B CELL RESPONSES TO HUMAN FLAVIVIRUS VACCINATION AND SARS-COV-2 INFECTION - From the Department of Medicine, Huddinge Karolinska Institutet ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
From the Department of Medicine, Huddinge
Karolinska Institutet, Stockholm, Sweden
B CELL RESPONSES TO HUMAN
FLAVIVIRUS VACCINATION AND
SARS-COV-2 INFECTION
Tyler Sandberg
Stockholm 2022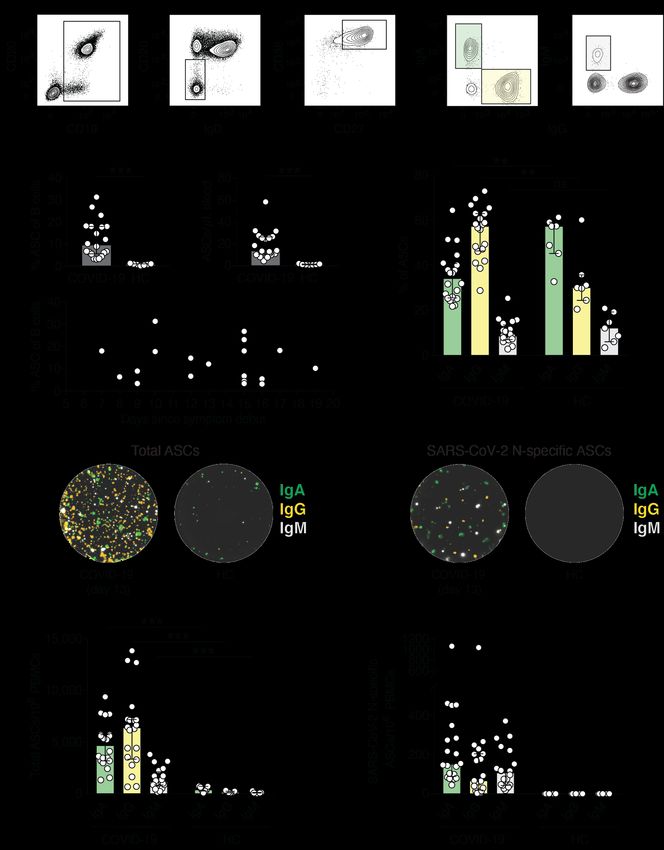
All previously published papers were reproduced with permission from the publisher. Published by Karolinska Institutet. Printed by Universitetsservice US-AB, 2022 © Tyler Sandberg, 2022 ISBN 978-91-8016-437-5 Cover illustration: Created in Adobe Illustrator by Tyler Sandberg
B cell responses to human flavivirus vaccination and
SARS-CoV-2 infection
THESIS FOR DOCTORAL DEGREE (Ph.D.)
By
Tyler Sandberg
The thesis will be defended in public at Karolinska Institutet ANA-Futura, Alfred Nobels Allè
8, 18th of March, 2022 at 9:30.
Principal Supervisor: Opponent:
Professor Hans-Gustaf Ljunggren Professor Tomas Bergström, MD-PhD
Karolinska Institutet University of Gothenburg
Department of Medicine, Huddinge Institute of Biomedicine
Center for Infectious Medicine Department of Infectious Diseases
Co-supervisor(s): Examination Board:
Kim Blom, PhD Associate Professor Guro Gafvelin
Karolinska Institutet Karolinska Institutet
Department of Medicine, Huddinge Department of Clinical Neuroscience
Center for Infectious Medicine Therapeutic Immune Design Unit
Professor Karin Loré Associate Professor Anna Lundgren
Karolinska Institutet University of Gothenburg
Department of Medicine, Solna Institute of Biomedicine
Division of Immunology & Allergy Department of Microbiology & Immunology
Associate Professor Anna Överby
Umeå University
Department of Clinical Microbiology
Section of Virology
This thesis is dedicated to my family. I wouldn’t have made it here without you.
POPULAR SCIENCE SUMMARY OF THE THESIS No other medical intervention has had such a significant impact on human health as that of vaccines. Vaccination has become a topic of interest since the debut of the COVID-19 pandemic. Vaccines are designed to teach the immune system to recognize critical parts of a pathogen so that if you were ever exposed to it in real life, your immune cells and antibodies could prevent infection or severe disease. There are several different vaccine platforms used today, the most common using weakened live viruses or inactivated viruses in vaccine formulations. During the COVID-19 pandemic, mRNA vaccine technology was approved for the first time in humans, saving an estimated 500,000 lives already. One of the most effective and successful vaccines in history is the yellow fever virus (YFV) vaccine. First developed by Max Theiler, who later won the Nobel Prize in Medicine for his discovery, the vaccine uses a weakened strain of the virus that, upon administration, leads to a mild infection. The immune events following vaccination lead to sustained antibody titers, memory T cells that can kill virus-infected cells and memory B cells that can quickly produce antibodies if infected with the virus. The YFV vaccine has long been considered a model vaccine as it provides lifelong immunity from a single dose. Scientists have long used the vaccine to learn more about viral infections and the immune responses following vaccination. In this thesis, we used the YFV vaccine to understand how human B cells respond to vaccination. In Paper I, we found that the YFV vaccine elicits a strong B cell response during the first two weeks after vaccination. The vaccine also leads to the development of protective levels of antibodies, and we were able to identify virus-specific memory B cells circulating in the donor samples by the end of the study. Similarly in Paper II, B cell responses to other vaccines including the Tick-borne encephalitis (TBEV) vaccine and Japanese encephalitis (JEV) vaccine were also studied. An important question we wanted to investigate in Paper II was how human B cell responses would be affected if the YFV vaccine was given at the same time as either TBEV or JEV vaccines. We found that most healthy donors in the studies developed antibodies against the respective viruses after vaccination with no differences in donors receiving two vaccines compared with donors receiving only one of the vaccines. At the beginning of the COVID-19 pandemic, our focus shifted towards studying human immune responses to SARS-CoV-2 infection in hospitalized patients. In Papers III and IV, we found that COVID-19 patients had strong early B cell and T cell responses with high levels of antibody responses. These early immune events led to detectable virus-specific memory B cells and memory T cells in addition to antibodies at 5- and 9-months after infection, likely contributing to protection from reinfection or severe disease. The findings in this thesis contribute to our understanding of how, specifically, B cells respond to vaccination and infection. This knowledge can be used to help design new vaccines and vaccination strategies or even improve upon current vaccines to prevent severe human infection and disease.
ABSTRACT Viral infections pose a major threat to global heath. As specific antiviral treatments are lacking against many human viruses, vaccination is the most effective medical intervention to prevent severe disease and death. Delineating the immune events following viral vaccination and infection can help in the design of new vaccines and therapeutics. The aims of this thesis were to characterize human B cell responses following yellow fever virus (YFV) vaccination (Paper I), following concomitant vaccination with YFV and the Tick-borne encephalitis virus (TBEV) vaccine or Japanese encephalitis virus (JEV) vaccine (Paper II), and during acute and convalescent SARS-CoV-2 infection (Papers III and IV). In Paper I, healthy volunteers were vaccinated with the YFV vaccine and blood samples were taken at up to five time points afterwards to characterize the magnitude, kinetics, and specificity of the humoral immune response. Activation in the Th1-polarized circulating T follicular helper cell population was observed 7 days following vaccination, coinciding with increased germinal center activity as measured by serum CXCL13 levels. Peak YFV-E specific plasmablast expansion was observed at day 14 following vaccination. Additionally, the frequencies of IgG+ plasmablasts at day 14 correlated with day 90 neutralizing antibody (nAb) titer magnitude, suggesting that plasmablasts may be used as an early marker indicating later protective immunity. YFV-E specific memory B cells were also detectable at day 28 and 90 as well as protective titers of nAbs. These findings provide insights into immune events that lead to the development of B cell immunity following YFV vaccination. In Paper II, we investigated the feasibility and effectiveness of concomitant vaccination with YFV vaccines and either TBEV or JEV vaccines. 145 healthy volunteers were recruited into a prospective open label, non-randomized clinical trial and received either YFV, TBEV, or JEV vaccines only or YFV vaccine with either TBEV or JEV vaccines. Blood and serum samples were taken at baseline and up to ten timepoint following vaccination. The development of virus-specific nAbs was not affected by concomitant vaccination when comparing the vaccine cohorts. Importantly adverse events were mild and not affected by concomitant vaccination suggesting that the vaccination strategy should be considered effective and safe. In Papers III and IV, early immune events and ultimately immune memory was investigated in COVID-19 patients during and after hospitalization. Increased germinal center activity with a Th1-polarized circulating T follicular helper cell activation was observed that coincided with SARS-CoV-2-specific expanded antibody secreting cells during acute COVID-19. The majority of patients also had detectable levels of nAbs during acute disease. In Paper IV, SARS-CoV-2-specific nAb titers persisted in patients at 5- and 9-months post infection as well as virus-specific polyfunctional memory T cells and memory B cells regardless of COVID-19 severity during hospitalization. Together, the findings in this thesis contribute to our understanding of humoral responses to different types of flavivirus vaccines as well as infection with SARS-CoV-2.
LIST OF SCIENTIFIC PAPERS
I. Sandberg JT, Ols S, Löfling M, Varnaitė R, Lindgren G, Nilsson O, Rombo
L, Kalén M, Loré K, Blom K, and Ljunggren HG. Activation and Kinetics of
Circulating T Follicular Helper Cells, Specific Plasmablast Response and
Development of Neutralizing Antibodies Following Yellow Fever Virus
Vaccination. Journal of Immunology, 2021, 207 (4), pp. 1033-1043.
II. Sandberg JT, Löfling M, Varnaitė R, Emgård J, Al-Tawil N, Lindquist L,
Gredmark-Russ S, Klingström J, Loré K, Blom K, and Ljunggren HG. Safety
and immunological responses of concomitant vaccination with flavivirus
vaccines: results from an open label non-randomized clinical trial. Manuscript.
III. Varnaitė R, Garcia M, Glans H*, Maleki KT*, Sandberg JT*, Tynell J, Christ
W, Lagerqvist N, Asgeirsson H, Ljunggren HG, Ahlén G, Frelin L, Sällberg
M, Blom K, Klingström J, and Gredmark-Russ S. Expansion of SARS-CoV-
2–Specific Antibody-Secreting Cells and Generation of Neutralizing
Antibodies in Hospitalized COVID-19 Patients. Journal of Immunology, 2020,
205 (9), pp. 2437-2446. *Authors contributed equally.
IV. Sandberg JT*, Varnaitė R*, Christ W, Chen P, Muvva JR, Maleki KT, Garcia
M, Dzidic M, Folkesson E, Skagerberg M, Ahlén G, Frelin L, Sällberg M,
Eriksson LI, Rooyackers O, Sönnerborg A, Buggert M, Björkström NK,
Aleman S, Strålin K, Klingström J, Ljunggren HG, Blom K, Gredmark-Russ
S, and The Karolinska COVID-19 Study Group. SARS-CoV-2-specific
humoral and cellular immunity persists through 9 months irrespective of
COVID-19 severity at hospitalization. Clinical & Translational Immunology,
2021, 10: e1306. *Joint first authors.SCIENTIFIC PAPERS NOT INCLUDED IN THE THESIS
SI. Blom K, Sandberg JT, Loré K, and Ljunggren HG. Prospects for induction of CD8 T
cell-mediated immunity to Zika virus infection by yellow fever virus vaccination.
Journal of Internal Medicine. 2017, 282 (3) pp. 206-208.
SII. Blom K, Cuapio A, Sandberg JT, Varnaite R, Michaëlsson J, Björkström NK,
Sandberg JK, Klingström J, Lindquist L, Gredmark Russ S, Ljunggren HG. Cell-
Mediated Immune Responses and Immunopathogenesis of Human Tick-Borne
Encephalitis Virus-Infection. Frontiers in Immunology. 2018, 9:2174.
SIII. Cornillet M, Strunz B, Rooyackers O, Ponzetta A, Chen P, Muvva JR, Akber M,
Buggert M, Chambers BJ, Dzidic M, Filipovic I, Gorin J-B, Gredmark-Russ S, Hertwig
L, Klingström J, Kokkinou E, Kvedaraite E, Lourda M, Mjösberg J, Maucourant C,
Norrby-Teglund A, Parrot T, Perez-Potti A, Rivera-Ballesteros O, Sandberg JK,
Sandberg JT, Sekine T, Svensson M, Varnaite R, , Eriksson LI, Aleman S, Strålin K,
Ljunggren H-G and Björkström NK (2021), COVID-19 specific metabolic imprint
yields insights into multi organ-system perturbations. European Journal of
Immunology. https://doi.org/10.1002/eji.202149626CONTENTS
1 INTRODUCTION ......................................................................................................... 3
1.1 Adaptive immunity............................................................................................... 3
1.1.1 B cells ....................................................................................................... 3
1.1.2 CD4+ T cells ............................................................................................. 4
1.2 Flaviviruses & flavivirus vaccines ....................................................................... 5
1.2.1 YFV .......................................................................................................... 6
1.2.2 TBEV ....................................................................................................... 6
1.2.3 JEV ........................................................................................................... 6
1.2.4 Flavivirus vaccines................................................................................... 6
1.3 SARS-CoV-2...................................................................................................... 10
1.3.1 Clinical manifestation ............................................................................ 10
2 RESEARCH AIMS ..................................................................................................... 11
3 MATERIALS AND METHODS ................................................................................ 13
3.1 Ethical considerations ........................................................................................ 13
3.2 Sample collection and processing ...................................................................... 14
3.3 Flow cytometry .................................................................................................. 14
3.4 Serological analyses ........................................................................................... 14
3.4.1 ELISA .................................................................................................... 15
3.4.2 Neutralization tests................................................................................. 15
3.5 FluoroSpot Assays ............................................................................................. 16
3.5.1 B cell FluoroSpot ................................................................................... 16
3.5.2 T cell FluoroSpot ................................................................................... 17
3.6 Analysis of soluble markers ............................................................................... 18
3.6.1 ELISA .................................................................................................... 18
3.6.2 Multiplex immunoassay......................................................................... 18
3.6.3 Proteomic analysis – Olink immune response panel............................. 19
3.6.4 Viral load – real-time PCR .................................................................... 19
3.7 Statistics.............................................................................................................. 19
4 RESULTS & DISCUSSION ....................................................................................... 21
4.1 B cell immune responses following flavivirus vaccination............................... 21
4.1.1 B cell and T cell activation .................................................................... 21
4.1.2 YFV replication and soluble factors ...................................................... 22
4.1.3 Germinal center assessment ................................................................... 23
4.1.4 Plasmablast expansion ........................................................................... 24
4.1.5 Serological responses ............................................................................. 26
4.1.6 Safety assessment................................................................................... 27
4.2 B cell immune responses to SARS-CoV-2 infection ........................................ 28
4.2.1 B cell and T cell activation during acute COVID-19 ............................ 28
4.2.2 Germinal center assessment during acute COVID-19 .......................... 29
4.2.3 Plasmablast expansion during acute COVID-19 ................................... 30
4.2.4 Serological outcomes during and following SARS-CoV-2
infection.................................................................................................. 32
4.2.5 Persistence of immunological memory ................................................. 33
5 CONCLUSIONS ......................................................................................................... 37
6 FUTURE PERSPECTIVEs ......................................................................................... 39
7 ACKNOWLEDGEMENTS ........................................................................................ 41
8 REFERENCES ............................................................................................................ 45LIST OF ABBREVIATIONS ASC antibody secreting cell ACE2 angiotensin-converting enzyme 2 COVID-19 coronavirus disease 2019 CXCL13 C-X-C motif chemokine ligand 13 CXCR C-X-C chemokine receptor cTfh circulating T follicular helper cell ELISA enzyme-linked immunoassay E protein envelope protein FACS fluorescence activated cell sorting FDC follicular dendritic cell IFN-ɣ interferon ɣ IL- interleukin- JEV Japanese encephalitis virus mASC memory B cell derived antibody secreting cell MHC-II major histocompatibility complex class II nAbs neutralizing antibodies N-protein nucleocapsid protein PBMC peripheral blood mononuclear cell PCR polymerase chain reaction RBD receptor binding domain RFFIT rapid fluorescent foci inhibition test SARS-CoV-2 severe acute respiratory syndrome coronavirus 2 S1-protein spike subunit 1 SHM somatic hyper mutation TBE Tick-borne encephalitis TBEV Tick-borne encephalitis virus Tfh T follicular helper cell TNF Tumor necrosis factor YFV yellow fever virus YFV 17D yellow fever virus vaccine 17D
1 INTRODUCTION
1.1 ADAPTIVE IMMUNITY
As a result of the COVID-19 pandemic, vaccines, the immune system, antibodies, B cells, and
T cells have become common topics of discussion in our everyday lives. The mechanisms
behind how we develop immune memory following vaccination or infection are complex and
still being delineated today. The human immune system can be broadly divided into two parts,
the innate and the adaptive immune systems. The innate immune system acts as the first
responder following infection or vaccination. Innate immune cells start inflammatory
responses, take up antigen, and bring them to the draining lymph nodes thereby acting as a
bridge to adaptive immune responses. The adaptive immune system consists of antigen-specific
responses from B cells and T cells that arise in the days or weeks following antigen exposure1.
A hallmark feature of the adaptive immune system is the development of immunological
memory so that future exposure to the pathogen leads to a more robust and rapid response,
contributing towards preventing severe disease.
1.1.1 B cells
Following infection or vaccination, antigen-specific B cells will, ideally, become activated and
differentiate into antibody secreting cells (ASCs) producing neutralizing antibodies (nAbs) or
memory B cells. The process starts when antigen makes its way to the draining lymph nodes
where B cells capable of binding the specific antigen via their surface B cell receptor (surface
IgD/IgM) undergo a quick activation (Figure 1). In an extrafollicular reaction, B cells rapidly
differentiate into ASCs such as plasmablasts that produce large amounts of antibodies within
the first weeks following vaccination or infection. Plasmablasts, which are short lived ASCs,
have been shown to play a role in clearing active infections and typically show up in circulation
one to two weeks following infection or vaccination secreting large amounts of pathogen-
specific antibodies2–5. A follicular reaction is often initiated when activated cognate CD4+
helper T cells provide costimulatory signals to B cells to enter or initiate the germinal center
reaction6,7. B cells upregulate CXCR5 to migrate towards higher concentrations of CXCL13
produced by follicular dendritic cells (FDC) and T follicular helper (Tfh) cells in the B cell
follicle8,9. During the germinal center reaction, activated B cells undergo rapid proliferation
and somatic hypermutations (SHM) in the dark zone to generate higher affinity B cell receptors.
With new mutations in the BCR, B cells exit to the light zone to test the BCR against antigen
carried by FDCs. Tfh cells provide signaling to B cells with advantages mutations to return to
the dark zone for more SHM or further differentiation. High affinity B cells are ultimately
chosen for further differentiation into memory B cells or ASCs like plasmablasts or long-lived
plasma cells6. Plasma cells migrate to the bone marrow where they produce antibodies and
memory B cells patrol secondary lymphoid organs for their specific antigen10. Memory B cells
that meet their antigen quickly differentiate into ASCs or even better memory B cells with
3higher affinity BCRs3,11. Together long-lived plasma cells and memory B cells are key players
in providing protective immunity from pathogens.
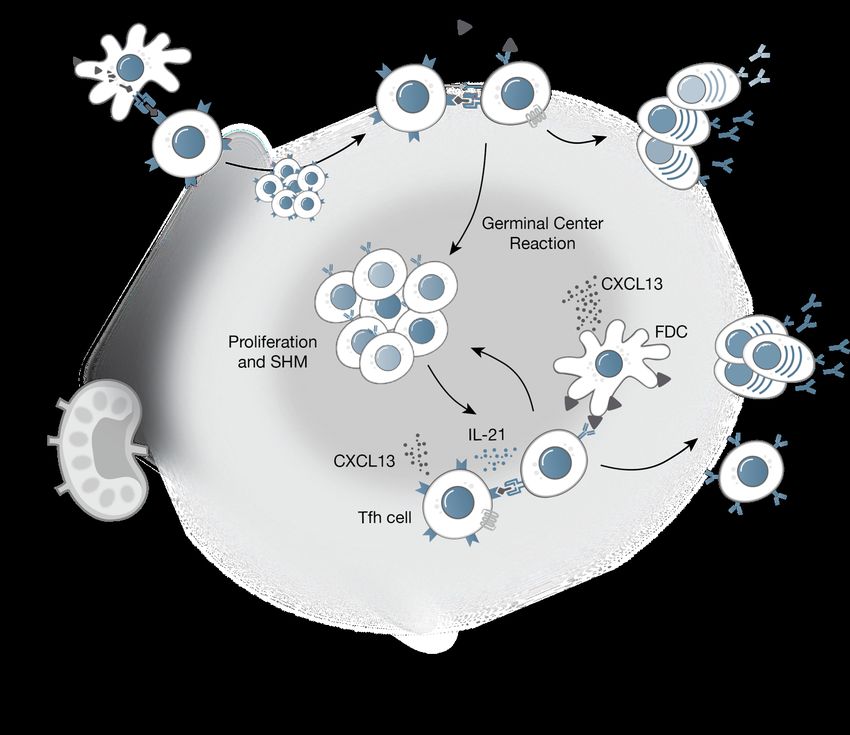
Figure 1 | Germinal center reaction. In response to antigen reaching secondary lymph organs, antigen-specific B cells become
activated. Dendritic cells (DCs) process and present antigen on MHC-II molecules to activate the cells that in turn signal cognate
B cells to enter germinal center (GC) reactions or differentiae into plasmablasts. B cells entering the dark zone of the GC undergo
mass proliferation and SHM honing the B cell receptor. B cells migrate towards the light zone to test the new B cell receptor
and present to cognate germinal center Tfh cells. Advantageous mutations are selected and signals from Tfh cells direct the B
cell to differentiate into memory B cells, plasmablasts, or long-lived plasma cells.
1.1.2 CD4+ T cells
The two main subsets of T cells carry out very different functions during and after vaccination
or infection. CD4+ T cells are commonly referred to as helper T cells and play a critical role in
coordinating adaptive immune responses. Their main function is to help other immune cells
carry out their effector functions through cytokine signaling12,13. Their distinct helper functions
are used in their classification as well including different T helper cells (Th1, Th2, Th17, and
Th22 cells), T regulator cells and Tfh cells14. As their name suggests, Tfh cells reside in follicles
of secondary lymphoid organs. Here they play an essential role in establishing germinal center
reactions. Vaccine- or infection-activated Tfh cells express the chemokine receptor CXCR5
and downregulate the lymph node homing receptor CCR7, directing them into B cell follicles
with higher concentrations of CXCL13 produced by FDCs15,16. There, Tfh cells also secrete
high levels of CXCL13 aiding in the organization of germinal centers17. Activated B cells that
have undergone SMH in the germinal center migrate to these CXCL13 rich areas. Here, Tfh
cells provide co-stimulation to activated cognate B cells via CD40-CD40-L interaction and the
production of IL-21. These signals drive B cell proliferation and is critical both in the selection
of high-affinity B cells and support memory B cell development6,18,19. Increases in peripheral
4CXCL13 levels have been shown to coincide with lymph node germinal center activity16. This
has suggested that serum or plasma CXCL13 levels can act as a biomarker indicating the
activation of germinal centers and thereby, adaptive immune system activation following
vaccination or infection16. Deeper insights into human germinal center reactions and Tfh cell
responses has been hampered due to the invasiveness of extracting human lymph nodes. More
recently, however, a subset of CD4+ T cells in the periphery have been identified that share
phenotypic and functional characteristics of germinal center Tfh cells that are believed to
correspond with their germinal center counterparts20. These circulating Tfh (cTfh) cells have
been shown to correlate with later neutralizing antibody titers and play an important role in
developing immune memory21,22. The expression of CXCR3 on cTfh cells has also revealed
that they can polarize towards Th1 functional cells following vaccination with HPV and
influenza vaccines making them a cell of interest when delineating immune responses
following infection or vaccination23,24.
1.2 FLAVIVIRUSES & FLAVIVIRUS VACCINES
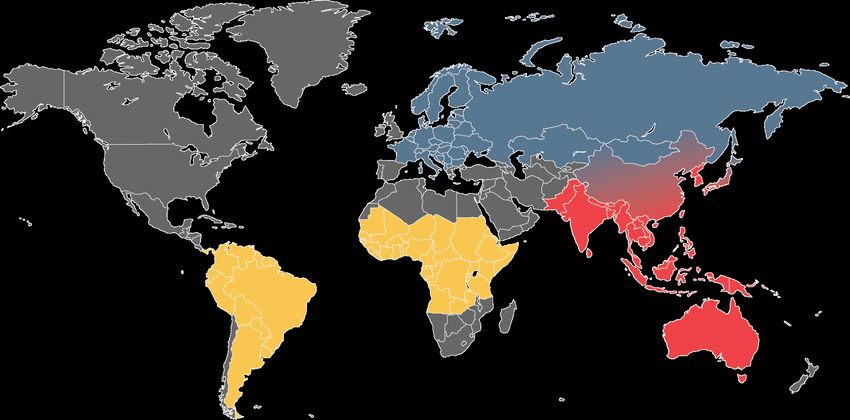
Arboviral infections continue to pose a global health threat affecting nearly every continent.
The virus family Flaviviridae is a large contributor to this threat with human pathogens such
as yellow fever virus (YFV), Tick-borne encephalitis virus (TBEV), Zika virus, Japanese
encephalitis virus (JEV), Dengue virus, and West Nile virus. The reach of flaviviruses are also
increasing with recent outbreaks of Zika and YFV in South America, TBEV across Europe and
northern Asia and JEV spreading in SE Asia and even reaching Australia25–28 (Figure 2). No
specific antiviral treatment exists against these viruses, so vaccination is the main strategy used
to control the spread and preventing severe infection. Fortunately, we do have vaccines against
several of these viruses, three of which are used in the current thesis; YFV, TBEV, and JEV
vaccines. The YFV vaccine uses a live attenuated strain of YFV that, after a single dose,
provides lifelong immunity. TBEV and JEV vaccines, on the other hand are whole, inactivated
Figure 2 | Global distribution of YFV, TBEV and JEV. Map adapted from the Center for Disease Control and the World
Health Organization epidemiological maps. Yellow denotes YFV endemic areas, blue denotes TBEV endemic areas, and red
denotes JEV endemic areas.
5virus vaccines using Alum as adjuvant and require multiple booster doses to provide and maintain immunity. The differences in immune responses and effectiveness from vaccination have inspired scientists to further unravel the complex mechanisms contributing to the development of immunity against these flavivirus infections. The following sections aim to provide an overview of the current understanding of humoral and B cell responses induced by flavivirus vaccines included in the thesis. 1.2.1 YFV YFV infection causes over 200,000 infections and 30,000 deaths annually. The YFV is transmitted to human through bites from infected Aedes aegypti mosquitos leading to varying symptoms depending on the severity of the infection29,30. Following a 3-6-day incubation period, flu-like symptoms arise including fever, headache, and joint pain typically lasting between three and four days before subsiding. In roughly 15% of cases, symptoms can worsen and include nausea, vomiting, renal failure, jaundice, and hemorrhaging leading to a 50% mortality rate within 10-14 days.31 1.2.2 TBEV An estimated 13,000 TBEV infections occur annually transmitted by bites from infected Ixodes ticks and even through ingestion of unpasteurized milk from infected livestock32,33. In two thirds of infection, symptoms arise after a 4 to 28-day incubation period. TBEV infection causes Tick-borne encephalitis (TBE), a biphasic disease with the first viremic stage causing flu-like symptoms including fever, fatigue, malaise, headache, and joint/body pain32,34. Most patients recover at this point and usually do not seek medical care, but in roughly a quarter of cases, following a 7-day symptom-free period, the second phase of disease starts with symptoms ranging from mild meningitis to severe encephalitis that may or may not include myelitis and spinal paralysis35. The majority of patients recover as the fatality rate is rather low (0-15%), but the consequences of infection can leave severe neurological problems and moderate to severe sequalae36,37. 1.2.3 JEV Like YFV, JEV is transmitted to humans via bites from infected mosquitoes (Culex genus) causing roughly 68,000 cases annually over an area with a population over 3 billion38. Most infections with JEV do not lead to symptomatic disease, but about 1 out of 250 infections lead to severe disease. Following a 4 to 14-day incubation period, flu-like symptoms arise including fever, chills, and headache with the addition of myalgia, confusion and even paralysis39. Patients can deteriorate quickly with more severe symptoms including encephalitis and mental disturbances that can lead to fatal outcomes (20-30% fatality rate). Chances of fatal outcomes also increase in young children40,41. 1.2.4 Flavivirus vaccines Vaccination is possibly the most successful medical intervention in modern history42. Their use has saved countless lives and revolutionized the control, and even contributed to the 6
elimination, of certain infectious diseases. There are many types of vaccines all with the same
goal of activating adaptive immune responses that, in turn, develop immune memory consisting
of memory cells and antibody responses42. Several different vaccines are available against
flavivirus infections and in this thesis, those currently available in Sweden are reviewed and
used in the included works.
1.2.4.1 YFV Vaccine
The yellow fever vaccine 17D (YFV 17D) has long been considered by many as one of the
best vaccines presently available due to a very high safety profile and effectiveness43. A single
dose has been shown to provide life-long immunity from YFV infection. The YFV 17D is a
live attenuated virus vaccine that started with isolated Asibi strain that was passaged through
mouse and chick embryo cultures more than 200 times44,45. The brain and spinal cords were
removed from embryo cultures to reduce neurovirulence and viscerotropism of the virus during
the attenuation process. Early clinical trials considered the vaccine safe and showed the
effective development of nAbs following vaccination. This early success lead to large scale
production for larger clinical trials in Brazil and between 1938 and 1941, over 2 million
individuals were vaccinated with YFV 17D becoming one of the first large scale vaccination
campaigns46. YFV 17D is currently produced from two selected substrains from the original
passaged vaccine strain in embryonated eggs under strict standardized procedures outlined by
the World Health Organization45. The YFV 17D is administered in the upper arm
subcutaneously at a volume of 0.5 mL containing 3.7-4.7 log10 IU per dose47. Due to vaccine
shortages during recent outbreaks, multiple studies have reported on the success of fractional
dosing with current YFV vaccines with similar seroconversion rates and efficacy48–52.
As the YFV 17D is replication competent, following administration, low levels of virus titers
are detectable in peripheral blood45,53. This generates a robust innate immune response peaking
on day 7 providing early expression of antiviral molecules that stimulates the adaptive immune
response54. Increases in dendritic and monocyte cells populations have been observed in
peripheral blood along with activated NK cell populations and increased IFN-ɣ levels during
the first week following YFV 17D vaccination likely aiding in the control of the mild infection
induced by the vaccine53,55. The activation of multiple TLRs and upregulation of MHC and
costimulatory molecules on dendritic cells give way to a mixed Th1/Th2 response bridging the
innate and adaptive responses13,56,57. The effective and robust innate response observed after
YFV 17D vaccination has been suggested to be the underlying factor to the strong and durable
adaptive response associated with the successful vaccine45.
The adaptive response to the YFV 17D has been extensively studied with more focus towards
antibody, innate and T cells responses, but only recently have specific B cell responses been
more heavily investigated. The earliest studies carried out while developing the vaccine
focused on antibody responses to test the effectiveness of YFV 17D. It was found that the
majority of human subjects seroconverted by day 14 following vaccination44,58, a finding
confirmed by later studies that also showed 99-100% of subjects seroconverted by day 2859.
The duration of protection, that is to say, the duration of detectable nAbs has been determined
7to be lifelong following a single dose of YFV 17D45,60,61. More recent studies have investigated
cellular responses that lead to the development of nAbs and memory cells. The YFV 17D
establishes a large pool of both memory B cells and memory T cells that, upon subsequent
exposure to YFV, become activated and carry out their specific effector functions. One week
following vaccination, naïve B cells, CD27- IgD+ cells, and memory B cells (CD27+ IgD-)
decrease in frequency in peripheral blood, but return to normal levels by the 28 day mark62.
The plasmablast population has been observed to increase at day 14 following vaccination that
coincides with an increase in nAbs before returning to baseline levels shortly after62,63.
Investigation of these plasmablasts revealed high amounts of somatic hypermutations in their
BCRs suggesting these cells are products of a germinal center reaction63. YFV E-protein
specific memory B cells arise already at day 14 following vaccination and are detectable, albeit
at lower levels, one year following vaccination63. It has also been suggested that humoral
responses elicited by the YFV 17D may be potentially cross reactive with other flaviviruses,
namely in the E protein when considering nAbs64.
1.2.4.2 TBEV Vaccines
Two vaccines against TBEV are currently available in Sweden, FSME-IMMUN (Neudörfl
strain, Pfizer) and Encepur (K23 strain, Bavarian Nordic) and both are recommended for
populations living in endemic areas65. Both vaccines are formaldehyde inactivated whole virus
vaccines combined with aluminum hydroxide as adjuvant. Production of the two vaccines is
essentially the same. The working virus stock was first isolated from Ixodes ticks and then
passaged several times through mouse brains and finally chick embryo cells66. This virus stock
is used to seed new production of the vaccine. The vaccines are administered intramuscularly
at a volume of 0.5 mL and include three primary immunizations at day 0, 1-3 months following
the first dose, and at 5-12 months following the second dose (Figure 3)67. Booster doses are
required to maintain immunity first at 3 years following dose three and then every 5 years
thereafter. It is recommended that the primary vaccination schedule consists of the same
vaccine, but then either FSME-IMMUN and Encepur can be given as booster doses, although
a recent study has shown that either vaccine can be used after two doses of the same68. Most
individuals receiving either of the vaccines seroconvert by the end of primary vaccination69.
Figure 3 | TBEV primary vaccination schedule. Primary vaccination consists of three vaccine doses followed
by a booster dose at 3 years and every 5 years thereafter.
Early effectiveness studies have deemed the vaccines 99% effective in protecting against
TBEV infection where 88% of the population had been vaccinated and 58% had received
regular vaccine boosters70. A more recent review, however, found seroconversion to be around
87%, still relatively high, albeit lower than original studies suggested71. Despite full
8vaccination, breakthrough infections can occur, mostly seen in those over 50 years of age,
although much more uncommon after a fifth dose72–74.
Both TBEV vaccines elicit humoral immune responses primarily targeting the E protein75.
Numerous studies have characterized antibody responses following vaccination76–78, but few
have characterized B cell or T cell responses. TBEV-specific CD4+ T cells respond in lower
frequencies with less polyfunctionality compared to their TBEV infection induced
counterparts79,80. More work is needed characterizing immunological events that lead to the
production of nAbs in order to fully understand current TBEV vaccines and even aid in
developing new vaccination strategies for more successful immunization.
1.2.4.3 JEV Vaccines
A single vaccine against JEV is available in Sweden and recommended for long-term travelers
to endemic regions. The JEV vaccine, Ixiaro (Valneva), is an inactivated whole virus vaccine
based on mouse brain passaged SA-14-14-2 wild type strain cultured in Vero cells. The strain
was chosen due to its high degree of neuroattenuation resulting in a higher safety profile81,82.
The vaccine formulation consists of formaldehyde inactive whole virus combined with
aluminum hydroxide as adjuvant. Each 0.5 mL dose contains roughly 6 µg of viral antigen and
is administered intramuscularly. Primary vaccination consists of two doses, the second dose
given 14-28 days following the first dose (Figure 4). Yearly booster doses are recommended
if staying in endemic areas83.
Figure 4 | JEV primary vaccination schedule. Primary vaccination consists of two vaccine doses followed by a
booster dose each year if remaining in endemic areas.
Similar to TBEV vaccines, seroconversion is quite high following primary vaccination84. NAbs
elicited by the vaccine have been shown to be protective by transferring human sera to mice
that were protected from lethal challenge85. The characterization of immune responses
following JEV vaccination have been limited to mostly serological studies including
persistence studies86. Age-related differences in immune responses have also been reported
with significantly diminished responses towards the vaccine in older popultions87. Reduced
IFN-ɣ and specific antibody titers among elderly individuals provide evidence for the need of
newer and better vaccination strategies against the virus. Other findings have shown the
antibody titers persists at least 5-6 years in vacinees86,88. In one of these studies, using JEV-
specific neutralization assays, it was confirmed that close to 86% of vaccinees still had
protective titers by month 60 who had also previously been vaccinated against TBEV
suggesting potential boosting effects from previous vaccination with another flavivirus.
These cross reactive flavivirus-specific immune responses could lead to new vaccination
strategies for travelers and those living in endemic areas. More work is needed, however, to
9confirm this hypothesis as well as too further characterize B cell and T cell responses to the inactivated JEV vaccine. 1.3 SARS-COV-2 Coronavirus disease (COVID-19), which is caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), first emerged in December 2019 in Wuhan, China and has since led to a global pandemic89. Two years after its first emergence, almost 300 million confirmed SARS-CoV-2 infections have led to more than 5.4 million deaths90. A massive amount of resources and time has gone into delineating how the virus works, immune responses to the infection as well as developing new vaccines in order to control the spread of the virus. 1.3.1 Clinical manifestation A main focus of the research surrounding COVID-19 has been to try to understand the varying levels of disease severity presented after infection with SARS-CoV-2. Many infections lead to asymptomatic disease, while some individuals present mild, moderate, severe, or critical disease that can lead to a fatal outcome91. Infection primarily occurs via infected droplets in the respiratory tract where virus gains entry into epithelial cells by binding to angiotensin- converting enzyme 2 (ACE2)92,93. Viral entry occurs via the virus’ spike protein’s receptor binding domain (RBD) that binds to ACE2 receptors expressed on epithelial cells of the respiratory tract, but also in other organs94. Active replication and release of virus in the respiratory tract often leads to common flu-like symptoms including fever, myalgia, headache and respiratory symptoms95. As previously stated, COVID-19 can present as asymptomatic disease, mild, moderate, severe, or critical disease with several risk factors contributing to more severe disease. These risk factors and co-morbidities include older age, hypertension, obesity, diabetes, cardiovascular disease, and/or chronic obstructive pulmonary disease92. Mild COVID-19 symptoms include flu-like symptoms such as cough, fever, fatigue, and sore throat. Moderate COVID-19 includes symptoms of mild disease with the addition of pneumonia96. Severe COVID-19 develops when oxygen supplementation is required following pneumonia and critical COVID-19 patients often require mechanical ventilation and can present with sepsis and multiple organ failure97. 10
2 RESEARCH AIMS
The original aim of this thesis was to characterize human humoral immune responses induced
by vaccination against different flaviviruses. When the COVID-19 pandemic struck, focus was
turned towards understanding immune responses towards SARS-CoV-2 infection. Many of the
methods used in the initial vaccine studies and knowledge gained there were used and
optimized for characterizing B cell responses in hospitalized COVID-19 patients.
The specific aims of this thesis were to:
• To characterize human B cell responses following YFV vaccination (Paper I).
• To assess the saftey and immune responses of concomittant vaccination with the YFV
vaccine and other inactivated flavivirus vaccines (Paper II).
• To assess and characterize SARS-CoV-2 specific humoral responses in hospitalized
COVID-19 patients (Paper III and IV).
• To assess the persistence and robustness of immunological memory induced by
moderate and severe SARS-CoV-2 infection (Paper IV).
113 MATERIALS AND METHODS
3.1 ETHICAL CONSIDERATIONS
The studies included in this thesis were all approved by local and national ethical review
authorities. Ethical evaluation was done and approved prior to the start of the studies.
Regarding ethical considerations for Paper I and Paper II, the volunteers involved in the
studies were healthy adult volunteers with no previous vaccination or known exposure to the
flaviviruses in question. As TBEV is endemic in many populous areas of Sweden, the TBEV
vaccine is highly recommended and routinely taken by a large portion of the Swedish
population. Similarly, the YFV and JEV vaccines are routinely administered to individuals
traveling to endemic areas and is even required for entry by some countries. Since these
licensed vaccines have previously gone through clinical trials, being deemed safe, and already
available to the public, no major ethical issues apart from what was brought up in applications
to the regional/national ethical committees were identified. Very few, if any, vaccinees follow
up their immunity status following vaccination. By enrolling in the vaccine studies, individuals
were informed about their possibility to get information of their antibody status. All study
volunteers gave informed consent after going through the consent with research and medical
personnel. The informed consent included risks and known side effects from vaccination,
description of the blood sampling, compensation, their rights as volunteers, requirements to
take part in the study and the information of the researchers involved. All volunteer information
and research data are saved and encrypted in approved drives and servers. Each volunteer’s
blood samples were given a unique, coded, ID that only the Karolinska Trial Alliance medical
staff knew to pseudonymize volunteer personal information. Study volunteers also had the right
to leave the study at any point for any reason.
In Papers III and IV, hospitalized COVID-19 patients gave written or oral informed consent
either before or following sampling due to the acuteness of the COVID-19 pandemic and the
urgency to delineate as much as we could about the infection and related immune responses.
Patients who were sampled prior to giving informed consent were able to remove themselves
from the study afterwards, and all collected clinical and research data was deleted entirely.
Patient samples were coded to pseudonymize patient information, and sensitive data was also
handled according to GDPR on encrypted drives and servers. Participation in the study did not
affect patient treatment and sample collection was often done in connection with routine blood
sampling for clinical use. Although there was no direct benefit for participation in the studies
for the patient, it would contribute to improve the understanding of COVID-19 and disease
severity that, in turn, could benefit future patients.
To conclude, ethical consideration was taken for the included studies and the studies were
carried out according to Good Clinical Practice.
133.2 SAMPLE COLLECTION AND PROCESSING Venous blood samples from study volunteers and patients were collected in serum and either EDTA or heparin tubes. PBMCs were isolated using gradient centrifugation and used immediately for fresh experiments or cryopreserved in FCS with 10% DMSO at -180°C for future analyses. Serum tubes were allowed to stand upright for 2 hours, and serum was isolated by centrifugation and stored at -80°C for future analysis. An overview of sample collection timepoints are available in the respective papers. 3.3 FLOW CYTOMETRY Throughout all four studies included in the thesis, flow cytometry was a key method. Fluorescence-activated cell sorting (FACS) is a common immunological method where surface and intracellular markers of cells are stained with fluorochrome-conjugated antibodies and then analyzed on a flow cytometer. The fluorophores are excited by lasers and their emission wavelengths are recorded by the instrument for assessment of molecule expression. Flow cytometry panels were designed to assess B cells, T cells and NK cells and their subsets as well as their expression of certain chemokine receptors, activation markers, and immunoglobulin expression. This method provides insights into cellular populations that can provide glimpses into human immunological responses following vaccination and during infection. In each of the flow cytometry panels used in the studies, both extracellular and intracellular stainings were used. First, either freshly isolated PBMCs or thawed cryopreserved PBMCs were incubated with a cocktail of fluorophore conjugated antibodies specific for different surface markers. Following incubation, the cocktail was removed, and the cells washed before fixation that permeabilizes the cells allowing for intracellular staining. After fixation, a cocktail of fluorophore conjugated antibodies specific for intracellular markers are added. Following this final incubation, cells are washed and resuspended for acquiring on a flow cytometer. FACS is an invaluable method for immunologists generating immense amounts of data from a small sample of cells. 3.4 SEROLOGICAL ANALYSES Following vaccination or infection, pathogen-specific antibodies are usually generated and aid in providing protective immunity from the pathogen in the future. The main methods used to assess the magnitude of these responses are enzyme-linked immunosorbent assays (ELISA) and live virus neutralization assays. ELISAs measure the presence and/or quantity of pathogen- specific antibodies in a given sample, while neutralization assays not only measure antibody titers, but also the antibody’s ability to neutralize infection and prevent infection. Neutralization assays are often referred to as the golden standard to assess specific antibody levels and their functionality but are not always practical and often require higher level safety facilities as one works with high titers of live infectious virus. ELISAs, although practical and don’t require specialized safety considerations, are not as sensitive and don’t assess functionality of preventing infection. The methods are often used in parallel to answer different questions about the serological response. 14
3.4.1 ELISA
Detection and quantification of antigen-specific immunoglobulins were assessed in the
included studies using indirect and antigen “sandwich” ELISA (Figure 5). Diluted serum
samples were added to RBD-antigen coated microplates and antigen-specific immunoglobulins
were captured on the plate-bound antigen. With the indirect ELISAs (TBEV-, JEV-, SARS-
CoV-2 S1-, and N-specific IgG) (Papers II and IV), HRP-conjugated anti-human IgG
antibody was added and bound to the Fc region of the captured antigen-specific IgG. With the
antigen “sandwich” ELISA (SARS-CoV-2 RBD-specific IgG/IgM), HRP-conjugated antigen
was added and bound by the captured specific IgG. In both ELISAs, a chromogen (TMB) was
added reacting with the HRP and giving off a blue color. The reaction is stopped with the
addition of sulfuric acid giving rise to a shift in color. The absorbance of the yellow color is
measured with a spectrophotometer and the value is proportional to the concentration of
amount of specific immunoglobulin present in the serum.
Figure 5 | Principles of indirect
and sandwich ELISAs. Antigen-
specific immunoglobulin binds to
antigen coated wells and is detected
by HRP enzyme conjugated antigen
or detection antibody. The enzyme
reacts with the TMB substrate gives
rise to a blue color and the reaction is
ended with the addition of sulfuric
acid shifting the color to yellow. This
color is proportional to the amount of
antigen-specific immunoglobulin
present in the sample read by a
spectrophotometer.
3.4.2 Neutralization tests
To test the neutralizing capacity of antibodies after vaccination or infection two different
neutralization assays were used in the included studies. The benefit of neutralization assays
over ELISAs is that they give actual insight into the functionally of the developed antibodies
and probably protection from infection. In Papers I & II, a rapid fluorescent focus inhibition
test (RFFIT) against YFV, TBEV and JEV were used. Serum from days 0, 30, and the final
timepoint were tested in serial dilutions with either YFV, TBEV, or JEV in microplates in a
BSL-3 laboratory. BHK-21 cells were added and incubated with the diluted serum and virus
mixture overnight. Plates were fixed with acetone followed by staining with virus-specific
detection antibodies and a fluorophore conjugated antibody. Virus infected cells were detected
using fluorescence microscopy. Titers were determined when wells with 50% reduction in
virus infected cells occurred. In Papers III & IV, the neutralizing capacity of SARS-CoV-2-
specific antibodies were assessed using a microneutralization assay. Patient serum was diluted
in serial dilutions and mixed with equal volumes of SARS-CoV-2. Following incubation, the
serum and virus dilutions were moved to microplates seeded with Vero E6 cells and incubated
15for four days. Plates were then examined with optical microscopy for cytopathic effect (CPE).
Titers were determined neutralizing if less than 50% of the cell layer showed signs of CPE.
3.5 FLUOROSPOT ASSAYS
The ELISpot assay was first developed in 1983 by Cecil Czerkinsky to detect antigen-specific
antibody secreting cells98. The method has since been adapted to assess not only antibody
secreting cells, but also specific T cell responses and with the inclusion of fluorophore
detection, multiple antigens or secreted analytes can be assessed in a single sample. The highly
sensitive ELISpot and FluoroSpot methods are still today considered a gold standard for
detection and enumeration of antigen specific B cells and T cells.
3.5.1 B cell FluoroSpot
In assessing adaptive immune responses to vaccination and infection, it is important to
determine if the observed responses are specific to the pathogen in question. The golden
standard of detection antigen-specific B cells has long been the ELISpot assay. In recent years,
the assay has been enhanced utilizing fluorophores instead of enzymatic detection methods and
deemed FluoroSpot. To detect the presence of vaccine- (Paper I) or infection-specific (Paper
III & IV) ASCs and memory B cells, modified B cell FluoroSpot assays were used. The B cell
FluoroSpot assay enumerates ASCs in a sample using a sandwich assay of capture and
detection antibodies or antigen (Figure 6). The assay allows for the detection of total ASCs in
a sample or only antigen specific cells. ASCs can be assessed directly from PBMC samples
following in vivo activation during infection or following vaccination. To assess memory B
cells, they first require ex vivo stimulation to differentiate into ASCs. A stimulation cocktail of
R848 (a TLR 7/8 antagonist) and IL-2 (aids in B cell differentiation) are added to PBMCs and
incubated for three days. This stimulates memory B cells in the sample to differentiate into
ASCs that are then detectable using FluoroSpot technology. To enumerate total ASCs in a
sample, a PVDF microplate is coated with capture antibody specific for either IgG, IgA, or
IgM. PBMCs are added to wells and incubated overnight. All immunoglobulin secreted by
ASCs fall straight down and are bound by capture antibodies. Cells are then removed, and a
fluorophore-conjugated detection antibody is added creating a sandwich around the target
Figure 6 | Principles of B cell FluoroSpot. PVDF membrane microplates are coated with capture antigen or antibody. In vivo
or ex vivo activated ASCs are added to the wells and secreted antibody is captured by the plate. Fluorophore conjugated antigen
or detection antibody is added and used to detect the number of ASCs present in each well as read by a FluoroSpot reader.
16antibody. Plates are read in a FluoroSpot reader where a spot for each cell present is detected
and counted via fluorescence excitement. Antigen specific cells were detected in two ways.
The first method replaces the capture antibody coating with the antigen of interest coated onto
the PVDF microplates. Once PBMCs are added, only antigen specific immunoglobulin binds
to the plate. The second method, rereferred to as reverse FluoroSpot, replaces the flutophone-
conjugated detection antibody with a fluorophore-conjugated antigen. This way, only antigen-
specific immunoglobulin is read in the reader. In Paper I, YFV E protein-specific ASCs and
memory B cells were assessed using reverse FluoroSpot following YFV vaccination. YFV E
protein was first conjugated with fluorophore and used for detection of specific-ASCs. In
Paper III, SARS-CoV-2 N-protein-specific ASCs were detected in patients with acute
COVID-19 by coating wells with N-protein. IgG, IgA, and IgM N-specific ASCs were directly
assessed from isolated PBMCs. In Paper IV, development of SARS-CoV-2 S1- or N-specific
memory B cells were assessed by coating wells with S1- or N-proteins. Memory B cells
required three-day stimulation for detection and only IgG and IgA cells were assessed.
3.5.2 T cell FluoroSpot
Similar to B cell FluoroSpots, the T cell FluoroSpot is another sensitive method used to detect
antigen-specific T cell responses to a specific pathogen. In Paper IV, the T cell FluoroSpot
assay was used to detect SARS-CoV-2-specific memory T cells. The assay allows for the
detection and enumeration of cells secreting IFN-ɣ, IL-2, TNF in a given sample (Figure 7).
PBMCs were stimulated with SARS-CoV-2 peptide pools (SNMO, S1-spanning overlapping
pool, or N-spanning overlapping pools) at optimized concentrations. PBMCs with stimulation
mixes were incubated in pre-coated microplates with capture antibodies against IFN-ɣ, IL-2,
TNF and incubated overnight. Cells responding to the peptide pools secrete one, two, or all
three analytes and are captured on the plate. Cells are removed and fluorophore-conjugated
detection antibodies specific for the three analytes are added. Spots are detected using a
FluoroSpot reader enumerating single cells as well as the relative amount of analyte secreted.
Figure 7 | Principles of T cell FluoroSpot. PBMCs are incubated overnight in PVDF membrane
microplates coated with capture analyte-specific antibodies. Analyte is secreted from cells and bound
to plates by capture antibodies during incubation. Cells are then removed, and bound analyte is
detected using fluorophore-conjugated detection antibodies that are read using a FluoroSpot reader.
One spot is the equivalent of one cell.
173.6 ANALYSIS OF SOLUBLE MARKERS
3.6.1 ELISA
In Papers I, II, and IV, a sandwich ELISA was used to assess the concentration of serum
CXCL13 (Figure 8). Undiluted serum was added to microplates pre-coated with capture
monoclonal antibody specific for CXCL13. Serum CXCL13 was captured and then detected
using a second CXCL13-specific, enzyme conjugated (HRP) monoclonal antibody. The
chromogen solution (TMB) was then added reacting with the HRP and giving off a blue color
and the reaction was stopped with the addition of sulfuric acid giving rise to a shift in color.
The absorbance of the yellow color is measured with a spectrophotometer and the value is
proportional to the concentration of CXCL13 present in the serum.
Figure 8 | Principle of sandwich ELISA.
Undiluted serum samples are added to
microplates coated with capture antibody specific
to the analyte of interest (CXCL13). Serum is
removed and HRP-conjugated detection
antibodies bind to bound CXCL13 creating a
“sandwich”. TMB chomogen is added and the
enzymatic reaction develops a blue color that is
stopped by addition of sulfuric acid shifting the
color to yellow. This color is proportional to the
amount of analyte present in the sample read by a
spectrophotometer.
3.6.2 Multiplex immunoassay
In Paper II, several soluble analytes were measured in the same sample using a Bioplex
multiplex immunoassay. The principle of the multiplex assay is similar to a sandwich ELISA,
but instead of the capture antibodies being bound to a microplate, they are coupled to
microbeads and multiple different microbeads are used simultaneously in the same sample. The
microbeads are coated with capture antibodies and red and infrared fluorophores used for
identification of the specific bead and ultimately analyte. Several different microbeads are
added to a sample and the analyte of interest binds to the microbead. A biotinylated conjugated
detection antibody is added followed by PE-streptavidin conjugates. The samples are read
using flow cytometry where each microbead emits its unique emission and the coupled PE
intensity is measured from each bead. Together these emissions are translated into a
concentration of the specific analyte present in the sample.
18You can also read

















































