Human impacts on two endemic cassowary species in Indonesian New Guinea
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Human impacts on two endemic cassowary species in
Indonesian New Guinea
J E D E D I A H F . B R O D I E and M A R G A R E T H A P A N G A U - A D A M
Abstract Cassowaries are important seed dispersers in reproductive traits in some plant taxa (Pannell, ;
tropical rainforests of New Guinea, but little is known Hall & Walter, ), and cassowaries were estimated to
about their population ecology or their responses to be the most important vertebrate disperser of plant
human disturbance. We used camera traps to measure the seeds at a floristically diverse site in New Guinea (Mack
occurrence, local abundance, and activity patterns of nor- & Wright, ).
thern cassowaries Casuarius unappendiculatus in lowland Of the three cassowary species the southern cassowary C.
forests near Nimbokrang, Papua, and dwarf cassowaries casuarius (height .–. m) is native to New Guinea and
Casuarius bennetti in the Arfak Mountains, West Papua. Australia, and the northern C. unappendiculatus (.–. m)
Our goals were to assess human impacts on cassowaries at and dwarf cassowaries C. bennetti (.–. m) are endemic
multiple spatial scales and to measure their activity patterns to New Guinea (heights from Pratt & Beehler, ).
over an elevational divide. At fine spatial scales local abun- Australian populations of the southern cassowary have
dance of cassowaries was strongly reduced in areas fre- been studied using sign surveys (Westcott, ), transects
quented by humans. At larger spatial scales the distance to (Moore, ), radio telemetry (Westcott et al., ) and
the nearest village or drivable road did not affect local abun- global positioning system telemetry (Campbell et al.,
dance but altered the stage structure of the individuals de- ). However, other than a preliminary study on the im-
tected, with a higher proportion of juveniles relative to pacts of logging on northern cassowaries (Pangau-Adam
adults. Local abundance of cassowaries was unrelated to et al., ) almost nothing is known about the population
site usage by introduced pigs. Both populations studied ecology of the species in New Guinea.
were strongly diurnal and their activity patterns were not The northern and southern species are categorized as
significantly different. Efforts to control hunting remain Vulnerable, and the dwarf as Near Threatened on the
critical to sustaining cassowaries and the seed dispersal ser- IUCN Red List (BirdLife International, a,b, ), and
vices they provide. all three are thought to have declining populations
(IUCN, ). This threat is largely attributable to their sus-
Keywords Activity patterns, bird conservation, climate
ceptibility to over-hunting (Johnson et al., ; Sales, )
change, exploitation, hunting, introduced species, Papua,
and, in some areas, habitat loss (Crome & Moore, ;
ratite
Campbell et al., ; Pangau-Adam et al., ).
We studied a population of C. unappendiculatus in low-
land forests and a population of C. bennetti in submontane
forests of Indonesian New Guinea using motion-triggered
Introduction
camera traps. Substantial habitat remains available in
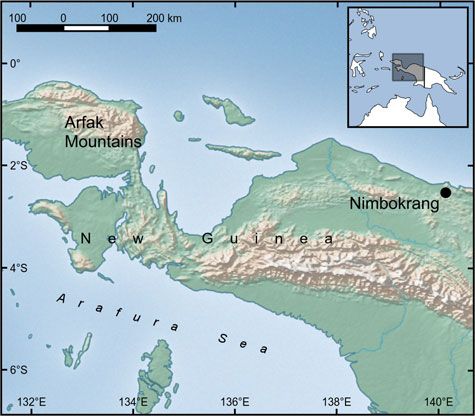
both areas (Miettinen et al., ; Fig. ) but hunting pres-
C assowaries Casuarius spp. are the largest avian frugi-
vores and the largest-bodied seed dispersing animals
in the tropical forests of New Guinea and Australia.
sure was thought to be high, based on proximity to
human settlements and conversations with local hunters
Dispersal of seeds by cassowaries is known to be important (Pangau-Adam & Noske, ). We sought to assess how
for many plant species (Mack, ; Westcott et al., ; both species responded to potential hunting pressure,
Bradford & Westcott, , ; Pangau-Adam & using accessibility (e.g. the distance to the nearest village
Muehlenberg, ) and even for the evolution of or road) as a proxy. We also tried to ascertain whether ac-
cessibility influenced cassowary population structure. In
heavily hunted populations it can become increasingly dif-
JEDEDIAH F. BRODIE* (Corresponding author) Departments of Zoology and ficult for juveniles to reach adulthood or for adults to sur-
Botany, University of British Columbia, Vancouver, Canada
E-mail brodie@biodiversity.ubc.ca vive, and thus the age structure can shift to a higher
MARGARETHA PANGAU-ADAM† Biology Department, Faculty of Natural Sciences
proportion of juveniles (Mills, ; Brodie et al., ).
and Mathematics, Cenderawasih University, Papua, Indonesia Other than hunting, mammal introductions in New
*Also at: Biodiversity Research Centre, University of British Columbia, Guinea are of conservation concern because the region
Vancouver, Canada lacks most placental mammal lineages. Exotic primates
†Also at: Department of Conservation Biology, University of Göttingen,
Germany
near Jayapura, Papua, for example, could have significant
effects on native birds not adapted to these nest predators
Received June . Revision requested July .
Accepted July . First published online November . (Corlett & Primack, ). There were no ungulates in
Oryx, 2017, 51(2), 354–360 © 2015 Fauna & Flora International doi:10.1017/S0030605315001039
Downloaded from https://www.cambridge.org/core. IP address: 46.4.80.155, on 30 Oct 2021 at 05:01:11, subject to the Cambridge Core terms of use, available at https://www.cambridge.org/core/terms.
https://doi.org/10.1017/S0030605315001039Human impacts on endemic cassowaries 355
Methods
Study system and field sampling
In June we deployed Reconyx HC camera-trap
units (Reconyx, Inc., Holmen, USA) at two humid tropical
forest sites: in the lowlands near Nimbokrang, Papua
Province, and in the Arfak Mountains, West Papua
Province (Fig. ). The cameras were retrieved – months
later. The Nimbokrang area has flat, alluvial forest, much
of which was flooded to a depth of a few centimetres at
the time of camera deployment; the area we worked in
was at – m elevation. The forests have not been com-
mercially logged, although illegal selective logging of mer-
bau Intsia bijuga (Fabaceae) is widespread. The northern
cassowary is the only cassowary in this area. In the Arfak
Mountains we worked near the village of Syoubri, at eleva-
tions of ,–, m in lower montane forests (Flannery,
FIG. 1 The study sites in the Arfak Mountains and Nimbokrang, ). The topography is steep; for example, one camera
in Indonesian New Guinea.
transect spanned vertical metres over , linear metres
(a mean slope of %). The forests have a lower canopy than
New Guinea prior to the introduction of pigs (Sus scrofa × S. in Nimbokrang, and denser understorey vegetation. There
celebensis) by humans sometime in the last few thousand has been no commercial logging, and forest clearance for
years (Flannery, ). Pigs breed quickly and are omnivor- small-scale subsistence agriculture is limited to areas near
ous, suggesting that they could possibly compete with casso- villages. The dwarf cassowary is the only cassowary there.
waries for fruit. We conducted a preliminary assessment of Subsistence hunting is common in both areas, and casso-
possible interference competition between these species by waries are preferred prey species (MP-A & JB, pers. obs.).
examining at fine spatial scales whether cassowaries avoided The cameras were arranged along transects radiating
areas frequented by pigs. from two villages and the nearby Korea Road in the
Climate change is another potential threat. Many tropical Nimbokrang area, and two villages in the Arfak
animals are thought to be near their thermal maxima Mountains. Cameras were set c. km apart along each tran-
(Tewksbury et al., ), implying that any near-future sect, usually on small game trails, stream beds, ridge tops, or
rises in temperature could force them to migrate to higher in other areas that animals might use for movement or for-
elevations, possibly leading to biodiversity loss in the low- aging. The distance from the cameras to human habitation
lands (Colwell et al., ). For many tropical bird species, or roads was .– km, with various distance categories ap-
ambient temperatures are already thought to exceed the proximately equally represented. Cameras were attached to
upper critical temperature beyond which they must expend the bases of trees, housed within protective metal cases.
energy to stay cool (Khaliq et al., ). We conducted They operated hours per day, recording colour images
a preliminary assessment of whether C. unappendiculatus during the day and black-and-white at night. Although cas-
in the lowlands were near their thermal maximum by sowaries may be active at any time, foraging is thought to be
examining whether they constrained their daily activity to strictly diurnal (Moore, ). Upon being triggered by mo-
mornings and evenings (i.e. avoiding the heat of the day) tion in front of the receptor, a camera recorded a sequence
relative to the activity patterns of C. bennetti at higher of three images and then did not record again for at least s.
elevations.
Specifically, we predicted that () local abundance of Data analysis
cassowaries would be higher at sites further from human
access and at sites with low usage by humans (predators) To test prediction () we used N-mixture models (Royle,
and pigs (introduced potential competitors), () the age ) to estimate whether the local abundance of cassowar-
structure of cassowary populations would be related to po- ies at specific sites (i.e. camera trap stations) was related to
tential hunting pressure, with a lower proportion of mature the distance from the site to the nearest drivable road or vil-
adults at sites closer to human access, and () northern lage (a metric of potential hunting pressure), and the usage
cassowaries in the lowlands would be less active during of the site by pigs and humans. We conducted these analyses
the hottest portion of the day than dwarf cassowaries in separately for each species and also pooled data on the two
the mountains. species, to increase our limited sample size. We accounted
Oryx, 2017, 51(2), 354–360 © 2015 Fauna & Flora International doi:10.1017/S0030605315001039
Downloaded from https://www.cambridge.org/core. IP address: 46.4.80.155, on 30 Oct 2021 at 05:01:11, subject to the Cambridge Core terms of use, available at https://www.cambridge.org/core/terms.
https://doi.org/10.1017/S0030605315001039356 J. F. Brodie and M. Pangau-Adam
TABLE 1 N-mixture model selection results for all models with ΔAIC , for the northern cassowary Casuarius unappendiculatus.
Top model 2nd model 3rd model 4th model Mean model
ΔAIC 0.00 1.56 1.80 1.96
Akaike weight 0.45 0.21 0.18 0.17
R2 0.39 0.41 0.40 0.39
Local abundance function1
Intercept 0.93 ± 0.42 0.96 ± 0.43 0.95 ± 0.43 0.93 ± 0.43 0.95 ± 0.43
Distance2 0.11 ± 0.24 0.02 ± 0.04
Pig relative abundance index3 −0.04 ± 0.24 −0.01 ± 0.04
Human relative abundance index3 0.14 ± 0.20 0.03 ± 0.04
Detection function1
Intercept −4.01 ± 3.02 −4.24 ± 5.15 −4.19 ± 4.55 −4.26 ± 5.94 −4.17 ± 4.27
Camera hours 5.89 ± 16.06 6.96 ± 27.52 6.70 ± 24.27 7.24 ± 31.78 6.55 ± 22.78
Coefficients ± SE from standardized variables
The linear distance from each camera trap site to the nearest village or drivable road
The number of independent detections per trap-days
for the large number of zero detections in the data, typical To test prediction () we compared the activity patterns
for camera-trapping studies, by pooling data into -day of the two cassowary species using Watson’s two-sample test
sampling intervals and using zero-inflated Poisson distribu- of homogeneity (Jammalamadaka & Sengupta, ). This
tions in the N-mixture models. These models are robust to is a non-parametric test analogous to a Kolmogorov–
variation in sampling effort across sites. We ran models re- Smirnov test but for use with circular (e.g. hours in a day,
presenting all possible combinations of the three local abun- months in a year, compass bearings) rather than linear
dance covariates in combination with no detection variables.
covariates, a time detection covariate (to account for trends
in detectability), and a camera hours detection covariate
(the total number of hours during which a camera operated Results
at each site during each -day sampling interval). All con-
tinuous variables were standardized prior to analysis. We We detected northern cassowaries on of a total of ,
used multi-model inference by averaging parameter coeffi- camera-days at Nimbokrang, and dwarf cassowaries on
cients across all models with ΔAIC , .. To assess site of , camera-days in the Arfak Mountains. The top
usage by humans and pigs we measured a relative abun- N-mixture model for C. unappendiculatus did not have
dance index for each species at each camera station as the any abundance covariates (Table ). The top models for C.
number of independent detections per camera days. bennetti and the two species pooled both had negative
Two detections were considered independent if they were human site usage terms, as did five of the seven models
non-consecutive (i.e. another species was detected at the with ΔAIC , for C. bennetti (Table ) and all four of the
camera station in between) or if they were . minutes models with ΔAIC , for the species pooled (Table ).
apart; these are increasingly standard criteria for analysis Relative usage of each camera site by pigs, and the distance
of camera trap data (e.g. O’Brien et al., ; Brodie & from each station to the nearest road or village were not re-
Giordano, ; Carter et al., ). lated to local abundance of either cassowary species indi-
We tested prediction () using logistic regression of dis- vidually or the two species pooled (Tables –). Camera
tance from road or village (predictor variable) vs casso- hours was an important detection covariate for C. unappen-
wary life stage ( for mature adults, for juveniles), again diculatus and the two species pooled but not for C. bennetti
conducting analyses for the two species separately and (Tables –).
pooled. As such, these were tests of the influence of We obtained independent detections of C. unappendi-
human access on cassowary stage structure, conditional culatus and of C. bennetti; % of C. unappendiculatus
upon detection. The life stages are easily distinguished, detections and % of C. bennetti detections were of mature
with adults having black body feathers with blue and red adults. Chicks were detected only once, when two northern
skin on the neck, whereas juveniles are uniformly brown. cassowary chicks were photographed with an adult in
We did not assess chicks (light coloured with black stripes) Nimbokrang. The proportion of independent detections
in this analysis. We analysed independent detections for that were adults was not significantly related to distance
both species, where independence was determined by the from the nearest road or village for C. unappendiculatus
same criteria used to calculate relative abundance indices (logistic regression, P = .) or C. bennetti (P = .) alone
for humans and pigs. (possibly because of limited sample size), but for the two
Oryx, 2017, 51(2), 354–360 © 2015 Fauna & Flora International doi:10.1017/S0030605315001039
Downloaded from https://www.cambridge.org/core. IP address: 46.4.80.155, on 30 Oct 2021 at 05:01:11, subject to the Cambridge Core terms of use, available at https://www.cambridge.org/core/terms.
https://doi.org/10.1017/S0030605315001039Human impacts on endemic cassowaries 357
TABLE 2 N-mixture model selection results for all models with ΔAIC , for the dwarf cassowary Casuarius bennetti. R values are calcu-
lated relative to the intercept-only model, and therefore that model does not have an R.
Top model 2nd model 3rd model 4th model 5th model 6th model 7th model Mean model
ΔAIC 0.00 0.92 1.34 1.39 1.52 1.92 1.95
Akaike weight 0.26 0.16 0.13 0.13 0.12 0.10 0.10
R2 0.17 0.20 0.20 0.19 0.17 0.05
Local abun-
dance
function1
Intercept −0.07 ± 1.12 0.23 ± 0.77 −1.05 ± 0.92 −0.69 ± 1.12 −0.70 ± 1.12 −0.59 ± 1.16 −0.46 ± 0.44 −0.56 ± 0.97
Distance2 −0.16 ± 0.51 −0.02 ± 0.05
Pig relative 0.28 ± 0.25 0.37 ± 0.26 0.07 ± 0.06
abundance
index3
Human rela- −3.01 ± 2.94 −2.42 ± 2.58 −3.06 ± 2.97 −3.08 ± 2.98 −3.22 ± 3.17 −2.19 ± 2.16
tive abun-
dance index3
Detection
function1
Intercept −2.22 ± 0.46 −2.17 ± 0.42 −2.08 ± 0.30 −2.22 ± 0.46 −2.07 ± 0.50 −2.31 ± 0.58 −2.03 ± 0.29 −2.17 ± 0.43
Camera 0.30 ± 0.48 0.04 ± 0.06
hours
Coefficients ± SE from standardized variables
The linear distance from each camera trap site to the nearest village or drivable road
The number of independent detections per trap-days
TABLE 3 N-mixture model selection results for all models with ΔAIC , for the northern and dwarf cassowaries pooled.
Top model 2nd model 3rd model 4th model Mean model
ΔAIC 0.00 1.80 1.97 1.97
Akaike weight 0.47 0.19 0.17 0.17
R2 0.26 0.27 0.15 0.26
Local abundance function1
Intercept 0.09 ± 0.67 −0.06 ± 0.75 0.09 ± 0.67 0.00 ± 0.83 0.04 ± 0.71
Distance2 0.11 ± 0.25 0.02 ± 0.05
Pig relative abundance index3 0.05 ± 0.28 0.01 ± 0.09
Human relative abundance index3 −2.08 ± 1.58 −1.95 ± 1.57 −2.08 ± 1.58 −2.05 ± 1.59 −2.05 ± 1.58
Detection function1
Intercept −2.67 ± 0.39 −2.59 ± 0.40 −2.61 ± 0.38 −2.63 ± 0.45 −2.64 ± 0.40
Camera hours 0.64 ± 0.52 0.64 ± 0.52 0.64 ± 0.52 0.53 ± 0.43
Coefficients ± SE from standardized variables
The linear distance from each camera trap site to the nearest village or drivable road
The number of independent detections per trap-days
species pooled this relationship was significant (slope = ., ). We assembled multiple lines of evidence related to
P , .; Fig. ). large- and small-scale distribution, local abundance, and
Both species were strictly diurnal in activity patterns (Fig. ) population stage structure of cassowaries rather than relying
and the activity patterns did not differ significantly between the on any single measure. Despite heavy hunting pressure
two species (Watson’s two-sample test statistic = ., critic- (Pangau-Adam & Noske, ) human accessibility was
al value = ., P . .). not strongly related to the local abundance of cassowaries.
This may be because we did not sample sufficiently remote
areas; all of the locations in which we worked could poten-
Discussion tially be heavily hunted. Human accessibility was related to
cassowary population stage structure, with mature adults
The impacts of unreported and unregulated hunting on comprising a significantly larger proportion of detections
cryptic animals can be difficult to assess (Brodie et al., at locations far from villages or drivable roads. It could be
Oryx, 2017, 51(2), 354–360 © 2015 Fauna & Flora International doi:10.1017/S0030605315001039
Downloaded from https://www.cambridge.org/core. IP address: 46.4.80.155, on 30 Oct 2021 at 05:01:11, subject to the Cambridge Core terms of use, available at https://www.cambridge.org/core/terms.
https://doi.org/10.1017/S0030605315001039358 J. F. Brodie and M. Pangau-Adam
FIG. 3 Activity patterns of the northern and dwarf cassowaries.
The patterns for the two species are not significantly different
according to Watson’s two-sample test of homogeneity for
circular predictor variables.
); our results expand upon this by identifying hunting
as another potentially critical threat.
We did not detect a strong influence of pigs on the dis-
tribution of cassowaries at fine spatial scales. This could in-
dicate that introduced pigs are not competing strongly with
cassowaries, at least via interference competition. However,
we note that absence of evidence does not constitute evi-
dence of absence; it remains possible that pigs affect casso-
wary populations by depressing the availability of fruit (i.e.
exploitative competition) or attracting humans, a shared
predator (i.e. apparent competition; sensu Holt, ).
The two cassowary populations did not differ significantly
in their activity patterns, suggesting that lowland northern
cassowaries do not avoid being active during the hottest
parts of the day any more than higher elevation dwarf casso-
waries. There is therefore no evidence that lowland cassowar-
ies are thermally stressed. Lowland populations also retain the
FIG. 2 Logistic trendlines for the life-stage structure of the ability to adjust behaviourally to rising temperatures (by shift-
northern Casuarius unappendiculatus and dwarf cassowaries
ing their activity patterns) and may not be highly susceptible
Casuarius bennetti as a function of human accessibility; dashed
lines represent % confidence intervals.
to modest near-term regional warming. However, we caution
that more information is needed on this topic. Cassowaries
and other lowland species may have abrupt thresholds in
that mature adults remain in the deep forest and exclude ju- their physiological responses to temperature, below which
veniles from their territories, forcing the latter to live closer they display no adverse effects but above which they cannot
to roads, where they are more vulnerable to hunting. On cope (Colwell et al., ).
their own, stage ratios are impossible to interpret in terms Some cassowary populations are known to migrate alti-
of the dynamics of the population (Caughley, ; Mills, tudinally, perhaps in response to shifting patterns of fruit
). However, given that local abundance does not change availability (Wright, ). This could potentially violate
in more inaccessible areas, where more of the individuals are our N-mixture model’s assumption that local abundance
immature, this could be a signal of exploitation. It also ap- was closed during the sampling periods (Royle, ), espe-
pears that, at fine spatial scales (i.e. , c. km), cassowaries cially given our long sampling intervals (up to months at a
avoid areas frequented by humans, which could be evidence few camera stations). However, none of our top models in-
of an impact of hunting on cassowary distribution. Previous cluded time as a detection covariate, suggesting that detect-
work has suggested that logging activities are a prime threat ability did not change significantly over the sampling period
to northern cassowary populations (Pangau-Adam et al., as would have been the case if a substantial portion of the
Oryx, 2017, 51(2), 354–360 © 2015 Fauna & Flora International doi:10.1017/S0030605315001039
Downloaded from https://www.cambridge.org/core. IP address: 46.4.80.155, on 30 Oct 2021 at 05:01:11, subject to the Cambridge Core terms of use, available at https://www.cambridge.org/core/terms.
https://doi.org/10.1017/S0030605315001039Human impacts on endemic cassowaries 359
population migrated into or out of the study areas during B R O D I E , J.F., G I O R D A N O , A.J., Z I P K I N , E.F., B E R N A R D , H.,
this period. M O H D -A Z L A N , J. & A M B U , L. () Correlation and persistence of
hunting and logging impacts on tropical rainforest mammals.
Cassowaries are functionally important as dispersers of
Conservation Biology, , –.
plant seeds (Mack, ; Wright, ; Bradford & B R O D I E , J.F., M U N T I F E R I N G , J., H E A R N , M., L O U T I T , B., L O U T I T , R.,
Westcott, , ). Their loss through overexploitation B R E L L , B. et al. () Population recovery of black rhinoceros in
could therefore have important consequences for forest re- north-west Namibia following poaching. Animal Conservation, ,
generation (McConkey et al., ) and ecosystem function- –.
C A M P B E L L , H.A., D W Y E R , R.G., F I T Z G I B B O N S , S., K L E I N , C.J.,
ing (Brodie & Gibbs, ). Efforts to control hunting and
L A U R I D S E N , G., M C K E O W N , A. et al. () Prioritising the
ensure its sustainability remain critical across New Guinea. protection of habitat utilised by southern cassowaries Casuarius
To this end, in August and November we orga- casuarius johnsonii. Endangered Species Research, , –.
nized interactive educational workshops for researchers, C A R T E R , N.H., S H R E S T H A , B.K., K A R K I , J.B., P R A D H A N , N.M.B. &
university staff and local communities in Nimbokrang on L I U , J. () Coexistence between wildlife and humans at fine
the importance of cassowaries for forest ecosystems and spatial scales. Proceedings of the National Academy of Sciences of the
United States of America, , –.
the sustainability of community-based wildlife hunting. C A U G H L E Y , G. () Interpretation of age ratios. The Journal of
We also employed hunters as local guides during our field- Wildlife Management, , –.
work, hoping to raise awareness about sustainable hunting. C O LW E L L , R.K., B R E H M , G., C A R D E L Ú S , C.L., G I L M A N , A.C. &
Finally, we prepared a list of recommendations concerning L O N G I N O , J.T. () Global warming, elevational range shifts, and
sustainable wildlife hunting, which has been disseminated lowland biotic attrition in the wet tropics. Science, , –.
C O R L E T T , R.T. & P R I M AC K , R.B. () Tropical Rain Forests: An
to local governments in Papua, Indonesia.
Ecological and Biogeographical Comparison. Wiley and Sons,
Chichester, UK.
C R O M E , F.H.J. & M O O R E , L.A. () Cassowaries in north-eastern
Acknowledgements Queensland: report of a survey and a review and assessment of their
status and conservation and management needs. Australian Wildlife
JB was supported by the Natural Sciences and Engineering Research, , –.
F L A N N E R Y , T. () Mammals of New Guinea. Reed Books,
Research Council of Canada and the Canadian Foundation
Chatsworth, Australia.
for Innovation. MP-A was supported by the Dorothea H A L L , J.A. & W A LT E R , G.H. () Relative seed and fruit toxicity of
Schlözer Fellowship Programme, University of Göttingen, the Australian cycads Macrozamia miquelii and Cycas ophiolitica:
Germany. We thank the local communities for permission further evidence for a megafaunal seed dispersal syndrome in cycads,
to conduct this work. We are grateful to Om Dance, Om and its possible antiquity. Journal of Chemical Ecology, , –.
H O LT , R.D. () Predation, apparent competition, and structure of
Augustinus and Om Zeth for assistance with field work.
prey communities. Theoretical Population Biology, , –.
IUCN () IUCN Red List of Threatened Species v. .. Http://
www.iucnredlist.org [accessed July ].
References J A M M A L A M A D A K A , S.R. & S E N G U P T A , A. () Topics in Circular
Statistics, Section .. World Scientific Press, Singapore.
B I R D L I F E I N T E R N AT I O N A L (a) Casuarius unappendiculatus. In J O H N S O N , A., B I N O , R. & I G A G , P. () A preliminary evaluation of
The IUCN Red List of Threatened Species . Http://dx.doi.org/. the sustainability of cassowary (Aves: Casuariidae) capture and
/IUCN.UK.-.RLTS.TA.en [accessed trade in Papua New Guinea. Animal Conservation, , –.
September ]. K H A L I Q , I., H O F , C., P R I N Z I N G E R , R., B O H N I N G -G A E S E , K. &
B I R D L I F E I N T E R N AT I O N A L (b) Casuarius casuarius. In The IUCN P F E N N I N G E R , M. () Global variation in thermal tolerances and
Red List of Threatened Species . Http://dx.doi.org/./ vulnerability of endotherms to climate change. Proceedings of the
IUCN.UK.-.RLTS.TA.en [accessed Royal Society B, , .
September ]. M AC K , A.L. () Distance and nonrandomness of seed dispersal by
B I R D L I F E I N T E R N AT I O N A L () Casuarius bennetti. In The IUCN the dwarf cassowary Casuarius bennetti. Ecography, , –.
Red List of Threatened Species . Http://dx.doi.org/./IUCN. M AC K , A.L. & W R I G H T , D.D. () The frugivore community and
UK.-.RLTS.TA.en [accessed September the fruiting plant flora in a New Guinea rainforest: identifying
]. keystone frugivores. In Tropical Fruits and Frugivores (eds J.L. Dew
B R A D FO R D , M.G. & W E S T C O T T , D.A. () Consequences of & J.P. Boubli), pp. –. Springer, Dordrecht, The Netherlands.
southern cassowary (Casuarius casuarius, L.) gut passage and M C C O N K E Y , K.R., P R A S A D , S., C O R L E T T , R.T., C A M P O S -A R C E I Z , A.,
deposition pattern on the germination of rainforest seeds. Austral B R O D I E , J.F., R O G E R S , H. & S A N TA M A R I A , L. () Seed dispersal
Ecology, , –. in changing landscapes. Biological Conservation, , –.
B R A D FO R D , M.G. & W E S T C O T T , D.A. () Predation of cassowary M I E T T I N E N , J., S H I , C., T A N , W.J. & L I E W , S.C. () land cover
dispersed seeds: is the cassowary an effective disperser? Integrative map of insular Southeast Asia in -m spatial resolution. Remote
Zoology, , –. Sensing Letters, , –.
B R O D I E , J.F. & G I B B S , H.K. () Bushmeat hunting as climate threat. M I L L S , L.S. () Conservation of Wildlife Populations. Blackwell
Science, , –. Publishing, Malden, USA.
B R O D I E , J.F. & G I O R D A N O , A.J. () Density of the Vulnerable M O O R E , L.A. () Population ecology of the southern cassowary
Sunda clouded leopard Neofelis diardi in a protected area in Sabah, Casuarius casuarius johnsonii, Mission Beach north Queensland.
Malaysian Borneo. Oryx, , –. Journal of Ornithology, , –.
Oryx, 2017, 51(2), 354–360 © 2015 Fauna & Flora International doi:10.1017/S0030605315001039
Downloaded from https://www.cambridge.org/core. IP address: 46.4.80.155, on 30 Oct 2021 at 05:01:11, subject to the Cambridge Core terms of use, available at https://www.cambridge.org/core/terms.
https://doi.org/10.1017/S0030605315001039360 J. F. Brodie and M. Pangau-Adam
O’B R I E N , T.G., K I N N A I R D , M.F. & W I B I S O N O , H.T. () T E W K S B U R Y , J.J., H U E Y , R.B. & D E U T S C H , C.A. () Putting the heat
Crouching tigers, hidden prey: Sumatran tiger and prey on tropical animals. Science, , –.
populations in a tropical forest landscape. Animal Conservation, , W E S T C O T T , D.A. () Counting cassowaries: what does cassowary
–. sign reveal about their abundance? Wildlife Research, , –.
P A N G A U -A D A M , M. & M U E H L E N B E R G , M. () Palm species in the W E S T C O T T , D.A., B E N T R U P P E R B Ä U M E R , J., B R A D F O R D , M.G. &
diet of the northern cassowary (Casuarius unappendiculatus) in M C K E OW N , A. () Incorporating patterns of disperser
Jayapura Region, Papua, Indonesia. Palms, , –. behaviour into models of seed dispersal and its effects on estimated
P A N G A U -A D A M , M., M U E H L E N B E R G , M. & W A LT E R T , M. () dispersal curves. Oecologia, , –.
Rainforest disturbance affects population density of the northern W R I G H T , D.D. () Diet, keystone resources and altitudinal
cassowary Casuarius unappendiculatus in Papua, Indonesia. Oryx, movement of dwarf cassowaries in relation to fruiting phenology in
, –. a Papua New Guinean rainforest. In Tropical Fruits and Frugivores
P A N G A U -A D A M , M. & N O S K E , R. () Wildlife hunting and bird (eds J.L. Dew & J.P. Boubli), pp. –. Springer, Dordrecht, The
trade in northern Papua. In Ethno-Ornithology: Global Studies in Netherlands.
Indigenous Ornithology, Culture, Society and Conservation (eds
S. Tidemann, A. Gosler & R. Gosford), pp. –. Earthscan,
London, UK. Biographical sketches
P A N N E L L , C.M. () A new, cassowary-dispersed, species of Aglaia
(Meliaceae, section Amoora) from Papua New Guinea. Kew J E D E D I A H B R O D I E studies the ecology and conservation of large ver-
Bulletin, , –. tebrates, particularly in tropical rainforests. He has focused on how an-
P R A T T , T.K. & B E E H L E R , B.M. () Birds of New Guinea. Princeton thropogenic stressors such as bushmeat hunting, logging and climate
University Press, Princeton, USA. change affect populations directly, and how they disrupt animal–plant
R O Y L E , J.A. () N-mixture models for estimating population size interactions such as seed dispersal. M A R G A R E T H A P A N G A U - A D A M
from spatially replicated counts. Biometrics, , –. is an ecologist with a particular interest in frugivory, seed dispersal
S A L E S , J. () Current conservation status of ratites. Journal of and forest regeneration, community-based wildlife management, and
Threatened Taxa, , –. biodiversity conservation.
Oryx, 2017, 51(2), 354–360 © 2015 Fauna & Flora International doi:10.1017/S0030605315001039
Downloaded from https://www.cambridge.org/core. IP address: 46.4.80.155, on 30 Oct 2021 at 05:01:11, subject to the Cambridge Core terms of use, available at https://www.cambridge.org/core/terms.
https://doi.org/10.1017/S0030605315001039You can also read

















































