Ice Age effects on genetic divergence of the American crocodile (Crocodylus acutus) in Panama: reconstructing limits of gene flow and ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Evolution, 2023, 77(1), 329–334
https://doi.org/10.1093/evolut/qpac006
Advance access publication 8 December 2022
Technical Note
Ice Age effects on genetic divergence of the American
crocodile (Crocodylus acutus) in Panama: reconstructing
limits of gene flow and environmental ranges: a reply to
O’Dea et al.
Jose Avila-Cervantes, PhD1, Hans C. E. Larsson, PhD1
Downloaded from https://academic.oup.com/evolut/article/77/1/329/6873140 by guest on 28 January 2024
McGill University, Montreal, Quebec, Canada
1
Corresponding author: McGill University, Redpath Museum 859 Sherbrooke Street West, Montreal, Quebec H3A 0C4, Canada.
Email: jose.avilacervantes@mail.mcgill.ca
Abstract
O’Dea et al. (2022) (Pleistocene sea level changes and crocodile population histories on the isthmus of panama: a comment on Avila-Cervantes
et al. (2020). Evolution, 76(11), 2778–2783. https://doi.org/10.1111/evo.14610) question our hypothesis that sea-level changes during the past
glaciation played a role in restricting gene flow between Pacific and Caribbean Crocodylus acutus in Panama. They argue that an error in sea-
level high-stand reconstruction during the last interglacial period (118–130 ka) does not support our hypothesis. Although they are correct in our
high-stand reconstruction error, overlooked the point in that we were presenting a model of restricted gene flow across the Panamanian Isthmus
during low sea levels. We review the assumptions of gene demographic methods, emphasizing that we were focusing on times of genetic
divergence. We expand here why gene flow between these coastal populations could have been restricted during the last glacial maximum
(19–26.5 ka) and the 50,000 years preceding it when sea levels were lower than today. O’Dea et al. suggest local climates may have played
larger roles than sea levels. We demonstrate that paleoclimate estimates for the past 3.3 Ma in Panama are within the bounds of extant C.
acutus. The importance of Ice Age Sea-level dynamics on Neotropical species was likely profound and should be incorporated into evolutionary
studies of these taxa.
Keywords: Crocodylus, Ice Age, relative sea levels, gene flow
We thank O’Dea et al. (2022) for their comments on our extending only to the Caribbean coast and no reported pop-
publication and for highlighting an error we made using ulations on the adjacent Pacific coast (Thorbjarnarson et al.,
maximum sea levels during the last interglacial (LIG) that 2006). In addition, we inferred the demographic history and
occurred about 130,000 years ago (ka) to 118 ka. We take calculated the divergence times of the sampled populations.
this opportunity to clarify key aspects of our paper, develop Demographic models use genetic and genomic data to esti-
a more extensive discussion on the importance of including mate population size, migration rates, and divergence times
environmental changes to population demographics, and em- between populations (Gutenkust et al., 2010). Divergence
phasize that any LIG sea-level change above current levels has times are estimated from generation times, so the mod-
no bearing on our results. els require some assumptions of average generation times
Our study analyzed the genetic variation of the American in those populations. We used two methods for the demo-
crocodile (Crocodylus acutus) on both sides of the Central graphic models: Approximate Bayesian Computation (ABC)
American Isthmus. We sampled several populations from the on DIYABC v.2 (Cornuet et al., 2014) and the Diffusion
Caribbean and Pacific coasts of Panama to estimate the impact approximation demographic inference (δaδi) (Gutenkunst et
of the formation of the Isthmus of Panama (about 3 million al., 2009). It is important to note that the calculated times
years ago) on the restriction of gene flow and construction of restricted gene flow can only be the most recent isolation
of the Panama Canal (about 100 years ago) on the presumed and, in cases where secondary contact is inferred, the penul-
resumption of gene flow between these coastal populations. timate isolation (Table 1).
These populations were hypothesized to be genetically sep- The results identified several periods of restricted gene
arated at least until the construction of the Panama Canal. flow with the most realistic ranging between 26.7 and 66.8
Species distribution maps of C. acutus in the Atrato Basin ka (Table 1). All were about 2 orders of magnitude younger
in northern Colombia sometimes reconstruct a continuous than the formation of the Isthmus of Panama. Our primary
range between the Caribbean and Pacific coast (e.g., Briggs- conclusion was that the Isthmus was not a barrier at all, and
Gonzalez et al., 2017; Rossi et al., 2020). However, these at best semipermeable to these large-bodied amphibious rep-
populations are restricted to the Atrato River with a range tiles. In an attempt to explore other factors that may have
Received October 2, 2021; revisions received September 4, 2022; accepted October 28, 2022
© The Author(s) 2022. Published by Oxford University Press on behalf of The Society for the Study of Evolution (SSE). All rights reserved. For permissions,
please e-mail: journals.permissions@oup.com330 Avila-Cervantes and Larsson
Table 1. Estimated divergence times for populations using DIYABC and δaδi. Only the most supported models from Avila-Cervantes et al. (2021) are
presented.
Divergence Barrier Population DIYABC Model 2 δaδi
pairs
Divergence in Divergence in Secondary contact in
thousands of years (ka) thousands of years (ka) thousands of years (ka)
Caribbean Sympatric GAL-BCI 7.82–9.77 5.8–7.3 3 - 3.7
Pacific Sympatric LAG- 11.04–13.8 1.1–1.4 1.1 - 1.4
COIB
Caribbean—Pacific Allopatric GAL-LAG 32.6–40.75 30.9–38.6 20.7 - 25.9
Caribbean—Pacific Allopatric BCI-LAG NA 53.2–66.8 26.7 - 33.4
been responsible for these relatively recent divergence dates isolation (Figure 1B). Prior global glaciations and intergla-
Downloaded from https://academic.oup.com/evolut/article/77/1/329/6873140 by guest on 28 January 2024
between the Pacific and Caribbean populations, we looked for cial periods that occurred throughout the Ice Age may have
temporal associations between sea-level lows and restricted had similar effects on crocodile gene flow across the Isthmus
gene flow. Extreme sea-level lows are expected to have made but are simply not recoverable due to the repeated periods
the Isthmus which was otherwise semipermeable to crocodile of admixture between the two coasts. Only the last major
gene flow, less permeable. restriction to gene flow event is recoverable with the present
The ABC and δaδi divergence times coincide within the genomic data. There is no fossil record of the species to cali-
period of the LGM (26.5–19 ka) and the penultimate gla- brate earlier divergences.
cial maximum (140 ka) (Colleoni et al., 2016). These periods O’Dea et al. suggest paleoclimates may have played a more
were characterized by extreme rises and falls in eustatic sea important role than RSL on C. acutus gene flow over the
levels. These levels varied from +6 m (Blanchon et al., 2009) Isthmus. Below, we present data that challenge this hypothe-
during the LIG to −130 to −120 m during the LGM (Gowan sis. Adaptations of the genus permit C. acutus to be the most
et al., 2021; Miller et al., 2020). widely distributed crocodile species in the Neotropics (Oaks,
The major argument O’Dea et al. have is our reconstruction 2011). The species inhabit mangrove-lined coastal lagoons
of sea levels around Panama during the LIG using the uncor- or estuaries, offshore cays and coral atolls, and rivers and
rected values presented by Rohling and colleagues (2017), and reservoirs. These environments range from hypersaline to
that our divergence times do not match exactly with the LGM. freshwater (Thorbjarnarson, 1989). Water salinities reported
However, in the context of our work, the absolute LIG sea-level for C. acutus can vary from 0 ppt in coastal lagoons to >50
height does not impact our results as our demographic models ppt in atolls, and offshore salinities range from 34 to 36 ppt
were only testing for the most recent discernible barrier to gene (Platt et al., 2013). The species inhabit environments with
flow between each coast and subsequent potential secondary contrasting temperatures, precipitation, and elevations over
contacts. Any sea-level drop over current levels is expected to a vast range that spans from the northwestern Pacific coast
reduce gene flow across the Isthmus. The beginning of the LGM of Mexico to Ecuador to the Caribbean islands. To explore
(19–26.5 ka) nearly overlaps with the recovered dates of popu- the bioclimatic profile of C. acutus we used 400-point obser-
lation divergences between the allopatric adjacent populations vations of the species (Thorbjarnarson et al., 2006) and
(26.7–66.8 ka) and we suggested this was a driver in genetic extracted the values of the eight bioclimatic variables from
isolation between the two coastal populations. Although these WorldClim 2 (Fick & Hijmans, 2017) of each locality, at
times do not completely overlap, the coincidence of the dra- a resolution of 30 arc seconds. We used R V.3.6.1 (R Core
matic sea level drops during the LGM was discussed as a pos- Team, 2021) and the packages DISMO V.1.3 (Hijmans et
sible mechanism of gene flow restriction (Avila-Cervantes et al., 2020) and MAPTOOLS V.1.2 (Bivand & Lewin-Kohn,
al., 2021). Any earlier barriers to gene flow during older glacial 2021). The results indicate a broad range of temperature
maxima cannot be inferred, as these events would have been and precipitation regimes for the species’ localities (Table 2).
erased by subsequent pulses of genetic flow between the coasts. These extreme arid to tropical and cool to hot environments
However, the LGM low stand sea levels occur at least 2–40 present a remarkably resilient species that subtle paleocli-
ka after the estimated genetic divergence between the popu- matic changes are unlikely to have affected. At no point in the
lations on the two coasts. This does not discount the possibil- past 3.3 million years has the paleoclimate of Panama been
ity that during the drop in relative sea level (RSL) preceding estimated to have passed beyond the bioclimatic variables
the LGM, a threshold was passed where sea levels were low C. acutus encounters within its current distribution. Table 3
enough to cause the Isthmus to switch from a semi-perme- indicates the profile of 70 localities of C. acutus in Panama
able barrier to an impermeable barrier to gene flow in these (Thorbjarnarson et al., 2006) for the bioclimatic variables
animals. To further explore this, we present models of inter- from PaleoClim (Brown et al., 2018) with a resolution of
mediate low RSLs that occurred throughout the time span 2.5 arc seconds during the last glacial maximum (21 ka), the
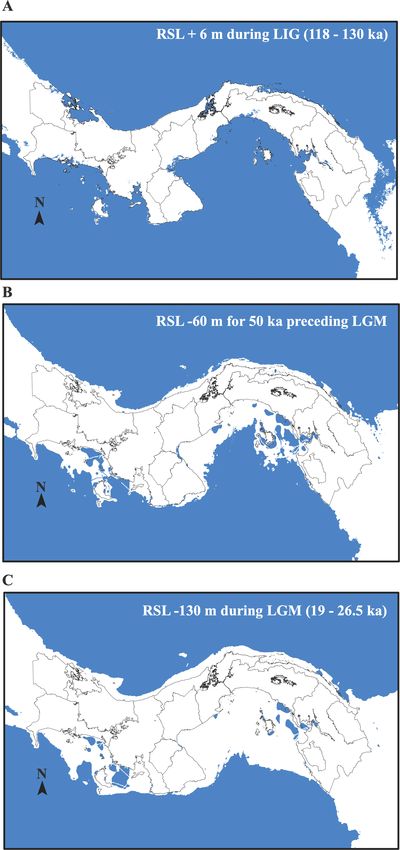
between the LIG and LGM (Figure 1). These sea-level drops LIG (130 ka), and the Pliocene (3.3 Ma). Temperature and
were nonlinear. Between approximately 20–70 ka were more precipitation values are well within the ranges of the species’
than 60 m below current levels (Rohling et al., 2017). We actual distribution today. The environmental variation and
hypothesize these lows were enough to significantly limit ranges among current populations of C. acutus are stunning
gene flow. This 50,000-year-long period preceding the LGM and highlight the robustness of this species.
overlaps the divergence estimates we recovered and suggests We agree that it is important to consider other drivers than
this was the threshold in RSL drop that factored into their changes in sea level to understand the biology and evolutionEvolution (2023), Vol. 77, No. 1 331
Downloaded from https://academic.oup.com/evolut/article/77/1/329/6873140 by guest on 28 January 2024
Figure 1. (A) Relative sea level (RSL) of 6 m above present during the LIG, about 118–130 ka (Blanchon et al., 2009). (B) RSL of 60 m below present,
about 50 ka preceding LGM (Rohling et al., 2017). Note that for most of this time RSL was even lower. (C) RSL of 130 m below present during the LGM,
about 19–26.5 ka (Gowan et al., 2021; Miller et al., 2020). We used the GEBCO grids (2021) and the Panama province boundaries map from the STRI
GIS data portal (https://stridata-si.opendata.arcgis.com/) to create the maps.332 Avila-Cervantes and Larsson
of C. acutus in the Isthmus of Panama. However, the spe- Lessios, 2008; Stange et al., 2018). However, more restricted
cies biology and its capacity to inhabit a wide range of envi- coastal populations, such as mangroves, are revealing star-
ronments (Table 2) make it resilient to the increased salinity tling regional demographics that coincide with the LGM
(discussed above) and climatic variation and rainfall changes (Ceron-Souza et al., 2015).
related to the glacial–interglacial cycles (Table 3). We did find that the genetic divergence of Crocodylus acutus
O’Dea et al. also suggest a series of other avenues to does not coincide with the formation of the Central American
explore and interpret crocodile gene flow disruption across Isthmus, 3 Ma, but instead coincides with the drop in RSL
the Isthmus such as paleontology and archeology. Although leading up to and during the LGM, about 20 ka. Although
there is a wealth of crocodyliform paleontological and arche- paleoclimate most certainly played a role in crocodile local
ological data we don’t see how the Miocene gavialoid or rela- evolution, we demonstrate that the extreme tolerances of
tively recent archeological records O’Dea et al. suggest could extant C. acutus make such claims for their Pacific-Caribbean
help interpret Crocodylus acutus gene flow across the Isthmus divergence unlikely. Rather, the drop in RSL leading to the
during the Pliocene and Quaternary. The species of gavialoid LGM seems like a plausible explanation that could have
suggested, Aktiogavialis caribesi, was recovered from the late caused the Isthmus to become a barrier to gene flow for these
Miocene Urumaco Formation (c. 10–9 Ma) (Salas-Gismondi amphibious taxa but further sampling and testing are needed
Downloaded from https://academic.oup.com/evolut/article/77/1/329/6873140 by guest on 28 January 2024
et al., 2019), predating the formation of the Isthmus by up to confirm this theory. The wide range of climate regimes for
to seven million years and long before the immigration of this species implies that past climates would have had little
Crocodylus from Africa, that occurred about 5 Ma (Avila- effect on its distribution; however, we expect that the relative
Cervantes & Larsson, 2018; Oaks, 2011). The archeological elevation and width of the Isthmus should. We highlight that
data postdates any of our inferred divergence dates by several Ice Age Sea-level dynamics may have played larger roles in the
thousand years and cannot offer clues to the factors acting evolution of Central American taxa and processes of genetic
on those divergences during the time span between the LIG disruption than previously appreciated. Moreover, C. acutus
and LGM. may be a remarkable taxa to begin examining these effects
In summary, we agree that we made an error in the recon- due to its broad climate distribution and amphibious habitat.
struction of the RSL high stand during the LIG. Sea-level
high stands are likely to facilitate gene flow for these animals.
The majority of terrestrial and marine systems studied in the Data availability
region recover little to no influence of the Ice Age on popu- There is no data to be archived.
lation demographics but focus on the original formation of
the Isthmus (Bacon et al., 2015; Cowman & Belwood, 2013;
Author contributions
J.A.C. wrote the manuscript. H.C.E.L. wrote the manuscript.
Table 2. Bioclimatic profile of C. acutus based on 400 localities in its All authors gave final approval for publication.
range of distribution (Thorbjarnarson et al., 2006) and the bioclimatic
variables from WorldClim 2 (Fick & Hijmans, 2017).
Funding statement
Bioclimatic variable Min Mean Max
Support for this project was provided by NSERC Discovery
Annual mean temperature (°C) 21.31 26.13 28.81 Grant to H.C.E.L.
Max temperature of warmest month (°C) 29.1 32.55 35.9
Min temperature of coldest month (°C) 9.9 19.54 23.5 Conflict of interest: The authors declare no conflict of in-
Temperature annual range (°C) 7.4 13.01 25.8
terest.
Annual precipitation (mm) 217 1,838 3,840
Precipitation of wettest month (mm) 70 307 734 Acknowledgments
Precipitation of driest month (mm) 0 29.1 132
This research was performed using the infrastructure of the
Elevation (m) 0 91.38 1,265
Integrated Quantitative Biology Initiative, funded by the
Quebec government, McGill University, and the Canadian
Table 3. Bioclimatic profile of C. acutus based on 70 localities in Panama (Thorbjarnarson et al., 2006) and the bioclimatic variables from PaleoClim
(Brown et al., 2018) for the last glacial maximum (21 ka) (Karger et al., 2021), the last interglacial 130 ka (Otto-Bliesner et al., 2006), and the Pliocene
(3.3 Ma) (Dolan et al., 2015).
Last glacilal maximum (21 ka) Last Intreglacial (130 ka) Pliocene M2 (3.3 Ma)
Bioclimatic variable Min Mean Max Min Mean Max Min Mean Max
Annual mean temperature (°C) 19.5 22.3 23.5 23.1 25.6 26.5 24.7 27.1 28.1
Max temperature of warmest month (°C) 22.4 26.6 30 26.9 29.5 30.5 NA NA NA
Min temperature of coldest month (°C) 16.2 18.9 20.4 20 22.4 24.1 NA NA NA
Temperature annual range (°C) 4 22.2 23.6 5.6 7.1 8.8 NA NA NA
Annual precipitation (mm) 1,358 2,121 3,214 1,380 2,381 3,340 1,908 2,766 3,753
Precipitation of wettest month (mm) 198 322.6 514 216 365 550 463 570.7 753
Precipitation of driest month (mm) 9 35.91 182 11 28.4 169 0 3.9 137Evolution (2023), Vol. 77, No. 1 333
Foundation of Innovation project 33122. We would like to Gowan, E. J., Zhang, X., Khosravi, S., Rovere, A., Stocchi, P., Hughes, A.
thank the reviewers for their thoughtful comments toward L. C., Gyllencreutz, R., Mangerud, J., Svendsen, J. I., & Lohmann,
improving our manuscript. G. (2021). A new global ice sheet reconstruction for the past 80
000 years. Nature Communications, 12(1), 1199–1199. https://doi.
org/10.1038/s41467-021-21469-w
References Gutenkunst, R. N., Hernandez, R. D., Williamson, S. H., Bustamante, C.
Avila-Cervantes, J., & Larsson, H. C. E. (2018). Crocodylus dispersion D., & Los Alamos National Lab. (LANL), Los Alamos, NM (Unit-
based on fossil calibration and DNA sequences, using individuals ed States). (2009). Inferring the joint demographic history of mul-
from wild populations. Latin American Congress of Vertebrate Pa- tiple populations from multidimensional snp frequency data. Plos
leontology. August 2018. Villa de Leyva, Colombia. Genetics, 5(10). https://doi.org/10.1371/journal.pgen.1000695
Avila-Cervantes, J., Arias, C., Venegas-Anaya, M., Vargas, M., Larsson, Gutenkunst, R., Hernandez, R., Williamson, S., & Bustamante, C.
H. C. E., & McMillan, W. O. (2021). Effect of the Central Ameri- (2010). Diffusion approximations for demographic inference:
can Isthmus on gene flow and divergence of the American crocodile DaDi. Nature Precedings, (20100630). https://doi.org/10.1038/
(Crocodylus acutus). Evolution; International Journal of Organic npre.2010.4594.1
Evolution, 75(2), 245–259. https://doi.org/10.1111/evo.14139 Hijmans, R. S., Phillips, S., Leathwick, J., & Elith, J. (2020). dismo:
Bacon, C. D., Silvestro, D., Jaramillo, C., Smith, B. T., Chakrabarty, Species Distribution Modeling. R package version 1.3-3. https://
P., & Antonelli, A. (2015). Biological evidence supports an early CRAN.R-project.org/package=dismo
Downloaded from https://academic.oup.com/evolut/article/77/1/329/6873140 by guest on 28 January 2024
and complex emergence of the Isthmus of Panama. Proceedings of Lessios, H. A. (2008). The great American schism: Divergence of ma-
the National Academy of Sciences of the United States of America, rine organisms after the rise of the Central American Isthmus. An-
112(19), 6110–6115. https://doi.org/10.1073/pnas.1423853112 nual Review of Ecology, Evolution, and Systematics, 39(1), 63–91.
Bivand, R., & Lewin-Koh, N. (2021). maptools: Tools for Handling https://doi.org/10.1146/annurev.ecolsys.38.091206.095815
Spatial Objects. R package version 1.1-2. https://CRAN.R-project. Karger, D. N., Nobis, M. P., Normand, S., Graham, C. H., & Zim-
org/package=maptools mermann, N. E. (2021). CHELSA-TraCE21k v1. 0. Downscaled
Blanchon, P., Eisenhauer, A., Fietzke, J., & Liebetrau, V. (2009). Rap- transient temperature and precipitation data since the last glacial
id sea-level rise and reef back-stepping at the close of the last in- maximum. Climate of the Past Discussions, 1–27.
terglacial high stand. Nature, 458(7240), 881–884. https://doi. Miller, K. G., Browning, J. V., Schmelz, W. J., Kopp, R. E., Mountain,
org/10.1038/nature07933 G. S., & Wright, J. D. (2020). Cenozoic sea-level and cryospher-
Briggs-Gonzalez, V., Bonenfant, C., Basille, M., Cherkiss, M., Beau- ic evolution from deep-sea geochemical and continental margin
champ, J., & Mazzotti, F. (2017). Life histories and conservation records. Science Advances, 6(20), 1346. https://doi.org/10.1126/
of long-lived reptiles, an illustration with the American crocodile sciadv.aaz1346
(Crocodylus acutus). Journal of Animal Ecology, 86(5), 1102– O’Dea, A., Flantua, S. G. A., Leray, M., Lueders-Dumont, J. A., & Tit-
1113. https://doi.org/10.1111/1365-2656.12723 comb, M. C. (2022). Pleistocene sea level changes and crocodile
Brown, J. L., Hill, D. J., Dolan, A. M., Carnaval, A. C., & Haywood, population histories on the isthmus of panama: a comment on Avi-
A. M. (2018). Paleoclim, high spatial resolution paleoclimate sur- la-Cervantes et al. (2020). Evolution, 76(11), 2778–2783. https://
faces for global land areas. Scientific Data, 5, 180254. https://doi. doi.org/10.1111/evo.14610
org/10.1038/sdata.2018.254 Oaks, J. R. (2011). A time-calibrated species tree of Crocodylia reveals
Cerón-Souza, I., Gonzalez, E. G., Schwarzbach, A. E., Salas-Leiva, D. E., a recent radiation of the true crocodiles. Evolution, 65, 3285–3297.
Rivera-Ocasio, E., Toro-Perea, N., Bermingham, E., & McMillan, https://doi.org/10.1111/j.1558-5646.2011.01373.x
W. O. (2015). Contrasting demographic history and gene flow pat- Otto-Bliesner, B. L., Cape, L. I. P. M., Marshall, S. J., Overpeck, J. T., Miller,
terns of two mangrove species on either side of the Central Amer- G. H., & Hu, A. (2006). Simulating Arctic climate warmth and icefield
ican Isthmus. Ecology and Evolution, 5(16), 3486–3499. https:// retreat in the last interglaciation. Science, 311(5768), 1751–1753.
doi.org/10.1002/ece3.1569 Platt, S. G., Thorbjarnarson, J. B., Rainwater, T. R., & Martin, D. R.
Colleoni, F., Wekerle, C., Jens-Ove, N., Brandefelt, J., & Masina, S. (2013). Diet of the American crocodile (Crocodylus acutus) in ma-
(2016). Constraint on the penultimate glacial maximum northern rine environments of coastal Belize. Journal of Herpetology, 47(1),
hemisphere ice topography (≈140 kyrs bp). Quaternary Science Re- 1–10. https://doi.org/10.1670/12-077
views, 137, 97–112. https://doi.org/10.1016/j.quascirev.2016.01.02 R Core Team. (2021). R: A language and environment for statistical
Cornuet, J. M., Pudlo, P., Veyssier, J., Dehne-Garcia, A., Gautier, M., computing. R Foundation for Statistical Computing. https://ww-
Leblois, R., Marin, J. M., & Estoup, A. (2014). Diyabc v2.0: A w.R-project.org/.
software to make approximate Bayesian computation inferences Rohling, E. J., Hibbert, F. D., Williams, F. H., Grant, K. M., Marino, G.,
about population history using single nucleotide polymorphism, Foster, G. L., Hennekam, R., de Lange, G. J., Roberts, A. P., Yu, J.,
DNA sequence and microsatellite data. Bioinformatics (Oxford, Webster, J. M., & Yokoyama, Y. (2017). Differences between the
England), 30(8), 1187–1189. last two glacial maxima and implications for ice-sheet, δ18o, and
Cowman, P. F., & Bellwood, D. R. (2013). Vicariance across major ma- sea-level reconstructions. Quaternary Science Reviews, 176.
rine biogeographic barriers: Temporal concordance and the relative Rossi, N. A., Menchaca-Rodriguez, A., Antelo, R., Wilson, B., McLaren,
intensity of hard versus soft barriers. Proceedings. Biological Sci- K., Mazzotti, F., Crespo, R., Wasilewski, J., Alda, F., Doadrio, I., Bar-
ences, 280(1768), 20131541–20131541. https://doi.org/10.1098/ ros, T. R., Hekkala, E., Alonso-Tabet, M., Yairen, A.-G., Lopez, M.,
rspb.2013.1541 Espinosa-Lopez, G., Burgess, J., Thorbjarnarson, J. B., Ginsberg, J.
Dolan, A. M., Haywood, A. M., Hunter, S. J., Tindall, J. C., Dowsett, R., … Chiang, T.-Y. (2020). High levels of population genetic differ-
H. J., Hill, D. J., & Pickering, S. J. (2015). Modelling the enigmatic entiation in the American crocodile (Crocodylus acutus). PLoS One,
late Pliocene glacial event—Marine isotope stage m2. Global and 15(7), 0235288. https://doi.org/10.1371/journal.pone.0235288
Planetary Change, 128, 47–60. https://doi.org/10.1016/j.glopla- Salas-Gismondi, R., Moreno-Bernal, J. W., Scheyer, T. M., Sánchez-Vil-
cha.2015.02.001 lagra Marcelo, R., & Jaramillo, C. (2019). New miocene Caribbean
Fick, S. E., & Hijmans, R. J. (2017). WorldClim 2: New 1-km spa- gavialoids and patterns of longirostry in crocodylians. Journal of
tial resolution climate surfaces for global land areas. International Systematic Palaeontology, 17(12), 1049–1075. https://doi.org/10.1
Journal of Climatology, 37, 4302–4315. https://doi.org/10.1002/ 080/14772019.2018.1495275
joc.5086 Stange, M., Sánchez-Villagra, M. R., Salzburger, W., & Matschiner, M.
GEBCO Compilation Group. (2021). GEBCO 2021 Grid. doi: (2018). Bayesian divergence-time estimation with genome-wide
10.5285/c6612cbe-50b3-0cff-e053-6c86abc09f8f. single-nucleotide polymorphism data of sea catfishes (ariidae)334 Avila-Cervantes and Larsson
supports miocene closure of the Panamanian Isthmus. Systematic Thorbjarnarson, J., Mazzotti, F., Sanderson, E., Buitrago, F., Laz-
Biology, 67(4), 681–699. https://doi.org/10.1093/sysbio/syy006 cano, M., Minkowski, K., Manuel, M., Ponce, P., Sigler, L.,
Thorbjarnarson, J. (1989). Ecology of the American crocodile, Croc- Soberon, R., Trelancia, A. M., & Velasco, A. (2006). Regional
odylus acutus. In: Hall, P. & Bryant, R. (Eds.), Crocodiles, their habitat conservation priorities for the American crocodile. Bio-
ecology, management and conservation a special publication of the logical Conservation, 128(1), 25–36. https://doi.org/10.1016/j.
crocodile specialist group (pp. 228–258). IUCN Publications. biocon.2005.09.013
Downloaded from https://academic.oup.com/evolut/article/77/1/329/6873140 by guest on 28 January 2024You can also read

















































