Comparative transcriptome analyses of the Drosophila pupal eye
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
2
G3, 2021, 11(1), jkaa003
DOI: 10.1093/g3journal/jkaa003
Advance Access Publication Date: 20 November 2020
Investigation
Comparative transcriptome analyses of the Drosophila
pupal eye
Miles W. DeAngelis, Joseph D. Coolon , and Ruth I. Johnson *
Department of Biology, Wesleyan University, 52 Lawn Avenue, Middletown, CT 06459, USA
Downloaded from https://academic.oup.com/g3journal/article/11/1/jkaa003/5995320 by guest on 28 October 2021
*Corresponding author: Department of Biology, Wesleyan University, 52 Lawn Avenue, Hall-Atwater Room 212, Middletown, CT 06459, USA.
rijohnson@wesleyan.edu
Abstract
Tissue function is dependent on correct cellular organization and behavior. As a result, the identification and study of genes that contrib-
ute to tissue morphogenesis is of paramount importance to the fields of cell and developmental biology. Many of the genes required for
tissue patterning and organization are highly conserved between phyla. This has led to the emergence of several model organisms and
developmental systems that are used to study tissue morphogenesis. One such model is the Drosophila melanogaster pupal eye that has
a highly stereotyped arrangement of cells. In addition, the pupal eye is postmitotic that allows for the study of tissue morphogenesis
independent from any effects of proliferation. While the changes in cell morphology and organization that occur throughout pupal eye
development are well documented, less is known about the corresponding transcriptional changes that choreograph these processes.
To identify these transcriptional changes, we dissected wild-type Canton S pupal eyes and performed RNA-sequencing. Our analyses
identified differential expression of many loci that are documented regulators of pupal eye morphogenesis and contribute to multiple
biological processes including signaling, axon projection, adhesion, and cell survival. We also identified differential expression of genes
not previously implicated in pupal eye morphogenesis such as components of the Toll pathway, several non-classical cadherins, and
components of the muscle sarcomere, which could suggest these loci function as novel patterning factors.
Keywords: pupae; eye; Drosophila; transcriptome; morphogenesis
Introduction rise to the eyes as well as the antennae, ocelli, and surrounding
head epithelium (Haynie and Bryant 1986). Establishment of the
The Drosophila pupal eye is a postmitotic pseudostratified neuroe-
eye disk and the subsequent eye field is dependent on a network
pithelium that is organized into 750 optical units known as
of highly conserved transcription factors known as the Retinal
ommatidia (Figure 1, A–D). Each ommatidium contains eight pho-
Determination Network (Kumar 2010, 2011; Treisman 2013). The
toreceptor neurons (R1–R8), four lens-secreting cone cells, and
major components of this network include Eyeless, Optix,
two pigment-producing primary (1 ) cells (Ready et al. 1976;
Cagan and Ready 1989a; Wolff and Ready 1991a, 1993; Carthew Dachshund, Twin of eyeless, Eyegone, Eyes absent, and Sine ocu-
2007). Surrounding each ommatidium are lattice cells, which also lis (Cheyette et al. 1994; Mardon et al. 1994; Quiring et al. 1994;
produce pigment. By 40 h after puparium formation (APF), the Serikaku and O’Tousa, 1994; Halder et al. 1995; Czerny et al. 1999;
pupal eye has achieved its stereotypical honeycomb organization Seimiya and Gehring 2000; Jang et al. 2003). At the beginning of
and lattice cells can be classified as either secondary (2 ) or ter- the third larval instar, a combination of Hedgehog (Hh) (Wolff
tiary (3 ) cells depending on the number of contacts they form and Ready 1991a; Ma et al. 1993; Ma and Moses 1995; Treisman
with adjacent ommatidia. Those classified as 2 cells have an and Rubin 1995; Greenwood and Struhl 1999; Curtiss and Mlodzik
elongated rectangular apical surface area that contacts two adja- 2000; Kenyon et al. 2003) and Ecdysone (Bate and Martı́nez-Arias
cent ommatidia, while the apical surface area of 3 cells is more 1993; Thummel 1996; Niwa et al. 2004) signaling leads to the initi-
hexagonal in shape and contacts three adjacent ommatidia ation and progression of the morphogenetic furrow. The furrow,
(Figure 1C). In addition, each eye contains 600 sensory bristle which is caused by ingression of the apical surface area of col-
groups that are present at the anterior vertex of each ommatid- umns of retinal progenitor cells, is initiated from the posterior of
ium with the exception of those along the edges of the eye the eye disk and progresses toward the anterior over the course
(Wigglesworth 1953; Waddington and Perry 1960; Cagan and of several hours. This progression leads to the formation a devel-
Ready 1989a). opmental gradient that persists throughout most of eye morpho-
During embryogenesis, the first cells are selected that will go genesis (Wolff and Ready 1991a).
on to form the eyes (Honn et al. 2016). These retinal progenitor Cell differentiation in the eye is stepwise, beginning during
cells proliferate to form the eye-antennal imaginal disks that give passage of the morphogenetic furrow with the specification of
Received: August 18, 2020. Accepted: October 08, 2020
C The Author(s) 2020. Published by Oxford University Press on behalf of Genetics Society of America.
V
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0/), which
permits unrestricted reuse, distribution, and reproduction in any medium, provided the original work is properly cited.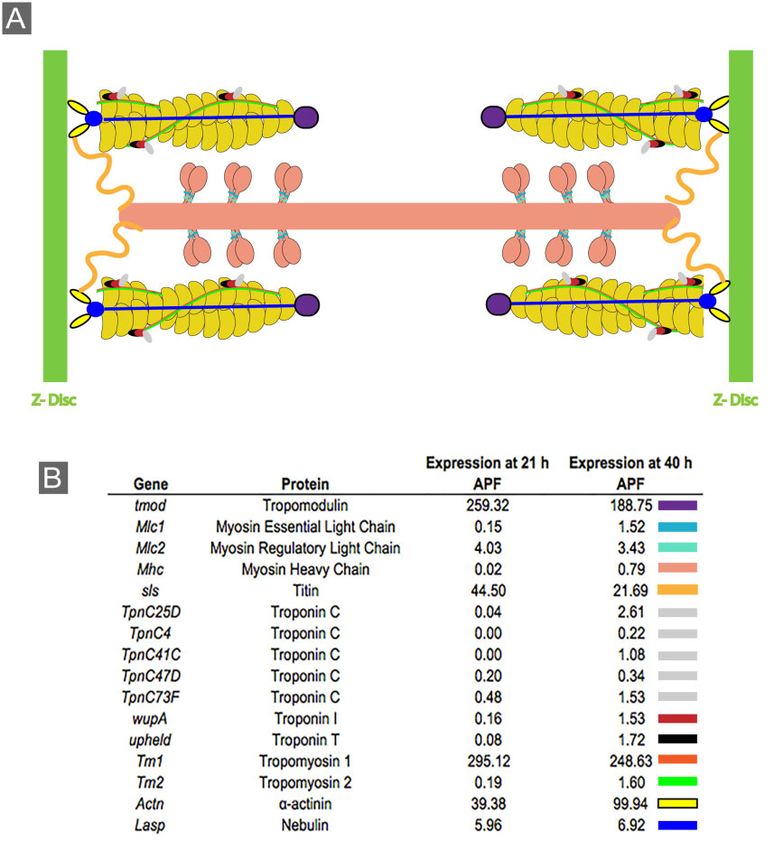
2 | G3, 2021, Vol. 11, No. 1
Downloaded from https://academic.oup.com/g3journal/article/11/1/jkaa003/5995320 by guest on 28 October 2021
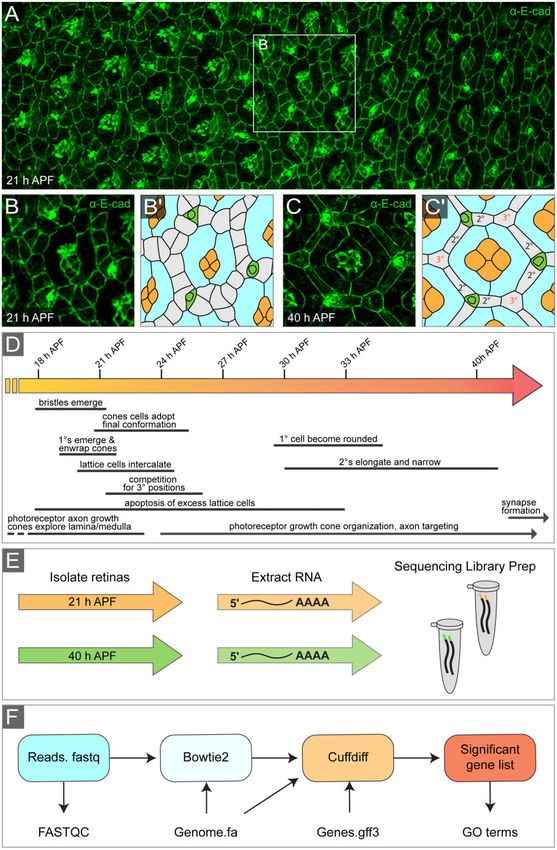
Figure 1 The Drosophila pupal eye and experimental design. (A) Mid-region of a pupal eye at 21 h APF, characterized by a developmental gradient.
Younger, anterior (less organized) tissue to the left and older (posterior) tissue is to the right. White box encloses a single ommatidium shown at higher
resolution in (B) with tracing (B0 ). (C) Representative ommatidium at 40 h APF and tracing (C0 ), with cone cells colored orange, 1 cells in blue, bristle
groups in green, and lattice cells in gray with 2 and 3 cells indicated. (D) Timeline of development of the pupal eye. Major morphogenetic events that
occur between 18 and 40 h APF are indicated. Times relate to development at 25 C. (E) Experimental workflow: eyes were dissected at 21 or 40 h APF,
total RNA extracted, and sequencing libraries prepared (cDNA synthesis and bar-coding). (F) Bioinformatics pipeline used to identify differentially
expressed genes and identify enriched gene ontology terms from raw sequencing reads.
M. W. DeAngelis et al. | 3
the eight photoreceptors (Reinke and Zipursky 1988; Van Vactor Yorkie activities, a lattice cell’s proximity to 1 cells, and the bal-
et al. 1991; Jarman et al. 1994, 1995; Fanto and Mlodzik 1999; ance of death and survival signals that a cell receives (Rusconi
Frankfort et al. 2001; Pepple et al. 2008). Shortly after their recruit- et al. 2000; Monserrate and Brachmann 2007; DeAngelis et al.
ment, photoreceptors begin to project their axons to the lamina 2020).
or the medulla (Gibbs and Truman 1998; Clandinin and Zipursky While the morphological events that occur during pupal eye
2002; Pepple et al. 2008). Axons will reach their target brain development have been well documented, little is known about
regions at 24 h APF and begin to form synapses with target neu- the corresponding transcriptional changes that facilitate and
rons at 50 h APF (Gibbs and Truman 1998). Over the course of choreograph them. Previous transcriptome studies of Drosophila
pupal development, photoreceptors undergo substantial mor- eyes have focused on the larval eye disk (Ikmi et al. 2014; Potier
phological changes. Beginning at 48 h APF, the apical poles of et al. 2014; Torres-Oliva et al. 2018), adult eye (Hall et al. 2017), or
each photoreceptor fold 90 toward one another. The central re- analyzed transcriptional changes throughout the entire pupal
gion of this apical pole will develop into the rhabdomere, which head (Ranade et al. 2008). In this study, we compare the transcrip-
contains rhodopsin-rich microvilli (Knust 2007). Recruitment of tomes of pupal eyes at 21 and 40 h APF using RNA-sequencing
Downloaded from https://academic.oup.com/g3journal/article/11/1/jkaa003/5995320 by guest on 28 October 2021
cone and 1 cells, as well as bristle groups, occurs after photore- (RNA-seq) to capture differences in the expression of genes asso-
ceptor specification. Photoreceptors initiate cone cell recruitment ciated with adhesion, cell death, axon projection, and the signal-
through a combination of epidermal growth factor receptor ing pathways that regulate these developmental processes. Our
(EGFR) and Notch signaling (Tomlinson and Ready 1987; Cagan analysis identified large-scale transcriptional differences be-
and Ready 1989b; Zak and Shilo 1992; Freeman 1996, 1997). Over tween the two developmental ages. Some of the differentially
the course of pupal eye development cone cell contacts become expressed genes we identified have previously been established
highly stereotyped, facilitated by the expression of N-cadherin as regulators of pupal eye morphogenesis, or have been impli-
(Hayashi and Carthew 2004). After cone cell specification, EGF se- cated in signal transduction, axon projection, adhesion, or cell
creted by photoreceptors initiates the expression of the Notch li- survival in the eye or other tissues. In addition, we identify many
gand Delta on the surface of cone cells (Nagaraj and Banerjee novel genes not yet associated with eye development. These in-
2007). The interaction between Delta and the Notch receptor, cluded members of the Toll signaling pathway, several non-
expressed on the surface of retinal progenitors, leads to the speci- classical cadherins, and genes associated with muscle structure
fication of two cells that become 1 cells (Cagan and Ready and development. We anticipate that the transcriptome data pre-
1989b; Nagaraj and Banerjee 2007). Once selected, the 1 cells sented here will be a valuable resource for Drosophila pupal eye
gradually enwrap the four cone cells. Formation of the bristle biologists and the broader morphogenesis field.
groups occurs at 18 h APF when two cells are selected to un-
dergo a final round of mitosis, giving rise to the four cells that
comprise each group (Cagan and Ready 1989a; Meserve and Materials and methods
Duronio 2017). This will be the final mitotic division to take place Fly stocks
during Drosophila eye development with all remaining retinal pro- Drosophila melanogaster Canton S cultures were maintained at 25 C
genitor cells adopting the lattice cell fate. on nutrient-rich medium.
Pupal eye development is dependent on the precise regulation
of adhesion between cells. In the eye, adhesion is directed by E- Immunofluorescence
cadherin (Grzeschik and Knust 2005; Larson et al. 2008; Seppa Pupal eyes were dissected and fixed as previously described
et al. 2008; Zaessinger et al. 2015) and the immunoglobulin (Ig) do- (DeAngelis and Johnson 2019). For 1 antibody staining, rat anti-
main adhesion molecules Roughest (Rst), Kin of Irre (Kirre), Hibris E-cad (1:20, DSHB, # 528120) was used to visualize cell bound-
R
(Hbs), and Sticks and Stones (Sns) (Gorski et al. 2000; Araujo et al. aries. Secondary antibodies conjugated to Alexa FluorV 488
2003; Bao and Cagan 2005; Grzeschik and Knust 2005; Mirkovic (Jackson ImmunoResearch) were used at 1:200. Dissected pupal
and Mlodzik 2006; Grillo-Hill and Wolff 2009; Bao et al. 2010). The eyes were imaged with a Leica DM5500 B fluorescence micro-
expression and interaction of Ig domain adhesion molecules is scope and corresponding software.
complimentary. Rst and Kirre are expressed in lattice cells, while
Hbs and Sns are expressed in 1 cells (Gorski et al. 2000; Bao and RNA-seq and bioinformatics analysis
Cagan 2005; Bao et al. 2010). In the eye, Rst interacts with Hbs, Between 50 and 70 eyes were dissected from 21 and 40 h APF
while Sns interacts with Kirre and these interactions are required Canton S pupae at the same time each day, and total RNA was
for cell sorting and maintaining preferential adhesion between 1 extracted from three biological replicates using the ReliaPrep
and lattice cells within the developing eye (Bao and Cagan 2005; RNA Tissue Miniprep System (Promega Corporation, Cat # M3001)
Grillo-Hill and Wolff 2009; Bao et al. 2010). Precise regulation of as described (DeAngelis and Johnson 2019). Barcoded cDNA li-
adhesion is also required for intercalation that organizes lattice brary prep was performed using the TruSeq library preparation
cells into a single row surrounding each ommatidium. kit, libraries were pooled, balanced pooling was confirmed using
Intercalation occurs between 18 and 27 h APF and is dependent qPCR, and 51-bp paired-end sequencing was performed by the
on interactions between Rst and Hbs, as well as interactions be- University of Michigan Sequencing Core Facility as described
tween Cindr and the ArfGAPs that regulate Arf6 (Johnson et al. (DeAngelis et al. 2020). All raw sequencing reads were imported
2008, 2011; Larson et al. 2008). into Galaxy (https://usegalaxy.org/). Quality control of sequence
Each pupal eye is equipped with more lattice cells than needed read outputs was performed using FASTQC (Andrews, 2010;
for eventual formation of the honeycomb lattice leading to the Afgan et al. 2018). The percentage of mapped reads was calcu-
apoptosis of approximately one-third of lattice cells between 18 lated using FlagStat (Li et al. 2009). Sequence reads were aligned
and 33 h APF. Lattice cell removal is directed by a combination of to the D. melanogaster reference genome available from Ensembl
Wingless, JNK, and Notch signaling (Cagan and Ready 1989b; (Zerbino et al. 2018) at the time of analysis: reference genome:
Wolff and Ready 1991b; Cordero et al. 2004; Bushnell et al. 2018). Drosophila_melanogaster.BDGP6.dna.toplevel.fa and gene anno-
Survival of individual lattice cells is dependent on EGFR and tation: Drosophila_melanogaster.BDGP6.93.gff3 (Zerbino et al.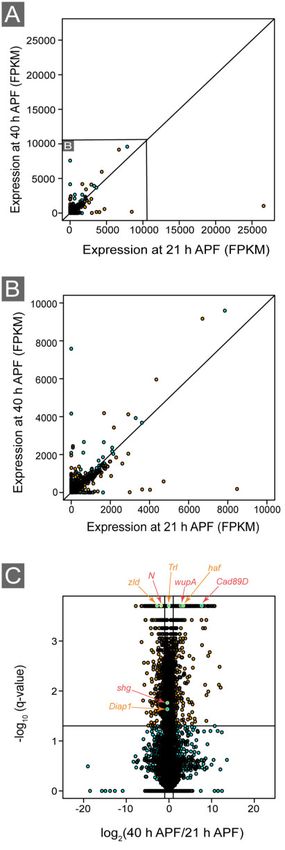
4 | G3, 2021, Vol. 11, No. 1
2018) using bowtie2 with default parameters (Langmead and
Salzberg 2012). Gene expression quantification along with corre-
sponding statistical analyses were performed using Cuffdiff.
Cuffdiff parameters included geometric normalization and tran-
script length correction where a bias length correction using the
reference genome was performed (Trapnell et al. 2012).
Significantly differentially expressed genes were identified as
those with false discovery rate corrected P-values (q-values) lower
than our predetermined threshold (q < 0.05). Gene ontology (GO)
analyses were performed using the Gene Ontology Consortium
(http://geneontology.org/). Volcano and scatter plots were cre-
ated with R-statistical software (Figure 2) (CRAN 2018).
Candidate regulatory transcription factors and transcription
Downloaded from https://academic.oup.com/g3journal/article/11/1/jkaa003/5995320 by guest on 28 October 2021
factor-binding motifs were identified with i-Cis Target analysis
(https://gbiomed.kuleuven.be/apps/lcb/i-cisTarget/index.php)
(Herrmann et al. 2012) and Analysis of Motif Enrichment (AME)
(http://meme-suite.org/tools/ame) (McLeay and Bailey 2010) us-
ing lists of candidate genes and default parameters. Prior to AME
analyses, the extended gene region for each significantly differ-
entially expressed gene was downloaded from FlyBase (https://fly
base.org/) using batch download (https://flybase.org/download/se
quence/batch/).
Data availability statement
Raw RNA-seq output files generated in this work are deposited
under accession number GSE160441 in Gene Expression
Omnibus.
Supplementary material is available at figshare DOI: https://
doi.org/10.25387/g3.12824789.
Results and discussion
Mapped reads and GO
To identify transcriptional changes during pupal eye morphogen-
esis, we dissected Canton S eyes at 21 and 40 h APF (Figure 1, A–C)
(Materials and methods). We chose to analyze differences in gene
expression at 21 h APF as the eye is in the midst of critical pat-
terning and developmental events (Figure 1D). These include 1
cell recruitment, lattice cell intercalation, the conclusion of axon
outgrowth into the brain, and the beginning of elimination of ex-
cess lattice cells by apoptosis. We selected 40 h APF for a compar-
ison point as apoptosis has ceased, the honeycomb lattice is
established, and axon growth cones are approaching their future
synaptic targets. After dissection, total RNA was extracted and
sequenced (Figure 1E) (Materials and methods). A total of 2.7
108 sequence reads were generated with a range of 40,214,252–
44,584,127 mapped reads per sample (92.65–93.42% mapped to
the reference genome) indicating appropriate read depth for
analysis (Table 1). Sequence reads were then subjected to an
established bioinformatics pipeline to identify differentially
expressed genes (Figure 1F) (Materials and methods) (Lanno et al.
2017). Our analyses identified 4636 loci in which expression was
significantly modified between 21 and 40 h APF (q-valueM. W. DeAngelis et al. | 5
Table 1 Total number of reads and mapped reads per sequencing
library This table has not been correctly formatted when
converted from the file submitted. There are two Rep 1 entries
under the heading 21 h APF. The second should be under the
heading 40 h APF instead.
Total reads Mapped reads % mapped
21 h APF
Rep1 44,711,158 41,768,811 93.42
Rep2 43,337,942 40,376,753 93.17
Rep3 43,677,528 40,768,017 93.34
Rep1 (this should be 48,119,480 44,584,127 92.65
underthe 40 h APF
40 h APF
Rep2 46,431,166 43,315,962 93.29
Downloaded from https://academic.oup.com/g3journal/article/11/1/jkaa003/5995320 by guest on 28 October 2021
Rep3 43,313,444 40,214,252 92.84
were most significant for upregulated genes included nucleic
acid metabolic process (GO:0090304), localization (GO:0051179),
transport (GO:0006810), and establishment of localization
(GO:0051234) (Figure 3B and Supplementary Table S3). For down-
regulated genes, the most significant GO terms included regula-
tion of biological process (GO:0050789), regulation of gene
expression (GO:0010468), macromolecule metabolic process
(GO:0060255), and regulation of metabolic process (GO:0019222)
(Figure 3C and Supplementary Table S4).
Differential expression of signaling pathway
components between 21 and 40 h APF
Since development of the pupal eye is dependent on signal trans-
duction, we assessed whether expression of core signaling pro-
teins differed at 21 and 40 h APF. Our analyses identified
differential expression of multiple loci broadly associated with
signaling (GO:0023052) (Supplementary Table S5). These loci
encompassed core components of several signaling pathways
with established roles during Drosophila pupal eye development
including the Notch, EGFR, Decapentaplegic (Dpp)/TGFb, and
Planar Cell Polarity pathways and also identified signaling path-
ways that have not been interrogated for their role in fly eye de-
velopment (Supplementary Table S5).
Notch signaling is vital during early pupal eye development
for photoreceptor, cone and 1 cell specification, patterning, and
inducing cell death (Cagan and Ready 1989b; Fortini et al. 1993;
Cordero et al. 2004; Grzeschik and Knust 2005; Nagaraj and
Banerjee 2007; Bao et al. 2010; Nagel and Preiss 2011; Xiong et al.
2013; Bao 2014). At 40 h relative to 21 h APF, we observed a reduc-
tion in the expression of the Notch receptor (N) and its ligand Delta
(Dl) (Figure 2C) (Vässin and Campos-Ortega 1987; Rebay et al. 1991;
Fleming et al. 1997) but an increase in the ligands Serrate (Rebay
et al. 1991) and weary (wry) (Kim et al. 2010). Expression of the
Notch regulators O-fucosyltransferase 1 (O-fut1) (Sasamura et al.
2003) and Notchless (Nle) (Royet et al. 1998) decreased by 40 h APF,
as did mind bomb 1 (mib1) (Lai et al. 2005) and the DNA-binding tran-
scription factor Suppressor of Hairless (Su(H)) (Gho et al. 1996) that
are also required for N signaling. Consistent with reduced Notch
pathway activity at 40 h APF, we observed a significant reduction
in the expression of 10 enhancers of split (E(spl)) genes that are tran-
scriptional targets of Notch signaling (Schrons et al. 1992; Jennings
et al. 1994) (Supplementary Table S1). Figure 3 Graphical representation of significant GO terms. The 30 most-
significant GO terms (A) when comparing all significant gene expression
The EGFR pathway is required for cell differentiation, adhe-
changes from 21 and 40 h APF, (B) loci with increased expression at 40 h
sion, and survival during early pupal eye development relative to 21 h APF, (C) loci with decreased expression at 40 h relative to
(Wasserman et al. 2000; Freeman and Bienz 2001; Brown and 21 h APF.6 | G3, 2021, Vol. 11, No. 1
Freeman 2003; Protzer et al. 2008; Martı́n-Bermudo et al. 2015; Differential expression of Ecdysone pathway
Malartre 2016). We therefore reasoned that, similar to compo- components and response elements
nents of the Notch pathway, we would observe reduced expres- Ecdysone is a steroid hormone required for insect metamorphosis
sion of EGFR pathway components at 40 h relative to 21 h APF as that facilitates the transition from the larval to pupal stage
differentiation, apoptosis, and patterning have concluded. (Becker 1959; Ashburner et al. 1974; Bate and Martı́nez-Arias 1993;
Consistent with this prediction, we detected significant reduction Thummel 1996). During pupal development, ecdysone levels
in the expression of several canonical Receptor Tyrosine Kinase peak at 20 h APF and progressively drop until adults eclose
(RTK) components including Egfr (Schejter and Shilo 1989), the (Thummel 2001). In addition, our analyses identified increased
adaptor proteins downstream of receptor kinase (drk), and SHC adap- expression of abrupt (ab) at 40 h relative to 21 h APF, which nega-
tor protein (Shc) (Stern et al. 1993; Lai et al. 1995), the GEF Son of sev- tively regulates the transcription of ecdysone target genes (Jang
enless (Sos) (Rogge et al. 1991), the small GTPase Ras oncogene at et al. 2009) (Supplementary Table S1). We therefore reasoned that
85D (Ras85D) (Neuman-Silberberg et al. 1984), the kinases Raf onco- expression of documented Ecdysone transcriptional target genes
Downloaded from https://academic.oup.com/g3journal/article/11/1/jkaa003/5995320 by guest on 28 October 2021
gene (Raf) (Mark et al. 1987) and Downstream of raf1 (Dsor1) (Lu et al. would be reduced at 40 h APF and this was indeed the case for
1993), and the transcriptional activator pointed (pnt) (Scholz et al. Edg91 (Apple and Fristrom 1991), Eip71CD (Savakis et al. 1980),
1993) (Supplementary Table S5). While the expression of Egfr and Eip63E (Stowers et al. 2000), ftz-f1 (Woodard et al. 1994), ImpE2 and
many core RTK signaling components declined at 40 h APF, ex- ImpL2 (Natzle et al. 1988), and Pep (Möritz et al. 1984)
pression of several other RTKs and their ligands increased (Supplementary Table S6). However, the expression of other ec-
(Supplementary Table S5) suggestive of a continuous require- dysone response element genes increased, including Eip93F
ment for RTK-components in the eye. These included PDFG and (Baehrecke and Thummel 1995), Eip74EF (Burtis et al. 1990),
VEGF receptor related (Pvr) (Heino et al. 2001) along with its ligands Eip78C (Stone and Thummel 1993), and Eip75B (Segraves and
PDFG and VEGF receptor-related factor 1 (Pvf1) (Duchek et al. 2001), Hogness 1984) (Supplementary Table S6). Since ecdysone signal-
Pvf2 (Munier et al. 2002), Pvf3 (Cho et al. 2002), and both branchless ing contributes to apoptosis in other Drosophila pupal tissues in-
(bnl) (Sutherland et al. 1996) and heartless (htl) (Beiman et al. 1996), cluding the salivary gland, midgut, and muscle (Denton and
which encode a fibroblast growth factor and fibroblast growth Kumar 2015; Nicolson et al. 2015; Xu et al. 2019), it is possible that
factor receptor (FGFR), respectively. Since FGFR signaling regu- ecdysone response genes identified in our analyses of 21 h APF
lates E-cadherin, crumbs, and actin expression in the developing eyes contribute to or even initiate the apoptosis of lattice cells,
Drosophila pupal eye, increased expression of bnl and htl could which begins at around 18–20 h APF, correlating with increased
contribute to the maintenance of adhesion during later develop- ecdysone (Thummel 2001).
mental stages (Mukherjee et al. 2012). In addition, these results
Gene expression changes associated with axon
could be suggestive of a prominent role for RTK’s such as pvr to
projection
mediate cell survival during the later stages of pupal eye develop-
Growth of photoreceptor axons begins during larval development
ment as is observed in blood and neural progenitor cells
and continues until axons reach the lamina or medulla at 24 h
(Brückner et al. 2004; Read 2018).
APF (Gibbs and Truman 1998). Axons then extend exploratory
Our data also revealed differential expression of signaling pro-
growth cones (from 27 h APF) so that synaptogenesis can begin
teins, which have not yet been implicated as regulators of pupal
from 50 h APF (Gibbs and Truman 1998; Clandinin and Zipursky
eye development. For example, we found that core components
2002). Accordingly, we identified differential expression of loci as-
of the Toll signaling pathway (Supplementary Table S5) were dif-
sociated with axon elongation and extension, growth cone forma-
ferentially expressed between 21 and 40 h APF including the se-
tion, synaptogenesis, and neuronal signaling (Supplementary
creted protease Spatzle-Processing Enzyme (SPE) (Jang et al. 2006),
Table S7). These included hattifattener (haf), which increased in
the spatzle cleaving enzyme easter (ea) (Chasan and Anderson
expression at 40 h relative to 21 h APF (Supplementary Table S7
1989), the receptors Toll (Tl) (Anderson et al. 1985), Tollo (Seppo
and Figure 2C). The haf locus encodes a protein required for axon
et al. 1999), Toll-6, Toll-7 (Tauszig et al. 2000), and 18 wheeler (18w)
targeting and increased expression is indicative of a maturing
(Williams et al. 1997), the scaffolding protein Myd88 (Horng and
neural system (Kurusu et al. 2008). Expression of several cell ad-
Medzhitov 2001), and the transcription factor dorsal (dl) (Lemaitre hesion molecules also increased at 40 h APF. These included the
et al. 1995). The Toll pathway has been widely studied in the con- neural-cadherins CadN (Iwai et al. 1997) and CadN2 (Yonekura
text of the immune response (Valanne et al. 2011; Satoh and et al. 2007), Turtle (Tutl) (Al-Anzi and Wyman 2009), and 10 differ-
Akira 2017; Vidya et al. 2018); however, it was first identified as a ent BEAT-family genes (Pipes et al. 2001). Collectively, these adhe-
regulator of dorsal ventral patterning of the embryo (Anderson sion molecules are required for axon projection and synapse
et al. 1985; Morisato and Anderson 1994). Toll signaling has also formation (Longley and Ready 1995; Iwai et al. 1997; Knust 2007)
been implicated during wound healing of the Drosophila embry- and increased expression of these loci is consistent with axon ex-
onic epidermis (Carvalho et al. 2014; Capilla et al. 2017) and for ep- tension.
ithelial integrity in the Drosophila salivary gland (Kolesnikov and Growth cone extension is an integral part of axon projection
Beckendorf 2007). In addition, the Toll receptors Toll-2 (encoded and is dependent on dynamic cytoskeletal rearrangements.
by 18w), Toll-6 (encoded by Toll-6) and Toll-8 (encoded by tollo) Accordingly, we found differential expression of genes known to
form transheterophillic complexes that facilitate cell intercala- be required for actin remodeling at 40 h relative to 21 h APF con-
tion in the embryonic epithelium by stimulating actin reorganiza- sistent with the actin architectural rearrangements necessary for
tion (Paré et al. 2014). Expression of each of these three toll growth cone extension (Supplementary Table S7). For example,
receptors increased at 40 h APF relative to 21 h APF we identified increased transcription of Capulet (Capt), which
(Supplementary Table S5) and studies that assess their role dur- inhibits actin filament growth (Wills et al. 2002), enabled (ena),
ing pupal eye morphogenesis are an interesting avenue of future which stimulates the addition of actin monomers and is required
research. for axon elongation (Wills et al. 1999; Bashaw et al. 2000), andM. W. DeAngelis et al. | 7
cherrio (cher), a filament protein that crosslinks actin filaments development as they do in the larval eye and oocyte. In particu-
(Sokol and Cooley 1999). We found decreased expression of chick- lar, our results showed a large increase in the expression of
adee (chic), which encodes profilin (Cooley et al. 1992), jitterbug Cad89D at 40 h relative to 21 h APF (log2-fold change ¼ 9.36), sug-
(jbug), which encodes an actin cross-linking protein (Oliva et al. gestive of a prominent role for this cadherin during later stages of
2015), and cdc42, which activates the WASP complex (Luo et al. pupal eye development. Predictive bioinformatics analyses sug-
1994). In addition to the differential expression of loci associated gest that it binds to the members of the myosin family (Gaudet
with actin regulation, we identified increased expression of loci et al. 2011) and it will be interesting for future studies to identify
associated with microtubules that promote axon elongation and its specific functions along with those of other non-classical cad-
maturation. These included Tau (Bolkan and Kretzschmar 2014), herins in the developing pupal eye.
which crosslinks microtubules in the axon, and Kinesin-like protein
at 64D (Klp64D), a motor protein associated with axonal transport Differential expression of cell survival genes
(Berger et al. 2008). Taken together, these data are consistent with Programmed cell death is a critical aspect of pupal eye morpho-
the dynamic cytoskeletal rearrangements consistent with growth genesis that occurs between 18 and 33 h APF and leads to the
Downloaded from https://academic.oup.com/g3journal/article/11/1/jkaa003/5995320 by guest on 28 October 2021
cone extension. culling of roughly one in three lattice cells (Miller and Cagan
1998; Rusconi et al. 2000; Cordero et al. 2004; Lin et al. 2004;
Differential expression in cell adhesion regulators Mendes et al. 2006; Monserrate and Brachmann 2007; Verghese
during pupal eye development et al. 2012; Denton and Kumar 2015; Bushnell et al. 2018;
Modulations in adhesion are vital as cells undergo complex shape DeAngelis et al. 2020). We therefore reasoned that we would ob-
and positional changes during morphogenesis (Collinet and serve decreased expression of established initiators or mediators
Lecuit 2013; Guillot and Lecuit 2013; Martin and Goldstein 2014; of apoptosis at 40 h relative to 21 h APF and indeed this was ob-
Takeichi 2014; McFaul and Fernandez-Gonzalez 2017; Pannekoek served for klumpfuss (klu) (Rusconi et al. 2004), head involution defec-
et al. 2019). Accordingly, we found significant changes in the ex- tive (hid) (Grether et al. 1995), reaper (rpr) (White et al. 1994), Death
pression of many cell adhesion loci (Supplementary Table S8). Regulator Nedd2-like caspase (Dronc) (Dorstyn et al. 1999), and Death
For example, at 40 h APF, we detected decreased expression of related ICE-like caspase (Drice) (Fraser and Evan 1997)
the Ig domain adhesion molecules rst (Ramos et al. 1993), hbs (Supplementary Table S9). Our analyses also identified a signifi-
(Dworak et al. 2001), and sns (Bour et al. 2000), which are required cant decrease in the expression of Diap1 (Supplementary Table
for pigment cell morphogenesis (Araujo et al. 2003; Bao and S9 and Figure 2C), an antagonist of apoptosis (Hay et al. 1995) that
Cagan 2005; Grzeschik and Knust 2005; Grillo-Hill and Wolff may be less critical at 40 h APF since apoptosis has abated and
2009; Bao et al. 2010) and photoreceptor axon guidance the transcription of known apoptotic inducing factors has de-
(Schneider et al. 1995; Sugie et al. 2010), although we did not iden- creased.
tify a significant change in the expression of the Sns ligand Kirre
(Strünkelnberg et al. 2001) (log2-fold change ¼ 0.05). Since Notch Genes associated with muscle development and
signaling regulates the expression of hbs (Bao 2014), the reduced the sarcomere are required for pupal eye
Notch activity at 40 h APF may explain the reduction in its ex- patterning
pression (Supplementary Tables S1, S5, and S8). We also identi- Our RNA-seq analyses identified changes in the expression of
fied decreased expression of shg (encodes E-cad) by 40 h APF many genes implicated in various aspects of muscle development
(Figure 2C) (Tepass et al. 1996), which is required for adhesion in or structure (Supplementary Table S10). These included rst, hbs,
the developing pupal eye (Grzeschik and Knust 2005; Larson et al. and sns, which are required for myoblast fusion during muscle
2008; Seppa et al. 2008; Zaessinger et al. 2015). Taken together, re- development (Bour et al. 2000; Artero et al. 2001; Strünkelnberg
duced expression of these adhesion molecules at 40 h APF may et al. 2001), and Klarsicht (Klar), which is required for nuclei posi-
reflect a lower requirement once the honeycomb lattice is estab- tioning in muscle cells (Elhanany-Tamir et al. 2012) and also has
lished. a documented role in photoreceptor morphogenesis (Mosley-
Our analyses also highlighted changes in the expression of Bishop et al. 1999; Patterson et al. 2004). Particularly, striking was
many non-classical cadherins. These included fat (ft) (Mahoney the component of the muscle sarcomere, which facilitates mus-
et al. 1991), dachsous (ds) (Clark et al. 1995), Calsyntenin-1 (cals) (Vogt cle contraction, that were expressed in the developing pupal eye
et al. 2001), and Cad89D, Cad87A, Cad74A, Cad96Ca, Cad96Cb, (Figure 4). Each sarcomere is composed of actin and myosin bun-
Cad99C, Cad88C, and Cad86C (Hynes and Zhao 2000; Tepass et al. dles along with regulatory proteins such as the troponins and tro-
2000; Hill et al. 2001) (Supplementary Table S8 and Figure 2C). pomyosins, which mediate the interactions between actin and
While the developmental roles for many of these non-classical myosin bundles (Henderson et al. 2011; Sweeney and Hammers
cadherins have not been well studied, others have established 2018; Mukund and Subramaniam 2019). Our analyses identified
roles in morphogenesis. Both Ft and Ds are regulators of planar increased expression of core sarcomere components including
cell polarity (Matakatsu and Blair 2004; Simon 2004) and are com- Myosin alkali light chain 1 (Mlc1) (Falkenthal et al. 1984), wings up A
ponents of the Hippo signaling pathway (Hariharan and Bilder (wupA), which encodes the orthologue of Troponin I (Figure 2C)
2006; Willecke et al. 2006; Zhao et al. 2013), which is required for (Prado et al. 1995), and Tropomyosin 2 (Tm2), which encodes a
pupal eye patterning (DeAngelis et al. 2020). During larval eye de- Tropomyosin that functions cooperatively with Troponin I during
velopment, Cad86C is a downstream target of Hh and Dpp signal- muscle contraction (Karlik and Fyrberg 1986; Naimi et al. 2001)
ing and is required for apical cell constriction during (Figure 4). Since sarcomere-like structures are not found in the
morphogenetic furrow progression, while both Cad99C and pupal eye, we predict that these proteins are repurposed for other
Cad74A are required for the organization of the follicular epithe- roles in Drosophila retinal cells. For example, wupA and Tm2 are
lium during development of the Drosophila oocyte (Schlichting required for the maintenance of chromosomal integrity and cell
and Dahmann 2008; Zartman et al. 2008; Chung and Andrew polarity in the syncytial embryo, S2 cells, and larval wing epithe-
2014). It could be that Cad86C, Cad99C, and Cad74A are required lia (Sahota et al. 2009; Casas-Tintó and Ferrús 2019). Previous
to maintain epithelial cell shape and integrity during pupal eye studies also showed that Mlc1, wupA, and Tm2 can interact in8 | G3, 2021, Vol. 11, No. 1
Downloaded from https://academic.oup.com/g3journal/article/11/1/jkaa003/5995320 by guest on 28 October 2021
Figure 4 Genes associated with muscle function are expressed in the pupal eye. (A) Schematic representation of a sarcomere, composed of actin
filaments (yellow), myosin bundles (peach) and a myriad of proteins that maintain sarcomere structure or function. (B) Relative expression (fragments
per kilobase of transcript per million mapped reads) of sarcomere genes in the Drosophila eye at 21 and 40 h APF. Colored rectangles (at right) indicate
color of protein in illustration.
contexts other than the muscle sarcomere such as in S2R cells genes associated with signaling, axon guidance, biological adhe-
(Guruharsha et al. 2011). Further analyses are needed to clarify sion, or muscle structure development, suggesting that these
the precise molecular functions of these muscle-associated pro- processes are regulated by a limited number of transcription fac-
teins during pupal eye morphogenesis. tors (Supplementary Table S12). In contrast, we did not identify
recurrent transcription factor regulatory motifs in the groups of
Computational identification of putative genes associated with ecdysone signaling or cell survival that
transcriptional regulatory factors changed in expression from 21 to 40 h APF. This suggests that in
Our transcriptome analyses identified 4636 differentially the eye the genes associated with cell survival/apoptosis and ec-
expressed loci that contribute to multiple biological processes dysone signaling are not a part of regulatory networks but in-
(Supplementary Tables S2–S4). To gain insight into common reg- stead their expression is regulated by multiple transcription
ulatory elements that might be utilized to facilitate changes in factors.
gene expression, we used full i-Cis target analysis (Herrmann The conserved GAGA transcription factor, Trithorax-like (Trl)
et al. 2012) to identify transcriptional regulatory networks within (Farkas et al. 1994), emerged in our i-Cis analysis as a transcrip-
groups of functionally related differentially expressed genes tional regulator of loci associated with multiple processes includ-
(documented in Supplementary Tables S5–S10). These analyses ing signaling, axon projection, adhesion, and muscle structure
identified transcription factor-binding sites that were common to and development (Supplementary Table S12 and Figure 2C). AM. W. DeAngelis et al. | 9
potential role for Trl was further suggested by using AME, which and, given the highly conserved nature of many genes associated
identified transcription factor binding motifs that were enriched with tissue patterning, for the broader morphogenesis field as
in the complete list of differentially expressed genes. These anal- well.
yses identified several transcription factors, including Trl, which
may contribute to transcriptional regulation during pupal eye de-
velopment (Supplementary Table S13) (McLeay and Bailey 2010). Acknowledgments
In a previous study of the role of Trl in the apoptosis of lattice We thank the University of Michigan Advanced Genomics and
cells, patterning defects are also evident in Trl mutant clones, RNA Sequencing Core Facility for RNA-sequencing. Zack Drum,
underscoring a role for Trl in pupal eye morphogenesis (Dos- Michael Weir, Cathie Pfleger, Jake Aronowitz, and members of
Santos et al. 2008). Trl can influence transcription by functioning the Johnson and Coolon laboratories provided helpful discussion
as either a transcriptional activator (Biggin and Tjian 1988; and commentary on this article.
Soeller et al. 1993; Farkas et al. 1994) or a repressor (Farkas et al.
1994; Horard et al. 2000; Busturia et al. 2001; Poux et al. 2001;
Downloaded from https://academic.oup.com/g3journal/article/11/1/jkaa003/5995320 by guest on 28 October 2021
Mahmoudi et al. 2003; Mishra et al. 2003), and interestingly, our Funding
analyses indicated that Trl expression declined significantly at 40 Our work was supported by National Institute of Health grant
h relative to 21 h APF (Supplementary Tables S12 and S13). This R15GM114729.
change in Trl expression likely contributes to the very different
transcriptional profile of the eye at 40 h APF (Supplementary Conflicts of interest: None declared.
Table S1).
In addition to Trl, our i-Cis analysis identified the transcription
Literature cited
factor Homothorax (Hth) as a putative regulator of genes associ-
ated with axon guidance and muscle structure as well as Zelda Afgan E, Baker D, Batut B, van den Beek M, Bouvier D, et al. 2018 The
(Zld), which may contribute to the expression of genes associated Galaxy platform for accessible, reproducible and collaborative bio-
with axon guidance and adhesion (Supplementary Table S12 and medical analyses: 2018 update. Nucleic Acids Res. 46:W537–W544.
Figure 2C). Frequently associated with the Hippo pathway (Peng Al-Anzi B, Wyman RJ. 2009 The Drosophila immunoglobulin gene
et al. 2009), Hth is also required for the establishment of the eye turtle encodes guidance molecules involved in axon pathfinding.
field and photoreceptor specification (Pai et al. 1998; Pichaud and Neural Dev. 4:31.
Casares 2000; Wernet et al. 2003; Singh et al. 2011). Hth has also Anderson KV, Bokla L, Nüsslein-Volhard C. 1985 Establishment of
been implicated in muscle fiber formation, which correlates with dorsal-ventral polarity in the Drosophila embryo: the induction
our association of Hth with the transcription of muscle- of polarity by the Toll gene product. Cell. 42:791–798.
associated genes in the pupal eye (Bryantsev et al. 2012). The role Andrews S. 2010. FastQC: a quality control tool for high throughput
of Zld during eye development has not been explicitly character- sequence data.
ized during the pupal stage; however, previous studies indicate Apple R, Fristrom JW. 1991 20-Hydroxyecdysone is required for, and
that it is expressed in the larval eye disk (Giannios and Tsitilou negatively regulates, transcription of Drosophila pupal cuticle
2013). Another study reported that mutations in zld led to small protein genes. Dev Biol. 146:569–582.
and deformed eyes (Hamm et al. 2017). Like Trl, both Hth and Zld Araujo H, Machado LC, Octacı’lio-Silva S, Mizutani CM, Silva MJ,
function as either transcriptional activators or repressors and Ramos, et al. 2003 Requirement of the roughest gene for differen-
both declined in expression at 40 h APF (Inbal et al. 2001; tiation and time of death of interommatidial cells during pupal
Kobayashi et al. 2003; Liang et al. 2008; McDaniel et al. 2019). stages of Drosophila compound eye development. Mech Dev. 120:
Future studies will be needed to identify the transcriptional tar- 537–547.
gets of Trl, Hth, and Zld during pupal eye morphogenesis. Artero RD, Castanon I, Baylies MK. 2001 The immunoglobulin-like
To conclude, the pupal eye is an effective model system to use protein Hibris functions as a dose-dependent regulator of myo-
to interrogate processes required for the patterning or organiza- blast fusion and is differentially controlled by Ras and Notch sig-
tion of epithelia as well as pathways that lead to cell differentia- naling. Development. 128:4251–4264.
tion, and photoreceptor morphogenesis and axon projection. Ashburner M, Chihara C, Meltzer P, Richards G. 1974 Temporal con-
However, while many of the morphological changes associated trol of puffing activity in polytene chromosomes. Cold Spring
with these events have been well documented, less is known of Harb Symp Quant Biol. 38:655–662.
the corresponding transcriptional changes that drive them. Baehrecke EH, Thummel CS. 1995 The Drosophila E93 gene from the
Here, we compare the transcriptomes of pupal eyes at two 93F early puff displays stage-and tissue-specific regulation by
distinct stages of development. We identified changes in the 20-hydroxyecdysone. Dev Biol. 171:85–97.
expression of loci that are documented regulators of pupal Bao S. 2014 Notch controls cell adhesion in the Drosophila eye. PLoS
eye development such as components of Notch and EGFR signal- Genet. 10:e1004087.
ing pathways (Supplementary Table S5), ecdysone response Bao S, Cagan R. 2005 Preferential adhesion mediated by Hibris and
targets (Supplementary Table S6), regulators of axon guidance roughest regulates morphogenesis and patterning in the
(Supplementary Table S7), adhesion (Supplementary Table S8), Drosophila eye. Dev Cell. 8:925–935.
and cell survival (Supplementary Table S9). In addition, we iden- Bao S, Fischbach K-F, Corbin V, Cagan RL. 2010 Preferential adhesion
tified numerous novel gene expression changes that have not maintains separation of ommatidia in the Drosophila eye. Dev
been studied in the context of pupal eye morphogenesis. These Biology. 344:948–956.
included components of the Toll pathway (Supplementary Table Bashaw GJ, Kidd T, Murray D, Pawson T, Goodman CS. 2000
S5), non-classical cadherins (Supplementary Table S8), and nu- Repulsive axon guidance: Abelson and Enabled play opposing
merous proteins required for muscle development and structure roles downstream of the roundabout receptor. Cell. 101:703–715.
(Supplementary Table S10). We anticipate that these data will be Bate M, Martı́nez-Arias A. 1993. The Development of Drosophila mel-
a rich resource for future research on pupal eye morphogenesis anogaster. Plainview, NY: Cold Spring Harbor Laboratory Press.10 | G3, 2021, Vol. 11, No. 1
Becker H. 1959. The puffs of salivary gland chromosomes of homeodomain-containing protein required for the development
Drosophila melanogaster. Part 1. Observations on the behavior of of the entire visual system. Neuron. 12:977–996.
a typical puff in the normal strain and in two mutants, giant and Cho NK, Keyes L, Johnson E, Heller J, Ryner L, Karim, et al. 2002.
lethal giant larvae. Chromosoma. 10:654–678. Developmental control of blood cell migration by the Drosophila
Beiman M, Shilo B-Z, Volk T. 1996. Heartless, a Drosophila FGF recep- VEGF pathway. Cell. 108:865–876.
tor homolog, is essential for cell migration and establishment of Chung S, Andrew DJ. 2014. Cadherin 99C regulates apical expansion
several mesodermal lineages. Genes Dev. 10:2993–3002. and cell rearrangement during epithelial tube elongation.
Berger J, Senti K-A, Senti G, Newsome TP, Åsling B, Dickson, et al. Development. 141:1950–1960.
2008. Systematic identification of genes that regulate neuronal Clandinin TR, Zipursky SL. 2002. Making connections in the fly visual
wiring in the Drosophila visual system. PLoS Genet . e1000085.4: system. Neuron. 35:827–841.
Biggin MD, Tjian R. 1988. Transcription factors that activate the Clark HF, Brentrup D, Schneitz K, Bieber A, Goodman C, Noll, et al.
Ultrabithorax promoter in developmentally staged extracts. Cell. 1995. Dachsous encodes a member of the cadherin superfamily
53:699–711. that controls imaginal disc morphogenesis in Drosophila. Genes
Downloaded from https://academic.oup.com/g3journal/article/11/1/jkaa003/5995320 by guest on 28 October 2021
Bolkan BJ, Kretzschmar D. 2014. Loss of Tau results in defects in pho- Dev. 9:1530–1542.
toreceptor development and progressive neuronal degeneration Collinet C, Lecuit T. 2013. Stability and dynamics of cell–cell junc-
in Drosophila. Dev Neurobiol. 74:1210–1225. tions. Prog Mol Biol Transl Sci, 116:25–47.
Bour BA, Chakravarti M, West JM, Abmayr SM. 2000. Drosophila SNS, Cooley L, Verheyen E, Ayers K. 1992. chickadee encodes a profilin re-
a member of the immunoglobulin superfamily that is essential quired for intercellular cytoplasm transport during Drosophila
for myoblast fusion. Genes Dev. 14:1498–1511. oogenesis. Cell. 69:173–184.
Brown KE, Freeman M. 2003. Egfr signalling defines a protective func- Cordero J, Jassim O, Bao S, Cagan R. 2004. A role for wingless in an
tion for ommatidial orientation in the Drosophila eye. early pupal cell death event that contributes to patterning the
Development. 130:5401–5412. Drosophila eye. Mech Dev. 121:1523–1530.
Brückner K, Kockel L, Duchek P, Luque CM, Rørth P, Perrimon, et al. CRAN. 2018. R: A Language and Environment for Statistical
2004. The PDGF/VEGF receptor controls blood cell survival in Computing. Vienna, Austria: R Foundation for Statistical
Drosophila. Dev Cell. 7:73–84. Computing.
Bryantsev AL, Duong S, Brunetti TM, Chechenova MB, Lovato TL, et Curtiss J, Mlodzik M. 2000. Morphogenetic furrow initiation and pro-
al. 2012. Extradenticle and homothorax control adult muscle fi- gression during eye development in Drosophila: the roles of
ber identity in Drosophila. Dev Cell. 23:664–673. decapentaplegic, hedgehog and eyes absent. Development. 127:
Burtis KC, Thummel CS, Jones CW, Karim FD, Hogness DS. 1990. The 1325–1336.
Drosophila 74EF early puff contains E74, a complex Czerny T, Halder G, Kloter U, Souabni A, Gehring WJ, Busslinger, et al.
ecdysone-inducible gene that encodes two ets-related proteins. 1999. twin of eyeless, a second Pax-6 gene of Drosophila, acts up-
Cell. 61:85–99. stream of eyeless in the control of eye development. Mol Cell. 3:
Bushnell HL, Feiler CE, Ketosugbo KF, Hellerman MB, Nazzaro VL, et 297–307.
al. 2018. JNK is antagonized to ensure the correct number of inter- DeAngelis MW, Johnson RI. 2019. Dissection of the Drosophila pupal
ommatidial cells pattern the Drosophila retina. Dev Biol. 433: retina for immunohistochemistry, western analysis, and RNA
94–107. isolation. J Vis Exp. 145:e59299.
Busturia A, Lloyd A, Bejarano F, Zavortink M, Xin H, Sakonju, et al. DeAngelis MW, McGhie EW, Coolon JD, Johnson RI. 2020. Mask, a
2001. The MCP silencer of the Drosophila Abd-B gene requires component of the Hippo pathway, is required for Drosophila
both Pleiohomeotic and GAGA factor for the maintenance of re- eye morphogenesis. Dev Biol. 464:53–70.
pression. Development. 128:2163–2173. Denton D, Kumar S. 2015. Studying apoptosis in Drosophila. Cold
Cagan RL, Ready DF. 1989a. The emergence of order in the Spring Harb Protoc. 2015.
Drosophila pupal retina. Dev Biol. 136:346–362. Dorstyn L, Colussi PA, Quinn LM, Richardson H, Kumar S. 1999.
Cagan RL, Ready DF. 1989b. Notch is required for successive cell deci- DRONC, an ecdysone-inducible Drosophila caspase. Proc Natl
sions in the developing Drosophila retina. Genes Dev. 3: Acad Sci USA. 96:4307–4312.
1099–1112. Dos-Santos N, Rubin T, Chalvet F, Gandille P, Cremazy F, et al. 2008.
Capilla A, Karachentsev D, Patterson RA, Hermann A, Juarez MT, et Drosophila retinal pigment cell death is regulated in a
al. 2017. Toll pathway is required for wound-induced expression position-dependent manner by a cell memory gene. Int J Dev Biol.
of barrier repair genes in the Drosophila epidermis. Proc Natl 52:21–31.
Acad Sci USA. 114:E2682–E2688. Duchek P, Somogyi K, Jékely G, Beccari S, Rørth P. 2001. Guidance of
Carthew RW. 2007. Pattern formation in the Drosophila eye. Curr cell migration by the Drosophila PDGF/VEGF receptor. Cell. 107:
Opin Genet Dev. 17:309–313. 17–26.
Carvalho L, Jacinto A, Matova N. 2014. The Toll/NF-jB signaling Dworak HA, Charles MA, Pellerano LB, Sink H. 2001.
pathway is required for epidermal wound repair in Drosophila. Characterization of Drosophila hibris, a gene related to human
Proc Natl Acad Sci USA. 111:E5373–E5382. nephrin. Development. 128:4265–4276.
Casas-Tintó S, Ferrús A. 2019. Troponin-I mediates the localization Elhanany-Tamir H, Yanxun VY, Shnayder M, Jain A, Welte M, et al. 2012.
of selected apico-basal cell polarity signaling proteins. J Cell Sci. Organelle positioning in muscles requires cooperation between two
132:jcs225243. KASH proteins and microtubules. J Cell Biol. 198:833–846.
Chasan R, Anderson KV. 1989. The role of Easter, an apparent serine Falkenthal S, Parker VP, Mattox WW, Davidson N. 1984. Drosophila
protease, in organizing the dorsal-ventral pattern of the melanogaster has only one myosin alkali light-chain gene which
Drosophila embryo. Cell. 56:391–400. encodes a protein with considerable amino acid sequence
Cheyette BN, Green PJ, Martin K, Garren H, Hartenstein V, Zipursky, homology to chicken myosin alkali light chains. Mol Cell Biol. 4:
et al. 1994. The Drosophila sine oculis locus encodes a 956–965.M. W. DeAngelis et al. | 11
Fanto M, Mlodzik M. 1999. Asymmetric Notch activation specifies Hall H, Medina P, Cooper DA, Escobedo SE, Rounds J, Brennan, et al.
photoreceptors R3 and R4 and planar polarity in the Drosophila 2017. Transcriptome profiling of aging Drosophila photoreceptors
eye. Nature. 397:523–526. reveals gene expression trends that correlate with visual senes-
Farkas G, Gausz J, Galloni M, Reuter G, Gyurkovics H, Karch, et al. cence. BMC Genomics. 18:894.
1994. The Trithorax-like gene encodes the Drosophila GAGA fac- Hamm DC, Larson ED, Nevil M, Marshall KE, Bondra ER, et al. 2017. A
tor. Nature. 371:806–808. conserved maternal-specific repressive domain in Zelda revealed
Fleming RJ, Gu Y, Hukriede NA. 1997. Serrate-mediated activation of by Cas9-mediated mutagenesis in Drosophila melanogaster.
Notch is specifically blocked by the product of the gene fringe in PLoS Genet. 13:e1007120.
the dorsal compartment of the Drosophila wing imaginal disc. Hariharan IK, Bilder D. 2006. Regulation of imaginal disc growth by
Development. 124:2973–2981. tumor-suppressor genes in Drosophila. Annu Rev Genet. 40:
Fortini ME, Rebay I, Caron LA, Artavanis-Tsakonas S. 1993. An acti- 335–361.
vated Notch receptor blocks cell-fate commitment in the devel- Hay BA, Wassarman DA, Rubin GM. 1995. Drosophila homologs of
oping Drosophila eye. Nature. 365:555–557. baculovirus inhibitor of apoptosis proteins function to block cell
Downloaded from https://academic.oup.com/g3journal/article/11/1/jkaa003/5995320 by guest on 28 October 2021
Frankfort BJ, Nolo R, Zhang Z, Bellen H, Mardon G. 2001. senseless re- death. Cell. 83:1253–1262.
pression of rough is required for R8 photoreceptor differentiation Hayashi T, Carthew RW. 2004. Surface mechanics mediate pattern
in the developing Drosophila eye. Neuron. 32:403–414. formation in the developing retina. Nature. 431:647–652.
Fraser AG, Evan GI. 1997. Identification of a Drosophila melanogaster Haynie JL, Bryant PJ. 1986. Development of the eye-antenna imaginal
ICE/CED-3-related protease, drICE. EMBO J. 16:2805–2813. disc and morphogenesis of the adult head in Drosophila mela-
Freeman M. 1996. Reiterative use of the EGF receptor triggers differ- nogaster. J Exp Zool. 237:293–308.
entiation of all cell types in the Drosophila eye. Cell. 87:651–660. Heino TI, Kärpänen T, Wahlström G, Pulkkinen M, Eriksson U, et al.
Freeman M. 1997. Cell determination strategies in the Drosophila 2001. The Drosophila VEGF receptor homolog is expressed in
eye. Development. 124:261–270. hemocytes. Mech Dev. 109:69–77.
Freeman M, Bienz M. 2001. EGF receptor/Rolled MAP kinase signal- Henderson CA, Gomez CG, Novak SM, Mi-Mi L, Gregorio CC. 2011.
Overview of the muscle cytoskeleton. Compr Physiol. 7:891–944.
ling protects cells against activated Armadillo in the Drosophila
Herrmann C, Van de Sande B, Potier D, Aerts S. 2012. i-cisTarget: an
eye. EMBO Rep. 2:157–162.
integrative genomics method for the prediction of regulatory fea-
Gaudet P, Livstone MS, Lewis SE, Thomas PD. 2011.
tures and cis-regulatory modules. Nucleic Acids Res. 40:
Phylogenetic-based propagation of functional annotations within
e114–e114.
the Gene Ontology consortium. Brief Bioinformatics. 12:449–462.
Hill E, Broadbent ID, Chothia C, Pettitt J. 2001. Cadherin superfamily
Gho M, Lecourtois M, Géraud G, Posakony JW, Schweisguth F. 1996.
proteins in Caenorhabditis elegans and Drosophila mela-
Subcellular localization of suppressor of hairless in Drosophila
nogaster. J Mol Biol. 305:1011–1024.
sense organ cells during Notch signalling. Development. 122:
Honn JS, Johansson L, Lestander ÅR. 2016. Regulation of twin of eye-
1673–1682.
less during Drosophila development. Gene Expr Patterns. 20:
Giannios P, Tsitilou SG. 2013. The embryonic transcription factor
120–129.
Zelda of Drosophila melanogaster is also expressed in larvae and
Horard B, Tatout C, Poux S, Pirrotta V. 2000. Structure of a polycomb
may regulate developmentally important genes. Biochem
response element and in vitro binding of polycomb group com-
Biophys Res Commun. 438:329–333.
plexes containing GAGA factor. Mol Cell Biol. 20:3187–3197.
Gibbs SM, Truman JW. 1998. Nitric oxide and cyclic GMP regulate ret-
Horng T, Medzhitov R. 2001. Drosophila MyD88 is an adapter in the
inal patterning in the optic lobe of Drosophila. Neuron. 20:83–93.
Toll signaling pathway. Proc Natl Acad Sci USA. 98:12654–12658.
Gorski S, Brachmann CB, Tanenbaum S, Cagan R. 2000. Delta and
Hynes RO, Zhao Q. 2000. The evolution of cell adhesion. J Cell Biol.
notch promote correct localization of irreC-rst. Cell Death Differ.
150:F89–F96.
7:1011–1013.
Ikmi A, Gaertner B, Seidel C, Srivastava M, Zeitlinger J, Gibson, et al.
Greenwood S, Struhl G. 1999. Progression of the morphogenetic fur-
2014. Molecular evolution of the Yap/Yorkie proto-oncogene and
row in the Drosophila eye: the roles of Hedgehog, elucidation of its core transcriptional program. Mol Biol Evol. 31:
Decapentaplegic and the Raf pathway. Development. 126: 1375–1390.
5795–5808. Inbal A, Halachmi N, Dibner C, Frank D, Salzberg A. 2001. Genetic ev-
Grether ME, Abrams JM, Agapite J, White K, Steller H. 1995. The head idence for the transcriptional-activating function of Homothorax
involution defective gene of Drosophila melanogaster functions during adult fly development. Development. 128:3405–3413.
in programmed cell death. Genes Dev. 9:1694–1708. Iwai Y, Usui T, Hirano S, Steward R, Takeichi M, Uemura, et al. 1997.
Grillo-Hill BK, Wolff T. 2009. Dynamic cell shapes and contacts in Axon patterning requires D N-cadherin, a novel neuronal adhe-
the developing Drosophila retina are regulated by the Ig cell ad- sion receptor, in the Drosophila embryonic CNS. Neuron. 19:
hesion protein hibris. Dev Dyn. 238:2223–2234. 77–89.
Grzeschik NA, Knust E. 2005. IrreC/rst-mediated cell sorting during Jang AC-C, Chang Y-C, Bai J, Montell D. 2009. Border-cell migration
Drosophila pupal eye development depends on proper localisa- requires integration of spatial and temporal signals by the BTB
tion of DE-cadherin. Development. 132:2035–2045. protein Abrupt. Nat Cell Biol. 11:569–579.
Guillot C, Lecuit T. 2013. Mechanics of epithelial tissue homeostasis Jang C-C, Chao J-L, Jones N, Yao L-C, Bessarab DA, Kuo, et al. 2003.
and morphogenesis. Science. 340:1185–1189. Two Pax genes, eye gone and eyeless, act cooperatively in
Guruharsha K, Rual J-F, Zhai B, Mintseris J, Vaidya P, Vaidya, et al. promoting Drosophila eye development. Development. 130:
2011. A protein complex network of Drosophila melanogaster. 2939–2951.
Cell. 147:690–703. Jang I-H, Chosa N, Kim S-H, Nam H-J, Lemaitre B, Ochiai, et al.
Halder G, Callaerts P, Gehring WJ. 1995. Induction of ectopic eyes by 2006. A Spätzle-processing enzyme required for toll
targeted expression of the eyeless gene in Drosophila. Science. signaling activation in Drosophila innate immunity. Dev Cell.
267:1788–1792. 10:45–55.You can also read

















































