Biogenic signals from plastids and their role in chloroplast development
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Journal of Experimental Botany, Vol. 73, No. 21 pp. 7105–7125, 2022
https://doi.org/10.1093/jxb/erac344 Advance Access Publication 25 August 2022
DARWIN REVIEW
Biogenic signals from plastids and their role in chloroplast
development
Monique Liebers1, Carolina Cozzi1, Finia Uecker1, Louise Chambon2, Robert Blanvillain2, and
Downloaded from https://academic.oup.com/jxb/article/73/21/7105/6674789 by guest on 13 December 2022
Thomas Pfannschmidt1,*,
1
Gottfried-Wilhelm-Leibniz-Universität Hannover, Naturwissenschaftliche Fakultät, Institut für Botanik, Pflanzenphysiologie, Herrenhäuser
Str. 2, D-30419 Hannover, Germany
2
Université Grenoble-Alpes, CNRS, CEA, INRA, IRIG-LPCV, F-38000 Grenoble, France
* Correspondence: T.Pfannschmidt@botanik.uni-hannover.de
Received 2 March 2022; Editorial decision 11 August 2022; Accepted 23 August 2022
Editor: Donald Ort, University of Illinois, USA
Abstract
Plant seeds do not contain differentiated chloroplasts. Upon germination, the seedlings thus need to gain photoau-
totrophy before storage energies are depleted. This requires the coordinated expression of photosynthesis genes
encoded in nuclear and plastid genomes. Chloroplast biogenesis needs to be additionally coordinated with the light
regulation network that controls seedling development. This coordination is achieved by nucleus to plastid signals
called anterograde and plastid to nucleus signals termed retrograde. Retrograde signals sent from plastids during
initial chloroplast biogenesis are also called biogenic signals. They have been recognized as highly important for
proper chloroplast biogenesis and for seedling development. The molecular nature, transport, targets, and signalling
function of biogenic signals are, however, under debate. Several studies disproved the involvement of a number of
key components that were at the base of initial models of retrograde signalling. New models now propose major roles
for a functional feedback between plastid and cytosolic protein homeostasis in signalling plastid dysfunction as well
as the action of dually localized nucleo-plastidic proteins that coordinate chloroplast biogenesis with light-dependent
control of seedling development. This review provides a survey of the developments in this research field, summarizes
the unsolved questions, highlights several recent advances, and discusses potential new working modes.
Keywords: Biogenic signals, cellular communication, gene expression, light regulation, PEP-associated proteins,
photomorphogenesis, plastids.
Introduction
Plastids are cell organelles that are characteristic for plant and endosymbiosis are far from fully understood, and phyloge-
algal cells. They originated from an event in which a mito- nomic analyses even suggest the involvement of Chlamydiae as
chondriated eukaryotic cell engulfed a cyanobacteria-like pho- a third partner (reviewed in Ball et al., 2016; Zeng and Dehesh,
tosynthetic prokaryote.This engulfment was of mutual benefit, 2021). However, it is agreed that all plastid lineages trace back
resulting in a stable endosymbiosis (reviewed in Archibald, to one endosymbiotic event forming a monophyletic group
2015). The circumstances leading to the establishment of this (reviewed in Delwiche, 1999; Stoebe and Maier, 2002).
© The Author(s) 2022. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email:
journals.permissions@oup.com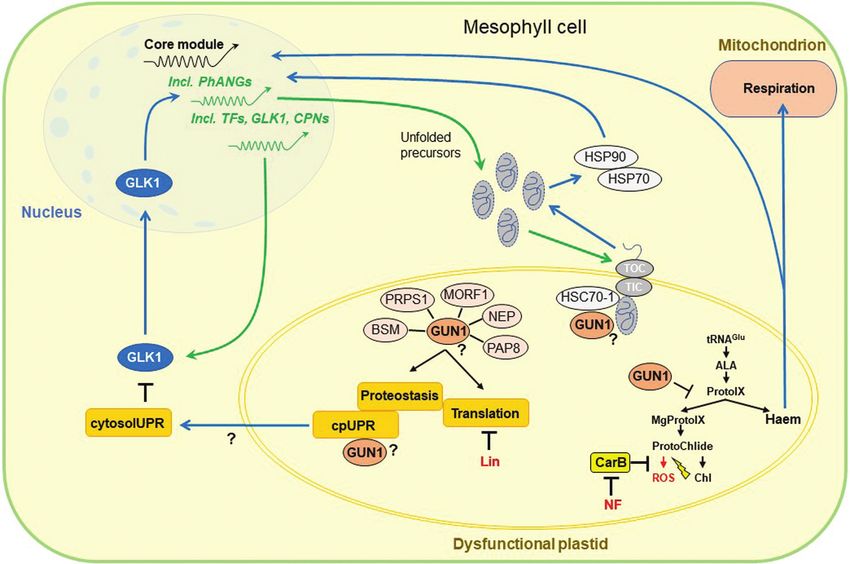
7106 | Liebers et al.
Plants and green algae derived from the green lineage and signals can be distinguished: (i) plastid pigments or pigment
possess primary chloroplasts that enable them to perform pho- precursors (tetrapyrroles, apocarotenoids, and carotenoids); (ii)
tosynthesis. Chloroplasts of even distant species share highly signals originating from plastid gene expression (PGE); (iii) re-
conserved features including a double envelope membrane, a active oxygen species (ROS); (iv) photosynthesis-related redox
protein import machinery (the TOC–TIC complex, translo- signals; and (v) changes in metabolite pools or fluxes (reviewed
con of the outer and inner membrane of the chloroplast, re- in Chan et al., 2016; de Souza et al., 2016; Hernandez-Verdeja
spectively), an internal thylakoid membrane system embedding and Strand, 2018). These signals do not all occur at the same
the photosynthetic apparatus, a plastid-specific genome (the time or in the same developmental context, and some signals
plastome), and fully functional transcription and translation may even appear under several conditions but causing dif-
machineries for the expression of this plastome. Many of these ferent effects. In order to respect the developmental and en-
features display prokaryotic elements or structures that are sig- vironmental context in which these retrograde signals become
Downloaded from https://academic.oup.com/jxb/article/73/21/7105/6674789 by guest on 13 December 2022
natures of the prokaryotic ancestry of chloroplasts such as pro- active, one can distinguish: (i) biogenic signals that are active
karyotic –35 and –10 promoter elements, 70S ribosomes, or an during early steps of chloroplast biogenesis (e.g. during pro-
Escherichia coli-like RNA polymerase (reviewed in Pyke, 2007; plastid to chloroplast or etioplast to chloroplast transition); (ii)
Jarvis and López-Juez, 2013). However, during the course of operational signals that are active when the chloroplast is fully
evolution, the endosymbiont was re-shaped in many aspects, functional and responds to the environment; and (iii) degra-
a process that allowed the final functional and structural inte- dational retrograde signals that are active when plastids are
gration into the host cell. One important step in this process degraded in response to age or pathogen attack (reviewed in
was the transfer of the largest part of the genome of the en- Pogson et al., 2008; Pfannschmidt et al., 2020). In this review,
dosymbiont into the host nuclear genome. This gene transfer we focus mainly on the field of biogenic signals where much
expanded the coding capacity of the host cell and provided progress has been obtained in the last years. For detailed in-
novel factors and elements for a functional enhancement of the formation about the other signals, the interested reader is re-
biochemical and regulatory properties in it (Martin et al., 2002; ferred to recent reviews (Chan et al., 2016; de Souza et al.,
reviewed in Bock and Timmis, 2008; Criscuolo and Gribaldo, 2016; Hernandez-Verdeja and Strand, 2018). Most of the prog-
2011). ress discussed below concerns studies using dictotyledonous
Today’s plant chloroplasts contain a highly conserved ge- Angiosperms (mostly Arabidopsis thaliana). Monocotyledonous
nome of ~120–160 kbp that carries ~120 genes encoding Angiosperms or Gymnosperms were investigated much less
components of the photosynthetic apparatus and protein sub- frequently; however, because of their distinct differences in
units of the ribosomes and the RNA polymerase, rRNAs, and chloroplast development, they provide important additions to
tRNAs (reviewed in Sugiura, 1992; Bock, 2007). Proteomic the field (Hills et al., 2015; Loudya et al., 2021).
analyses, however, identified 2500–3500 different proteins
in chloroplasts, exceeding by far the coding capacity of the
plastome (von Zychlinski et al., 2005; Zybailov et al., 2008; Plastid development during early steps of
Ferro et al., 2010). Thus, the vast majority of plastid-localized the plant life cycle
proteins are encoded in the nucleus, translated in the cytosol Plastids are morphologically and functionally very diverse, and
as pre-proteins, and finally imported into the organelle (Bauer chloroplasts represent only one specific form of them. In multicel-
et al., 2000; reviewed in Soll and Schleiff, 2004; Strittmatter lular plants, one can observe chromoplasts, amyloplasts, etioplasts,
et al., 2010; Nakai, 2018). Intriguingly, all major protein com- or elaioplasts. They all develop from the same undifferentiated
plexes in chloroplasts are composed of a mix of nuclear- and precursor, the proplastid, that is inherited between generations
plastid-encoded subunits, indicating that their expression needs and that is found in meristematic stem cells. However, the tissue
to be coordinated, allowing a successful assembly of these pro- context of the cell decides which type of plastid is formed and,
tein complexes. For many years, it was believed that the nu- therefore, a given cell usually contains only one type of plastid. In-
cleus exclusively controls chloroplast biogenesis by providing dependent of the respective form, all plastid types contain the same
all critical protein factors and structural components; however, plastome, allowing them interconversions of these types in case
in recent years, it became increasingly clear that the chloroplast developmental or environmental conditions change (reviewed in
itself also provides essential regulatory signals that contribute Lopez-Juez and Pyke, 2005; Liebers et al., 2017).
to this coordination. In order to distinguish between these two
types of regulation, the terms ‘anterograde control’ (describing Plastid development during embryogenesis and early
the nucleus to chloroplast signalling) and ‘retrograde control’ seedling development, and the impact of plastid
(describing the chloroplast to nucleus signalling) (Mochizuki transcription
et al., 2001) were coined (reviewed in Woodson and Chory,
2008). Retrograde signalling from plastids has attracted much Plastids cannot be formed de novo, but are inherited from pa-
interest in the last three decades, and considerable progress has rental progenitor cells (reviewed in Pyke, 2007). During their
been made in its understanding. So far, five different classes of division, the inherited proplastids are randomly distributed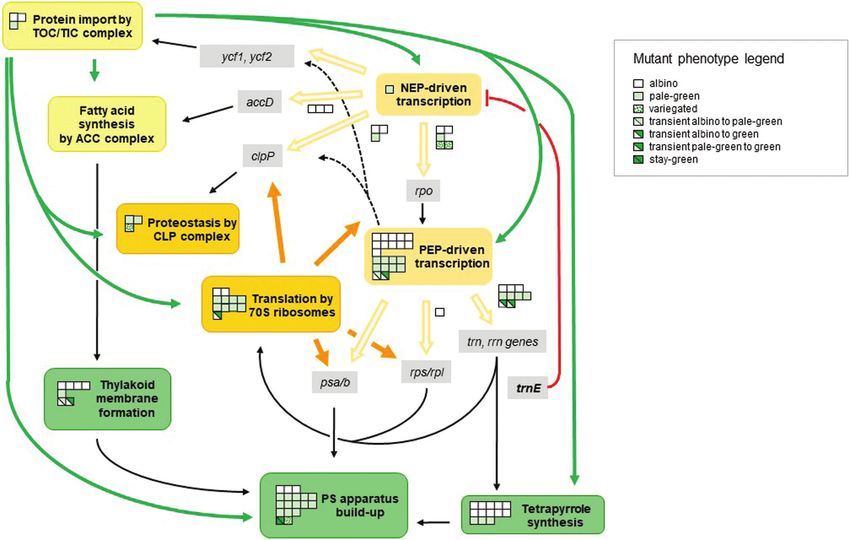
Retrograde biogenic signals | 7107
to the daughter cells in which they subsequently divide and (Rodermel and Bogorad, 1985; Demarsy et al., 2012). Fol-
multiply by fission, using a prokaryotic-type division apparatus lowing the increase in transcript abundance, protein abundance
(reviewed in Osteryoung and Pyke, 2014). In vascular plants, is also rising (Kanervo et al., 2008). However, RNAP activity
all types and interstages of plastid transmission, such as ma- as well as RNA stability need to be considered as both change
ternal, biparental, or paternal inheritance, as well as maternal during chloroplast formation.This becomes especially apparent
inheritance with parental leakage were found (reviewed in in the developmental gradient along the leaf blade of mono-
Timmis et al., 2004; Pyke, 2007; Greiner et al., 2015). Most cotyledonous plants that grow from a basal meristem. Cells at
Angiosperms inherit plastids uniparentally, but the underlying the base are the youngest, with poorly developed chloroplasts.
mechanisms vary greatly, even between close taxa (reviewed in Towards the leaf tip, cell age increases and chloroplasts become
Greiner et al., 2015). mature, allowing the determination of transcription rates and
After fertilization, a characteristic developmental pro- transcript stabilities at different developmental stages (Mullet
Downloaded from https://academic.oup.com/jxb/article/73/21/7105/6674789 by guest on 13 December 2022
gramme takes place that comprises embryogenesis, maturation, and Klein, 1987; Baumgartner et al., 1993; Cahoon et al., 2004).
and desiccation, ultimately leading to the formation of mature
embryos (Le et al., 2010). In oilseed plants, such as A. thali- Plastid development during photomorphogenesis
ana, this includes a transient formation of photosynthetically
active chloroplasts, starting at the globular stage (Tejos et al., In many Angiosperms, germination occurs when a species-
2010). With the progressing ripening of the seed, these plas- dependent combination of temperature, moisture, and light is
tids de-differentiate to non-photosynthetic, colourless eoplasts perceived by the quiescent embryo. Whenever light is required
that, similar to proplastids, are able to re-differentiate into all to break dormancy, a minimum of a few hours of exposure to
plastid types (Allorent et al., 2013; reviewed in Liebers et al., specific wavelengths, such as red light, is necessary to activate
2017). The conversion between plastid types involves changes the phytochromes until germination is irreversibly launched.
in nuclear gene expression and significant changes in plastid Then photoreceptors can be converted back to their inactive
transcriptional activities. There exist two types of plastid RNA forms; naturally and slowly in the dark (mimicking night con-
polymerases (RNAPs): (i) a nuclear encoded RNA poly- ditions) or artificially by far-red light exposure. The latter were
merase (NEP), which is a single-subunit T3/T7 phage-type defined as ‘true-dark conditions’ that correspond to the photo-
polymerase and (ii) a plastid encoded RNA polymerase (PEP) conversion of phytochromes into the inactive stage; although
that forms a multisubunit complex of prokaryotic origin this is only achievable under laboratory conditions and not
(Lerbs-Mache, 1993; Hedtke et al., 1997; reviewed in Borner fully complete since Pr also absorbs some far-red light (Leivar
et al., 2015). The two types of RNAPs have distinct as well as et al., 2008). Once germination has started, the seedling enters
overlapping functions, and their relative contribution to the a direct photomorphogenetic programme, or engages in a dark
expression of the plastome varies greatly over time and with developmental programme called skotomorphogenesis. In di-
developmental conditions. The often-found notion that NEP rect photomorphogenesis, chloroplast biogenesis has been fol-
is responsible for the expression of house-keeping genes, while lowed through fluorescent studies (Yadav et al., 2019) showing
PEP is responsible for the expression of photosynthesis genes, that cell specificity of chloroplast biogenesis in above-ground
thus is a simplification that does not reflect the complexity tissues (mostly cotyledons) starts after the emergence of the
in the distribution of work between the two plastid RNAP root tip from the seed coat. However, plastids of the cotyledons
activities. Plastidial genes are classified according to the pres- transitorily lose their chlorophyll autofluorescence during early
ence of NEP and/or PEP recognition sites in their promot- development while ribosomes are present in the stroma (Liang
ers (Hajdukiewicz et al., 1997; reviewed in Liere et al., 2011; et al., 2018) and the plastome expression machinery is active
Pfannschmidt et al., 2015). Class I genes are solely transcribed (Dubreuil et al., 2018). The inner membrane of the plastid en-
by PEP (e.g. psbA, psbB, psbK, rbcL, petB, and ndhA), class II velope invaginates to form tubulovesicular thylakoids, then the
genes are recognized by both RNAPs (e.g. atpB, atpI, ndhB, plastids increase in size while the inner membranes become flat
ndhF, clpP, and ycf1), and class III genes are only NEP de- and thin. This membrane remodelling is accompanied by the
pendent (e.g. ycf2, accD, and rpoB/C1/C2). During the proplas- association of polysomes on thylakoids, and protein translation
tid to chloroplast transition in developing embryos, both NEP of the photosynthetic apparatus while penetrating within the
and PEP genes are up-regulated. However, while NEP-depen- membrane. Subunits of PSII appear and gradually accumulate
dent transcripts only slightly increase, photosynthesis-associ- throughout the development of the light-grown seedling. The
ated genes are tremendously up-regulated (Allorent et al., 2013; flattened membranes then produce small pre-grana and serve
Kremnev and Strand, 2014). A massive re-programming, in- as nucleation points for subsequent stacking (Liang et al., 2018).
cluding a genome-wide up-regulation of photosynthesis genes, From embryogenesis up to the first 2 d post-imbibition, the
was also observed during germination in long-day conditions shoot apical meristem (SAM) and epidermis are completely
as well as during the etioplast to chloroplast transition, where devoid of chlorophyll, regardless of their light environment
the maximal transcript accumulation is reached after radicle (Tejos et al., 2010; Yadav et al., 2019). In these areas, plastids
outgrowth or 10–44 h after a dark to light shift, respectively are proplastids with only a few vesicles, no thylakoid structure
7108 | Liebers et al.
except for a few non-photosynthetic tubules, and display re- compounded roles of the different phytochromes with the dif-
sidual photosynthesis proteins (Charuvi et al., 2012). In the ferent PIFs allow a fine-tuning of the developmental responses
course of development depending on the specific cell lineages, to their light environment, including de-etiolation and shade
the SAM can differentiate rudimentary chloroplasts, except in avoidance. Furthermore, PIFs are also negative regulators of
the L2 central zone as well as some cells below the L3 layer chloroplast biogenesis, having direct effects on rate-limiting
(Charuvi et al., 2012; Dalal et al., 2018;Yadav et al., 2019). In the steps of the tetrapyrrole biosynthetic pathway (Stephenson et al.,
peripheral zone, stacked internal membrane systems develop 2009). The phytochrome-mediated light signalling represses
within photosynthetic plastids that will either regress or further the COP1-mediated degradation of HY5, thereby leading to
develop, depending on their position and subsequent cell fate. the accumulation of a different post-translationally modified
During skotomorphogenesis, etiolated tissues produce a typ- HY5. This stabilized transcription factor (together with fur-
ical round and small plastid that contains starch, plastoglobuli, ther factors) is then responsible for a profound reprogramming
Downloaded from https://academic.oup.com/jxb/article/73/21/7105/6674789 by guest on 13 December 2022
and an imposing structure named the prolamellar body (PLB) of the transcriptional activities with respect to its numerous
as well as proto-thylakoids (PTs). These etioplasts do not stack target genes (Hajdu et al., 2018). Notably, HY5 initiates the
internal membranes; they are devoid of chlorophyll, while the expression of photomorphogenic factors (Lee et al., 2007).
presence of carotenoids is largely responsible for the yellow Meanwhile, light exposure triggers the transcriptional activa-
colour of the cotyledons. Etioplasts are developmentally halted tion of golden2-like myb transcription factors 1 and 2 (GLK1
and can be regarded as a precursor stage capable of acquiring and 2) that are responsible for the proper expression of nuclear
a chloroplast structure in just a few hours after illumination. photosynthesis genes (reviewed in Waters and Langdale, 2009;
Many studies of chloroplast biogenesis, therefore, use a dark Waters et al., 2009) required to build up the photosynthetic
to light shift (Armarego-Marriott et al., 2020). The PLB has machinery in the plastids. The rapidity of the responses at gene
a unique lipid–protein–pigment composition in a hexagonal expression and subsequent morphological levels prevented a
para-crystalline structure sharing a continuous lumen (Flo- precise dissection of the sequence of events that lead eventually
ris and Kuhlbrandt, 2021). Most of the prolamellar proteome to a green photomorphogenic seedling. However, recent de-
corresponds to the light-dependent protochlorophyllide oxi- tailed studies provided significant progress on this question (see
doreductase (LPOR) that is linked in a tertiary complex to below for more details) (Dubreuil et al., 2018; Loudya et al.,
NADPH and protochlorophyllide (reviewed in Solymosi and 2021; Pipitone et al., 2021). The functional involvement of cy-
Schoefs, 2010) as well as some photosynthesis precursor pro- tosolic and nuclear regulators in this particular step of seedling
teins (Blomqvist et al., 2008). The presence of these subunits in development appears to be well understood, but several studies
etioplasts allows rapid synthesis of chlorophylls and construc- revealed that retrograde signals from plastids are also required.
tion of the photosynthetic apparatus after illumination. The relative contribution of these signals as well as the coordi-
The genetic control of the dark or light developmental nation with the nucleo-cytosolic system, however, remains to
programme involves two major interacting pathways corre- be elucidated and is a major topic of this review.
sponding to two classes of isolated mutants; long hypocotyl
(hy) and constitutively photomorphogenic (cop). Most im-
portant, in the dark, photomorphogenesis is inhibited by a Retrograde biogenic signals in chloroplast
degradational module (the COP9 signalosome) (reviewed in biogenesis
Sullivan et al., 2003; Lau and Deng, 2012; Seluzicki et al., 2017).
Initial experimental identification of signals and mutants
The E3 ubiquitin ligase COP1 destabilizes the basic domain/
leucine zipper (bZIP) transcription factor elongated hypocotyl Retrograde biogenic signals were originally identified in
5 (HY5) and its closest homologue HYH (Osterlund et al., plants with dysfunctional plastids. The first notion came
2000; Holm et al., 2002).Then light converts the cytosolic pool from studies with the barley mutant albostrians that exhib-
of inactive phytochrome B (Pr) into its active state (Pfr), trig- its defects in plastid ribosome formation in cells of its basal
gering its nuclear translocation (Yamaguchi et al., 1999; Chen leaf meristem. A recently identified genetic defect in the nu-
et al., 2003). Specific domains of the phytochromes interact clear gene for a CCT domain protein (Li et al., 2019) results
with phytochrome-interacting factors (PIFs) (Huq et al., 2004), in white striped leaves with variable degrees of variegation
leading to mutual degradation or inactivation of the partners in this mutant. While the green tissues harbour fully func-
(reviewed in Leivar and Monte, 2014). PIFs comprise four tional chloroplasts (making the mutant viable), white tissues
basic helix–loop–helix (bHLH) transcription factors (PIF1, cannot perform photosynthesis. Interestingly, the expression
3, 4, and 5) acting as transcriptional repressors of photomor- of not only plastid proteins was diminished by the ribo-
phogenesis and activators of skotomorphogenesis (Leivar et al., some defect but also that of nuclear-encoded photosynthesis
2008). The interaction of PIFs with photoactivated phyto- proteins such as RBCS, leading to the proposal of a plastid
chromes leads to their degradation (Al-Sady et al., 2006), sub- factor that reports the functional state of the plastids to the
sequently releasing the repression of the photomorphogenic nucleus (Bradbeer et al., 1979). This concept was supported
programme (reviewed in Jiao et al., 2007).The antagonistic and by studies in plants in which carotenoid biogenesis wasRetrograde biogenic signals | 7109
inhibited either through pharmacological or genetic were studied in great detail, and gun2–gun6 were all identified
approaches during the very early steps of seedling develop- to contain defects in enzymes of the plastid tetrapyrrole bi-
ment, leading to photo-oxidation. Lack of carotenoids in this osynthesis pathway (TBP). GUN2 encodes haem oxygenase
very sensitive developmental stage in which thylakoid mem- and GUN3 encodes phytochromobilin synthase (Mochizuki
branes still have to be established results in a light-induced et al., 2001). Both enzymes are located on the haem branch
destruction of internal plastid structures, and thus prevents of the TBP pathway and catalyse essential steps in the bio-
the biogenesis of chloroplasts, resulting in dysfunctional plas- synthesis of the phytochrome chromophore phytochromobilin
tids like those in the albostrians mutant. Maize and mustard (Kohchi et al., 2001). GUN4 encodes an activating regulator
seedlings with such photo-oxidized, dysfunctional plastids of the Mg-chelatase and GUN5 the H-subunit of the Mg-
displayed a repression of the nuclear-encoded genes LHCB chelatase complex (Mochizuki et al., 2001; Larkin et al., 2003).
(formerly called CAB) and RBCS (Mayfield and Taylor, 1984; Thus, both enzymes are involved in chlorophyll biosynthesis.
Downloaded from https://academic.oup.com/jxb/article/73/21/7105/6674789 by guest on 13 December 2022
Oelmuller and Mohr, 1986). Based on these results, it was The gun6 mutant overexpresses ferrochelatase 1 (FC1) that
assumed that plastids could communicate their developmental forms haem from protoporphyrin IX (Woodson et al., 2011),
and functional status in order to adapt nuclear photosynthesis and thus belongs to the haem branch of the TBP. Because of
gene expression (reviewed in Oelmuller, 1989; Taylor, 1989). the high proportion of TBP enzymes among the GUN com-
Pharmaceutical inhibition of chloroplast biogenesis in many ponents, it was assumed that metabolic intermediates of the
studies is achieved by the application of norflurazon (NF), a TBP pathway, such as Mg-protoporphyrin-IX (Mg-proto-
potent inhibitor of the phytoene desaturase, the key entry en- IX), or the endpoint product haem could serve as retrograde
zyme of carotenoid biosynthesis (Chamovitz et al., 1991). The signals (Mochizuki et al., 2001; Strand et al., 2003; Woodson
inhibitor, in addition, has been recently reported also to inhibit et al., 2011). It was reported that amino-laevulinic acid (ALA)
FAD2, an enzyme involved in fatty acid desaturation (Abrous- feeding prior to NF treatment made Mg-proto-IX detectable
Belbachir et al., 2009), and it therefore may also have a direct in the cytosol (Ankele et al., 2007). However, further analyses
effect on the formation of thylakoid membrane lipids. Dark- could not identify significant correlations between changes in
or light-grown seedlings treated with this inhibitor exhibit nuclear gene expression and the accumulation of this interme-
white cotyledons, demonstrating the effective inhibition of diate, ruling out that the levels of Mg-proto-IX per se influence
carotenoid biosynthesis; however, photo-oxidative destruction nuclear gene expression (Mochizuki et al., 2008; Moulin et al.,
occurs only in the presence of illumination. Alternatively, chlo- 2008). Nevertheless, it cannot be excluded that Mg-proto-IX
roplast biogenesis in germinating seedlings can be blocked by could function as a transmitter of plastid signals by activation
application of inhibitors of prokaryotic gene expression. Chlo- and/or interaction with other potential retrograde signals, such
ramphenicol, erythromycin, or lincomycin (Lin) have been re- as ROS and abscisic acid (ABA) (Voigt et al., 2010). For haem,
ported to block plastid translation, resulting in seedlings of pale the situation is somewhat different since it has been known
appearance that display reduced levels of LHCB and RBCS for a long time that haem is exported from plastids and trans-
transcripts (Oelmuller et al., 1986; Gray et al., 1995).Treatments ported to the mitochondria where it serves as a redox cofactor
with the transcription inhibitors tagetitoxin and rifampicin in mitochondrial electron transport (Thomas and Weinstein,
were found to be equally effective (Rapp and Mullet, 1991; 1990). Indeed, analysis of the gun6-1D mutant of Arabidopsis
Pfannschmidt and Link, 1997). Such inhibitor experiments re- indicated that increased flux through the FC1-mediated haem
vealed several important facts: (i) the plastid signal can orig- synthesis pathway promoted expression of photosynthesis-
inate from both plastid translation and transcription; (ii) the associated nuclear genes (PhANGs), supporting the assumption
plastid signal is also required in the dark, before first illumina- that the haem pool (or a fraction of it) represents a retrograde
tion (not observable with NF) (Pfannschmidt and Link, 1997; signal (Woodson et al., 2011).
Sullivan and Gray, 1999); and (iii) the plastid signal appears to The gun1 mutant is an exception within the gun mutant col-
be required in the first 48–72 h after germination since later lection since it was found to be the only mutant that exhibits
application of the inhibitors was found to no longer be effec- decoupling of nuclear gene expression not only upon NF but
tive. It was therefore concluded that the plastid signal involves also upon Lin treatment. GUN1 was identified to encode a
a product of early PGE that affects a specific developmental plastid pentatricopeptide repeat (PPR) protein that contains
step essential for the initiation of chloroplast biogenesis (Gray a small MutS-related domain (Koussevitzky et al., 2007; Cot-
et al., 2003). tage et al., 2010). PPR proteins are very numerous in plants,
Further progress in the understanding of the plastid signal and plastid PPR proteins are known to be involved in various
came from a mutant screen in Arabidopsis that revealed that steps of RNA metabolism (reviewed in Schmitz-Linneweber
the nuclear gene expression could be decoupled from the and Small, 2008). Furthermore, GUN1 was reported to in-
developmental state of the plastid. These genomes uncoupled teract with many proteins, probably being involved in protein
(gun) mutants displayed a partial de-repression of LHCB ex- homeostasis (Tadini et al., 2016), and also to bind tetrapyrroles
pression despite photo-oxidation of plastids by NF applica- in order to reduce haem and protochlorophyllide synthesis
tion (Susek et al., 1993). In subsequent years the gun mutants (Shimizu et al., 2019), as well as enzymes of the TBP pathway7110 | Liebers et al.
(Tadini et al., 2019). In fact, the precise molecular function (iv) Chloroplast biogenesis of Angiosperms is under strong
of GUN1 is still under investigation, but it appears to act as light control, and anterograde signalling is a major deter-
a central integrator of several signalling routes (see below for minant in the build-up of the photosynthetic apparatus
more details) (Koussevitzky et al., 2007; reviewed in Colombo during early seedling development as well as in meristems
et al., 2016). of fully grown plants. Since retrograde biogenic signals are
In sum, the gun mutant screen had two important outcomes, equally important in chloroplast biogenesis, an interaction
even though the precise molecular involvement in retrograde sig- between anterograde and retrograde signalling appears
nalling of these mutants is still debated: (i) it demonstrated that the likely, but the potential interactions remain to be under-
plastid signal can be genetically interrupted (thus the low expres- stood, representing an interesting field for future research.
sion of nuclear target genes is not simply a general negative feed- (v) The most important question is probably the one re-
back effect of the dysfunctional plastids in the affected cells) and garding the physical nature of the biogenic signal(s) that
Downloaded from https://academic.oup.com/jxb/article/73/21/7105/6674789 by guest on 13 December 2022
(ii) it showed that genetic defects causing perturbations at different is(are) transmitted by the plastid. Unambiguous identifica-
sites in the plastid result in the same nuclear response (suggesting tion of one or several signal molecules would provide not
either merging of several plastid signals into one pathway or a mu- only a better mechanistic understanding of the signalling
tual molecular influence of the different affected processes). route/network, but also a means for further experimental
approaches.
Central open questions in the understanding of The answers to each of these topics are tightly connected and
biogenic signals the current progress in our understanding in each area, there-
fore, is now discussed in an integrated manner.
The initial studies paved the way to more comprehensive
approaches in recent years, and the use of plant models with
dysfunctional plastids (either induced genetically or pharma- Gene targets and qualitative character of plastid signals
ceutically) in combination with physiological assays became a Initial studies of retrograde biogenic signals focused on the
standard approach for studying biogenic signals in depth. The expression behaviour of a limited number of PhANGs and
research field has seen tremendous progress in the last 15 years the proposals of the positive or negative character of these
mostly focusing on five central aspects. signals in early models based largely on the observed expres-
sion changes in these reporter systems. A major problem in
(i) Initial studies used only a very limited number of nu- this context is the fact that only relative expression values
clear genes as the reporter for the activity of the plastid could be analysed and the true action of the signal remained
signal, mostly RBCS, LHCB, and a few other PhANGs. ambiguous; for example, low expression of PhANGs upon a
This term became a common synonym for the target of block in chloroplast biogenesis in comparison with normal
biogenic signals. In between it became clear that many development could indicate either active repression (a nega-
other genes (also encoding non-plastid-localized proteins) tive signal) or the lack of promotion (a positive signal) by the
are also under biogenic control and that in addition other dysfunctional plastid. At the same time, only a few transcrip-
levels of gene expression such as translation or protein tional regulators could be assigned a potential role in retro-
stability are affected. This indicates that retrograde signals grade signalling, and mechanistic regulatory models remained
represent a major regulatory determinant in early seedling highly speculative, leading to a controversial discussion about
development, but its precise position within the regula- the positive or negative character of plastid signals (reviewed
tion networks responsible is far from understood. in Pfannschmidt, 2010; Terry and Smith, 2013). Thus, without
(ii) Another discussion focused on the qualitative action of a detailed understanding of the molecular mechanism, a
the plastid signal(s), namely whether lower nuclear gene hypothesized positive or negative action of (a) plastid signal(s)
expression in plants with dysfunctional plastids represents must be regarded as a purely conceptual categorization that,
the result of (1) active negative signalling (that might be however, might be helpful in generating working hypotheses.
de-repressed in gun mutants, a molecular reaction in re- Furthermore, the ‘gun phenotype’ as the molecular read-out in
cent studies often called the gun phenotype) or (2) the the presence of NF or Lin was used for many mutants to test
lack of (a) positive signal(s) (not produced by the plastid the potential (positive or negative) involvement of the respec-
due to its dysfunction). tive inactivated component in retrograde signalling. This way
(iii) The origin of biogenic retrograde signals has been attrib- of classifying mutants as ‘new’ gun mutants, however, produced
uted to two major processes, TBP and PGE, where the a number of cases with controversial results, leading to much
latter is comprised of two subprocesses: transcription and debate about whether or not a specific component is part of
translation.The question arose of whether all processes in- retrograde control (for more details, see further below). The
itiate independent signals or whether they all converge study of the expression behaviour of single genes without ad-
into one common signalling pathway as proposed for the ditional experimental lines, therefore, should be highly disfa-
function of GUN1, a debate not yet finished. voured.Retrograde biogenic signals | 7111
Much progress in this aspect was obtained by using full ge- cytosolic protein homeostasis exists and that retrograde control
nome transcriptomic approaches in order to reveal the true is strongly exerted at this level (for more details, see below)
regulatory impact of biogenic signals during early seedling de- (reviewed in Wu and Bock, 2021). A sole focus on transcript
velopment of Arabidopsis. Virtually thousands of genes were changes might therefore miss important aspects and targets of
found to be changed in expression when chloroplast biogenesis retrograde control for a given gene, and parallel analysis at dif-
was blocked genetically or by NF or Lin treatments (Ruckle ferent levels of expression appears recommendable in future
et al., 2012; Grubler et al., 2017; Page et al., 2017b), indicat- analyses.
ing that plastid signals (i) represent a major regulatory ele-
ment in this early developmental stage, (ii) affect many more Functional connections between TBP- and PGE-derived
gene groups other than PhANGs, and (iii) also control genes signals
that encode products not located in plastids. The last aspect The TBP pathway is completely localized within plastids; how-
Downloaded from https://academic.oup.com/jxb/article/73/21/7105/6674789 by guest on 13 December 2022
is largely unexplored, but appears highly interesting with re- ever, all enzymes of this pathway are encoded in the nucleus
spect to potential influences of plastid signals on overall plant and must be imported from the cytosol (reviewed in Tanaka
morphology (e.g. leaf shape) (reviewed in Tiller and Bock, and Tanaka, 2007). Thus, establishment of the TBP enzyme ac-
2014) and metabolism. A meta-analysis comparing datasets tivities does not require direct PGE activity. In turn, all PGE
from these studies identified a core module of nuclear genes activities known so far can be performed without requiring
responding under all three conditions (Grubler et al., 2021). any product from TBP. At first sight, therefore, one could as-
It is comprised of 152 genes that could be subdivided into sume that both processes are independent from each other and
seven major functional categories comprising photosynthesis, that retrograde biogenic signals emerging from them follow
carbohydrate metabolism and transport, redox regulation, de- independent and distinct signalling routes.This, however, is not
velopment, transcription, proteins and stress, as well as lipids the case since TBP and PGE affect each other in multiple ways.
and hormones, representing the broad functional impact of bi- This mutual influence starts already with the import of the
ogenic signals at this stage of seedling development. The group TBP enzymes. The TOC/TIC machinery involves one sub-
of PhANGs appeared to be regulated highly uniformly in all unit that is encoded by ycf1, a plastid class II gene transcribed
three approaches, while other groups revealed more differential by NEP and PEP (Fig. 1) (Zhelyazkova et al., 2012; reviewed
responses, suggesting that one and the same plastid signal can in Lyska et al., 2013). Furthermore, the plastid class III gene
cause gene-specific effects of both a positive and negative re- ycf2 that is transcribed only by NEP encodes a component of
sponse. A qualitative classification of retrograde signals thus can the AAA-ATPase import motor complex that interacts with
be done only in a gene-specific manner. the TIC complex and drives the ATP-dependent translocation
Key activators of PhANG expression are GLK1 and GLK2 of pre-proteins over the inner membrane. Ycf2 contains an
(Fitter et al., 2002; Waters et al., 2009; reviewed in Waters and NAD-malate dehydrogenase activity enabling ATP production
Langdale, 2009). GLK1 in particular appears to be a target also in the dark or in non-photosynthetic plastids (such as in
for retrograde control since it was also found within the core meristems or in dark-grown seedlings) (Kikuchi et al., 2018).
module displaying an expression behaviour like the PhANGs. Thus, establishment and function of the TOC/TIC complex is
Interestingly GLK1- and GLK2-overexpressing Arabidopsis dependent on the activity of PGE and, as a consequence, also
lines exhibit a gun phenotype (Leister and Kleine, 2016; Martin the import of all cytosolic pre-proteins. There is still some un-
et al., 2016), and recent data indicate that GLK1 acts down- certainty about the possible existence of different subtypes of
stream of GUN1-mediated retrograde signalling, suggesting import complexes that could possess different substrate speci-
the action of a tentative GUN1/GLK1 module. This module ficities, and it is not yet fully understood whether or not Ycf1
is proposed to control not only PhANG expression in the nu- is required for the import of all nuclear-encoded components
cleus but also developmental processes in skotomorphogenesis (de Vries et al., 2015; reviewed in Nakai, 2015).
and seedling de-etiolation, providing a potential way in which Another important link between PGE and TBP relates to
chloroplast biogenesis and early seedling development could the production of the entrance substrate of the TBP pathway,
be coordinated (Hernandez-Verdeja et al., 2022; Veciana et al., the glutamyl-tRNA (the amino acid glutamate linked to
2022). In addition, a recent study was able to demonstrate a tRNA-Glu/trnE) that serves as precursor for the production of
strong post-transcriptional effect on GLK1 protein accumu- ALA (von Wettstein et al., 1995). The proper timely and quan-
lation. While GLK1 mRNA accumulates to normal levels titative expression of the trnE gene is absolutely essential for
in NF- or Lin-treated gun1-101 mutants, no GLK1 protein sufficient functioning of the TBP pathway (Fig. 1). The trnE
accumulated, suggesting that GLK1 is controlled by plastid sig- gene is transcribed by the PEP enzyme that is the dominant
nals at the protein level. Additional experiments with prote- transcription machinery for the expression of plastid tRNA
asome inhibitors could confirm this assumption (Tokumaru genes (Williams-Carrier et al., 2014). PEP requires the interac-
et al., 2017), pointing to cytosolic protein homeostasis as an tion with the nuclear-encoded sigma factor 2 (Sig2) for recog-
important target for retrograde signals. Several recent studies nition of the trnE promoter, as demonstrated by the pale-green
indeed confirmed that a connection between plastid and phenotype of SIG2 inactivation mutants (Hanaoka et al., 2003;7112 | Liebers et al.
Downloaded from https://academic.oup.com/jxb/article/73/21/7105/6674789 by guest on 13 December 2022
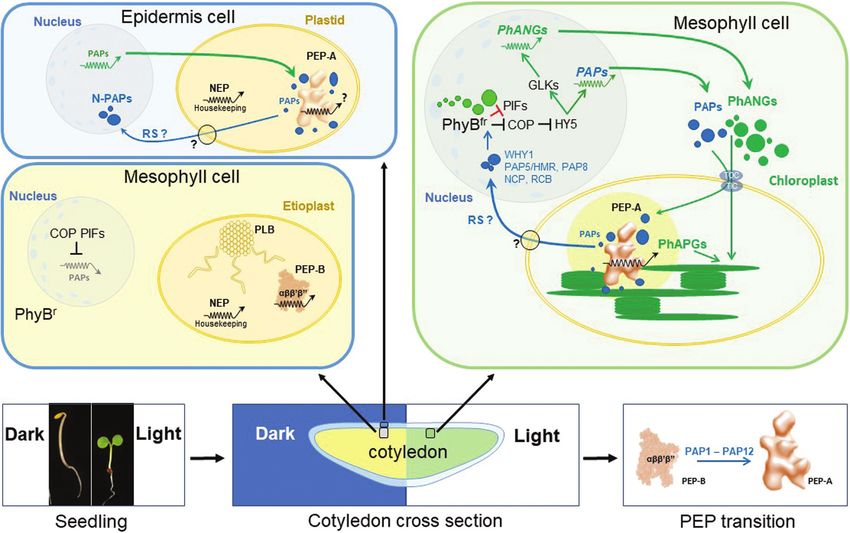
Fig. 1. Structural and functional integration of essential plastid processes during chloroplast biogenesis. The scheme depicts an overview of the
molecular connections between the major biosynthetic processes within the plastid required for the build-up of the photosynthetic apparatus during
chloroplast biogenesis. Light orange boxes: transcriptional activities. Dark orange boxes: processes related to protein synthesis and homeostasis. Yellow
boxes: envelope-located processes. Green boxes: processes directly involved in formation of photosynthetic structures. Grey boxes: genes transcribed
by nuclear- and plastid-encoded RNA polymerases (NEP and PEP). Green arrows: delivery of proteins by import through TOC–TIC from the cytosol. Light
orange arrows: transcript maturation. Black arrows: delivery of plastid-synthesized products to subsequent processes. Broken black arrows: additional
transcriptional involvement of PEP activity. Orange arrows: translation of plastid transcripts at 70S ribosomes. Red arrow: negative feedback from the
trnE pool. Squares indicate mutant alleles identified in Arabidopsis resulting in defects of the respective process in which the squares are located. The
phenotype caused by the genetic defect is indicated by the colour code of each square. The corresponding key is given in the right margin. For gene
identities of the alleles, refer to Table S1 at Zenodo. For details see main text.
Woodson et al., 2013). PEP transcription thus performs strong ment of a high transcriptional PEP activity, especially during
regulatory control over the substrate availability and the me- the critical early phases in the onset of chloroplast biogen-
tabolite flux through the TBP pathway.Therefore, SIG mutants esis. Plastid transcription and plastid translation, therefore, are
have also been regarded as substitute mutants for TBP retro- highly intertwined, generating a chicken and egg problem
grade signalling (Woodson et al., 2013). Interestingly, a feed- concerning the mutual dependency.
back control has been proposed in which increasing levels of The faithful generation of a photosynthetic apparatus
trnE production lead to down-regulation of NEP activity by during chloroplast biogenesis requires a high coordination
physical interaction, thereby providing a means for the shift in the production of photosynthesis proteins, chlorophylls,
from NEP to PEP activity during early chloroplast biogenesis and thylakoid membranes. Any disturbance of this coordina-
(Hanaoka et al., 2005). tion can generate severe consequences, leading eventually to
The trn genes transcribed by PEP are, in addition, essential an interruption of the complete process as observed in var-
for effective functioning of plastid translation (Fig. 1). Further- ious mutants with defects in distinct regulatory steps of plastid
more, PEP transcribes rrn genes as well as genes for the large gene expression, tetrapyrrole biosynthesis, or lipid formation
and small ribosome subunit proteins (rpl and rps, respectively), (see Fig. 1 and Table S1 at Zenodo https://doi.org/10.5281/
providing structural components of the 70S ribosomes, and zenodo.6674852, Liebers et al., 2022). The plastid-encoded
last but not least a high number of photosynthesis genes. Since genes accD and clpP each encode a single subunit of the mul-
the RNA polymerase (rpo) core subunits of PEP are plastid tisubunit acetyl-CoA carboxylase (ACC) and Clp protease
encoded, in turn plastid translation is essential for the establish- (ClpP) complexes, respectively. The ACC complex synthesizesRetrograde biogenic signals | 7113
malonyl-CoA, an essential precursor for plastid lipid produc- this mutant type the availability of pigment-binding proteins
tion (required for thylakoid lipid production) while the ClpP or complexes (assuming full functionality of the TBP) probably
complex is required for the maintenance of a proper protein determines the degree of colouration, also allowing gradual
homeostasis (proteostasis) in plastids (required to remove dam- variations, while defects in pigment synthesis appear to result
aged or misfolded proteins). Correct expression and assembly more in all-or-nothing phenotypes. Thus, during the build-up
of the plastid-encoded subunits with the nuclear-encoded of the photosynthetic apparatus, expression of TBP-associated
subunits for both complexes have been shown to be achieved genes as well as PhANGs requires a high coordination in order
by retrograde signalling (Babiychuk et al., 2011; Llamas et al., to avoid mismatches in the relative abundance of their prod-
2017). These processes thus are most likely to be coupled to ucts.
TBP (see below for more details). This is supported by the
observation that the transcriptional regulation of the nuclear- Functional connections between biogenic signals and
Downloaded from https://academic.oup.com/jxb/article/73/21/7105/6674789 by guest on 13 December 2022
encoded key enzymes of thylakoid membrane lipid synthesis, light regulation
MGDG1 and DGD1, is coordinated with chlorophyll biosyn- Due to the strong impact of light on the biogenesis of chlo-
thesis involving control by light and plastid signals (Kobayashi roplasts, it appears very likely that retrograde signals and the
et al., 2014). light signalling network do interact (reviewed in Hernan-
In sum, PGE and TBP pathways are tightly interlinked, and a dez-Verdeja et al., 2020). Indeed, already early investigations
clear-cut separation between retrograde signals from PGE and of nuclear gene expression reported a convergence of signals
TBP often remains difficult. In fact, this close linkage may ex- from plastids and the light regulation network at the level of
plain why many mutants with defects in diverse steps within or promoter usage (Kusnetsov et al., 1996). Later transcriptomic
related to PGE or TBP reveal similar phenotypes with defects approaches indicated that there exists a strong mutual influence
in proper chloroplast biogenesis. To obtain an overview of the between light and retrograde signals even at the system level
distribution of functional defects related to these phenotypes, (Ruckle et al., 2007, 2012; Ruckle and Larkin, 2009; Grubler
we performed a database and literature survey and identified et al., 2017), and more recent studies confirmed a convergence
122 mutants in Arabidopsis that exhibit an albino or pale- of both regulatory networks (Martin et al., 2016). In particular,
green phenotype sometimes with spatial (variegation) or tem- it was proposed that the GUN1/GLK1 module acts antago-
poral (virescence) variation (Table S1 at Zenodo). This survey nistically to the phytochrome/PIF signalling hub and inhibits
might miss non-viable mutants leading to embryo lethality or the proper development of cotyledons when chloroplast for-
mutants in which the gene defect can be compensated by pa- mation is prevented or compromised by the application of Lin
ralogous genes. Therefore, the total number of critical compo- or high light stress. Biogenic signals from plastids thus can act
nents is likely to be higher. Many of these mutant alleles have as developmental triggers that impact light-controlled seedling
orthologous alleles in maize which may survive because of the morphogenesis besides their role as PhANG expression regula-
nuclear location of the ACCD gene in monocotyledonous tors (Martin et al., 2016).
plants (Belcher et al., 2015). In all mutant types, but especially The impact of light on the total transcriptome during seed-
in albino mutants, defects in PGE were found to be domi- ling development is massive. Almost a third of all genes exhibit
nant in number (Fig. 1). Detailed functional analyses indicate significant expression changes upon the light-induced shift
that intrinsic components of both the transcription and trans- from skoto- to photomorphogenesis (Ma et al., 2001). The
lation complexes as well as plastid-localized regulatory factors resulting molecular and developmental changes appear rap-
controlling their respective expression are involved. The PGE idly, within hours, rendering it difficult to dissect the respective
system thus exhibits a surprising low capability to compensate regulatory impact of each signalling system. However, recent
for functional deficiencies despite its high complexity, pointing studies on slowly greening Arabidopsis single-cell cultures
to a low level of redundancy in the system. This might be (Dubreuil et al., 2018) as well as careful analyses along the de-
explained by the endosymbiotic origin of plastids that exerts a velopmental cell gradient in wheat leaves (Loudya et al., 2021)
high selective pressure on autotrophy which, in turn, may lead uncovered a bi-phasic programme for chloroplast biogenesis.
to a high specialization in regulation and function of factors Although the two systems and the resulting observations are
involved in PGE. Consequently, such a high specialization in not fully coherent, a simplified two-step model for chloroplast
PGE results in a low flexibility towards defects, dysfunctions, or biogenesis can be proposed. The first phase begins with organ-
functional imbalances, and a proper stoichiometric production elle proliferation followed by establishment and build-up of the
of the components required for chloroplast biogenesis requires plastid genetic system in meristematic and non-differentiated
a tight coupling to the nuclear gene expression system. Pre- cells. After that, a second build-up phase follows in which the
cisely that is achieved by the biogenic retrograde control path- photosynthetic apparatus is established within more differenti-
ways. Defects in pigment biosynthesis were found to be the ated cells. Any severe disturbance of the first phase thus will
second largest group causing albinism, but apparently are less prevent a successful transition into the second phase, result-
important in pale-green mutants where defects in photosyn- ing in developmentally arrested plastids unable to perform ef-
thesis become more important (Fig. 1). This implies that in ficient photosynthesis. This bi-phasic programme provides a7114 | Liebers et al.
reasonable explanation for the observation that application of More importantly, besides metabolites, a number of pro-
inhibitors of prokaryotic gene expression is effective only in a teins may act as retrograde signalling compounds, although
small time window after germination (see above). currently no specific protein export from plastids could be
Interestingly, gymnosperms are able to perform greening demonstrated. Nevertheless, a steadily increasing number of
already in the dark since they possess a different type of dually localized nucleo-plastidic proteins has been identified
protochlorophyllide reductase enzyme that works light inde- (reviewed in Krause and Krupinska, 2009), and for several of
pendently (Forreiter and Apel, 1993). Using pine seedlings as a them a direct trafficking from the plastid towards the nucleus
natural tool to separate plastid signals from the light signalling has been proposed (see below for a detailed discussion). These
network, a recent study could provide independent evidence proteins are reported to affect the expression of PhANGs and
for the assumption that retrograde control of nuclear genes other genes either directly or indirectly. Two general scenar-
evolved before the light signalling network took control over ios have been proposed so far: (i) plastid-localized proteins or
Downloaded from https://academic.oup.com/jxb/article/73/21/7105/6674789 by guest on 13 December 2022
photomorphogenesis (Hills et al., 2015). Retrograde biogenic protein complexes that mediate or support the transition of a
signals thus need to be considered as an intrinsic component of retrograde signal (subsequently referred to as ‘stationary me-
the basal developmental programmes triggering early seedling diator proteins’) and (ii) proteins that act as genuine signal(s)
development rather than being a part of the light signalling by leaving the plastid or its surface and moving to the nucleus
network. They, however, provide a potential connection point (referred to as ‘mobile signalling proteins’).
that couples both signalling networks.
Stationary mediator proteins
The physical nature of retrograde biogenic This class of proteins resides inside the plastid compartment.
signals These proteins may move within the compartment, but never
leave it. The most prominent representative of this protein
Plastids are involved in most metabolic and biosynthesis pathways
group is GUN1, a highly investigated receiver and transmitter
of plant cells and, therefore, possess a huge variety of envelope-
protein for retrograde signals which interacts with several other
localized transporters that are involved in shuttling of meta-
mediator proteins. The GUN1 transcript accumulates to sig-
bolic intermediates between the cytosol and the plastid stroma
nificant amounts in all tissues, but the protein is found to ac-
(reviewed in Rolland et al., 2012). In virtually all developmental
cumulate only in cotyledons during proplastid to chloroplast
phases plastids are linked to cytosolic processes by exchange of
transition and in the SAM. Furthermore, it accumulates in re-
molecules providing many potential candidates that may act as
sponse to NF treatments; thus the protein is present precisely
retrograde signals.This tight connection with the cytosol reflects
at the developmental stage in which biogenic signals are most
the deep integration of plastids into the cellular metabolism that
important (Wu et al., 2018). Co-immunoprecipitation, yeast
was established during endosymbiosis, and it has been discussed
two-hybrid, and proteomic analyses indicated that GUN1 is
that even metabolite signatures may have a signalling function
functionally connected to or involved in (i) the TBP, (ii) the
(reviewed in Pfannschmidt, 2010). So far a role as a retrograde
PGE, (iii) plastid proteostasis, and (iv) protein import (Tadini
signal has been attributed to metabolites mostly in the context of
et al., 2016; Wu et al., 2019; Wang et al., 2021) (see Fig. 2).
operational signalling.The functions of methyl-erythritol-cyclo-
diphosphate (MEcPP), phospho-adenosine-phosphate (PAP), (i) GUN1 has been reported to bind haem and other porphy-
and apocarotenoids as signalling molecules in stress responses rins, as well as enzymes of the TBP, such as the D subunit
have been studied in great detail (reviewed in Chi et al., 2013; of Mg-chelatase (CHLD) and FC1, and thus may affect
Chan et al., 2016; de Souza et al., 2016), and only a small number the flux through the TBP, resulting in changes in the TBP-
of observations suggest that metabolites may also be involved in dependent retrograde signalling (reviewed in Colombo
biogenic retrograde signalling. Of special interest in this context et al., 2016; Shimizu et al., 2019). Additionally, GUN1-
are sugars, anthocyanins, and the various precursor molecules for mediated signals influence the expression of GLK1 and
plant hormones that are produced in plastids, such as those for GLK2, which modifies the expression of key TBP genes
ABA or jasmonates (Cottage et al., 2010; Richter et al., 2020). (Kakizaki et al., 2009). Recent studies have extensively
Chloroplast biogenesis is well known to be strongly promoted analysed in particular the impact of GUN1-mediated sig-
by cytokinins (Cortleven et al., 2016) and an interplay with nals on GLK1, leading to the proposal of a GUN1/GLK1
auxin and light (Kobayashi et al., 2012). Well-defined networks module (compare above), that is of importance for both
that describe the connections between light, hormones, and nu- seedling development and chloroplast biogenesis (Martin
clear transcriptional regulators in early chloroplast biogenesis et al., 2016). Moreover, GUN1 is proposed to aid plastid
have been developed in recent years (reviewed in Cackett et al., protein import (see below), and proteins being imported
2022). Elucidating the input of biogenic retrograde signals into in a GUN1-dependent manner include TBP enzymes,
these networks will be of highest interest for the understanding thus underpinning an influence of GUN1 on the level
of chloroplast biogenesis, representing a promising prospect. of haem and Mg-proto-IX (Wu et al., 2019). This mayYou can also read

















































