Tumor-Derived Oxidative Stress Triggers Ovarian Follicle Loss in Breast Cancer
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

The American Journal of Pathology, Vol. 193, No. 5, May 2023
ajp.amjpathol.org
TUMORIGENESIS AND NEOPLASTIC PROGRESSION
Tumor-Derived Oxidative Stress Triggers
Ovarian Follicle Loss in Breast Cancer
Yongqi Wu,* Jieqiong Huang,* Hui Chen,* Huan Tao,y Yunbiao He,z Guang Yang,* Qingbing Zha,y Gendie E. Lash,x and
Ping Li*{
From the Departments of Pathology* and Medical Statistics,z Jinan University School of Medicine, Guangzhou; The First Affiliated Hospital of Jinan
University,y Guangzhou; the Guangzhou Women and Children’s Medical Center,x Guangzhou Institute of Pediatrics, Guangzhou Medical University,
Guangzhou; and the Department of Gynecology and Obstetrics,{ The Fifth Affiliated Hospital of Jinan University, Heyuan, China
Accepted for publication
January 6, 2023. Breast cancer is a common indication for ovarian cryopreservation. However, whether the grafting
ovarian tissue meets functional requirements, as well as the need for additional interventions,
Address correspondence to Ping
Li, Ph.D., Department of Pathol- remains unclear. The current study demonstrates abnormal serum hormones in breast cancer in
ogy, Jinan University School of humans and breast cancer cell lineederived tumor-bearing mice, and for the first time shows tumor-
Medicine, No. 601 Huangpu Rd., induced loss of primordial and growing follicles, and the number of follicles being lost to either
Guangzhou 510632, China; or growth or atresia. A gene signature of tumor-bearing mice demonstrates the disturbed regulatory
Gendie E. Lash, Ph.D., Guangz- network of steroidogenesis, which links to mitochondria dysfunction in oocytes and granulosa cells
hou Women and Children’s via the phosphatidylinositol 3-kinase signaling pathway. Notably, increased reactive oxygen species
Medical Center, Guangzhou were identified in serum and ovarian tissues in tumor-bearing mice. Furthermore, supplementation
Institute of Pediatrics, Guangzhou with vitamin C promoted follicular quiescence, repairing tumor-induced follicle loss via inactivation
Medical University, No. 9 Jinsui
of the phosphatidylinositol 3-kinaseeAktemammalian target of rapamycin pathway, indicating the
Rd., Guangzhou 510632, Chi-
potential of antioxidants as a fertility therapy to achieve higher numbers of healthy follicles ready
na. E-mail: pinger355@126.com
or gendie.lash@hotmail.com. for ovarian cryopreservation. (Am J Pathol 2023, 193: 608e623; https://doi.org/10.1016/
j.ajpath.2023.01.015)
Breast cancer is the most common malignant tumor and the embryos and/or oocytes is the current commonly used
leading cause of cancer deaths among women, with an fertility preservation before cancer therapy. However, these
estimated 2.3 million new cases and 685,000 deaths protocols may increase the risk of relapse because of the
worldwide in 2021.1 The overall breast cancer onset in hormonal therapies used.7 An alternative for fertility pres-
China is at a younger age than in many Western countries.2 ervation for women with breast cancer is cryopreservation
Approximately 13.6% of all women diagnosed annually of ovarian tissue without the need for hyperstimulation.8,9
with breast cancers in China were aged
Tumor-Triggered Ovarian Follicle Loss
induction for preserving fertility.11 Because both the quality contraceptives. None of them had a history of disease of the
and the number of follicles are linked to pregnancy out- endocrine organs or hepatitis.
comes, it was questioned whether the grafting ovarian tissue
has normal endocrine ovarian function, as well as the need Hormone Assays
for additional interventions before ovarian transplantation.
It has been well established that cancer cells are under For human, fasting blood samples were collected on the
constant oxidative stress, as reflected by elevated reac- second or third day of the menstrual cycle for all patients,
tive oxygen species (ROS). ROS are a group of highly before operation or any other treatment. Serum levels of
reactive oxygen-containing molecules that are patholog- estradiol, progesterone, follicle-stimulating hormone (FSH),
ically elevated in breast cancer.12 The tumor-derived luteinizing hormone, and prolactin were analyzed using an
ROS can hyperactivate phosphatidylinositol 3-kinase automated chemiluminescent immunoassay analyzer (Beck-
(PI3K)eAkt by inactivating their upstream negative man Coulter UniCel DXI 800; Beckman Coulter, Brea, CA)
regulator phosphatase regulator and tensin homolog at Jinan University, The First Affiliated Hospital laboratory
(PTEN) in tumor cells.13 Furthermore, ROS can regulate (Guangzhou, China). For mice, measurement was performed
the PI3K-Akt signaling pathway in metabolic disorders, using enzyme-linked immunosorbent assay kits and recorded
such as type 2 diabetes and obesity, by directly targeting by microplate reader (Tecan Safire 2, Grödig, Austria).
follicles and thus affecting female fertility.14 Studies
have shown cryopreservation-induced oxidative damage Cell Culture and in Vivo Xenograft Models
in human ovarian tissues of patients with cancer.15
However, it is unclear whether a direct relationship ex- Female BALB/c nude mice (n Z 70; aged 4 to 6 weeks;
ists, and more importantly, whether increased ROS are weight, 18.1 0.1 g) were obtained from Beijing Vital
the cause of ovarian damage in patients with breast River Laboratory Animal Technology Co., Ltd. (Beijing,
cancer. The current study addressed the need for patients China). All mice were maintained under a 12:12-hour light/
with breast cancer to consider therapies to improve fol- dark cycle at a constant temperature of approximately 25 C
licle health before ovarian cryopreservation. The eleva- and humidity between 35% and 75%. MDA-MB-231 cells
tion of follicle-stimulating hormone (FSH) and decrease (BNCC, Beijing, China) were cultured in Dulbecco’s
of estradiol indicated premature ovarian insufficiency modified Eagle’s medium (HyClone, Logan, UT) supple-
associated with breast cancer in humans and mice. The mented with 5% fetal bovine serum (5% CO2, 20% O2,
tumor-bearing ovaries had low follicle density, suggest- 37 C). For experiments, MDA-MB-231 cells were resus-
ing that an underlying cause of these conditions may be pended and subcutaneously injected to the right forelimb of
a tumor-induced alteration in the ovaries. The study also female BALB/c nude mice. Mice were monitored for
indicated occurrence of oxidative stress in patients with development of primary xenograft tumors and sacrificed
breast cancer and in tumor-bearing mice. It showed that when tumors reached 10% of body weight. Experimental
the inactivation of PI3K-Aktemammalian target of protocols were approved by the Laboratory Animal Com-
rapamycin (mTOR) pathway by an antioxidant may be a mittee of Jinan University (number 00290215).
promising strategy for the derivation of healthy follicles
and functional ovarian tissues for patients with breast Estrus Cycle
cancer. Further studies are needed to define the role of
breast tumor-derived defects in the ovary and provide The estrus cycle of mice was monitored continuously by
safer and more efficient therapeutic options. vaginal smear for 20 days. Vaginal cytology was stained
with Crystal Violet Staining Solution (Beyotime, Shanghai,
China). The estrous cycle stage was determined according to
the cell types observed in the vaginal smears.
Materials and Methods
Human Subject’s Serum Collection Histology
Premenopausal women, aged 20 to 40 years (n Z 150; stage The mouse ovaries were fixed, embedded in paraffin,
I to III; TNM system), with newly diagnosed primary breast sectioned (5 mm thick), and stained with hematoxylin and
cancer were enrolled. Premenopausal status was defined as eosin. The follicles in the ovarian histologic sections were
having a regular vaginal bleeding history at the time of classified as primordial, primary, secondary, or antral folli-
diagnosis. Patients had no distant metastasis, any other cles, according to the morphologic structure of surrounding
malignancies, or major medical illness. Controls (n Z 103) somatic cells. Atretic preantral follicles were recognized
were randomly selected from the outpatient clinic with according to the morphologic criteria, as described by
routine hormonal measurements. Women are nonsmokers, Myers et al16 and Kitajima et al.17 Every fifth and sixth
and they did not take any dietary supplements of vitamin C histologic section were selected for comparison and
(VC) or E or use any alcohol, hormones, or oral evaluation.17
The American Journal of Pathology - ajp.amjpathol.org 609
Wu et al
Table 1 Antibodies Used for Immunostaining, Immunofluorescence, and Western Blot Analysis
Antibodies Source Identifier: catalog no.
Rabbit anti-DDX4 Abcam (Cambridge, UK) ab13840
Rabbit antiecleaved capspase-3 Cell Signaling Technology (Danvers, MA) 9961
Rabbit anti-FOXO3a (phosphorylated S253) Abcam ab47285
Rabbit anti-FSHR Proteintech (Wuhan, China) 22665-1-AP
Rabbit anti-Akt (phosphorylated Thr308) Cell Signaling Technology 13038
Rabbit anti-Akt Cell Signaling Technology 4691
Rabbit anti-p-RPS6 (phosphorylated Ser235/236) Cell Signaling Technology 4858
Rabbit anti-RPS6 Cell Signaling Technology 2217
Mouse anti-Mullerian hormone Invitrogen (Waltham, MA) PA5-35851
Mouse anti-PCNA Cell Signaling Technology 2586
Mouse anti-FOXO3a Proteintech 66428-1-lg
Mouse antieb-actin Affinity Biosciences (Cincinnati, OH) T0022
DDX4, DEAD-box helicase 4; FOXO3a, forkhead box O3a; FSHR, follicle-stimulating hormone receptor; PCNA, proliferating cell nuclear antigen; p-RPS6,
phosphorylated ribosomal protein S6.
Immunohistologic Staining and Qualification (https://bioconductor.org/packages/clusterProfiler) of R.
Gene set enrichment analysis software version 4.2.3 [http://
Immunohistochemistry and immunofluorescence staining were www.gsea-msigdb.org/gsea/downloads.jsp (registration
performed as previously described.18 Antibodies are presented required)] was used to identify whether a set of genes
in Table 1. The intensity was quantified and compared between associated with specific GO terms or pathways showed
each group with the follicles only at the same development. significant differences between the two groups. Enrichment
scores and P values were calculated with default parameters.
Real-Time Quantitative PCR and Western Blot Analysis
Transmission Electron Microscopy
Real-time quantitative PCR and Western blot analysis were
performed as previously described.18 The specific primers Ovarian specimens were prefixed with glutaraldehyde (Mack-
and antibodies are presented in Table 2. The expression of lin, Shanghai, China) and post-fixed in osmium acid solution.
genes was normalized to b-actin. The results are represen- After dehydration in acetone and embedding in epoxy resin,
tative of three independent experiments. ultrathin sections were cut and loaded onto copper screens.
These sections were stained with uranium acetate and lead
RNA Sequencing and Analysis
Table 2 Primer Sets Used for Real-Time Quantitative PCR
RNA isolated from ovarian tissues of control and tumor-bearing Gene name Primer sequence
mice (n Z 3 per group; age, postnatal day 80) was constructed ACTB Forward: 50 -AGAGAAGCTGTGCTATGTTGCT-30
into libraries using an MGI Easy mRNA Library Prep Kit (BGI, Reverse: 50 -GGAACCGCTCGTTGCCAATA-30
Shenzhen, China). The sequencing library was used for cluster BMP15 Forward: 50 -TTATACCATCGTTCGGCTGAC-30
generation and sequencing on the MGIseq2000 platform (BGI). Reverse: 50 -GAAAGTCCAGGGTCTGTACATG-30
Differentially expressed genes (DEGs) between different sam- CYP11A1 Forward: 50 -TGGGACTTAAGGCAGAAGCG-30
ples were detected as those with a fold change 2 and a P < 0.05 Reverse: 50 -CCTGCCAGCATCTCGGTAAT-30
using the R package DESeq2 version 1.34.0 (https://github. CYP19A1 Forward: 50 -TGATCATGGGCCTCCTTCTC-30
com/mikelove/DESeq2), and the DEGs were further analyzed Reverse: 50 -CCCAGACAGTAGCCAGGACCT-30
by the Gene Ontology (GO) and Kyoto Encyclopedia of GDF9 Forward: 50 -CAGTCCACCTGGAGGCCTTTA-30
Genes and Genomes databases to assess their functional Reverse: 50 -GAGCGGATGGCTTTCTGCCCT-30
GPX1 Forward: 50 -ATCAGTTCGGACACCAGGAG-30
enrichment. The RNA-sequencing data of control and tumor-
Reverse: 50 -TCACCATTCACTTCGCACTTC-30
bearing mice were submitted to the Gene Expression GSTM2 Forward: 50 -ATGGTTTGCAGGGAACAAGGT-30
Omnibius (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi? Reverse: 50 -CTTCAGGCCCTCAAAGCGAC-30
accZGSE2081 79; accession number GSE208179). HSD17B2 Forward: 50 -AATCATCAGACAGGAGCTTGAC-30
Reverse: 50 -CCTCTCTTTCAAGGTCGGGAT-30
Data Analysis IDH1 Forward: 50 -ATGCAAGGAGATGAAATGACACG-30
Reverse: 50 -GCATCACGATTCTCTATGCCTAA-30
GO and Kyoto Encyclopedia of Genes and Genomes pathway STAR Forward: 50 -AACGGGGACGAAGTGCTAAG-30
enrichment analysis for DEGs was performed using the Reverse: 50 -CCTCTGCAGGACCTTGATCTC-30
org.Hs.eg.db package version 3.15.0 (https://bioconductor.org/ VNN1 Forward: 50 -AACTGGATACCCTGTGATAACCC-30
Reverse: 50 -GTCTCCCATGTTCGCCACAA-30
packages/org.Hs.eg.db) and clusterProfiler version 4.4.4
610 ajp.amjpathol.org - The American Journal of Pathology
Tumor-Triggered Ovarian Follicle Loss
Figure 1 Dysregulated serum hormones in human and mice with breast cancer. A: Serum hormonal levels in premenopausal women with breast cancer,
aged 20 to 40 years (stage I to III), and matched healthy controls were measured by chemiluminescence. B: The experimental scheme of the tumor-bearing
mouse model (left panel). Human invasive breast cancer cells MDA-MB-231 were injected subcutaneously for 4 weeks in female BALB/c nude mice. Plot
indicating average mice weights (right panel). C: Crystal Violet staining of vaginal smears from mice in four different stages of the estrous stage; the relative
proportions of epithelial cells and leukocytes in vaginal smears were used for cycle stage determination. D: Representative estrous cycle of mice during 20
consecutive days and quantitative analysis of estrous cycles. E: Representative ovary gross images (left panel). Statistics of ovary/body weight ratio in control
and tumor-bearing mice (right panel). F: Serum hormonal levels in control and tumor-bearing mice were detected by enzyme-linked immunosorbent assay.
n Z 150 premenopausal women with breast cancer (A); n Z 103 healthy controls (A). *P < 0.05, **P < 0.01, and ***P < 0.001 indicate significant
differences between control and experimental groups. Scale bar Z 50 mm (C). E2, estradiol; AMH, anti-Mullerian hormone; FSH, follicle-stimulating hormone;
LH, luteinizing hormone; PRL, prolactin.
citrate, and then examined by transmission electron microscopy was used for multiple comparisons of data from more than two
at the Analysis and Testing Center of Jinan University. groups. Results are presented as means SEM, and P < 0.05
was considered statistically significant.
Statistical Analysis Results
GraphPad Prism 9 Software (GraphPad Software, San Diego, Alteration in Hormone Levels in Breast Cancer
CA) was used for all analyses. Comparisons between groups
were performed by unpaired t-test, and data with unequal The women in the tumor group had increased levels of FSH
variance were compared with the U-test. Analysis of variance and luteinizing hormone and decreased levels of estradiol
The American Journal of Pathology - ajp.amjpathol.org 611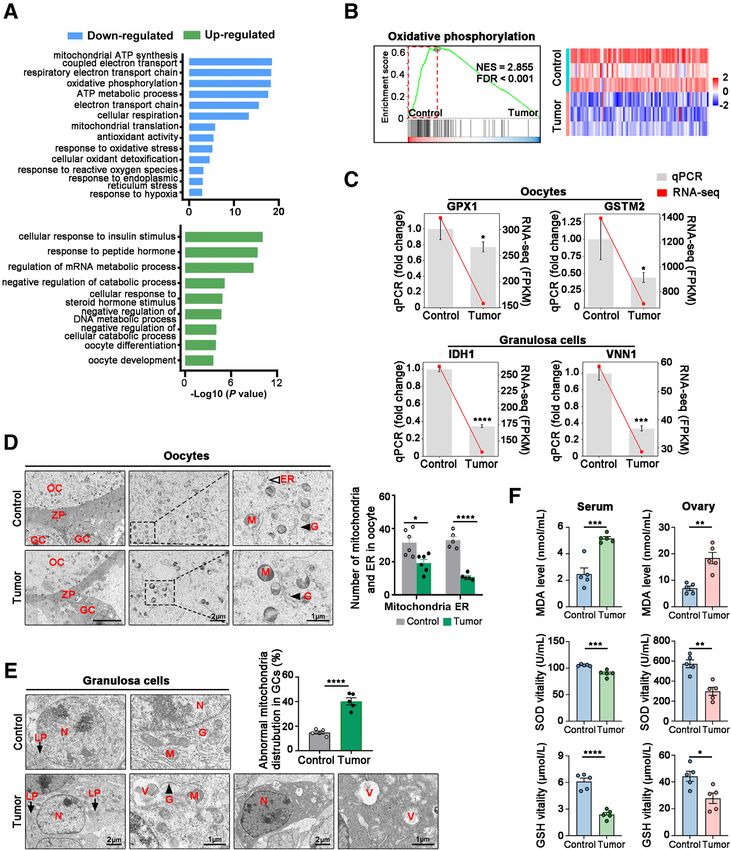
Wu et al and testosterone as compared to the healthy controls. There healthy premenopausal women, and there was no significant was no difference in progesterone and prolactin between the difference in overall hormone levels between hormone two groups (Figure 1A). The hormone levels in the cohorts receptorepositive and hormone receptorenegative tumors with different breast cancer subtypes were similar to those in (Table 3). To examine the impact of the in vivo tumor- 612 ajp.amjpathol.org - The American Journal of Pathology

Tumor-Triggered Ovarian Follicle Loss
Table 3 Serum Hormone Levels in Hormone ReceptorePositive and Hormone ReceptoreNegative Female Subjects with Breast Cancer
Hormone receptorepositive subjects (n Z 114) Hormone receptorenegative
Serum sex hormone levels Luminal A (n Z 37 Luminal B (n Z 43) HER2 (n Z 34) subjects (n Z 36) P value
Estradiol, pg/mL 42.7 (18.1e97.0) 53.3 (17.4e109.5) 0.7237
Progesterone, ng/mL 0.6 (0.3e1.45) 0.7 (0.4e1.5) 0.6478
Testosterone, ng/mL 0.4 (0.3e0.5) 0.4 (0.3e0.5) 0.3416
Prolactin, ng/mL 12.5 (9.3e20.6) 15.2 (7.6e24.2) 0.8531
FSH, mIU/mL 8.9 (6.0e31.8) 8.2 (4.4e19.7) 0.2478
LH, mIU/mL 7.1 (3.9e20.2) 5.1 (3.1e18.6) 0.2858
The values are median (25th percentilee75th percentile) for variables with skewed distribution, and data were analyzed using U-test.
FSH, follicle-stimulating hormone; HER2, human epidermal growth factor receptor 2; LH, luteinizing hormone.
derived microenvironment on the molecular and cellular primordial and secondary follicles (Figure 2, A and B).
properties of the ovary, the study established xenograft tu- Proportionally, there was a greater impact on the primordial
mors of the invasive breast cancer line MDA-MB-231 in follicles than on the early growing follicles, resulting in a
female BALB/c nude mice19 (Figure 1B and Supplemental higher ratio of growing versus primordial follicles
Figure S1). The tumor-bearing and control mice did not (Figure 2C). Positive DEAD-box helicase 4 (DDX4) stain-
have differences in body weight (Figure 1B). The estrous ing was detected in oocytes within follicles at all stages of
cycle is generally divided into the four stages of proestrus, developing ovaries, and immunofluorescence staining
estrus, metestrus, and diestrus (Figure 1C).20 The control showed an obvious decrease in the number of primordial
mice had a regular estrous cycle, and the average estrous follicles in tumor-bearing mice (Figure 2D) compared to
cycle stage lasted 4 to 5 days. The tumor-bearing mice had control mice, suggesting a direct destruction of primordial
prolonged metestrus and diestrus stages, as indicated by follicles.
microscopic analysis of the predominant cell types in the Follicle atresia is the main process responsible for the loss of
vaginal smears (Figure 1D). After 4 weeks, tumor-bearing ovarian follicles, and apoptosis is the underlying mecha-
mice exhibited smaller ovary size and disturbed hormone nism.22 Terminal deoxynucleotidyl transferase-mediated
levels (Figure 1, E and F). In tumor-bearing mice, the serum dUTP nick-end labeling staining analysis showed that signif-
estradiol and prolactin levels were decreased by 52% and icantly more atretic follicles were present in ovaries of tumor-
43%, respectively, whereas the serum FSH, luteinizing bearing mice than in controls (25% versus 7%) (Figure 2H).
hormone, and progesterone levels were elevated by 40%, The percentage of growing atretic follicles to total atretic fol-
22%, and 7%, respectively (Figure 1F), as compared to licles was higher compared with the percentage of primordial
control mice. atretic follicles (total, 27% versus 13%) (Figure 2, E and F).
Atresia was further confirmed by immunostaining for cleaved
Tumor-Induced Follicle Loss in Breast Cancer caspase-3, which showed atretic follicles, including primordial
follicles and growing follicles, in both tumor and control
Despite the known effects of estrogen and progesterone on groups (Figure 2, E and G), suggesting a hyperactivated
breast cancer,21 there have been no reports on ovarian apoptosis signaling pathway in tumor-bearing mice.
physiology in breast cancer. Follicles are the morphofunc- There was intense nuclear proliferating cell nuclear anti-
tional unit of the ovary; the tumor-bearing mice displayed a gen staining in the granulosa cells (GCs) of growing folli-
decrease in the number of total follicles, particularly the cles in both groups. While the proportion of overall positive
Figure 2 Tumor-induced follicle loss may be linked to follicular activation and apoptosis. A: Representative histology images of ovary sections from
control (left panels) and tumor-bearing mice (tumor; right panels). B: Bar chart showing the quantification of total, primordial, primary, secondary, and
antral follicles in the two groups. C: Comparison of the ratio of growing (primary plus secondary)/primordial follicles in ovaries of each group. D: Images of
immunofluorescence, with dashed lines showing the nuclei of the oocyte of primordial follicles were detected using an antieDEAD-box helicase 4 (anti-DDX4)
antibody in each treatment group. All of the nuclei were stained with DAPI. E: Representative hematoxylin and eosin (H&E; top panels) and cleaved caspase-3
staining (bottom panels) of atretic follicles (dashed lines) at different development stages. Top and bottom panels: Insets: Higher magnifications of the
dotted boxed areas. F: Bar chart showing the quantification of atretic follicles within primordial, primary, secondary, and antral follicles from the two groups.
G: Immunofluorescence staining of cleaved caspase-3epositive granulosa cells (GCs; white arrowheads) in growing follicles, and quantification of positive GC
rate per follicle in the two groups. Right panels: The regions in dotted boxed areas (middle panels) are enlarged. H: Immunofluorescence staining of
terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL)epositive GCs (white arrowheads) in growing follicles, and quantification of
positive GC rate per follicle in the two groups. Right panels: The regions in dotted boxed areas (middle panels) are enlarged. I: Immunofluorescence staining
of proliferating cell nuclear antigen (PCNA)epositive GCs (white arrowheads) in follicles at different development stage, and quantification of positive GC rate
per follicle in the two groups. Top and bottom left panels: Insets: Higher magnifications of the dotted boxed areas. *P < 0.05, ****P < 0.0001 indicate
significant differences between control and experimental groups. Scale bars: 200 mm (A, G, and H); 50 mm (D, E, and I); PDF, primordial follicle. SF, secondary
follicle.
The American Journal of Pathology - ajp.amjpathol.org 613
Wu et al follicles was decreased (70% versus 30%), the proportion of proliferating ability. On the other hand, the increase in primordial follicles was increased in tumor-bearing mice primary follicles and increased ratio of growing/primordial compared with that in controls (21% versus 63%) follicles (Figure 2B) suggested that the tumor was causing (Figure 2I), suggesting that the follicles were viable with primordial follicles to undergo recruitment and growth. 614 ajp.amjpathol.org - The American Journal of Pathology
Tumor-Triggered Ovarian Follicle Loss
Activation of PI3K-Akt-mTOR in Tumor-Bearing Mice ribosomal protein S6 (RPS6), and FOXO3a in the tumor-
bearing group (Figure 3H). These findings support the
To investigate the molecular mechanism by which breast idea that the PI3K-Akt-mTOR pathway, which has been
cancer tumors induced ovarian follicle loss, bulk RNA shown to be involved in follicular activation, recruitment,
sequencing was used to examine DEGs in the tumor-bearing and growth,24e27 was activated in the tumor-bearing
mice (Figure 3A and Supplemental Figure S2). Functional group.
enrichment analysis showed the gene cluster enrichment of
categories, such as the AMP-activated protein kinase
signaling pathway, forkhead box O (FOXO) signaling Gene Expression Analysis Suggests Changes in
pathway, mammalian target of rapamycin (mTOR) Hormonal Regulation
signaling pathway, and insulin signaling pathway, which are
associated with growth and development of ovarian follicles Gene set enrichment analysis indicated that functional
(Figure 3B). The gene set enrichment analysis results genes from steroid hormone biosynthesis were down-
showed significant dysregulation of PI3K-Akt and mTOR regulated in the tumor-bearing group compared with con-
pathways (Figure 3C). trols (Figure 4A). In the hypothalamus-pituitary-gonadal
PTEN acts a negative regulator of PI3K-Akt signaling axis, FSH binds to its receptor in GCs to regulate secre-
in primordial follicle activation.23 Immunofluorescence tion of sex steroid hormones. Anti-Mullerian hormone
staining detected PTEN expression in the cytoplasm of (AMH) modulates sensitivity of early growing follicles to
GCs from primordial to growing follicles in ovaries of FSH.28 Immunostaining results showed that both FSH re-
control, whereas the relative intensity of PTEN in GCs ceptor and AMH were expressed in GCs of different fol-
was decreased 33% in the tumor-bearing group compared licle stages in ovaries from both groups (Figure 4B). In
with that in controls (Figure 3D), as determined by primordial follicles, there was no difference in GC FSH
Western blot analysis (Figure 3E). FOXO3 is regulated by receptor expression, whereas in secondary and antral fol-
PTEN. On phosphorylation, FOXO3a is exported to the licles in the tumor-bearing group, there was a significant
cytoplasm of oocytes during primordial follicle activa- reduction in expression compared with that in the control
tion.23 Immunofluorescence analysis demonstrated that group (Figure 4B). AMH expression in GCs was signifi-
FOXO3a was highly expressed in the oocyte nuclei of cantly decreased in secondary follicles in the tumor-
primordial follicles double stained with DDX4 in con- bearing group (Figure 4B). Oocyte-derived factors, bone
trols, and there was increased nuclear exclusion of morphogenetic protein 15, and growth differentiation
FOXO3a in the ovaries of tumor-bearing group (Figure 3, factor-9 have been shown to play key roles in GCs and to
F and G). Consistent with these data, phosphorylated induce AMH,29 and their mRNA expression was increased
FOXO3a was increased and exported from the nuclei to in the tumor-bearing group, as determined by RNA-
the cytoplasm of primordial follicles in the tumor-bearing sequencing analysis and real-time quantitative PCR
group (Figure 3G). Meanwhile, the nuclear localization of (Figure 4C).
phosphorylated FOXO3a was also observed in the GCs of Analysis of microarray data showed that DEGs were
growing follicles (Figure 3, F and G). Western blot involved in estrogen synthesis, steroid signaling, and es-
analysis showed decreased expression of PTEN and trogen metabolism (Figure 4D). The rate of steroid
increased phosphorylation of key activation proteins Akt, biosynthesis is controlled by steroidogenic acute regulatory
Figure 3 Tumor triggers activation of phosphatidylinositol 3-kinase (PI3K)eAktemammalian target of rapamycin (mTOR). A: A heat map of 2482
differentially expressed genes (DEGs) by RNA sequencing in ovaries of control and tumor-bearing mice. B: Enriched Kyoto Encyclopedia of Genes and Genomes
(KEGG) pathways of DEGs in ovaries of control and tumor-bearing mice. The size of the dots represents the number of genes in each category. PI3K-Akt
signaling and mTOR signaling (red text) were up-regulated, whereas oxidative phosphorylation (blue text) was down-regulated, in the enrichment anal-
ysis. C: Immunofluorescence staining of phosphatase regulator and tensin homolog (PTEN) in granulosa cells (GCs) in primordial follicles, and quantification of
the percentages of PTENþ GCs (white arrows) per follicle within ovaries in the two groups. Right panels: Higher magnifications of the dotted boxed areas
(middle panels). D: Gene set enrichment analysis of transcriptome data indicating that the gene sets PI3K-Akt signaling and mTOR signaling are enriched in
ovaries of the tumor-bearing mice. E: Western blot analyses of PTEN in the ovaries of control and tumor-bearing mice (left panel), and protein quantification
normalized to b-actin (right panel). F: Immunofluorescence showing localization of forkhead box O3a (FOXO3a; Alexa488; green) and the oocyte maker DDX4
(Alexa555; red). All of the nuclei were stained with DAPI. FOXO3a was localized to cytoplasm of oocytes of the activated oocytes (white arrows). Top and
bottom right panels: Insets: Higher magnifications of the dotted boxed areas. G: Immunostaining for nuclear shuttling of FOXO3a and phosphorylated
FOXO3a (p-FOXO3a) expression and increased cytosolic localization of p-FOXO3a in activated primordial follicles (red circles). Insets: Higher magnifications of
the dashed boxed areas. H: Representative immunoblot analysis (left panel) and quantification (right panel) of key proteins of PI3K-Akt and mTOR signaling
in ovaries of tumor-bearing and control mice. *P < 0.05, **P < 0.01, and ***P < 0.001 indicate significant differences between control and experimental
groups. Scale bars Z 50 mm (C, F, and G). AMPK, AMP-activated protein kinase; DDX4, DEAD-box helicase 4; FDR, false discovery rate; Max, maximum; Min,
minimum; NES, Normalized Enrichment Score; p-Akt, phosphorylated Akt; PPAR, peroxisome proliferator-activated receptor; p-RPS6, phosphorylated ribosomal
protein S6; TGF-b, transforming growth factor-b.
The American Journal of Pathology - ajp.amjpathol.org 615Wu et al protein,30 and the down-regulation of estrogen and they work together to conduct steroidogenesis. These synthesiserelated genes was validated by real-time findings suggested that the disturbed regulatory network of quantitative PCR (Figure 4E). The oocyte is directly sur- steroidogenesis was responsible for abnormal sex hormone rounded by layers of GCs, followed by thecal cells (TCs), production in the tumor-bearing mice (Figure 4F). 616 ajp.amjpathol.org - The American Journal of Pathology
Tumor-Triggered Ovarian Follicle Loss
Oxidative Stress Regulation in the Ovary with Breast antioxidant VC was used to determine the relationship be-
Cancer tween ovarian oxidative stress and follicle development ca-
pacity in mice (Figure 6A and Supplemental Figure S3). The
ROS exert effects on follicle growth, oocyte maturation, and antioxidant effects were assessed by the serum and ovarian
ovarian steroid biosynthesis.14 In ovaries of mice, enrichment oxidative stresserelated parameters. Compared with tumor-
GO analysis showed that the down-regulated genes were bearing mice, there was an increase in superoxide dismut-
enriched in the terms mitochondrial ATP synthesis, oxidative ase activity and glutathione levels in serum (13% and 69%,
phosphorylation, and response to oxidative stress, and the up- respectively) and in ovarian tissues (39% and 40%, respec-
regulated genes were enriched in hormone and oocyte tively), whereas there was a reduction in malondialdehyde
development (Figure 5A). Gene set enrichment analysis also levels in serum (50%) and ovarian tissue (56%) in the VC-
demonstrated that functional genes from oxidative phos- treated group, indicating that the antioxidant VC could
phorylation were down-regulated in the tumor-bearing group reduce the oxidative stress levels of tumor-bearing mice
(Figure 5B). Consistent with the results of RNA sequencing, (Supplemental Figure S4). The size and weight of ovaries
several of the enzymatic antioxidant enzymes, such as from VC-treated mice were rescued compared with control
oocyte-specific glutathione peroxidase and glutathione S- (Figure 6B). VC treatment significantly reduced total pri-
transferase mu 2,31,32 and GC-specific isocitrate dehydroge- mordial tumor-derived follicle loss (Figure 6, C and D).
nase and vanin-132,33 were significantly down-regulated in Quantification of the number of the different follicle pop-
the tumor-bearing group (Figure 5C). The ultrastructural ulations in the ovaries of mice treated with VC showed that
changes in mitochondria of the ovaries were assessed using the total number of follicles and the numbers of primary and
transmission electron microscopy.34 In the tumor-bearing secondary follicles matched those in ovaries from
group, the number of mitochondria within oocytes was nonetumor-bearing mice (Figure 6C). Immunofluorescence
decreased by 39% compared with that in controls, accom- staining of DDX4 showed an obvious increase in the number
panied with a decrease in endoplasmic reticulum and Golgi of primordial follicles in VC-treated mice (Figure 6D).
bodies (Figure 5D). In contrast, GCs had higher levels of Proliferating cell nuclear antigen staining showed the pro-
abnormal mitochondria compared with controls, which were portion of positive GCs per follicle increased in secondary
vacuolized, and a few mitochondria in which cristae formed follicles (50% versus 29%), whereas they were decreased in
concentric whorls, the amounts of lipid droplets and swollen the primordial follicles (20% versus 69%) in the VC group
Golgi apparatuses were also significantly altered, and GCs compared with that of untreated ovaries (Figure 6E). VC
were condensed with more obvious apoptotic bodies treatment also caused a decrease in the number of atresia, but
(Figure 5E). Mitochondria dysfunction is often associated not in antral follicles (Figure 6F). Terminal deoxynucleotidyl
with increased ROS production by the organelle itself; transferase-mediated dUTP nick-end labeling and cleaved
therefore, changes in serum ROS levels in mice were caspase-3 immunofluorescence staining in ovaries from the
assessed. Superoxide dismutase activity and glutathione VC group contained fewer apoptotic follicles compared with
levels were significantly decreased, whereas the malondial- untreated mice (Figure 6, G and H).
dehyde levels were significantly increased, indicating high PTEN expression in GCs was increased in the VC group,
ROS levels and low antioxidant activity in both serum and as determined by immunofluorescence (Supplemental
ovarian tissues in tumor-bearing mice (Figure 5F). Figure S5) and Western blot analysis (Figure 7A). VC
treatment also inhibited shunting of FOXO3a to the cyto-
Antioxidant Treatment Alleviates Tumor-Derived plasm and attenuated nuclear phosphorylated FOXO3a
Follicle Loss (Figure 7B). Consistent with these data, Western blot analysis
results showed that VC treatment increased the phosphory-
To confirm whether the excessive ROS contributed to follicle lation of activation proteins Akt and RPS6 compared with the
loss and the underlying mechanisms in breast cancer, the untreated group (Figure 7C). These results suggest that VC
Figure 4 Tumor affects the transcriptional signature of oocyte and granulosa cells (GCs) in ovarian tissues. A: Gene set enrichment analysis results for the
gene sets steroid hormone biosynthesis using RNA-sequencing (RNA-seq) data from ovaries of control and tumor-bearing mice. B: Immunostaining for follicle-
stimulating hormone receptor (FSHR) and anti-Mullerian hormone (AMH) expression in GCs in follicles at different development stage, and quantification of
the percentages of positive GCs per follicle within ovaries in the two groups. Left panels: Insets: Higher magnifications of the dashed boxed areas. C: Real-
time quantitative PCR (qPCR) validating and quantifying changes in oocyte-derived transcripts [bone morphogenetic protein 15 (BMP15) and growth dif-
ferentiation factor-9 (GDF9)] selected from RNA-seq data. D: Heat map illustrating differences in ovarian tissues between control and tumor-bearing mice in
the expression of a cohort of transcripts involved in estrogen synthesis, including cytochrome P450 (CYP)11, CYP17, CYP19, and hydroxysteroid (HSD)17b; in
steroid signaling, including estrogen receptor (ESR)1, ESR2, progesterone receptor (PGR), and sex hormone binding globulin (SHBG); and in estrogen
metabolism, including CYP1A1, CYP1b1, and CYPIb1. E: qPCR validating and quantifying changes in steroid biosynthesiserelated transcripts [steroidogenic
acute regulatory protein (StAR), CYP11A, CYP19A, and HSD17b2] selected from RNA-seq data. F: The proposed working model of regulatory network of cell
typeespecific genes involved in steroidogenesis between oocytes, GCs, and thecal cells, which could contribute to abnormal hormonal regulation. *P < 0.05,
**P < 0.01, and ****P < 0.0001 indicate significant differences between control and experimental groups. Scale bars Z 50 mm (B). FDR, false discovery rate;
FPKM, fragments per kilobase million; FSH, follicle-stimulating hormone; LH, luteinizing hormone; LHR, LH receptor; NES, Normalized Enrichment Score; UGT,
uridine diphosphate glucosyltransferase.
The American Journal of Pathology - ajp.amjpathol.org 617Wu et al Figure 5 Mitochondria dysfunction and increased oxidation state in breast cancer and in ovary with breast cancer. A: Gene Ontology terms of down- regulated (blue) and up-regulated genes (green) in ovarian tissues from control versus tumor-bearing mice. B: Enrichment plot oxidative phosphorylation (left panel) for the comparison of control versus tumor group and corresponding heat map (right panel) of the core enrichment genes. C: Real-time quantitative PCR (qPCR) validating and quantifying changes in oocyte-derived transcripts [glutathione peroxidase (GPX1) and glutathione S-transferase mu 2 (GSTM2)] and granulosa cell (GC)ederived transcripts [isocitrate dehydrogenase (IDH1) and vanin-1 (VNN1)] selected from RNA-sequencing (RNA-seq) data. D and E: Representative transmission electron microscopy images of ovarian tissues and quantification of number of mitochondria in oocytes and abnormal mitochondria in GCs, as assessed by transmission electron microscopy in control versus tumor group. D: Right panels: Expanded images of the regions highlighted by the boxed areas (middle panels). Hollow arrowhead indicates the endoplasmic reticulum (ER); arrowheads, Golgi apparatuses (Gs); and arrows, lipid droplet (LP). F: The levels of reactive oxygen species in serum and ovarian tissues of control and tumor-bearing mice were detected by enzyme-linked immunosorbent assay. *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001 indicate significant differences between control and experimental groups. Scale bars: 500 mm (D, left panels); 2 mm (D, middle panels, and E, left panels); 1 mm (D and E, right panels). FDR, false discovery rate; FPKM, fragments per kilobase million; GSH, glutathione; M, mitochondria; MDA, malondialdehyde; N, nucleus; NES, Normalized Enrichment Score; OC, oocyte cytoplasm; SOD, superoxide dismutase; V, vacuole; ZP, zona pellucida. 618 ajp.amjpathol.org - The American Journal of Pathology
Tumor-Triggered Ovarian Follicle Loss Figure 6 Antioxidant treatment reduces tumor-derived follicle loss and restores hormonal balance. A: Schematic of experimental design whereby female BALB/c nude mice (aged 4 to 6 weeks) were split into two groups: control and tumor (i.p. injections MDA-MB-231 for 4 weeks). The tumor-bearing mice were subjected to treatment of i.p. injections of vitamin C (VC) as VC group, and the tumor and control group were glucose treated for 2 weeks. B: The repre- sentative ovary gross images and statistics of ovary/body weight ratio in control, tumor, and VC groups. C: Quantification of ovarian follicles in the three groups. D: Images of immunofluorescence showed the nuclei of the oocyte of primordial follicles (dashed lines) were detected using an antieDEAD-box helicase 4 (anti-DDX4) antibody in each treatment group. All of the nuclei were stained with DAPI. E: Immunofluorescence staining of proliferating cell nuclear antigen (PCNA)epositive granulosa cells (GCs; white arrowheads) in follicles at different development stage, and quantification of positive GC rate per follicle in the three groups. Left panels: Insets: Higher magnifications of the dotted boxed areas. F: Quantification of atretic follicles in the three groups. G: Immunofluorescence staining of terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL)epositive GCs in growing follicles, and quantification of positive GC rate per follicle in the three groups. H: Immunofluorescence staining of cleaved caspase-3epositive GCs in growing follicles, and quantification of positive GC rate per follicle in the three groups. n Z 5 per group (F). *P < 0.05, ***P < 0.001, and ****P < 0.0001 indicate significant differences between the three groups. Scale bars: 50 mm (D and E); 200 mm (G and H). AF, antral follicle; D, day; PDF, primordial follicle; SF, secondary follicle. The American Journal of Pathology - ajp.amjpathol.org 619
Wu et al
Figure 7 A: Western blot analyses of phosphatase regulator and tensin homolog (PTEN) in the ovaries of three groups, and protein quantification was
analyzed by being normalized to b-actin. B: Immunostaining for nuclear localization of forkhead box O3a (FOXO3a) and phosphorylated FOXO3a (p-FOXO3a)
expression (dashed circles) and decreased localization shuttling of p-FOXO3a after vitamin C (VC) treatment. Insets: Higher magnifications of the dotted
boxed areas. C: Representative immunoblot analysis (left panel) and qualification (right panel) of key proteins of phosphatidylinositol 3-kinaseeAkt and
mammalian target of rapamycin signaling in ovaries of control, tumor, and VC-treated mice. *P < 0.05, **P < 0.01, and ***P < 0.001 indicate significant
difference between the three groups. Scale bar Z 50 mm (B). p-Akt, phosphorylated Akt; p-RPS6, phosphorylated ribosomal protein S6.
prevents tumor-derived follicle loss via suppression of the High serum FSH levels induce the acceleration of follicular
PI3K-Akt-mTOR pathway. initiation and subsequent follicle loss.39 Estradiol reflects the
function of the preantral follicles, and has a negative feed-
back on FSH.40 Most young women with breast cancer
Discussion experience estrogen deficiency even before starting cancer
treatment,13 and approximately one-sixth of them are diag-
Although cryopreservation of ovarian tissue is helpful to nosed with triple-negative tumors.41 In the current study, a
young women with breast cancer, the preferred cryopreser- triple-negative breast cancer cell line, MDA-MB-231,
vation treatment for a successful outcome is yet to be deter- xenograft model was generated to study the mechanisms
mined. While normal ovarian function is restored in involved in follicular growth or loss in the grafting ovaries of
approximately 85% of patients undergoing auto- patients with breast cancer.19 In this study, the elevation of
transplantation, only approximately 40% of them achieve FSH and decrease of estradiol indicated premature ovarian
pregnancy and delivery.13,35,36 Individual ovarian reserve and, insufficiency associated with breast cancer in humans and
thus, follicle density, at the time of harvesting has the largest mice. The tumor-bearing ovaries had low follicle density,
influence on success rates of pregnancy and live births.37 suggesting that an underlying cause of these conditions may
However, the quantity of the ovarian reserve before cancer be a tumor-induced alteration in the ovaries.
treatment has not been considered, with most attention An uncontrolled and increased activation of the primor-
focusing on ovarian failure after chemotherapy. Using a mouse dial follicles leads to the development of premature ovarian
model, the current study demonstrates, for the first time, insufficiency.38 Herein, in tumor-bearing ovaries, follicle
tumor-induced loss of primordial and growing follicles, count and ovary size assessment demonstrated destruction
resulting from abnormal hormone regulation in breast cancer. of both primordial and growing follicles. The increase in the
The number of follicles were shown to be lost to either growth ratio of growing/primordial follicles also reflected the loss
or atresia, which was linked to oxidative stressetriggered of a greater number in primordial follicles than growing
activation of the PI3K-Akt-mTOR pathway. Therefore, pa- follicles, suggesting that the primordial follicles were
tients with breast cancer may need to consider therapies to largely responsible for this follicular depletion.42 Primordial
improve follicle health before ovarian cryopreservation. follicle activation is a process by which primordial follicles
Premature ovarian insufficiency pertains to ovarian are selected into the growing follicle pool, and is charac-
dysfunction for women aged 25 mIU/mL) and low levels of estradiol.38 number of proliferating GCs in tumor-being ovaries.
620 ajp.amjpathol.org - The American Journal of PathologyTumor-Triggered Ovarian Follicle Loss PI3K-Akt and mTOR signaling pathways can be acti- primordial and growing follicles and reduced follicular vated in oocytes or GCs, and a two-celletwo-pathway density in ovaries of tumor-bearing mice. model has been proposed to control follicle activation.43,44 Cancer appears to impair the cellular mitochondrial In general, active mTOR signaling enhances the expres- network leading to excess ROS production. Increased sion of KIT proto-oncogene receptor tyrosine kinase (KIT) oxidative stress levels and low antioxidant status are ligand in pre-GCs and KIT ligandeKIT signaling serves as detected in serum samples of patients with breast can- the bridge between the pre-GCs and the oocyte, resulting in cer.12,49 In the current study, similar findings were found in activation of intra-oocyte PI3K-Akt signaling (Figure 4F).44 the ovarian tissues of tumor-bearing mice. ROS generation In this study, FOXO3a was highly expressed in primordial can result from oxidative phosphorylation occurring in the follicles and shunting of FOXO3a to the cytoplasm, and the mitochondria. Transmission electron microscopy assess- relative level of PTEN was significantly decreased, whereas ment showed damage to mitochondria in both the oocyte phosphorylation of FOXO3a, Akt, and RPS6 were and GCs. Enriched GO terms of down-regulated genes were increased, suggesting activation of primordial follicles mainly related to mitochondrial function and ROS response, through PI3K-Akt and mTOR signaling in ovaries of tumor- and Kyoto Encyclopedia of Genes and Genomes pathway bearing mice. analysis showed that oxidative phosphorylation was in the After activation, the follicles that are not selected for down-regulated group in ovarian tissues of tumor-bearing further development undergo atresia. Herein, the ovaries of mice. The involvement of this process was confirmed by tumor-bearing mice showed high levels of atretic features, the determination of serum and ovarian oxidative stress and and atretic follicles (GCs and/or oocytes) were terminal antioxidant response levels, thus suggesting oxidative stress deoxynucleotidyl transferase-mediated dUTP nick-end la- occurs in patients with breast cancer and tumor-bearing beling- and cleaved caspase-3epositive. Although apoptosis mice. is minimal in preantral follicles of normal ovaries,16 it occurs Although ROS play a positive role in ovarian biology, in follicles destined to be recruited to the growing pool and excessive ROS can directly damage the intraovarian envi- likely to enter atresia. These results suggested that follicles ronment, just as they do in the oocyte and GCs/TCs.13 are lost from the pool either indirectly by activation of the Vitamin C is a known oxidation-reduction catalyst that can growth pathway or directly through induction of apoptotic reduce and neutralize ROS in GCs in vitro, as well as in a cell death in tumor-bearing ovaries. premature ovarian insufficiency mouse model.50,51 Fertility The balance of the hormones and intra-ovarian signals is benefits of vitamin C include regulation of steroidogenesis critical to determine whether follicles will complete folli- via steroidogenic acute regulatory protein signaling in GCs culogenesis or undergo atresia. The GCs and TCs are a site in vitro50 and follicle growth in cattle ovarian fragments.52 for steroidogenesis. Indeed, steroidogenesis is initiated by Here, breast cancereinduced ovarian alterations included steroidogenic enzymes in TCs and then subsequently accelerated loss of ovarian reserve, folliculogenesis dysre- aromatized in GCs.45 Within follicles, interactions between gulation, and steroid disorder, and ROS signaling was shown oocytes and GCs/TCs regulate folliculogenesis and the to contribute to oocyte and GC/TC interaction via PI3K- development of dominance through paracrine factors, such AKT-mTOR signaling inducing cellular proliferation/differ- as secretion of growth factors, modulation of KIT ligands, entiation and apoptosis. Furthermore, current studies in the and steroidogenesis.46,47 Evidence from human follicular mouse model showed that supplementation with vitamin C fluid supports that decreased estradiol production is the promoted follicular quiescence, repaired tumor-induced principal cause for the secondary increase in FSH, and the ovarian damage through restoration of the total number of increase in plasma FSH controls selection of dominant follicles, and maintained normal serum FSH and estrogen follicles.48 More importantly, PI3K-Akt and mTOR levels, thereby preserving ovarian function (Supplemental signaling may function downstream of FSH. Signaling Figures S6 and S7). The PI3K-Akt-mTOR pathway may be mediated by PTEN, FOXO3a, and transforming growth the major potential target to prevent follicle activation and factor-b family members (bone morphogenetic protein 15, loss following ovarian transplantation, and PTEN inhibitor growth differentiation factor-9, and AMH) is involved in the increases DNA damage capacity in oocyte in vitro activa- regulation of proliferation, differentiation, and apoptosis of tion.27 Thus, inactivation of PI3K-Akt-mTOR pathway by GCs in follicles.46 Therefore, the disturbed balance between using an antioxidant may have additional value as a prom- proliferation and apoptosis in GCs and TCs can influence ising strategy for the derivation of healthy follicles and folliculogenesis or follicle atresia. Atresia of primordial functional ovarian tissues for patients with breast cancer. follicles is believed to follow oocyte death, whereas atresia There are several limitations to the current study. First, a of growing follicles primarily begins in apoptotic GCs.24 link between the lowered hormone levels observed in The current results indicate a decrease in expression of human breast cancer and ovarian reserve has not been AMH, FSH receptor, and steroidogenic enzymes, suggest- shown, and further studies are required to confirm the ing that the altered cell-cell interactions between oocytes findings from the mouse model. Second, the efficacy of only and GCs/TCs are associated with impaired folliculogenesis, one antioxidant (vitamin C) was examined within the mouse which, in turn, results in increased follicular atresia in both model of breast cancereinduced loss of ovarian reserve. The American Journal of Pathology - ajp.amjpathol.org 621
Wu et al
Finally, the health of cryopreserved ovarian tissue after 11. Ladanyi C, Mor A, Christianson MS, Dhillon N, Segars JH: Recent
vitamin C supplementation was not examined. advances in the field of ovarian tissue cryopreservation and oppor-
tunities for research. J Assist Reprod Genet 2017, 34:709e722
The current study demonstrates in a xenograph mouse 12. Jelic MD, Mandic AD, Maricic SM, Srdjenovic BU: Oxidative stress
model a possible link between breast cancer and ovarian and its role in cancer. J Cancer Res Ther 2021, 17:22e28
reserve that may be enhanced by vitamin C supplementa- 13. Zhang T, Zhu X, Wu H, Jiang K, Zhao G, Shaukat A, Deng G, Qiu C:
tion. However, a link between altered hormone levels in Targeting the ROS/PI3K/AKT/HIF-1a/HK2 axis of breast cancer
breast cancer and ovarian reserve in women has not been cells: combined administration of polydatin and 2-deoxy-d-glucose. J
Cell Mol Med 2019, 23:3711e3723
established and needs additional investigation. 14. Dri M, Klinger FG, De Felici M: The ovarian reserve as target of
insulin/IGF and ROS in metabolic disorder-dependent ovarian dys-
functions. Reprod Fertil 2021, 2:R103eR112
Author Contributions 15. Fabbri R, Vicenti R, Martino NA, Dell’Aquila ME, Pasquinelli G,
Macciocca M, Magnani V, Paradisi R, Venturoli S: Confocal laser
Y.W., J.H., H.C., and G.Y. performed the experiments; scanning microscopy analysis of bioenergetic potential and oxidative
Y.H. analyzed and interpreted the data; H.T. and Q.Z. stress in fresh and frozen-thawed human ovarian tissue from onco-
logic patients. Fertil Steril 2014, 101:795e804
recruited the patients and obtained serum specimens; G.E.L. 16. Myers M, Britt KL, Wreford NG, Ebling FJ, Kerr JB: Methods for
edited the manuscript; and P.L. conceived the study, pro- quantifying follicular numbers within the mouse ovary. Reproduction
vided financial support, and wrote the manuscript. 2004, 127:569e580
17. Kitajima M, Dolmans MM, Donnez O, Masuzaki H, Soares M,
Donnez J: Enhanced follicular recruitment and atresia in cortex derived
Supplemental Data from ovaries with endometriomas. Fertil Steril 2014, 101:1031e1037
18. Wang Y, Zhang Y, Wu Y, He Y, Xiang J, Huang J, Lash GE, Li P:
SIRT1 regulates trophoblast senescence in premature placental aging
Supplemental material for this article can be found at
in preeclampsia. Placenta 2022, 122:56e65
http://doi.org/10.1016/j.ajpath.2023.01.015. 19. Huang Z, Yu P, Tang J: Characterization of triple-negative breast
cancer MDA-MB-231 cell spheroid model. Onco Targets Ther 2020,
13:5395e5405
References 20. Cora MC, Kooistra L, Travlos G: Vaginal cytology of the laboratory
rat and mouse: review and criteria for the staging of the estrous cycle
1. Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, using stained vaginal smears. Toxicol Pathol 2015, 43:776e793
Jemal A, Bray F: Global cancer statistics 2020: GLOBOCAN esti- 21. Bernhardt SM, Dasari P, Walsh D, Townsend AR, Price TJ,
mates of incidence and mortality worldwide for 36 cancers in 185 Ingman WV: Hormonal modulation of breast cancer gene expression:
countries. CA Cancer J Clin 2021, 71:209e249 implications for intrinsic subtyping in premenopausal women. Front
2. Fan L, Strasser-Weippl K, Li JJ, St Louis J, Finkelstein DM, Yu KD, Oncol 2016, 6:241
Chen WQ, Shao ZM, Goss PE: Breast cancer in China. Lancet Oncol 22. Regan SLP, Knight PG, Yovich JL, Leung Y, Arfuso F,
2014, 15:e279ee289 Dharmarajan A: Granulosa cell apoptosis in the ovarian follicle-a
3. Co M, Kwong A: Young onset breast cancer in Southern China - a 5- changing view. Front Endocrinol 2018, 9:61
year clinico-pathological study from a multi-centre database. Cancer 23. John GB, Gallardo TD, Shirley LJ, Castrillon DH: Foxo3 is a PI3K-
Treat Res Commun 2020, 24:100182 dependent molecular switch controlling the initiation of oocyte
4. Hickey M, Peate M, Saunders CM, Friedlander M: Breast cancer in growth. Dev Biol 2008, 321:197e204
young women and its impact on reproductive function. Hum Reprod 24. Reddy P, Liu L, Adhikari D, Jagarlamudi K, Rajareddy S, Shen Y,
Update 2009, 15:323e339 Du C, Tang W, Hämäläinen T, Peng SL, Lan ZJ, Cooney AJ,
5. Gonçalves V, Sehovic I, Quinn G: Childbearing attitudes and de- Huhtaniemi I, Liu K: Oocyte-specific deletion of Pten causes premature
cisions of young breast cancer survivors: a systematic review. Hum activation of the primordial follicle pool. Science 2008, 319:611e613
Reprod Update 2014, 20:279e292 25. Maidarti M, Clarkson YL, McLaughlin M, Anderson RA, Telfer EE:
6. Wan Q, Han L, Liu J, Li H, Li Y, Long QM, Tan L, Li JJ: Fertility Inhibition of PTEN activates bovine non-growing follicles in vitro but
preservation among young breast cancer patients: a single-center increases DNA damage and reduces DNA repair response. Hum
experience in China. Taiwan J Obstet Gynecol 2021, 60:827e830 Reprod 2019, 34:297e307
7. Arecco L, Blondeaux E, Bruzzone M, Ceppi M, Latocca MM, 26. De Felici M, Klinger FG: PI3K/PTEN/AKT signaling pathways in
Marrocco C, Boutros A, Spagnolo F, Razeti MG, Favero D, germ cell development and their involvement in germ cell tumors and
Spinaci S, Condorelli M, Massarotti C, Goldrat O, Del Mastro L, ovarian dysfunctions. Int J Mol Sci 2021, 22:9838
Demeestere I, Lambertini M: Safety of fertility preservation tech- 27. Terren C, Nisolle M, Munaut C: Pharmacological inhibition of the
niques before and after anticancer treatments in young women with PI3K/PTEN/Akt and mTOR signalling pathways limits follicle acti-
breast cancer: a systematic review and meta-analysis. Hum Reprod vation induced by ovarian cryopreservation and in vitro culture. J
2022, 37:954e968 Ovarian Res 2021, 14:95
8. Arav A, Patrizio P: Techniques of cryopreservation for ovarian tissue 28. Devillers MM, Petit F, Cluzet V, François CM, Giton F, Garrel G,
and whole ovary. Clin Med Insights Reprod Health 2019, 13: Cohen-Tannoudji J, Guigon CJ: FSH inhibits AMH to support
1179558119884945 ovarian estradiol synthesis in infantile mice. J Endocrinol 2019, 240:
9. Gjeterud J, Kristensen SG, Fedder J: [Indications for cryopreservation 215e228
and autotransplantation of ovarian tissue] Norwegian. Tidsskr Nor 29. Roy S, Gandra D, Seger C, Biswas A, Kushnir VA, Gleicher N,
Laegeforen 2021, 141. doi:10.4045/tidsskr.21.0030 Kumar TR, Sen A: Oocyte-derived factors (GDF9 and BMP15) and
10. Hossay C, Donnez J, Dolmans MM: Whole ovary cryopreservation FSH regulate AMH expression via modulation of H3K27AC in
and transplantation: a systematic review of challenges and research granulosa cells. Endocrinology 2018, 159:3433e3445
developments in animal experiments and humans. J Clin Med 2020, 30. Manna PR, Ahmed AU, Yang S, Narasimhan M, Cohen-Tannoudji J,
9:3196 Slominski AT, Pruitt K: Genomic profiling of the steroidogenic acute
622 ajp.amjpathol.org - The American Journal of PathologyYou can also read

















































