commensal microbiota produces a myriad of metab-
tion of M-cell numbers by OPG is critical to maintaining the
olites as the products of united host and micro-organism
equilibrium between the risk of infection and the ability to per-
interactions, although most of these are assumed to be un-
form immunosurveillance at the mucosal surface.
identified, and a large proportion of known metabolites have
yet to be defined functionally. Commensal microbiota-derived
The requirement for food intake metabolites regulate the development and function of the in-
Recent studies have illustrated a significant link between testinal immune system, albeit to a lesser extent than bac-
immune responses and metabolism. In epidemiological terial structural components.
studies, low nutritional status attenuates the efficacy of vac- Thus, our understanding of the interplay between micro-
cination and thus increases the prevalence of infectious bial metabolites and host physiology may still be at a pre-
diseases. Signaling via mTOR and AMPK, the principal nu- mature state. Nonetheless, by any metric, the research field
tritional sensors, regulates metabolic reprogramming of im- regarding the interplay between microbial metabolites and
Downloaded from https://academic.oup.com/intimm/article/33/12/755/6347559 by guest on 17 December 2021
mune cells. Consequently, the nutritional status markedly the immune system has enormously exploded in the past
affects the differentiation and functions of immune cells. decade. Not all the findings can be covered in this section,
We found that 36-h fasting induces apoptosis in PP GC B but we will highlight the current state of knowledge, mainly
cells because of decreased mTOR signaling (20). Hence, focusing on our discoveries about SCFAs and polyamines.
the production of antigen-specific IgA was severely affected It is noteworthy that SCFAs support the regulatory arm of the
even after refeeding. Notably, fasting also influenced the cel- immune system while enhancing the effector arm fighting
lular dynamics in PPs by down-regulating the production of against infection without causing excessive inflammatory re-
chemokines such as CXCL13 by stromal cells. sponses. On another front, it is well recognized that SCFAs
Metabolic reprogramming similar to the Warburg effect is and polyamines facilitate healthy host energy metabolism.
required for CXCL13 production. Therefore, the arrest of the Comparatively speaking, little is known about the contribu-
glycolysis-dependent metabolism during fasting led to the tion of commensal bacteria-derived polyamines in the host
down-regulation of CXCL13 expression in PPs. In contrast, immune system.
CXCL13 expression in the bone marrow (BM) increased only SCFAs, namely acetate, propionate and butyrate, are mi-
during fasting by an unknown mechanism, resulting in a tran- crobial anaerobic fermentation products of indigestible food
sient accumulation of naive B cells in the BM. Upon refeeding, components, such as resistant starch and soluble dietary
the naive B cells migrated from the BM to PPs, indicating that fibers. Most acetate and propionate producers belong to
the BM serves as a temporary refuge for naive B cells. the phylum Bacteroidetes, whereas butyrate producers are
In addition to naive B cells, the dynamics of CD8+ memory members of the phylum Firmicutes, particularly Clostridium
T (Tmem) cells and monocytes are also altered under a low clusters IV and XIVa (25, 26). In a pregnant dam, maternal
energy status. CXCL12–CXCR4 signaling and sphingosine microbiota-derived SCFAs influence the immune system of
1-phosphate (S1P)–S1P receptor (S1PR1) signaling are es- the offspring. Moreover, we found that SCFAs are sensed by
sential for T-cell migration. These pathways are enhanced in G protein-coupled receptor 41 (GPR41) and GPR43 of the
mice subjected to 50% calorie restriction (CR) (21). Moreover, embryo, influencing prenatal development of the metabolic
the concentration of glucocorticoids, which cause T-cell and neural systems, which is critical to prevent metabolic
apoptosis, is lower in the BM than in the peripheral tissue syndrome in adulthood (Fig. 2) (27). Among SCFAs, butyrate
during CR, whereas long-chain fatty acids essential for the has been considered the most immunologically effective me-
maintenance of Tmem cells are abundant in the BM in as- tabolite. The immunomodulatory effect of butyrate is high-
sociation with an increase in adipocytes. Such microenviron- lighted by the induction of regulatory T (Treg) and follicular
ments in the BM during CR offer a niche for CD8+ Tmem cells. Treg (Tfr) cells (28, 29). This effect is mainly attributed to the
Monocytes also migrate from the bloodstream to the BM inhibition of histone lysine deacetylases by butyrate. Butyrate
during short-term (4–20 h) fasting (22). Fasting suppresses promotes histone acetylation at the Foxp3 promoter and en-
the CCL2–CCR2 axis, a prerequisite for the egress of mono- hancer regions in Treg cells and the Bcl6 promoter region in
cytes from the BM into the bloodstream by activating AMPK/ Tfr cells (Fig. 2).
PPARα signaling in hepatocytes. Additionally, butyrate enhances the bactericidal activity of
As mentioned above, GC B cells and IgA+ B cells in PPs intestinal epithelial cells and macrophages, which decreases
were eliminated and replaced by naive B cells from the BM the dissemination of luminal bacteria to the systemic tissue. It
throughout fasting–refeeding. Considering that a subset of is also worth noting that butyrate accelerates class-switch re-
GC B cells differentiates into memory B cells, the elimination combination to IgA in a T-cell-independent fashion, increasing
of GC B cells may lead to the loss of immune memory for the concentration of colon luminal secretory IgA (30). Of note,
oral antigens. Indeed, fasted mice failed to generate antigen- butyrate is generated by fermentation of dietary fibers and
specific IgA, IgM and IgG upon repeated oral immunization resistant starches, and mucin O-glycans as an endogenous
with ovalbumin. Additionally, the supply of food antigens is substrate. The utilization of mucin O-glycans is compromised
vital for the homeostasis of PP lymphocytes (23, 24). Thus, in the microbial community of patients with ulcerative colitis
food intake that provides antigens and nutrients critically (UC) (31). Although oral administration of butyrate has been
contributes to lymphocyte accumulation and immunological applied as a treatment in animal models of colitis, the thera-
memory formation in PPs. peutic effect in human UC of rectal enema or oral application758 Immune regulation in the intestine
Downloaded from https://academic.oup.com/intimm/article/33/12/755/6347559 by guest on 17 December 2021
Fig. 2. Schematic diagram of immune regulation by microbial metabolites. Commensal bacteria produce SCFAs via the microbial fermenta-
tion of indigestible carbohydrates and mucin O-glycans in the large intestine. SCFAs produced by maternal commensal bacteria determine
the fate of metabolic and neural system differentiation through embryonic GPR41 and GPR43 during pregnancy. Butyrate has the most potent
activity to promote the differentiation of peripheral Treg (pTreg) cells and Tfr cells through the inhibition of class I histone lysine deacetylases.
Butyrate enhances the accumulation of histone acetylation (H3Kac) at the Foxp3 promoter/enhancer and Bcl6 promoter regions. Butyrate also
upregulates the production of transforming growth factor β1 (TGF-β) and all-trans RA (atRA) by CD103+CD11b+ dendritic cells (DCs) through
GPR41 and GPR109a, and the inhibition of class I HDACs. Polyamines, specifically putrescine, derived from commensal bacteria, are utilized
by the colon epithelial cells (CECs) and increase intracellular spermidine and spermine levels, enhancing the proliferation of CECs. Putrescine
also promotes the development of anti-inflammatory macrophages in the colon. Endogenous polyamine synthesis is required for the GATA-
binding protein 3 (GATA3) expression during Th2 cell differentiation.
is controversial, probably due to the difficulty of controlling of CX3CR1+ intestinal macrophages with anti-inflammatory
the concentration in the site of inflammation. Unlike previous properties (36). Bacterial putrescine is absorbed and metab-
methods, polyvinyl butyrate nanoparticles have been pro- olized to spermidine, a bioactive polyamine, by the host, thus
posed as a promising strategy to deliver butyrate to the in- contributing to the maintenance of the mucosal barrier and
flammation site in the large intestine (32). immune homeostasis in the intestine.
Polyamines (i.e. putrescine, spermidine and spermine) are
aliphatic compounds containing more than two amino groups,
Conclusion and perspectives
which interact with negatively charged biomolecules such as
DNA and RNA. Polyamine metabolism is closely linked to The Ancient Greek physician Hippocrates claimed that ‘all
many aspects of the immune system. Notably, recent studies disease begins in the gut’. Indeed, altered commensal micro-
revealed that ornithine decarboxylase (ODC), the rate-limiting biota, termed dysbiosis, has been associated with multiple
enzyme of polyamine biosynthesis, is indispensable for CD4+ diseases, including chronic inflammatory diseases, metabolic
T-cell lineage fidelity. In particular, a large portion of ODC- disorders, psychiatric disorders, allergy and autoimmune dis-
deficient CD4+ T cells produces IFN-γ even under T helper eases. The food–commensal microbiota axis plays a critical
2 (Th2)-skewing, Th17-skewing or Treg-skewing conditions, role not only in the regulation of immune response but also in
whereas IL-17 production is suppressed under Th17-skewing the establishment and maintenance of epithelial barrier func-
conditions (33, 34). Similarly, we observed that the inhibition tions (37). Therefore, dysbiosis often causes epithelial barrier
of ODC compromises the fidelity of the Th2 subset, con- dysfunction, leading to leaky gut syndrome (LGS). LGS trig-
verting it into Th9-like cells (35). gers inflammatory responses in the local and systemic tissue,
Meanwhile, exogenous polyamine supplementation by which exacerbates epithelial barrier dysfunction by affecting
commensal bacteria has an unignorable role in the gut im- the integrity of tight junctions. This vicious cycle may be impli-
mune system. We have provided evidence that commensal cated in the pathogenesis of dysbiosis-associated diseases.
bacteria-derived putrescine is utilized for epithelial prolifer- Moreover, accumulating studies have revealed that par-
ation. The exogenous putrescine also promotes differentiation ticular bacterial species serve as predisposing factors forImmune regulation in the intestine 759
several autoimmune diseases. M-cell-dependent transcytosis 15 Kanaya, T., Sakakibara, S., Jinnohara, T. et al. 2018. Development
of these disease-causing bacteria followed by activation of of intestinal M cells and follicle-associated epithelium is regulated
by TRAF6-mediated NF-κB signaling. J. Exp. Med. 215:501.
mucosal immune responses in PPs seems to facilitate the de- 16 Kanaya, T., Hase, K., Takahashi, D. et al. 2012. The Ets transcrip-
velopment of systemic autoimmune responses. Collectively, tion factor Spi-B is essential for the differentiation of intestinal
it is reasonable to consider that the intestinal microbiota microfold cells. Nat. Immunol. 13:29.
and gut immune system could be promising therapeutic tar- 17 Kimura, S., Kobayashi, N., Nakamura, Y. et al. 2019. Sox8 is es-
gets. Further investigations on the host–microbe interplay on sential for M cell maturation to accelerate IgA response at the
early stage after weaning in mice. J. Exp. Med. 216:831.
mucosal surfaces should provide insight into pathological 18 Kimura, S., Yamakami-Kimura, M., Obata, Y. et al. 2015.
mechanisms of dysbiosis-associated diseases and the de- Visualization of the entire differentiation process of murine M
velopment of new modalities. cells: suppression of their maturation in cecal patches. Mucosal
Immunol. 8:650.
19 Kimura, S., Nakamura, Y., Kobayashi, N. et al. 2020.
Funding Osteoprotegerin-dependent M cell self-regulation balances gut
infection and immunity. Nat. Commun. 11:234.
Downloaded from https://academic.oup.com/intimm/article/33/12/755/6347559 by guest on 17 December 2021
This work was supported by the Japan Society for the Promotion 20 Nagai, M., Noguchi, R., Takahashi, D. et al. 2019. Fasting-
of Science (20H05876 and 20H00509 to K.H.) and AMED-Crest refeeding impacts immune cell dynamics and mucosal immune
(21gm1310009h0002 to K.H.). responses. Cell 178:1072.
21 Collins, N., Han, S. J., Enamorado, M. et al. 2019. The bone
marrow protects and optimizes immunological memory during
Acknowledgements dietary restriction. Cell 178:1088.
We would like to thank Ian Newman for editing the manuscript. 22 Jordan, S., Tung, N., Casanova-Acebes, M. et al. 2019. Dietary
intake regulates the circulating inflammatory monocyte pool. Cell
Conflicts of interest statement: the authors declared no conflicts of 178:1102.
interest. 23 Kim, K. S., Hong, S. W., Han, D. et al. 2016. Dietary antigens limit
mucosal immunity by inducing regulatory T cells in the small in-
testine. Science 351:858.
References 24 Hara, S., Sasaki, T., Satoh-Takayama, N. et al. 2019. Dietary
1 Nakamura, Y., Kimura, S. and Hase, K. 2018. M cell-dependent antigens induce germinal center responses in Peyer’s Patches
antigen uptake on follicle-associated epithelium for mucosal im- and antigen-specific IgA Production. Front. Immunol. 10:2432.
mune surveillance. Inflamm. Regen. 38:15. 25 Louis, P. and Flint, H. J. 2009. Diversity, metabolism and microbial
2 Sender, R., Fuchs, S. and Milo, R. 2016. Revised estimates for ecology of butyrate-producing bacteria from the human large in-
the number of human and bacteria cells in the body. PLoS Biol. testine. FEMS Microbiol. Lett. 294:1.
14:e1002533. 26 Reichardt, N., Duncan, S. H., Young, P. et al. 2014. Phylogenetic
3 Lederberg, J. 2000. Infectious history. Science 288:287. distribution of three pathways for propionate production within the
4 Iwata, M., Hirakiyama, A., Eshima, Y. et al. 2004. Retinoic acid human gut microbiota. ISME J. 8:1323.
imprints gut-homing specificity on T cells. Immunity 21:527. 27 Kimura, I., Miyamoto, J., Ohue-Kitano, R. et al. 2020. Maternal gut
5 Nikawa, T., Odahara, K., Koizumi, H. et al. 1999. Vitamin A pre- microbiota in pregnancy influences offspring metabolic pheno-
vents the decline in immunoglobulin A and Th2 cytokine levels type in mice. Science 367:eaaw8429.
in small intestinal mucosa of protein-malnourished mice. J. Nutr. 28 Takahashi, D., Hoshina, N., Kabumoto, Y. et al. 2020. Microbiota-
129:934. derived butyrate limits the autoimmune response by promoting
6 Bockman, D. E. and Cooper, M. D. 1973. Pinocytosis by epithe- the differentiation of follicular regulatory T cells. EBioMedicine
lium associated with lymphoid follicles in the bursa of Fabricius, 58:102913.
appendix, and Peyer’s patches. An electron microscopic study. 29 Furusawa, Y., Obata, Y., Fukuda, S. et al. 2013. Commensal
Am. J. Anat. 136:455. microbe-derived butyrate induces the differentiation of colonic
7 Owen, R. L. and Jones, A. L. 1974. Epithelial cell specialization regulatory T cells. Nature 504:446.
within human Peyer’s patches: an ultrastructural study of intes- 30 Isobe, J., Maeda, S., Obata, Y. et al. 2020. Commensal-bacteria-
tinal lymphoid follicles. Gastroenterology 66:189. derived butyrate promotes the T-cell-independent IgA response
8 Hase, K., Ohshima, S., Kawano, K. et al. 2005. Distinct gene in the colon. Int. Immunol. 32:243.
expression profiles characterize cellular phenotypes of follicle- 31 Yamada, T., Hino, S., Iijima, H. et al. 2019. Mucin O-glycans fa-
associated epithelium and M cells. DNA Res. 12:127. cilitate symbiosynthesis to maintain gut immune homeostasis.
9 Hase, K., Kawano, K., Nochi, T. et al. 2009. Uptake through glyco- EBioMedicine 48:513.
protein 2 of FimH(+) bacteria by M cells initiates mucosal immune 32 Mu, Y., Kinashi, Y., Li, J. et al. 2021. Polyvinyl butyrate
response. Nature 462:226. nanoparticles as butyrate donors for colitis treatment. ACS Appl.
10 Hase, K., Kimura, S., Takatsu, H. et al. 2009. M-Sec promotes Bio Mater. 4:2335.
membrane nanotube formation by interacting with Ral and the 33 Puleston, D. J., Baixauli, F., Sanin, D. E. et al. 2021. Polyamine
exocyst complex. Nat. Cell Biol. 11:1427. metabolism is a central determinant of helper Tcell lineage fidelity.
11 Nakato, G., Fukuda, S., Hase, K. et al. 2009. New approach for Cell 184:4186. doi:10.1016/j.cell.2021.06.007
m-cell-specific molecules screening by comprehensive tran- 34 Wagner, A., Wang, C., Fessler, J. et al. 2021. Metabolic modeling
scriptome analysis. DNA Res. 16:227. of single Th17 cells reveals regulators of autoimmunity. Cell
12 Matsumura, T., Sugawara, Y., Yutani, M. et al. 2015. Botulinum 184:4168. doi:10.1016/j.cell.2021.05.045
toxin A complex exploits intestinal M cells to enter the host and 35 Nakamura, A., Takahashi, D., Nakamura, Y. et al. 2020. Polyamines
exert neurotoxicity. Nat. Commun. 6:6255. polarized Th2/Th9 cell-fate decision by regulating GATA3 expres-
13 Nakato, G., Hase, K., Suzuki, M. et al. 2012. Cutting Edge: sion. Arch. Biochem. Biophys. 693:108587.
Brucella abortus exploits a cellular prion protein on intestinal M 36 Nakamura, A., Kurihara, S., Takahashi, D. et al. 2021. Symbiotic
cells as an invasive receptor. J. Immunol. 189:1540. polyamine metabolism regulates epithelial proliferation and
14 Knoop, K. A., Kumar, N., Butler, B. R. et al. 2009. RANKL is macrophage differentiation in the colon. Nat. Commun. 12:1.
necessary and sufficient to initiate development of antigen- 37 Kinashi, Y. and Hase, K. 2021. Partners in leaky gut syndrome:
sampling M cells in the intestinal epithelium. J. Immunol. intestinal dysbiosis and autoimmunity. Front. Immunol. 12. Article
183:5738. no.: 673708. doi:10.3389/fimmu.2021.673708You can also read

















































