Intra- and Interspecific Prey Theft in Cicada Killers (Hymenoptera: Apoidea: Sphecius) - Semantic Scholar
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Journal of Insect Science, (2019) 19(1): 13; 1–7
doi: 10.1093/jisesa/iez004
Research
Intra- and Interspecific Prey Theft in Cicada Killers
(Hymenoptera: Apoidea: Sphecius)
J. R. Coelho,1,4 C. W. Holliday,2 and J. M. Hastings3
1
Institute for Franciscan Environmental Studies, Biology Program, Quincy University, Quincy, IL 62301, 2Department of Biology,
Lafayette College, Easton, PA 18042, 3Department of Biological Sciences, Northern Kentucky University, Highland Heights, KY 41099,
and 4Corresponding author, e-mail: coelhjo@quincy.edu
Subject Editor: Phyllis Weintraub
Received 27 November, 2018; Editorial decision 5 January, 2019
Abstract
We studied prey theft in two cicada killer aggregations: Ruby, Arizona (Sphecius convallis Patton) and Easton,
Pennsylvania (Sphecius speciosus Drury). Many prey (Tibicen parallelus Davis [Hemiptera: Cicadidae]) were
stolen from S. convallis by kingbirds and Greater Roadrunners at Ruby. Seventy percent of kingbird attacks on
provisioning wasps were successful. Using sand-filled trap nests baited with a cicada, we tested the hypothesis that
conspecific females might kleptoparasitize by laying an egg on the cicada and closing the nest cell. At Ruby, 45%
were so appropriated, and at Easton, 52%. Easton data showed that the longer a nest cell was left open, the higher
the rate of kleptoparasitism. Hence, intraspecific kleptoparasitism likely occurs at high rates in both populations.
Not needing to dig a burrow, or to hunt, capture, and carry a paralyzed cicada favors intraspecific kleptoparasitism
in cicada killers. Low cicada availability and intense avian kleptoparasitism of cicada killers may intensify selection
pressure for this behavior at the Arizona site. Pirating cicadas may be the only viable reproductive outlet for females
that are small or in environments with few prey. We suggest that provisioned nest cell kleptoparasitism may have
evolved in cicada killers as an alternative strategy to standard provisioning, given the dual uncertainties of adult
body size and prey availability.
Estudiamos el robo de presas en dos agregaciones asesinas de cigarra: Ruby, Arizona (Sphecius convallis Patton) y
Easton, Pennsylvania (S. speciosus Drury). Muchas presas (Tibicen parallelus Davis [Hemiptera: Cicadidae]) fueron
robadas de S. convallis por los tiranos y los correcaminos grandes en Ruby. El setenta por ciento de los ataques de
las aves kingbird a las avispas de aprovisionamiento tuvieron éxito. Usando nidos de trampas rellenos de arena
cebados con una cigarra, probamos la hipótesis de que las hembras conespecíficas podrían cleptoparasitar poniendo
un huevo en la cigarra y cerrando la célula nido. En Ruby, el 45% era tan apropiado, y en Easton, el 52%. Los datos de
Easton mostraron que cuanto más tiempo se dejaba abierta una célula nido, mayor era la tasa de cleptoparasitismo.
Por lo tanto, el cleptoparasitismo intraespecífico probablemente ocurra a tasas altas en ambas poblaciones. No
tener que cavar una madriguera, o cazar, capturar y transportar una cigarra paralizada favorece el cleptoparasitismo
intraespecífico en los asesinos de la cigarra. La baja disponibilidad de cigarras y el intenso cleptoparasitismo aviar
de los asesinos de cigarras pueden intensificar la presión de selección para este comportamiento en el sitio de
Arizona. Las cigarras piratas pueden ser la única salida reproductiva viable para las hembras que son pequeñas o
en ambientes con pocas presas. Sugerimos que el cleptoparasitismo aprovisionado de células nido puede haber
evolucionado en los asesinos de cigarra como una estrategia alternativa al aprovisionamiento estándar, dadas las
incertidumbres duales del tamaño del cuerpo adulto y la disponibilidad de presas.
Key words: foraging, alternative strategy, kleptoparasitism, solitary wasp, ecological interactions
Solitary, predaceous wasps paralyze their prey, providing a ready to have effects on wasp population dynamics, reproductive strate-
opportunity for other organisms to steal this resource. Indeed, gies, and evolved behavior.
parasitization by satellite flies and mutillids are well documented Members of the genus Sphex are known to have their large
(Dambach and Good 1943, Curran 1962, Lin 1963, Evans 1966, orthopteran prey stolen by House Sparrows and Grey Catbirds
Schmidt 2013). However, prey theft from wasps by conspecifics and (Brockmann 1980, Benttinen and Preisser 2009, Lechner 2010).
vertebrates has been described in only a few cases. Prey theft is likely Acorn Woodpeckers and whiptail lizards (Cnemidophorus spp.)
© The Author(s) 2019. Published by Oxford University Press on behalf of Entomological Society of America.
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-NoDerivs licence (http://creativecommons.
1
org/licenses/by-nc-nd/4.0/), which permits non-commercial reproduction and distribution of the work, in any medium, provided the original work is not
altered or transformed in any way, and that the work is properly cited. For commercial re-use, please contact journals.permissions@oup.com
2 Journal of Insect Science, 2019, Vol. 19, No. 1
have also been observed stealing cicadas from provisioning western the course of trying to find or access her burrow. There they are
cicada killers (Sphecius grandis Say; J. M. H., unpublished observa- left to rot, frequently consumed by ants or yellowjackets (Vespula
tions). The authors have observed various birds (including House spp.) (J. R. C., C. W. H., and J. M. H., unpublished observations).
Sparrow, J. R. C. and C. W. H.) attempting to steal cicadas from However, at sites with extremely high nest density (e.g., Will County,
Sphecius speciosus (Drury 1773) in this way, but never in substantial IL; Hastings et al. 2008), females may pounce on others carrying
numbers. Correspondents contributing to a cicada killer biogeog- cicadas, and dropped cicadas are immediately picked up by the pro-
raphy study (Coelho et al. 2011) noted House Sparrows, Brewer’s visioner or other female cicada killers (Lin 1963). These anecdotal
Blackbirds, Northern Cardinals, and American Crows taking prey observations suggested to us that, under the right conditions, compe-
cicadas (C. W. H., unpublished correspondence). Say’s Phoebe has tition for prey may become intense among cicada killers.
also been observed stealing prey from cicada killers (BBC 1993). In the course of many summers of field research on Sphecius, we
Cicada killers are particularly likely to be targets of vertebrate prey have often noticed female cicada killers entering the burrows of other
theft, as North American cicadas are large prey items (0.15–3.50 g females. Such ‘intruders’ spend from several seconds to an hour or
live mass) and generally contain no defensive chemicals. more in the burrow they are visiting and are sometimes vigorously
In the course of investigations on the ecology of Pacific cicada kil- evicted by the ‘owner’ female inside the burrow or when she returns
lers (Sphecius convallis Patton 1879) in Ruby, Arizona, we observed from foraging for food or cicada prey. Similar behavior is described
various vertebrate predators systematically robbing cicada killers of by various other authors (Dambach and Good 1943, Lin 1963, Evans
their prey. One goal of this study was to quantify this activity and 1966, Lin and Michener 1972, Brockman and Dawkins 1979, Pfennig
attempt to estimate its impact on the cicada killer population. and Reeve 1989) . We have often wondered whether or not such visits
Cicada killers in the genus Sphecius (Dahlbom 1843) are large might result in the in situ piracy by an intruder of an open nest cell
apoid wasps (Sann et al. 2018), which provision their young with containing one or more paralyzed cicadas put in place by the bur-
one to as many as nine paralyzed ‘annual’ cicadas. Cicada killers row’s owner; demonstrating this behavior was one of the main pur-
have a typical, solitary wasp life cycle that has been described nu- poses of our study. We designate this form of conspecific prey piracy
merous times (e.g., Dambach and Good 1943). Females emerge in ‘provisioned nest cell kleptoparasitism’ (PNCK) by cicada killer
early summer, mate, dig a burrow, and begin provisioning nest cells. females. A second purpose of this study was to assess the presence
Females hunt cicadas in the neighboring trees and sting them, re- and frequency of PNCK in two cicada killer populations in the United
sulting in complete, irreversible paralysis. The cicada is flown back States: the Pacific cicada killer, Sphecius convallis population at Ruby,
to the burrow, although overloaded wasps may be required to climb Arizona, and the Eastern cicada killer, Sphecius speciosus, population
trees or other objects to gain altitude and resume flying. The cicada on the Lafayette College campus in Easton, Pennsylvania.
is sequestered in a cell in the burrow. The flight of the female wasp
to the burrow while carrying a cicada provides the opportunity for
Materials and Methods
prey theft by birds.
High-value prey also provide opportunities for theft by conspe- Field sites
cifics. Evans (1957) reported prey-stealing in five species of Bembix. Ruby, Arizona
Villalobos and Shelly (1996) state that in the related wasp, Stictia The two cicada killer populations studied had a minimum of 50 or
heros (Fabr.), intraspecific prey theft is common. These observations more nesting female wasps. A large population of Pacific cicada killers
suggest that prey theft might occur among cicada killers as well. In nesting on a 3.3-ha field of mine tailings in Ruby (Santa Cruz County),
most of the United States, one or two cicadas in the genus Tibicen are Arizona (31°27′33.18″ N, 111°14′02.77″ W, elevation 1268 m), here-
used as nest cell provisions in nearly all Sphecius populations that after referred to as Ruby, was the subject of a number of ecological
have been investigated (Holliday et al. 2009, Hastings et al. 2010); studies in August to September 2009. Approximately 3.25 ha of fine,
Florida Sphecius spheciosus frequently use the tiny Neocicada hier- sandy tailings from an abandoned lead mine form a nearly level sand
oglyphica and compensate for the cicada’s small body mass by pro- flat blocking two south-draining valleys, each of which has a small
visioning nest cells with up to nine cicadas. Male cicada killers are lake formed upstream of the tailings. These tailings are 2–5 m above
much smaller than females and male eggs are provisioned with one the water levels of the lakes and provide a suitable site for the nest
or more cicadas; female eggs are provisioned with two or more cica- burrows of a breeding aggregation of several thousand Pacific cicada
das (Dow 1942). Female wasps know the sex of each egg before they killers. In 2009, this aggregation provisioned its offspring exclusively
lay it, as they dig larger underground nest cells for female eggs than with Tibicen parallelus (Davis 1923 Hemiptera: Cicadidae) cicadas,
for male eggs (Holliday, unpublished data from trap nest studies). which had an average body mass of 1.127 g; female S. convallis in this
They provision female eggs in the larger nest cells with more cicada population had an average body mass of 0.991 g (Coelho et al. 2012).
prey than male eggs, usually one Tibicen for a male egg and two for During the course of our 2-mo stay at Ruby, we made field notes of all
a female (Holliday et al. 2009). When more than one cicada is to cases of attacks on provisioning cicada killers. A focal area of the aggre-
be placed in a nest cell, the female leaves the burrow open with the gation area was chosen to observe prey theft by kingbirds (Tyrannus
first cicada within while she is away hunting for additional cicadas. spp.). This site had high burrow density (418 burrows along a 60-m
This behavior is in sharp contrast to that of the many progressively transect) and was directly adjacent to vegetation that kingbirds used
provisioning, solitary sand wasp species which close and even cam- heavily as perch sites. Observations were carried out primarily in the
ouflage the burrow opening while they are away hunting for prey morning hours when provisioning and prey theft were most frequent.
with which to feed their growing grubs (Evans and O’Neill 2007). Cases of kingbird attack on cicada killer/cicada pairs were recorded,
Such burrow closure presumably protects the developing wasp grub as well as whether the attack was successful. If the attack was success-
and reduces prey theft by conspecifics and by other arthropods, as ful, the behavior of the bird while handling the cicada was recorded.
well as reducing kleptoparasitism by egg-laying mutillid wasps and Successful provisioning events by cicada killers were also recorded.
by several families of flies. Most kingbirds were photographed for confirmation of identity with
At most cicada killer nesting aggregation sites, where nest density a Canon EOS 40D digital SLR camera and Sigma 150–500 mm lens.
is relatively low, cicadas are occasionally dropped by the female in Data are reported as mean ± SEM (N), unless otherwise indicated.
Journal of Insect Science, 2019, Vol. 19, No. 1 3
Easton, Pennsylvania The nest cell was connected to the surface of the ground by the
Sphecius speciosus was studied at Lafayette College, Easton, 75 cm, 1.8-cm diameter hole through the center of the packed sand
Northampton County, Pennsylvania (40°41′52″ N, 75°12′28″ W; in the PVC tube (Fig. 2). Trap nests were installed in the ground at
elevation 103 m), hereafter referred to as Easton, in August 2008 an ~45° angle, with their upper ends about 1 cm above the surface
and 2010. Data from 1989 to 1999 show that female wasps pre- of the ground. A female cicada killer entering the trap nest burrow at
sent at the southeast corner of Pardee Hall had a live body mass of the soil surface of the tube could easily walk down the burrow to its
0.965 g ± 0.0103 (416); the corresponding numbers for males were bottom, find the simulated nest cell containing the freshly paralyzed
0.424 g ± 0.0048 (451). Females of this population dig nest burrows cicada and then decide whether or not to appropriate the nest cell
in architectural berms around and next to the south side of Pardee and kleptoparasitize the cicada by laying an egg on it and closing the
Hall. The clay soil in these berms is relatively fine but contains small nest cell with sand; that is, commit what we term PNCK.
stones which may cause cicada killers to abandon a partly finished Natural cicada killer burrows have a tumulus of excavated soil at
burrow. This population provisions its offspring with two species of their mouths and we simulated this tumulus with an approximately
cicadas, Tibicen linnei (Smith and Grossbeck 1907), with an average 8-cm-high pile of damp sand at the mouth of the trap nest, complete
body mass of 1.280 g, and Tibicen tibicen [Linne 1758; formerly with the 2-cm-deep furrow leading away from the burrow opening
Tibicen chloromerus (Walker 1850)], with an average body mass of that is characteristic of natural tumuli and through which the female
1.660 g (Holliday, unpublished data). wasp excavating a burrow pushes excavated soil to the edge of the
tumulus (Fig. 3).
Trap Nest Method Over the course of each day, the artificial tumuli were sprayed
To estimate the rate of conspecific PNCK by female cicada killers, we lightly with water from a hand sprayer as necessary to keep them
used sand-filled trap nests ‘baited’ with a freshly paralyzed cicada, from drying and, presumably, to promote the resemblance of the
taken from a cicada killer returning to provision a nest-cell, and tumuli to freshly-excavated sand. Because natural cicada killer bur-
placed in a simulated nest cell at the bottom of the trap nest. Trap rows are often constructed under or next to isolated rocks, wood, or
nests were made using 75-cm length of 7.5 cm inside-diameter pol- other objects on the surface of the ground, a rock or a piece of wood
yvinyl chloride water pipe (J.M. Eagle Corp.). The pipe was split was placed on the uphill side of the burrow opposite the tumulus
along its length, reassembled with cellophane tape and filled with to serve as a landmark for visiting cicada killers. In rainy weather, a
cleaned, damp sand (‘Play Sand’, Home Depot Corp., 600 ml deion- styrofoam cover (the upper or lower half of a 24 × 24-cm ‘clamshell’
ized water/1,100 cm3 sand); during this filling, the sand was twice takeout food package) was erected as a rain shield over the trap
tamped down firmly using the end of the handle of an insect net. nest’s upper end using bamboo shish kebab skewers inserted through
A pilot hole was pushed into the center of sand-filled tube through- it at an angle into the ground and arranged so as to allow free access
out its length, again using the 1.8-cm diameter handle of an insect by wasps to the burrow. Trap nests were installed between 09:00
net, and the hole in the sand was beveled out halfway to the edge and 15:00 each day. Each trap nest at Easton was exposed to PNCK
of the plastic tube at one end (bevel diameter 3.75 cm), and at that by the natural population of cicada killers in the area for 25 min,
end, a 7.5 cm-diameter plastic cup was attached to the tube with 45 min, or 24 h. The trap nests at Ruby were checked after 23–30 h
cellophane tape. This cup was also filled with damp sand and had a of placing paralyzed cicadas in them.
simulated nest cell impressed into the sand in it using the rounded To determine whether PNCK might be an alternative strategy
end of a 4-cm-diameter trowel handle. Just before final assembly, a for smaller females, we compared provisioning females (owners)
freshly paralyzed cicada taken from a female cicada killer attempting to intruder females. We captured female wasps as they were pro-
to naturally provision a nearby burrow was added to the simulated visioning nests, anesthetized them with CO2, and weighed them to
nest cell with its head directed toward the bottom of the nest cell. the nearest 0.001 g using an Ohaus Adventurer-Pro electronic bal-
Thus, the complete trap nest was a 75-cm column of damp sand con- ance. We also attached a unique number tag to the thorax of each
tained in a 7.5-cm diameter PVC tube containing a freshly paralyzed wasp with cyanoacrylate glue before releasing it and used a match-
cicada in a 4-cm diameter simulated nest cell at its lower end (Fig. 1). ing numbered flag to mark its nest. We defined intruders as females
that entered a nest marked with the number of another female. We
observed intruding females enter marked nest burrows, waited for
them to exit the burrows, then captured, anesthetized, and weighed
them. Intruders were not marked; therefore, some could have been
measured more than once. However, given the extremely large pop-
ulation size at Ruby, such repeated measures are likely to be few.
Results
Bird Theft
In total, 765 min (12.75 h) were spent making observations at the
focal site for bird theft in Ruby over seven observation days from
3–15 September 2009. Cassin’s Kingbirds (Tyrannus vociferans
[Passeriformes: Tyrannidae]) carried out the majority of attacks on
provisioning cicada killers. Thick-billed Kingbirds (Tyrannus crassi-
rostris) were less common in the area, carrying out only three suc-
cessful and two unsuccessful attacks. Attacks by both species were
very similar, except that Thick-billed Kingbirds were never observed
to beat the cicada; hence, both species were lumped into the follow-
Fig. 1. Diagram of the trap nest. Dimensions provided in the text. ing analysis.
4 Journal of Insect Science, 2019, Vol. 19, No. 1
Fig. 2. Examples of the trap nests used in the study, removed from underground: (A) cut longitudinally to show the artificial tunnel and (B) opened at the bottom
to reveal the false burrow and provisioned cell within.
or mesquite, though not always the same one from which the sally
originated. The kingbird often subsequently beat the cicada against
a branch (67% of cases, Fig. 4); sometimes a piece of the cicada fell
off during this process. At times, the bird tossed the cicada in the
air, presumably to reposition it for swallowing, which was always
head first. We never observed kingbirds foraging on any other insects
while at Ruby. On a few occasions, kingbirds were observed taking
the wasp, but never during the focal observations.
We also documented nine cases of prey theft by Greater
Roadrunners (Geococcyx californianus [Cuculiformes: Cuculidae])
during 32 d of working at Ruby. However, their method of attack
differed greatly from that of kingbirds. Roadrunners walked the
periphery of the mine tailings, where most of the burrows were
located, generally staying in the shade of mesquites and other
trees. When a provisioning wasp landed, the roadrunner ran to it,
grasped the cicada in its bill before the wasp could enter a burrow,
and ran back into the shade (Fig. 5) to eat it. The roadrunner some-
Fig. 3. A female Pacific cicada killer walks on the tumulus at the entrance to times tossed the cicada a few times before swallowing it. We also
a buried trap nest. observed a single case of a woodpecker (most likely Ladder-backed
Woodpecker) stealing a cicada.
Although few specimens were recovered during this particu-
We recorded 56 instances of kingbirds (both Cassin’s and Thick-
lar study, all prey were presumed to be Tibicen parallelus Davis
billed) attacking provisioning female cicada killers. Of these, 39
(Hemiptera: Cicadidae), as hundreds of specimens taken in the
(70%) were successful. Kingbirds carried out a successful theft for
course of other studies (Coelho et al. 2012, 2016) at this time proved
every 19.4 min of observation on average, and the number of suc-
to be.
cessful attacks was equal to the number of successful provisionings
(39) recorded during the same observation periods. The kingbirds
usually perched on top of a mesquite bush (generally velvet mes- PNCK
quite, Prosopis velutina) or near the top of a dead agave (Agave All eggs found on paralyzed cicadas placed in trap nest cells were
angustifolia) inflorescence at the periphery of the sand dune, pro- visually indistinguishable from those laid by female cicada killers
viding them with a good view of approaching provisioning cicada in naturally provisioned trap nests used in other experiments in
killers. At times, there were as many as five kingbirds thus perched. 2007–2011 (Fig. 6). Since other Sphecius species or other genera of
On a successful sally, a kingbird generally flew out, seized the cicada large wasps which provision their nest cells with cicadas were never
in its bill; the cicada killer surrendered its cicada at this point and observed in either study area, we assume that all of the eggs we
the kingbird returned to a perch with it, landing on an agave mast found on cicadas in our trap nests were laid by cicada killers of
Journal of Insect Science, 2019, Vol. 19, No. 1 5
Fig. 5. A Greater Roadrunner runs away with a cicada after snatching it from
a Pacific cicada killer.
Fig. 4. A Cassin’s Kingbird with a cicada stolen from a Pacific cicada killer.
The cicada’s shape is distorted from being beaten against the agave mast.
the appropriate species. PNCK rates were similar for S. convallis at
Ruby and S. speciosus at Easton at ~50% (Table 1).
The longer a burrow holding a cicada was left unattended, the
greater the odds that the cicada would be kleptoparasitized (Table 2).
At Ruby, natural burrow owners were not significantly different
in body size from intruders (Table 3, t-test, P = 0.67). We never
recorded a numbered female (an owner) entering the nest of another.
Discussion
Tibicen parallelus is a large (1.1 g, Coelho et al. 2012), palatable
insect, representing a fine prize for a bird. Loaded wasps are slow,
loud, and the visual profile of the pair is much larger than either
alone. Furthermore, the provisioning female is not very maneuvera-
ble. One simple measure of maneuverability is the ratio of flight mus-
Fig. 6. A paralyzed Tibicen tibicen from a trap nest in Easton, PA, with an egg
cle mass to body mass (flight muscle ratio [FMR]; Marden 1987). of the eastern cicada killer laid under right middle leg.
The mean FMR of unladen S. convallis females is 0.401, indicating a
high degree of maneuverability (Coelho et al. 2012). However, once
the wasp is carrying a cicada, the maneuverability is much lower as Sibley 2000). The beating behavior is curious. Kingbirds are known
the FMR drops to a mean of 0.187, which is very near the marginal to beat large insects, such as caterpillars, in order to discard the
level for flight in Hymenoptera (0.18, Marden 1987). Flycatchers, skin (Ball 1936). As the cicadas were completely paralyzed, it was
having evolved as aerial predators, should find heavily loaded pro- needless to kill or incapacitate them. The kingbirds were perhaps
visioning cicada killers easy to capture. Nesting in colonies makes carrying out innate behaviors that they use to immobilize large prey.
cicada killers favorable targets, as a bird may easily intercept multiple Alternatively, the beating may make the cicadas easier to swallow
wasps. It is not surprising that generalist predators such as kingbirds and digest, as the head of T. parallelus is quite wide (10.7 ± 0.09 (32)
and roadrunners have learned to exploit such an abundant and con- mm, Coelho, unpublished data).
spicuous food item. A ladder-backed woodpecker was also observed Kleptoparasitism of wasp prey by birds is sufficiently rare that
attacking a provisioning wasp, and Say’s Phoebe is known to steal Brockmann (1980), referring to Sphex ichneumoneus, claimed, ‘No
cicadas from S. convallis (BBC 1993); we did not observe this species other species of digger wasps are known to be harassed by birds
attacking, though they were in the area. These bird species would in this way’. Avian kleptoparasitism was later documented on its
otherwise be unlikely to capture many cicadas, as T. parallelus is congener Sphex pensylvanicus (Benttinen and Preisser 2009, Lechner
cryptically colored, seldom flies, and remains largely inconspicuous 2010). In body mass, Sx. ichneumoneus averages 0.303 g, Sx. pen-
in the environment (except for calling by the males). The FMR of sylvanicus 0.489 g, whereas their prey average 0.462 and 0.425 g,
T. parallelus is only 0.225 (Coelho et al. 2012), indicating that, in an respectively (Coelho 2011). Sphex and Sphecius are large wasps with
aerial contest, the bird should have no difficulty capturing the cicada large prey, which presumably makes them worth the time and energy
alone. Provisioning cicada killers eliminate crypsis and its attendant spent by birds to kleptoparasitize them. The weak stings of both
advantages to the cicada. The authors confirmed the palatability of genera probably do little to deter the birds’ thievery.
T. parallelus by themselves consuming a number of freshly paralyzed While House Sparrows attacking prey-loaded Sx. ichneumoneus
specimens, roasted on a gas grill, without ill effect. most often approach them directly in flight, the prey are only stolen
It is typical for kingbirds to engage in this style of foraging: sit- once the wasp has been driven down to or dropped it on the ground.
ting on a high perch and hawking insects (Palmer and Fowler 1975, House Sparrows, not surprisingly, lack the aerial ability of kingbirds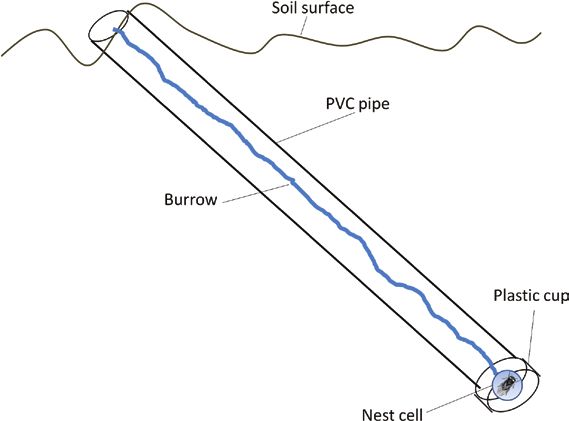
6 Journal of Insect Science, 2019, Vol. 19, No. 1
Table 1. Apparent kleptoparasitism rates after 22–30 h in two species of Sphecius
Species Location No. kleptoparasitized Total Percent kleptoparasitized
Sphecius convallis Ruby, AZ 13 29 45
Sphecius speciosus Easton, PA 27 52 52
Table 2. The effect of trap nest exposure time on kleptoparasitism Table 3. Body mass of owner and intruder female S. convallis at
rate in S. speciosus at Easton, PA Ruby, AZ
Time No. kleptoparasitized Total Percent Role Avg body mass (g) SEM N
kleptoparasitized
Intruder 0.963 0.173 100
25 min 9 49 18 Owner 0.981 0.127 162
45 min 10 37 27
24 h 27 52 52 Owners were not significantly different in size from intruders (t-test,
P = 0.67).
to take the prey in flight. Occasionally, House Sparrows attack the
provisioning wasp on the ground, as does the Greater Roadrunner, Sphecius convallis appears to be better defended, yielding a value
though House Sparrows approach by flying instead of running. of 1. Finally, the foul odor produced by cicada killers on capture (J.
Overall, the 70% success rate recorded here for kingbirds attempt- R. C., C. W. H., and J. M. H., unpublished observations) appears
ing to kleptoparasitize S. convallis is remarkably similar to the 79% to be stronger in Sphecius convallis than in the two other species.
rate of House Sparrows on Sx. ichneumoneus (Brockmann 1980), These three admittedly anecdotal observations could have evolved
60% rate of House Sparrows on Sx. pensylvanicus and 70% rate of in response to predation and prey theft by birds.
Grey Catbirds on Sx. pensylvanicus (Benttinen and Preisser 2009). Assuming that our trap nests are treated as genuine cicada killer
More significantly, the 47% reduction in successful provisioning by nests by females, in Arizona and Pennsylvania Sphecius populations,
S. convallis owing to bird kleptoparasitism far exceeds the 30% rate about half of cicadas left in open nest cells in open burrows for
for Sx. pensylvanicus or 25% for Sx. ichneumoneus. Our results thus 22–30 h are kleptoparasitized and used in situ as egg provisions by
suggest that avian kleptoparasitism was a significant factor in wasp females other than the one that dug the nest cell. This trend probably
fecundity during the season of our study. accounts for the large number of intruder females we see.
Theft of cicada prey by birds is substantial in this population of Few females performing PNCK add a second cicada to the one
cicada killers. Frequent attacks by birds were readily documented they pirate, and the 7 of 46 pirating females that did so (15%) were
in this situation, as opposed to the sporadic, anecdotal cases known only in Easton. This strategy is probably attributable to the real risk
from the literature and seen by us elsewhere. It is likely that large, at Easton (24 min for an average successful hunt) that the original
localized populations of cicada killers attract predators to the site. female will return with her second cicada, lay an egg. and close the
While woodpeckers and Greater Roadrunners probably have little nest cell while the ‘hopeful’ pirate is out hunting for a second cicada
impact on the population, kingbirds may do so, especially during the of her own. In normal nests, eggs laid on a single cicada are nearly
year of our study. In this particular field season, wasps were abun- always male (Dow 1942). Hence, one might predict that eggs laid
dant, with an estimated 6,000 males (via mark-recapture, unpub- during PNCK are predominantly male. Alternatively, half of the eggs
lished), and 2,000–3,000 females (based on a fluctuating number laid on single cicadas could be female, leading to an ‘underclass’ of
of burrows). However, cicadas were not abundant. Singing male small females which may be forced to rely mostly on PNCK to repro-
cicadas were few, as compared with other locales we visited in the duce. However, there was no size difference between PNCK females
region. Ruby Sphecius convallis had a provisioning frequency of and nest owners, suggesting that the ‘single cicada produces a male’
54 min/cicada (C. M. H., unpublished), as compared with 24 min/ rule applies to PNCK as well. When PNCK is frequent, it should lead
cicada in S. speciosus in Pennsylvania (Coelho and Holliday 2008) to strongly male-biased sex ratio, as the fecundity of both owner and
and 90 min in New Jersey (Grant 2006). Thus, foraging success was intruder will increase if they lay male eggs.
relatively low at Ruby. With kingbirds taking nearly half of the prey PNCK should be favored by natural selection, as it increases a
at times, the impact of prey theft on the population is expected to female’s fecundity by increasing her potential number of offspring
be large. while simultaneously lowering the time/energy investment in the
The high avian kleptoparasitism pressure experienced by Pacific resulting offspring. Although females engaging in PNCK might
cicada killers in the Ruby, Arizona, population may have led to some sometimes encounter agonistic behavioral defense by the resident
evolutionary adaptations that we noted subjectively, but perhaps are female from whom they are attempting to steal a provisioned nest
deserving of further study. First, the cuticle of S. convallis appears to cell (Lin 1963), we have never seen an interaction result in the death
be harder and/or thicker than that of S. speciosus or S. grandis. The of one of the females. This effect should be greater at Ruby, with its
authors have completed extensive thermoregulation studies on all 54.5 ± 9.9(19) min successful hunt duration, versus 24 min at Easton
three species (Coelho 2001, Coelho et al. 2007, 2016). After stab- (C. W. H., unpublished data), but that was not shown, as the rates of
bing hundreds of each in the head, thorax, and abdomen with a PNCK were about the same after 22–30 h at both sites.
26-ga thermocouple, we noted that the exoskeleton of S. convallis is Small females should have more difficulty capturing and return-
much more difficult to penetrate than that of the other two species, ing with cicadas of a given size than large ones (Lin and Michener
and occasionally bends the thermocouple. Second, having worked 1972, Coelho 1997, Coelho et al. 2012), so PNCK should be favored
with S. speciosus for decades, we find its sting effect to be rela- as possibly the only way that small females can reproduce in areas
tively mild, ~0.5 on the Schmidt scale of sting pain (Schmidt 2016). where only larger cicadas are available. However, this notion is notJournal of Insect Science, 2019, Vol. 19, No. 1 7
supported by data from Ruby showing that owners and intruders Coelho, J. R., C. W. Holliday, and J. M. Hastings. 2011. The geographic dis-
are not significantly different in size. Perhaps, all females perform tributions of cicada killers (Sphecius; Hymenoptera, Crabronidae) in the
PNCK when they can, but it may be the only way that small ones Americas. Open Entomol. J. 5: 31–38.
Coelho, J. R., J. M. Hastings, and C. W. Holliday. 2012. Near-optimal forag-
can reproduce at all.
ing in the pacific cicada killer sphecius convallis patton (Hymenoptera:
At high female population densities and/or in times of low prey
Crabronidae). Insects. 3: 133–140.
availability (i.e., Ruby in 2009), PNCK should be far more advan-
Coelho, J. R., C. W. Holliday, J. M. Hastings, and C. M. Phillips. 2016.
tageous than ‘honest’ provisioning of a wasp’s own nest cells and, Thermal biology of Pacific cicada killers, Sphecius convallis Patton, in the
thus, selected for. Cicada kleptoparasitism by birds at Ruby would Upper Sonoran Desert. J. Therm. Biol. 57: 101–109.
make conspecific PNCK even more advantageous. However, we saw Curran, C. H. 1962. Insects in your life. Sheridan House, New York.
slightly greater rates of nest cell piracy at Easton in summer 2010 Dambach, C. A., and E. Good. 1943. Life history and habits of the cicada
than at Ruby in 2009, so avian kleptoparasitism is not what was killer in Ohio. Ohio J. Sci. 43: 32–41.
driving PNCK in Easton, as House Sparrow kleptoparasitism of Davis, W. T. 1924. Cicada killing wasps and flies. J. New York Entomol. Soc.
cicadas carried by CKs has been seen only three times at Easton over 32: 113.
Dow, R. 1942. The relation of the prey of Sphecius speciosus to the size and sex
about 20 yr of observations.
of the adult wasp (Hym.: Sphecidae). Ann. Entomol. Soc. Am. 35: 310–317.
It appears that the large, paralyzed prey of cicada killers are attrac-
Evans, H. E. 1966. The comparative ethology and evolution of the sand wasps.
tive to a variety of thieves. Interspecific prey theft is common among
Harvard University Press, Cambridge, MA.
birds (Brockmann and Barnard 1979), and it is not surprising that Evans, H. E., and K. M. O’Neill. 2007. The sand wasps: natural history and
agile, generalist predators such as birds have discovered how to exploit behavior. Harvard. Cambridge, MA.
the provisioning system of Sphecius. Intraspecific kleptoparasitism is, Grant, P. R. 2006. Opportunistic predation and offspring sex ratios of cica-
however, much more subtle. If we had not utilized trap nests, we would da-killer wasps (Sphecius speciosus Drury). Ecol. Entomol. 31: 539–547.
not have known that it was occuring at all. This cryptic alternative Hastings, J. M., J. R. Coelho, and C. W. Holliday. 2008. Mating at high pop-
strategy provides a mechanism by which all females can potentially ulation density in a colonial territorial wasp, Sphecius speciosus Drury
reproduce, despite large differences in their adult sizes and without (Hymenoptera: Crabronidae). J. Kansas Entomol. Soc. 81(3): 301–308.
Hastings, J. M., C. W. Holliday, A. Long, K. Jones, and G. Rodriguez.
regard to the size range of cicada prey available in their environment.
2010. Size-specific provisioning by cicada killers, Sphecius speciosus,
(Hymenoptera: Crabronidae) in North Florida. Florida Entomol. 93(3):
412–421.
Acknowledgments
Holliday, C. W., J. M. Hastings, and J. R. Coelho. 2009. Cicada prey of
We thank Howard M. Frederick, owner of Ruby Mines, for access to the prop- New World cicada killers, Sphecius spp. (Hymenoptera: Crabronidae).
erty and Sundog Hunter for his hospitality and assistance in plant identification. Entomol. News. 120(1): 1–17.
Lechner, G. K. 2010. A second report of avian kleptoparasitism of the great
black wasp, Sphex pensylvanicus L. (Hymenoptera: Sphecidae: Sphecinae).
References Cited Entomol. News 121(3): 302–303.
Ball, A. E. 1936. American Land Birds. Dodd, Mead and Company, Inc., New Lin, N. 1963. Observations of suspected density dependent fighting between
York. females of the cicada killer wasp Sphecius speciosus. Bull. Brooklyn
(BBC) British Broadcasting Corporation. 1993. The Ghosts of Ruby. Rubin Entomol. Soc. 58: 121–123.
Tarrant Productions, Cambridge, MA. https://youtu.be/wKcPTpi1syg Lin, N., and C. D. Michener. 1972. Evolution of sociality in insects. Q. Rev.
Benttinen J., and E. L. Preisser 2009. Avian kleptoparasitism of the digger Biol. 47: 131–159
wasp Sphex pensylvanicus. Can. Entomol. 141: 604–608 Marden, J. H. 1987. Maximum lift production during takeoff in flying ani-
Brockmann, H. J. 1980. House sparrows kleptoparasitize digger wasps. mals. J. Exp. Biol. 130: 235–248.
Wilson Bull. 92(3): 394–398. Palmer, E. L., and H. S. Fowler. 1975. Fieldbook of natural hstory. McGraw-
Brockmann, H. J., and C. J. Barnard. 1979. Kleptoparasitism in birds. Anim. Hill, New York.
Behav. 27: 487–514. Pfennig, D. W., and H. K. Reeve. 1989. Neighbor recognition and con-
Brockmann, H. J., and R. Dawkins. 1979. Joint nesting in a digger wasp as text-dependent aggression in a solitary wasp, Sphecius speciosus
an evolutionarily stable preadaptation to social life. Behaviour. 71: 3–4. (Hymenoptera:Sphecidae). Ethology. 80: 1–18.
Coelho, J. R. 1997. Sexual size dimorphism and flight behavior in cicada kill- Sann, M., O. Niehuis, R. S. Peters, C. Mayer, A. Kozlov, L. Podsiadlowski, S.
ers (Sphecius speciosus). Oikos. 79: 371–375. Bank, K. Meusemann, B. Misof, C. Bleidorn, et al. 2018. Phylogenomic
Coelho, J. R. 2001. Behavioral and physiological thermoregulation in male analysis of Apoidea sheds new light on the sister group of bees. BMC Evol.
cicada killers (Sphecius speciosus) during territorial behavior. J. Therm. Biol. 18: 71.
Biol. 26: 109–116. Schmidt, J. O. 2013. Sand dune of Ruby, Arizona, an anthropogenically cre-
Coelho, J. R. 2011. Effects of prey size and load carriage on the evolution ated biodiversity hotspot for wasps and their velvet ant parasitoids, pp.
of foraging strategies in wasps, pp. 23–38. In C. Polidori (ed), Predation 172–174. In G. J. Gottfried, P. F. Ffolliott, B. S. Gebow, L. G. Eskew, and
in the Hymenoptera: an evolutionary perspective. Transworld Research L. C. Collins (eds). Merging science and management in a rapidly changing
Network publishing, Kerala, India. world: biodiversity and management of the Madrean Archipelago III; 1–5
Coelho, J. R., and C. W. Holliday. 2008. The effect of hind-tibial spurs on May 2012; Tucson, AZ. Proceedings, RMRS-P-67. Fort Collins, CO.
digging rate in female eastern cicada killers. Ecol. Entomol. 33: 1–5. Schmidt, J. O. 2016. The sting of the wild. Johns Hopkins University Press,
Coelho, J. R., C. W. Holliday, J. M. Hastings, E. Maty, M. Swigart, and Baltimore, MD.
A. Mendell. 2007. Thermoregulation in male western cicada killers Sibley, D. A. 2000. The sibley guide to birds. Alfred A. Knopf, New York.
(Sphecius grandis Say) in the Chihuahuan desert. J. Therm. Biol. 32(5): Villalobos, E. M., and T. Shelly. 1996. Intraspecific nest parasitism in the sand wasp
270–275. Stictia heros (Fabr.) (Hymenoptera: Sphecidae). J. Insect Beh. 9: 105–119.You can also read

















































