TRP Channels and Migraine: Recent Developments and New Therapeutic Opportunities - MDPI
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

pharmaceuticals
Review
TRP Channels and Migraine: Recent Developments
and New Therapeutic Opportunities
Silvia Benemei 1 and Greg Dussor 2, *
1 Headache Centre, Careggi University Hospital, Viale Pieraccini 18, 50139 Florence, Italy;
silvia.benemei@unifi.it
2 School of Behavioral and Brain Sciences, Center for Advanced Pain Studies,
The University of Texas at Dallas, Richardson, TX 75080, USA
* Correspondence: gregory.dussor1@utdallas.edu; Tel.: +1-972-883-2385
Received: 1 March 2019; Accepted: 4 April 2019; Published: 9 April 2019
Abstract: Migraine is the second-most disabling disease worldwide, and the second most common
neurological disorder. Attacks can last many hours or days, and consist of multiple symptoms
including headache, nausea, vomiting, hypersensitivity to stimuli such as light and sound, and in
some cases, an aura is present. Mechanisms contributing to migraine are still poorly understood.
However, transient receptor potential (TRP) channels have been repeatedly linked to the disorder,
including TRPV1, TRPV4, TRPM8, and TRPA1, based on their activation by pathological stimuli
related to attacks, or their modulation by drugs/natural products known to be efficacious for migraine.
This review will provide a brief overview of migraine, including current therapeutics and the
link to calcitonin gene-related peptide (CGRP), a neuropeptide strongly implicated in migraine
pathophysiology. Discussion will then focus on recent developments in preclinical and clinical
studies that implicate TRP channels in migraine pathophysiology or in the efficacy of therapeutics.
Given the use of onabotulinum toxin A (BoNTA) to treat chronic migraine, and its poorly understood
mechanism, this review will also cover possible contributions of TRP channels to BoNTA efficacy.
Discussion will conclude with remaining questions that require future work to more fully evaluate
TRP channels as novel therapeutic targets for migraine.
Keywords: ion channel; TRP; cortical spreading depression; meninges; dura mater; botulinum toxin
A; reactive oxygen species; reactive nitrogen species; neurogenic inflammation; CGRP
1. Migraine: Epidemiology and Symptoms
Migraine is one of the most common diseases worldwide affecting over 14% of the adult
population [1], with a gendered distribution in favor of females (2:1), and an increased prevalence
in reproductive age for both genders [2]. Importantly, migraine is the most common severe primary
headache, characterized by long-lasting (4–72 hours) attacks of pain, accompanied by nausea,
vomiting, and sensitivity to light and sound [1]. In current clinical practice, diagnosis of a migraine
is made in accordance to the criteria of the International Headache Society (IHS) [3], which also
allow the homogenous identification of the migraine phenotype worldwide for research purposes.
According to the current version of the International Classification of Headache Disorders (ICHD-3) [3],
the migraine can be with or without an aura (i.e., transient neurological symptoms, including visual
or somatosensory or speech disturbances, that precede and/or accompany and/or follow the pain)
and, in case of attacks for more than 15 days for 3 consecutive months, it can be classified as chronic.
The most recent results from the Global Burden of Disease (GBD) study, promoted by the World Health
Organization, reported migraine as the most disabling neurological disorder, and the second leading
cause of years lived with disability (YLDs) worldwide [4]. The high disability, combined with the
Pharmaceuticals 2019, 12, 54; doi:10.3390/ph12020054 www.mdpi.com/journal/pharmaceuticalsPharmaceuticals 2019, 12, 54 2 of 17
high prevalence, causes a significant socio-economic burden. In the European Union, where migraine
is estimated to affect 75 million patients, each year it costs ~111 billion €, of which 93% is given for
absence from work and reduced productivity [5].
2. Migraine: Therapeutics and Pathophysiology
Therapeutic options for the treatment of migraine include drugs for both the attacks, aimed
to abort ongoing pain and reduce migraine-related symptoms, and prophylaxis. Acute treatment
of attacks is recommended in any episode in any patient, while prophylaxis is recommended in
the case of patients suffering more than four attacks/days of headache per month. Currently
available and recommended drugs for the acute treatment of attacks encompass analgesics (i.e.,
paracetamol), non-steroidal anti-inflammatory drugs (NSAIDs), triptans, antiemetics and combinations
of analgesics; while ergot derivatives and opioids maintain only a marginal role in therapy. Preventive
treatments are to a large extent borrowed from other therapeutic areas, including antihypertensive
agents (e.g., angiotensin-converting enzyme [ACE] inhibitors, angiotensin receptor blockers, beta
blockers), antiepileptic drugs (e.g., topiramate, sodium valproate, gabapentin), tricyclic antidepressants
(e.g., amitriptyline, nortriptyline), but there are also some drugs used only with migraine prevention
objectives, such as flunarizine and pizotifene [1]. Importantly, recent clinical evidence has supported
the inclusion of onabotulinum toxin A (BoNTA) among the recommended drugs for migraine
prevention, but only for chronic patients [6,7].
Mechanisms contributing to migraine are multifactorial, complex, and not yet completely
clarified [8–10]. Notwithstanding the huge number of studies, including genome-wide association
studies (GWAS), the role of genetic factors still remains poorly defined [11], with the exception of
hemiplegic migraine, which has, for a good portion of cases, a classical mendelian transmission.
After years of open debate that has counter posed the neuronal and the vascular theory, migraine
is now considered a neurovascular disorder, with a primary role of the trigeminovascular system
in the mechanism of pain [12]. Following its first proposal, the neurovascular hypothesis has been
corroborated by preclinical and clinical data showing the involvement of neurogenic inflammation
and, in particular, of the neuropeptide calcitonin gene related peptide (CGRP) [12].
Neurogenic inflammation is a sterile inflammation mediated by the activation of trigeminal
perivascular fibers that release neuropeptides, such as substance P and—most importantly in
humans—CGRP, that induce increased blood flow and edema [12,13]. Importantly, it has been shown
in preclinical models that the activation of meningeal nociceptors could stimulate the trigeminal
ganglia [14], thus in turn perpetuating the release of vasoactive peptides, including CGRP.
3. CGRP and Migraine
CGRP, a neuropeptide produced from alternative splicing of the calcitonin gene, acts as a relevant
signaling peptide in mammalian biology, with a crucial role in both physiological and pathological
conditions. In particular, because of its potent action as a vasodilator, it is involved in the physiological
regulation of vascular tone and blood pressure, and some evidence has been collected for a role in
cardiovascular diseases and extra-cardiovascular conditions [15].
There are two CGRP forms differently expressed in humans, alpha-CGRP (primary sensory
neurons of the dorsal root ganglia, vagal ganglia and trigeminal system) and beta-CGRP (intrinsic
enteric neurons) [16–18]. According to the expression pattern, alpha-CGRP is primarily involved in
migraine pathogenesis. CGRP acts via a G protein-coupled receptor constituted by the calcitonin
receptor-like receptor (CLR), the receptor activity-modifying protein 1 (RAMP1) and a receptor
component protein (RCP), all of which are necessary to form the functional CGRP receptor [12].
In the last decades, both preclinical and clinical evidence has accumulated supporting a role of CGRP
in migraine mechanisms. First, CGRP levels are increased during a migraine attack [19] and in
chronic migraine patients also in the pain-free interval [20], returning to normal levels after triptan
administration associated with headache resolution [21–24]. Second, intravenous infusion of CGRP canPharmaceuticals 2019, 12, 54 3 of 17
induce migraine-like attacks in migraine patients [25,26]. Third, animal data suggest that CGRP can
induce the generation of light intolerance, which may be considered a parallel of human photophobia,
a phenomenon typically reported during migraine attack [27]. According to the abovementioned
evidence, CGRP is considered a relevant target for migraine, and many efforts have been made to
show that CGRP antagonism, by small molecules antagonists of CGRP receptor [28–33], an anti-CGRP
receptor antibody [34–36] and anti-CGRP peptide antibodies [37–45] are all efficacious for the treatment
or prevention of migraine.
4. Recent Findings on TRP Channels and Migraine Pain
While the mechanisms leading to the pain phase of migraine are unclear, the most likely
explanation is that activation of nociceptors innervating the cranial meninges (and possibly the
cerebral/meningeal vasculature) is responsible for the generation of headache [12,46]. Dilation of
meningeal vessels on the headache side during migraine attacks triggered by cilostazol has recently
been found, a finding that the authors suggest is an indicator of meningeal nociceptor activation [47],
but this can only be taken as correlation and not causation of headache at this point. However,
little information currently exists as to what specific mechanisms cause the activation of nociceptors in
the meninges and/or near vessels. This includes a lack of knowledge of the extracellular events within
the tissues themselves that occur during migraine attacks and the receptors expressed on nociceptors
that respond to these events.
Transient receptor potential (TRP) channels are a family of cation channels expressed primarily
on the cell membrane that cluster into six families including TRPA, TRPC, TRPM, TRPP, TRPL,
and TRPV [48–50]. These channels are likely to contribute to a number of different physiological
processes ranging from thermosensation and pain to regulation of Ca2+ levels in the endoplasmic
reticulum. Of relevance to the current discussion, they have been repeatedly hypothesized to contribute
to migraine, specifically as an activation mechanism of meningeal nociceptors. Reasons for the interest
in a TRP channel contribution to migraine are largely due to their expression on meningeal nociceptors
and their responsiveness to a variety of endogenous and exogenous stimuli that may be of relevance to
migraine attacks (Figure 1). Further, activation of TRP channels is well-known to promote the release
of CGRP from sensory nerve endings, and they have been used extensively as probes for the function
of CGRP in various processes [15,51,52]. Given the role of CGRP in migraine described above, and the
as yet unclear mechanism by which CGRP is released during migraine, TRP channels remain a focus
of interest for their potential contribution to attacks. A number of reviews on the topic of TRP channels
and migraine have been published in recent years documenting the evidence that exists supporting
their role in meningeal nociceptor activation [51,53–58], so only brief discussion of older work will be
given here, while the focus will be on newer TRP channel migraine studies.
4.1. TRPV1
Based primarily on preclinical animal studies, it is clear that sensory neurons that innervate the
meninges express TRPA1, TRPV1, TRPV4, and TRPM8 [51,53]. This is not likely to be an all-inclusive
list as other TRP channels have not yet been extensively studied in this context. One of the first
TRP channels to be investigated was TRPV1, which is activated by capsaicin and also sensitive to
endocannabinoids, endovanilloids, lipoxygenase metabolites, nerve-growth factor, and prostaglandins,
among other factors many of which may be relevant for migraine [48]. TRPV1 was hypothesized to be
present on meningeal sensory fibers based on early studies with capsaicin application to the meninges
(e.g. [59]). It was later directly demonstrated on these neurons using immunohistochemistry [60] and
recently shown on nerve fibers in the arterial wall of scalp vessels from chronic migraine patients,
where channel expression was higher compared to healthy controls [61]. Mechanisms leading to
increased TRPV1 expression in chronic migraine patients are unclear, but preclinical studies using
repeated 30-day administration of eletriptan or indomethacin found increased TRPV1 (and TRPA1) inPharmaceuticals 2019, 12, 54 4 of 17
the trigeminal ganglia, so migraine itself may upregulate the channel and/or migraine therapeutics
may also have this potential [62].
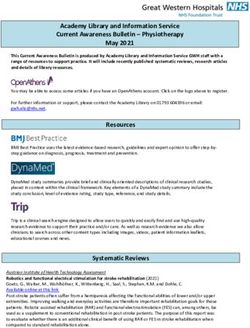
Figure 1. TRP channels on trigeminal afferents and their potential role in migraine pathology.
Multiple TRP channels are expressed on trigeminal sensory neurons innervating the meninges
including TRPV1, TRPA1, TRPV4, and TRPM8. These channels respond to stimuli implicated in
migraine, both from a pathology perspective (e.g. acrolein on TRPA1) and a therapeutic perspective
(e.g. parthenolide on TRPA1). Additional modulators are listed below their respective TRP channels.
Activation of TRP channels on meningeal afferents leads to action potential signaling into the trigeminal
nucleus caudalis (left) and ultimately to headache. Activation of TRP channels on these neurons also
leads to the release of neuropeptides such as CGRP, activating CGRP receptors on blood vessels
(right and bottom), causing vasodilation and contributing to neurogenic inflammation. Although not
shown, TRP channels are also expressed on the central terminals of meningeal afferents, and CGRP
is released as a transmitter in this synapse, both of which may also contribute to signaling within
this circuit. Multiple migraine therapeutics may act in this circuit, including: BoNTA, which may
indirectly contribute to decreased CGRP release and possibly inhibit recruitment of TRP channels to the
membrane; GEPANTs, which block the CGRP receptor; antiCGRP mAbs, which sequester extracellular
CGRP; and antiCGRP-R mAbs, which bind to and block the CGRP receptor. Abbreviations are found
in the lower right corner.
There have been numerous studies using capsaicin and TRPV1 antagonists to probe meningeal
afferent and vascular function and these studies suggest a role for the channel in headache
mechanisms [51]. Recent studies using this approach have found differential capsaicin-induced
meningeal blood flow changes in the presence of diet-induced obesity [63] in mice and suggest a role
for TRPV1 signaling in the increased prevalence of migraine with obesity (consistent with the increase
in capsaicin-induced nociception and photophobia with obesity [64]; a similar finding was also seen
with TRPA1 [65]). Capsaicin stimulation of facial skin in human has also been used as a surrogate
to study trigeminal-mediated dural vasodilation [66]. Using this assay, increased dermal blood flow
was seen in female (but not male) migraine patients compared to controls, suggesting that changesPharmaceuticals 2019, 12, 54 5 of 17
occur in TRPV1 fiber neurovascular responses in migraine [67]; similar changes in migraine patients in
capsaicin-induced dermal blood flow were also recently published [68].
The above studies still do not address the mechanisms by which TRPV1 may be activated
in the meninges during migraine attacks, they only show that the function of the channel or
TRPV1-expressing fibers are altered with migraine-like conditions. In one of the few recent studies
addressing a potential mechanism for TRPV1 activation, the authors used two-photon in vivo imaging
of anesthetized mouse to examine macrophage morphology and dendritic cell motility in the meninges
in relation to cortical-spreading depression (CSD), which is thought to be the underlying basis of
migraine aura and has been shown to activate meningeal nociceptors [69,70]. In time with CSD,
meningeal macrophages changed to an active-like shape and migration of dendritic cells stopped [71].
The functional consequences of either of these events is not clear, but both cell types were found
near TRPV1-expressing neurons, suggesting that CSD may alter meningeal immune cell function,
which subsequently signal to this population of nociceptors. These studies are consistent with earlier
findings on meningeal immune cells where CSD caused degranulation of mast cells [72], and together
further implicate [73] changes in meningeal immune cell function during migraine as one mechanism
of activation of dural nociceptors. Ultimately, while there may be a role for this channel in meningeal
afferent signaling and in headache, the TRPV1 antagonist SB-705498 failed in a clinical migraine
study [74]. This does not exclude a possibility for TRPV1 in migraine pathology, but it casts serious
doubt on selective TRPV1 antagonism as a therapeutic approach and suggests that activation of TRPV1
alone is not sufficient to generate the headache phase of migraine.
4.2. TRPV4
The TRPV4 channel responds to a number of stimuli including changes in osmolarity
(cell swelling) and the resulting mechanical forces imposed on the cell membrane [75–77], suggesting
that this channel serves as part of a mechanosensory complex. It is thus interesting in the context of
migraine as headache is known to be influenced by changes in intracranial pressure e.g. when coughing,
sneezing, standing or sitting, or exercising [78]. TRPV4 is also expressed in the vasculature where its
activity likely contributes to vascular permeability, among other functions [79]. Expression of TRPV4
on meningeal nociceptors was shown using application of channel activators to retrogradely-labeled
trigeminal neurons in vitro, which produced currents consistent with TRPV4, and activation of this
channel in the dura caused headache behavioral responses in rats [80]. Unfortunately, little work
has been done directly examining TRPV4 and headache since this publication. TRPV4 function
in sensory neurons is known to be modulated downstream of the protease-activated receptor 2
(PAR2) [81–83] and PAR2 activation sensitizes meningeal nociceptors to mechanical stimulation [84].
Until recently, preclinical behavioral studies had not been conducted with PAR2 activation in the
meninges. Using more selective PAR2 agonists than those in prior studies, it was found that dural
PAR2 activation causes headache behaviors in mice that were blocked by a selective PAR2 antagonist
and absent in PAR2 knockout mice [85]. These studies provide further indirect evidence that TRPV4
activity on meningeal nociceptors may contribute to headache, but it remains unclear whether TRPV4
plays a direct role in the ability of meningeal afferents to detect pressure changes. More studies are
needed to better explore the potential role for this channel in migraine.
4.3. TRPM8
Non-noxious cool temperature, natural products such as menthol, and synthetic compounds
such as icilin are all known to activate TRPM8; this channel has been implicated in pain signaling
in numerous prior studies [86,87]. It has also been the subject of much interest in the migraine
community, initially based on the repeated identification of single-nucleotide polymorphisms (SNPs)
and variants in the channel gene in numerous GWAS [53,87], most recently in a Taiwanese population
study [88]. TRPM8 is found on trigeminal nerve fibers innervating the dura and appears to undergo
developmental regulation where decreased expression occurs with age in mice such that relatively fewPharmaceuticals 2019, 12, 54 6 of 17
neurons express TRPM8 in adult [89,90]. There have been conflicting reports as to what behavioral
consequences are present when TRPM8 is activated in the meninges since studies have shown both
increases [91] and decreases [89] in headache behavior in rodents following dural application of
agonists. Ultimately, the context of when channel activation occurs may be relevant; if TRPM8 is
activated alone it may cause headache as in the case of “ice cream headaches” while if it is activated
along with ongoing inflammation, it may serve as an analgesic mechanism as is the case for topical
menthol [53]. Consistent with this idea is a recent publication demonstrating that when TRPM8 is
activated in the presence of an ongoing inflammatory state induced by a cocktail of inflammatory
mediators, the effect was a reduction in headache-related behavior [92]. These authors also found
increased overlap of TRPV1 and TRPM8 in the same trigeminal neurons innervating the dura in the
presence of meningeal inflammation, suggesting changes in channel expression occur under these
conditions. Interestingly, while TRPV1 and TRPM8 are not co-expressed in dorsal root ganglia [93],
co-expression has been found in trigeminal neurons [94], although as mentioned above, its expression
is subject to developmental regulation. Using a PC12 cell-based assay, they also found that TRPM8
activation could inhibit function of TRPV1 when both channels were expressed by the cells. Together,
these new studies support the context-specific role of TRPM8 in migraine. The context of channel
activation during native migraine attacks is not yet clear, so it remains unknown what role the channel
plays. This latter point is particularly relevant for drug discovery as it is not known whether an
agonist or antagonist would show efficacy for migraine. Nonetheless, Amgen recently published
preclinical pharmacology and human safety data on a novel TRPM8 antagonist proposed to be tested
for migraine [95].
One of the more intriguing studies related to TRPM8 and migraine in recent years was the finding
that the TRPM8 SNP rs10166942 (either C or T at chr2:234835093), which has been identified in GWAS of
migraine populations [53], is differentially expressed across the Earth depending on latitude [96]. It was
previously reported that humans who carry the T;T allele have a higher risk of migraine than those
carrying the rarer versions C;C or C;T [5,97]. The authors of this new study found that the frequency
of T-allele expression was only 5% in populations native to Nigeria while it was 88% in those native to
Finland, with South and East Asian populations between these extremes at 48% and 36%, respectively.
Thus, T-allele frequency seems to be predicted by a latitudinal cline, suggesting that human migraine to
the colder climates of Northern Europe introduced positive-selection pressure on TRPM8. Additionally,
this further suggests that the variants in the channel may respond differentially to temperature (possibly
with lower activity in colder climates which would facilitate adaptation to those environments),
although it is not yet known what these variants do to channel function/expression. Nonetheless,
the latitudinal cline of T-allele expression (higher in colder climates, lower in warmer climates) matches
with the epidemiology of migraine which shows higher prevalence in Europe, lower prevalence in
Africa, and intermediate prevalence in Asia (discussed in [96]). Better understanding of what the SNPs
at rs10166942 do to the expression/function of TRPM8 will likely provide much insight into how the
channel may be contributing to migraine, whether by greater or lesser activation during attacks.
4.4. TRPA1
The TRPA1 channel has probably received the most attention in preclinical headache/migraine
studies as it is activated by an extensive list of endogenous and exogenous stimuli that may have
relevance for migraine. These include reactive oxygen and nitrogen species, reactive prostaglandins,
and many environmental irritants such as chlorine, formaldehyde, cigarette smoke, and acrolein [51,55,
56,98]. For a recent review of TRPA1 and its role in numerous pain states, including migraine, see [58]
in this same Special Issue of Pharmaceuticals. Although the TRPA1 activator mustard oil had been used
as a probe to cause pain responses from the meninges in rodents for a number of years (see [99–101]),
investigation of this channel in migraine expanded following a couple of key studies. One was the
link between exposure to volatile oils from the Umbellularia californica tree (the “Headache Tree”) and
headache attacks [102] which was followed by later studies showing that umbellulone activates TRPA1Pharmaceuticals 2019, 12, 54 7 of 17
and activation of TRPA1 in the dura by umbellulone and mustard oil causes headache responses in
rats [103–105]. Umbellulone was also recently shown to facilitate propagation of CSD [106], so its
actions, and that of TRPA1 in general, may not be restricted to peripheral sensory neurons and the
meninges. This also led to several subsequent studies examining TRPA1-desensitizing compounds such
as parthenolide from the feverfew herb and isopetasin from butterbur for their ability to desensitize
meningeal nociceptors [107,108], potentially explaining the efficacy of these herbs in the treatment of
migraine. Both of these studies show that these natural-product modulators of TRPA1 lead to functional
desensitization of both the channel and the nerve ending expressing the channel. Consequences of
this desensitization are decreased responses of nociceptive nerve endings to subsequent stimulation,
which essentially acts as an inhibitor of neuronal function, and in the case of migraine, may lead to
decreased noxious input from the meninges. The results of these studies suggest that compounds
capable of desensitizing TRPA1 and TRPA1-expressing nerve endings may be viable new therapeutics
for migraine.
The other key study leading to increased interest in TRPA1 and migraine was the finding that
this channel mediates the vasodilatory response to inhalation of acrolein [109]. This was the first in a
line of publications by these authors documenting and interaction between environmental irritants
and TRPA1 that may lead to headaches following exposure. More recent work from these authors
has shown that repeated exposure to acrolein potentiates meningeal vasodilatory responses to TRPA1
and TRPV1 activation [110] and leads to migraine-like cutaneous hypersensitivity and increased
responses of the trigeminal afferent system to touch [111]. Further, lipidomic studies after acute or
chronic inhalation of acrolein in rats found increased levels of a number of factors known to activate
TRPV1 and TRPV4 [112]. These studies provide important mechanistic information for how repeated
inhalation of environmental irritants can lead to headache disorders and also propose TRPA1 (as well
as TRPV1 and TRPV4) antagonists as potential therapeutics in this area.
A number of studies over the last several years have been aimed at determining a role for TRPA1
in responses of the meningeal nociceptive system to other irritants such as hydrogen sulfide (H2 S) as
well as to nitric oxide (NO), both of which are well known to cause headaches. Importantly, these two
compounds may act together as H2 S and NO combine to form nitroxyl (HNO) that then reacts with
and activates TRPA1 by covalent modification [113]. This reaction contributes to modulation of
vascular tone downstream of H2 S and NO via the HNO-activated and TRPA1-mediated release of
CGRP. A recent study has demonstrated that this pathway contributes to activation of trigeminal
afferent signaling [114] and can increase meningeal blood flow following topical administration if
either NO or H2 S donors onto the dura via the production of HNO [115]. However, this mechanism is
far from fully explained as H2 S-NO-TRPA1 pathway can also decrease activity in neurons of the spinal
trigeminal nucleus, sometimes after prior activation [116]. Further, activation of TRPA1 alone (at least
using acrolein) may not be sufficient to initiate afferent signaling from the meninges but may require
co-activation of other channels such as TRPV1 [117]. Needless to say, these mechanisms are likely to
be complex.
NO donors have long been recognized as migraine triggers, provoking attacks in approximately
75% of patients within 6 hours of administration [118,119]. However, the mechanisms by which
this occurs remain poorly understood. Although one potential mechanism was described above
(via formation of HNO), many other possibilities exist downstream of NO including activation of
guanylyl cyclases, MAP kinases, nitrosylation of proteins, activation of cyclooxygenase enzymes,
and production of peroxynitrite radicals [120–125]. As discussed above and also shown here [126],
TRPA1 can mediate the nociceptive effects of NO, but given the diverse mechanisms downstream
of NO and the diffusion of NO throughout all cells, its actions are likely complex. This question
was extensively addressed recently using glyceryl trinitrate (GTN) administration to mice in the
presence of pharmacological and genetic tools against TRPA1, aldehyde dehydrogenase, reactive
oxygen and aldehyde species, and NADPH oxidase enzymes [127]. The authors examined both
cutaneous facial vasodilation and allodynia following GTN and found that the initial vasodilation isPharmaceuticals 2019, 12, 54 8 of 17
TRPA1-independent, but is mediated by aldehyde dehydrogenase (which liberates NO from GTN)
and a direct subsequent effect of NO on vessels. In contrast, the delayed allodynia following GTN was
entirely dependent on TRPA1 expression. Activation of TRPA1 likely occurs due to direct NO-mediated
interactions with the channel and also due to generation of reactive oxygen and carbonylic species
from increased activity of NADPH oxidase in the trigeminal ganglion. Consistent with this study was
another report showing that nitroglycerin can increase hyperalgesia in the orofacial formalin test in
rats, an effect that was blocked by a TRPA1 antagonist, along with an increase in channel expression in
the trigeminal ganglia, cervical spinal cord, and medulla [128]. These studies further implicates TRPA1
in migraine by directly demonstrating that it plays a role in headache-related behavior following
exposure of one of the most reliable triggers of attacks.
5. BoNTA, CGRP and TRP Channels
Interestingly, recent evidence points to the possible existence of interconnections between
the action of BoNTA, the release of CGRP and the TRP channels. In particular, in a rat
model of inflammation induced by complete Freund’s adjuvant (CFA), it was demonstrated that
pericranially-injected BoNTA is taken up by local sensory nerve endings, axonally transported to the
trigeminal ganglion and transcytosed to dural afferents, where BoNTA-cleaved SNAP-25 colocalized
with CGRP [129]. Importantly, BoNTA, similar to sumatriptan, reduced both the mechanical allodynia
and the dural neurogenic inflammation evoked by CFA and this is likely to happen by the suppression
of CGRP signalling [129]. Notably, it has been shown in migraine patients that CGRP levels after
BoNTA treatment are significantly lower in comparison to the CGRP levels obtained before BoNTA
treatment [130,131] and, more importantly, that the CGRP decrease is associated to the responsiveness
to treatment. In fact, in responder patients, pretreatment CGRP levels were significantly higher than in
nonresponders and, after treatment, the CGRP levels significantly decreased only in responders [130].
TRP channels, and especially TRPA1 and TRPV1, may also play a role in this scenario. It has been
demonstrated that BoNTA inhibits TRPV1 trafficking to the plasma membrane in primary trigeminal
ganglion neurons [132]. It has also been recently shown that the syntaxin 1-interacting protein,
Munc18-1, is necessary for TRPV1-triggered CGRP release and for the response to the proinflammatory
cytokine TNFα. Importantly, TNFα induces surface trafficking of TRPV1 and TRPA1 by means of
a synaptic vesicle membrane protein, VAMP1, which is also essential for the exocytosis of CGRP.
BoNTA inhibits the TNFα-elevated delivery and, consequently, abolishes the enhancement of Ca2+
influx through the upregulated surface-expressed TRPV1 and TRPA1 channels [133]. In addition, in a
rodent model, pretreatment with BoNTA significantly reduced both capsaicin- and allyl isothiocyanate
(AITC)-induced pain for at least 21 days, suggesting a durable pain-preventative effect of BoNTA
due to decreased responsiveness of TRPV1 and TRPA1 toward their selective agonists [134]. Finally,
BoNT-A administered to tissues outside the calvaria, is able to inhibit responses of C-type meningeal
nociceptors to stimulation of their intracranial dural receptive fields with capsaicin or mustard oil.
BoNTA effects on capsaicin responses are greater when the dose is injected along the suture lines,
instead of being divided between muscles and sutures [135], which is interesting from a translational
point of view given that BoNTA is injected in both locations in patients.
Although conclusive evidence is still needed, one may hypothesize that the abovementioned
mechanisms related to TRP channels contribute to the decreased release of CGRP associated to BoNTA;
initial evidence of the role of this circuitry has also been emerging for other drugs. Both valproic acid
and topiramate have been shown to inhibit the capsaicin-induced elevation of CGRP immunoreactivity
in the trigeminal ganglia and CGRP depletion in the dura mater of rats, even if with some differences
among adult and pediatric rats [136]. Finally, the pretreatment with valproic acid has been shown to
attenuate the enhanced blood flow responses observed after inhalation of the TRPA1 agonist acrolein
in a rodent model of chronic migraine [111]. Together, these studies support the concept that decreases
in TRP channel-mediated release of CGRP and other neuropeptides may contribute to the efficacy of
many pharmacological agents for migraine.Pharmaceuticals 2019, 12, 54 9 of 17
6. Concluding Remarks
There is currently a great deal of excitement in the migraine field due to the approval of several,
new CGRP-based therapeutics for use in humans. These novel agents add to the toolkit of the
headache specialist and offer new options with distinct mechanisms of action for patients. However,
it is already clear that these new therapeutics are not a panacea as many patients get suboptimal relief
from the monoclonal antibodies (approximately 35% showing less than 50% relief in an open-label
study of erenumab [137]; similar results have been observed in studies with other antibodies [138]).
While mechanisms determining what patients will or will not experience relief with these CGRP
therapies remain unclear, there is still great need for new therapeutics and TRP channels may represent
additional targets for these new drugs. Questions remain regarding the viability of TRP channels as
drug targets for migraine and these questions require additional work among the research community.
Although TRPV1 antagonists failed in a prior human trial for migraine, there is still a clear role for
TRPV1 in the activation of meningeal nociceptors and thus potential for alternate therapeutics to show
efficacy. Intranasal administration of the TRPV1 agonist Civamide was efficacious in humans, likely via
desensitization of nociceptors [139], providing proof of concept for TRPV1 modulating therapeutics,
in this case with an agonist. Little work currently exists surrounding TRPV4 and migraine and this is
the area that needs to most future exploration to evaluate this channel as a potential anti-migraine
drug target. The repeated identification of TRPM8 variants in GWAS supports continued interest in
this channel for the disorder, but it remains unknown whether therapeutics should be agonists or
antagonists. Better understanding of the impact of the channel mutations on expression/function
will help determine this answer. Finally, TRPA1 represents one of the most promising targets for new
therapeutics based on its potential role in headache following exposure to endogenous substances,
reactive oxygen species, environmental irritants, and migraine triggers such as NO donors as well
as its potential role in treating attacks when desensitized by natural products. As of this writing,
few TRPA1 antagonists have been developed for testing in humans, but there is now strong preclinical
rationale for testing of these agents, although no clinical trials are currently conducted to this aim
(www.clinicaltrials.gov accessed on February 17, 2019). Taken together, the data currently available
surrounding TRP channels and migraine continue to make a compelling case for further development
of new therapeutics based on modulation of these channels.
Funding: GD receives support from the National Institutes of Health (NS104200, NS104990, NS107058,NS098826)
and grant support from Alder Biopharmaceuticals and Teva Pharmaceuticals.
Conflicts of Interest: The authors declare no conflicts of interest in relation to this review.
Abbreviations
TRP Transient-receptor potential
CGRP Calcitonin gene-related peptide
BoNTA onabotulinum toxin A
IHS International Headache Society
ICHD International Classification of Headache Disorders
GBD Global Burden of Disease
NSAID Non-steroidal anti-inflammatory drugs
ACE Angiotensin-converting enzyme
GWAS Genome-wide association study
SNP Single nucleotide polymorphism
RAMP Receptor-activity modifying protein
RCP Receptor component protein
PAR-2 Protease-activated receptor type 2
H2 S Hydrogen sulfide
NO Nitric oxidePharmaceuticals 2019, 12, 54 10 of 17
HNO Nitroxyl
MAP Mitogen-activated protein
GTN Glyceryl trinitrate
NADPH Nicotinamide adenine dinucleotide phosphate
CFA Complete Freund’s adjuvant
SNAP-25 Synaptosomal-associated protein 25
Munc Mammalian uncoordinated
VAMP Vesicle-associated membrane protein
TNF Tumor necrosis factor
References
1. Dodick, D.W. Migraine. Lancet 2018, 391, 1315–1330. [PubMed]
2. Vetvik, K.G.; MacGregor, E.A. Sex differences in the epidemiology, clinical features, and pathophysiology of
migraine. Lancet Neurol. 2019, 16, 76–87.
3. Headache Classification Committee of the International Headache Society (IHS). The International
Classification of Headache Disorders, 3rd edition. Cephalalgia 2018, 38, 1–211.
4. Stovner, L.J.; Nichols, E.; Steiner, T.J.; Abd-Allah, F.; Abdelalim, A.; Al-Raddadi, R.M.; Ansha, M.G.; Barac, A.;
Bensenor, I.M.; Doan, L.P.; et al. Global, regional, and national burden of migraine and tension-type
headache, 1990-2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2018,
17, 954–976.
5. Freilinger, T.; Anttila, V.; de Vries, B.; Malik, R.; Kallela, M.; Terwindt, G.M.; Pozo-Rosich, P.; Winsvold, B.;
Nyholt, D.R.; van Oosterhout, W.P.; et al. Genome-wide association analysis identifies susceptibility loci for
migraine without aura. Nat. Genet. 2012, 44, 777–782. [CrossRef]
6. Blumenfeld, A.M.; Stark, R.J.; Freeman, M.C.; Orejudos, A.; Manack Adams, A. Long-term study of
the efficacy and safety of OnabotulinumtoxinA for the prevention of chronic migraine: COMPEL study.
J. Headache Pain 2018, 19, 13. [PubMed]
7. Dodick, D.W.; Turkel, C.C.; DeGryse, R.E.; Aurora, S.K.; Silberstein, S.D.; Lipton, R.B.; Diener, H.C.; Brin, M.F.
OnabotulinumtoxinA for treatment of chronic migraine: Pooled results from the double-blind, randomized,
placebo-controlled phases of the PREEMPT clinical program. Headache 2010, 50, 921–936. [PubMed]
8. Charles, A. The pathophysiology of migraine: Implications for clinical management. Lancet Neurol. 2018, 17,
174–182.
9. Borsook, D.; Maleki, N.; Becerra, L.; McEwen, B. Understanding migraine through the lens of maladaptive
stress responses: A model disease of allostatic load. Neuron 2012, 73, 219–234.
10. Noseda, R.; Burstein, R. Migraine pathophysiology: Anatomy of the trigeminovascular pathway and
associated neurological symptoms, CSD, sensitization and modulation of pain. Pain 2013, 154 (Suppl. 1),
S44–S53. [CrossRef]
11. Gormley, P.; Anttila, V.; Winsvold, B.S.; Palta, P.; Esko, T.; Pers, T.H.; Farh, K.H.; Cuenca-Leon, E.; Muona, M.;
Furlotte, N.A.; et al. Meta-analysis of 375,000 individuals identifies 38 susceptibility loci for migraine.
Nat. Genet. 2016, 48, 856–866. [CrossRef] [PubMed]
12. Edvinsson, L.; Haanes, K.A.; Warfvinge, K.; Krause, D.N. CGRP as the target of new migraine
therapies—Successful translation from bench to clinic. Nat. Rev. Neurol. 2018, 14, 338–350. [CrossRef]
[PubMed]
13. de Queiroz, B.F.; de Almeida, M.P.; Bakhle, Y.S.; Francischi, J.N. Calcitonin-gene related peptide is a potent
inducer of oedema in rat orofacial tissue. Neuropeptides 2018, 68, 43–48. [CrossRef]
14. Lukacs, M.; Haanes, K.A.; Majlath, Z.; Tajti, J.; Vecsei, L.; Warfvinge, K.; Edvinsson, L. Dural administration
of inflammatory soup or Complete Freund’s Adjuvant induces activation and inflammatory response in the
rat trigeminal ganglion. J. Headache Pain 2015, 16, 79. [CrossRef] [PubMed]
15. Russell, F.A.; King, R.; Smillie, S.J.; Kodji, X.; Brain, S.D. Calcitonin gene-related peptide: Physiology and
pathophysiology. Physiol. Rev. 2014, 94, 1099–1142. [CrossRef] [PubMed]
16. Mulderry, P.K.; Ghatei, M.A.; Bishop, A.E.; Allen, Y.S.; Polak, J.M.; Bloom, S.R. Distribution and
chromatographic characterisation of CGRP-like immunoreactivity in the brain and gut of the rat. Regul. Pept.
1985, 12, 133–143. [CrossRef]Pharmaceuticals 2019, 12, 54 11 of 17
17. Gibson, S.J.; Polak, J.M.; Bloom, S.R.; Sabate, I.M.; Mulderry, P.M.; Ghatei, M.A.; McGregor, G.P.; Morrison, J.F.;
Kelly, J.S.; Evans, R.M.; et al. Calcitonin gene-related peptide immunoreactivity in the spinal cord of man
and of eight other species. J. Neurosci. 1984, 4, 3101–3111. [CrossRef] [PubMed]
18. Eftekhari, S.; Salvatore, C.A.; Calamari, A.; Kane, S.A.; Tajti, J.; Edvinsson, L. Differential distribution of
calcitonin gene-related peptide and its receptor components in the human trigeminal ganglion. Neuroscience
2010, 169, 683–696. [CrossRef]
19. Gallai, V.; Sarchielli, P.; Floridi, A.; Franceschini, M.; Codini, M.; Glioti, G.; Trequattrini, A.; Palumbo, R.
Vasoactive peptide levels in the plasma of young migraine patients with and without aura assessed both
interictally and ictally. Cephalalgia 1995, 15, 384–390. [CrossRef]
20. Cernuda-Morollon, E.; Larrosa, D.; Ramon, C.; Vega, J.; Martinez-Camblor, P.; Pascual, J. Interictal increase
of CGRP levels in peripheral blood as a biomarker for chronic migraine. Neurology 2013, 81, 1191–1196.
[CrossRef]
21. Goadsby, P.J.; Edvinsson, L.; Ekman, R. Vasoactive peptide release in the extracerebral circulation of humans
during migraine headache. Ann. Neurol. 1990, 28, 183–187. [CrossRef]
22. Goadsby, P.J.; Edvinsson, L. The trigeminovascular system and migraine: Studies characterizing
cerebrovascular and neuropeptide changes seen in humans and cats. Ann. Neurol. 1993, 33, 48–56. [CrossRef]
23. Sarchielli, P.; Alberti, A.; Codini, M.; Floridi, A.; Gallai, V. Nitric oxide metabolites, prostaglandins
and trigeminal vasoactive peptides in internal jugular vein blood during spontaneous migraine attacks.
Cephalalgia 2000, 20, 907–918. [CrossRef]
24. Juhasz, G.; Zsombok, T.; Jakab, B.; Nemeth, J.; Szolcsanyi, J.; Bagdy, G. Sumatriptan causes parallel decrease
in plasma calcitonin gene-related peptide (CGRP) concentration and migraine headache during nitroglycerin
induced migraine attack. Cephalalgia 2005, 25, 179–183. [CrossRef]
25. Lassen, L.H.; Jacobsen, V.B.; Haderslev, P.A.; Sperling, B.; Iversen, H.K.; Olesen, J.; Tfelt-Hansen, P.
Involvement of calcitonin gene-related peptide in migraine: Regional cerebral blood flow and blood flow
velocity in migraine patients. J. Headache Pain 2008, 9, 151–157. [CrossRef]
26. Hansen, J.M.; Hauge, A.W.; Olesen, J.; Ashina, M. Calcitonin gene-related peptide triggers migraine-like
attacks in patients with migraine with aura. Cephalalgia 2010, 30, 1179–1186. [CrossRef]
27. Recober, A.; Kuburas, A.; Zhang, Z.; Wemmie, J.A.; Anderson, M.G.; Russo, A.F. Role of calcitonin
gene-related peptide in light-aversive behavior: Implications for migraine. J. Neurosci. 2009, 29, 8798–8804.
[CrossRef]
28. Olesen, J.; Diener, H.C.; Husstedt, I.W.; Goadsby, P.J.; Hall, D.; Meier, U.; Pollentier, S.; Lesko, L.M. Calcitonin
gene-related peptide receptor antagonist BIBN 4096 BS for the acute treatment of migraine. N. Engl. J. Med.
2004, 350, 1104–1110. [CrossRef]
29. Ho, T.W.; Ferrari, M.D.; Dodick, D.W.; Galet, V.; Kost, J.; Fan, X.; Leibensperger, H.; Froman, S.;
Assaid, C.; Lines, C.; et al. Efficacy and tolerability of MK-0974 (telcagepant), a new oral antagonist
of calcitonin gene-related peptide receptor, compared with zolmitriptan for acute migraine: A randomised,
placebo-controlled, parallel-treatment trial. Lancet 2008, 372, 2115–2123. [CrossRef]
30. Connor, K.M.; Shapiro, R.E.; Diener, H.C.; Lucas, S.; Kost, J.; Fan, X.; Fei, K.; Assaid, C.; Lines, C.; Ho, T.W.
Randomized, controlled trial of telcagepant for the acute treatment of migraine. Neurology 2009, 73, 970–977.
[CrossRef]
31. Voss, T.; Lipton, R.B.; Dodick, D.W.; Dupre, N.; Ge, J.Y.; Bachman, R.; Assaid, C.; Aurora, S.K.; Michelson, D.
A phase IIb randomized, double-blind, placebo-controlled trial of ubrogepant for the acute treatment of
migraine. Cephalalgia 2016, 36, 887–898. [CrossRef]
32. Hewitt, D.J.; Aurora, S.K.; Dodick, D.W.; Goadsby, P.J.; Ge, Y.J.; Bachman, R.; Taraborelli, D.; Fan, X.;
Assaid, C.; Lines, C.; et al. Randomized controlled trial of the CGRP receptor antagonist MK-3207 in the
acute treatment of migraine. Cephalalgia 2011, 31, 712–722. [CrossRef] [PubMed]
33. Marcus, R.; Goadsby, P.J.; Dodick, D.; Stock, D.; Manos, G.; Fischer, T.Z. BMS-927711 for the acute treatment of
migraine: A double-blind, randomized, placebo controlled, dose-ranging trial. Cephalalgia 2014, 34, 114–125.
[CrossRef]
34. Sun, H.; Dodick, D.W.; Silberstein, S.; Goadsby, P.J.; Reuter, U.; Ashina, M.; Saper, J.; Cady, R.; Chon, Y.;
Dietrich, J.; et al. Safety and efficacy of AMG 334 for prevention of episodic migraine: A randomised,
double-blind, placebo-controlled, phase 2 trial. Lancet Neurol. 2016, 15, 382–390. [CrossRef]Pharmaceuticals 2019, 12, 54 12 of 17
35. Tepper, S.; Ashina, M.; Reuter, U.; Brandes, J.L.; Dolezil, D.; Silberstein, S.; Winner, P.; Leonardi, D.; Mikol, D.;
Lenz, R. Safety and efficacy of erenumab for preventive treatment of chronic migraine: A randomised,
double-blind, placebo-controlled phase 2 trial. Lancet Neurol. 2017, 16, 425–434. [CrossRef]
36. Goadsby, P.J.; Reuter, U.; Hallstrom, Y.; Broessner, G.; Bonner, J.H.; Zhang, F.; Sapra, S.; Picard, H.; Mikol, D.D.;
Lenz, R.A. A Controlled Trial of Erenumab for Episodic Migraine. N. Engl. J. Med. 2017, 377, 2123–2132.
[CrossRef]
37. Dodick, D.W.; Goadsby, P.J.; Silberstein, S.D.; Lipton, R.B.; Olesen, J.; Ashina, M.; Wilks, K.; Kudrow, D.;
Kroll, R.; Kohrman, B.; et al. Safety and efficacy of ALD403, an antibody to calcitonin gene-related peptide, for
the prevention of frequent episodic migraine: A randomised, double-blind, placebo-controlled, exploratory
phase 2 trial. Lancet Neurol. 2014, 13, 1100–1107. [CrossRef]
38. Bigal, M.E.; Dodick, D.W.; Rapoport, A.M.; Silberstein, S.D.; Ma, Y.; Yang, R.; Loupe, P.S.; Burstein, R.;
Newman, L.C.; Lipton, R.B. Safety, tolerability, and efficacy of TEV-48125 for preventive treatment of
high-frequency episodic migraine: A multicentre, randomised, double-blind, placebo-controlled, phase 2b
study. Lancet Neurol. 2015, 14, 1081–1090. [CrossRef]
39. Bigal, M.E.; Dodick, D.W.; Krymchantowski, A.V.; VanderPluym, J.H.; Tepper, S.J.; Aycardi, E.; Loupe, P.S.;
Ma, Y.; Goadsby, P.J. TEV-48125 for the preventive treatment of chronic migraine: Efficacy at early time
points. Neurology 2016, 87, 41–48. [CrossRef]
40. Silberstein, S.D.; Dodick, D.W.; Bigal, M.E.; Yeung, P.P.; Goadsby, P.J.; Blankenbiller, T.; Grozinski-Wolff, M.;
Yang, R.; Ma, Y.; Aycardi, E. Fremanezumab for the Preventive Treatment of Chronic Migraine. N. Engl. J.
Med. 2017, 377, 2113–2122. [CrossRef] [PubMed]
41. Dodick, D.W.; Silberstein, S.D.; Bigal, M.E.; Yeung, P.P.; Goadsby, P.J.; Blankenbiller, T.; Grozinski-Wolff, M.;
Yang, R.; Ma, Y.; Aycardi, E. Effect of Fremanezumab Compared With Placebo for Prevention of Episodic
Migraine: A Randomized Clinical Trial. Jama 2018, 319, 1999–2008. [CrossRef]
42. Dodick, D.W.; Goadsby, P.J.; Spierings, E.L.; Scherer, J.C.; Sweeney, S.P.; Grayzel, D.S. Safety and efficacy
of LY2951742, a monoclonal antibody to calcitonin gene-related peptide, for the prevention of migraine:
A phase 2, randomised, double-blind, placebo-controlled study. Lancet Neurol. 2014, 13, 885–892. [CrossRef]
43. Skljarevski, V.; Oakes, T.M.; Zhang, Q.; Ferguson, M.B.; Martinez, J.; Camporeale, A.; Johnson, K.W.; Shan, Q.;
Carter, J.; Schacht, A.; et al. Effect of Different Doses of Galcanezumab vs Placebo for Episodic Migraine
Prevention: A Randomized Clinical Trial. JAMA Neurol. 2018, 75, 187–193. [CrossRef]
44. Stauffer, V.L.; Dodick, D.W.; Zhang, Q.; Carter, J.N.; Ailani, J.; Conley, R.R. Evaluation of Galcanezumab for
the Prevention of Episodic Migraine: The EVOLVE-1 Randomized Clinical Trial. JAMA Neurol. 2018, 75,
1080–1088. [CrossRef]
45. Skljarevski, V.; Matharu, M.; Millen, B.A.; Ossipov, M.H.; Kim, B.K.; Yang, J.Y. Efficacy and safety of
galcanezumab for the prevention of episodic migraine: Results of the EVOLVE-2 Phase 3 randomized
controlled clinical trial. Cephalalgia 2018, 38, 1442–1454. [CrossRef]
46. Bernstein, C.; Burstein, R. Sensitization of the trigeminovascular pathway: Perspective and implications to
migraine pathophysiology. J. Cl.in Neurol. 2012, 8, 89–99. [CrossRef]
47. Khan, S.; Mohammad Amin, F.; Emil Christensen, C.; Ghanizada, H.; Younis, S.; Olinger, A.C.R.;
de Koning, P.J.H.; Larsson, H.B.W.; Ashina, M. Meningeal contribution to migraine pain: A magnetic
resonance angiography study. Brain 2019, 142, 93–102. [CrossRef]
48. Vriens, J.; Appendino, G.; Nilius, B. Pharmacology of vanilloid transient receptor potential cation channels.
Mol. Pharmacol. 2009, 75, 1262–1279. [CrossRef]
49. Julius, D. TRP channels and pain. Annu. Rev. Cell Dev. Biol. 2013, 29, 355–384. [CrossRef]
50. Jardin, I.; Lopez, J.J.; Diez, R.; Sanchez-Collado, J.; Cantonero, C.; Albarran, L.; Woodard, G.E.; Redondo, P.C.;
Salido, G.M.; Smani, T.; et al. TRPs in Pain Sensation. Front. Physiol. 2017, 8, 392. [CrossRef]
51. Dussor, G.; Yan, J.; Xie, J.Y.; Ossipov, M.H.; Dodick, D.W.; Porreca, F. Targeting TRP channels for novel
migraine therapeutics. ACS Chem. Neurosci. 2014, 5, 1085–1096. [CrossRef]
52. Veldhuis, N.A.; Poole, D.P.; Grace, M.; McIntyre, P.; Bunnett, N.W. The G protein-coupled receptor-transient
receptor potential channel axis: Molecular insights for targeting disorders of sensation and inflammation.
Pharmacol. Rev. 2015, 67, 36–73. [CrossRef]
53. Dussor, G.; Cao, Y.Q. TRPM8 and Migraine. Headache 2016, 56, 1406–1417. [CrossRef]
54. Geppetti, P.; Benemei, S.; De Cesaris, F. CGRP receptors and TRP channels in migraine. J. Headache Pain 2015,
16, A21. [CrossRef]Pharmaceuticals 2019, 12, 54 13 of 17
55. Benemei, S.; De Cesaris, F.; Fusi, C.; Rossi, E.; Lupi, C.; Geppetti, P. TRPA1 and other TRP channels in
migraine. J. Headache Pain 2013, 14, 71. [CrossRef]
56. Benemei, S.; Fusi, C.; Trevisan, G.; Geppetti, P. The TRPA1 channel in migraine mechanism and treatment.
Br. J. Pharmacol. 2014, 171, 2552–2567. [CrossRef]
57. Dux, M.; Santha, P.; Jancso, G. The role of chemosensitive afferent nerves and TRP ion channels in the
pathomechanism of headaches. Pflug. Arch. 2012, 464, 239–248. [CrossRef]
58. Koivisto, A.; Jalava, N.; Bratty, R.; Pertovaara, A. TRPA1 Antagonists for Pain Relief. Pharmaceuticals 2018,
11, 117. [CrossRef]
59. Moskowitz, M.A.; Brody, M.; Liu-Chen, L.Y. In vitro release of immunoreactive substance P from putative
afferent nerve endings in bovine pia arachnoid. Neuroscience 1983, 9, 809–814. [CrossRef]
60. Shimizu, T.; Toriumi, H.; Sato, H.; Shibata, M.; Nagata, E.; Gotoh, K.; Suzuki, N. Distribution and origin of
TRPV1 receptor-containing nerve fibers in the dura mater of rat. Brain Res. 2007, 1173, 84–91. [CrossRef]
61. Del Fiacco, M.; Quartu, M.; Boi, M.; Serra, M.P.; Melis, T.; Boccaletti, R.; Shevel, E.; Cianchetti, C. TRPV1,
CGRP and SP in scalp arteries of patients suffering from chronic migraine. J. Neurol. Neurosurg. Psychiatry
2015, 86, 393–397. [CrossRef]
62. Buonvicino, D.; Urru, M.; Muzzi, M.; Ranieri, G.; Luceri, C.; Oteri, C.; Lapucci, A.; Chiarugi, A. Trigeminal
ganglion transcriptome analysis in 2 rat models of medication-overuse headache reveals coherent and
widespread induction of pronociceptive gene expression patterns. Pain 2018, 159, 1980–1988. [CrossRef]
63. Marics, B.; Peitl, B.; Pazmandi, K.; Bacsi, A.; Nemeth, J.; Oszlacs, O.; Jancso, G.; Dux, M. Diet-Induced
Obesity Enhances TRPV1-Mediated Neurovascular Reactions in the Dura Mater. Headache 2017, 57, 441–454.
[CrossRef]
64. Rossi, H.L.; Broadhurst, K.A.; Luu, A.S.; Lara, O.; Kothari, S.D.; Mohapatra, D.P.; Recober, A. Abnormal
trigeminal sensory processing in obese mice. Pain 2016, 157, 235–246. [CrossRef]
65. Marics, B.; Peitl, B.; Varga, A.; Pazmandi, K.; Bacsi, A.; Nemeth, J.; Szilvassy, Z.; Jancso, G.; Dux, M.
Diet-induced obesity alters dural CGRP release and potentiates TRPA1-mediated trigeminovascular
responses. Cephalalgia 2017, 37, 581–591. [CrossRef]
66. Ibrahimi, K.; Vermeersch, S.; Danser, A.; Villalon, C.M.; van den Meiracker, A.H.; de Hoon, J.;
MaassenVanDenBrink, A. Development of an experimental model to study trigeminal nerve-mediated
vasodilation on the human forehead. Cephalalgia 2014, 34, 514–522. [CrossRef]
67. Ibrahimi, K.; Vermeersch, S.; Frederiks, P.; Geldhof, V.; Draulans, C.; Buntinx, L.; Lesaffre, E.;
MaassenVanDenBrink, A.; de Hoon, J. The influence of migraine and female hormones on capsaicin-induced
dermal blood flow. Cephalalgia 2017, 37, 1164–1172. [CrossRef]
68. Kamshilin, A.A.; Volynsky, M.A.; Khayrutdinova, O.; Nurkhametova, D.; Babayan, L.; Amelin, A.V.;
Mamontov, O.V.; Giniatullin, R. Novel capsaicin-induced parameters of microcirculation in migraine patients
revealed by imaging photoplethysmography. J. Headache Pain 2018, 19, 43. [CrossRef]
69. Zhang, X.; Levy, D.; Noseda, R.; Kainz, V.; Jakubowski, M.; Burstein, R. Activation of meningeal nociceptors
by cortical spreading depression: Implications for migraine with aura. J. Neurosci. 2010, 30, 8807–8814.
[CrossRef]
70. Zhang, X.C.; Levy, D.; Kainz, V.; Noseda, R.; Jakubowski, M.; Burstein, R. Activation of Central
Trigeminovascular Neurons by Cortical Spreading Depression. Ann. Neurol. 2011, 69, 855–865. [CrossRef]
71. Schain, A.J.; Melo-Carrillo, A.; Borsook, D.; Grutzendler, J.; Strassman, A.M.; Burstein, R. Activation of pial
and dural macrophages and dendritic cells by cortical spreading depression. Ann. Neurol. 2018, 83, 508–521.
[CrossRef]
72. Karatas, H.; Erdener, S.E.; Gursoy-Ozdemir, Y.; Lule, S.; Eren-Kocak, E.; Sen, Z.D.; Dalkara, T. Spreading
Depression Triggers Headache by Activating Neuronal Panx1 Channels. Science 2013, 339, 1092–1095.
[CrossRef]
73. Levy, D. Endogenous mechanisms underlying the activation and sensitization of meningeal nociceptors:
The role of immuno-vascular interactions and cortical spreading depression. Curr. Pain Headache Rep. 2012,
16, 270–277. [CrossRef]
74. Palmer, C.B.; Lai, R.; Guillard, F.; Bullman, J.; Baines, A.; Napolitano, A.; Appleby, J. A randomised,
two-period cross-over study to investigate the efficacy of the TRPV1 antagonist SB-705498 in acute migraine.
In Proceedings of the yje Pain in Europe VI, Lisbon, Portugal, 9–12 September 2009; p. 702.Pharmaceuticals 2019, 12, 54 14 of 17
75. Liedtke, W.; Choe, Y.; Marti-Renom, M.A.; Bell, A.M.; Denis, C.S.; Sali, A.; Hudspeth, A.J.; Friedman, J.M.;
Heller, S. Vanilloid receptor-related osmotically activated channel (VR-OAC), a candidate vertebrate
osmoreceptor. Cell 2000, 103, 525–535. [CrossRef]
76. Liedtke, W.; Tobin, D.M.; Bargmann, C.I.; Friedman, J.M. Mammalian TRPV4 (VR-OAC) directs behavioral
responses to osmotic and mechanical stimuli in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2003,
100 (Suppl. 2), 14531–14536. [CrossRef]
77. Vriens, J.; Watanabe, H.; Janssens, A.; Droogmans, G.; Voets, T.; Nilius, B. Cell swelling, heat, and chemical
agonists use distinct pathways for the activation of the cation channel TRPV4. Proc. Natl. Acad. Sci. USA
2004, 101, 396–401. [CrossRef]
78. Burstein, R.; Cutrer, M.F.; Yarnitsky, D. The development of cutaneous allodynia during a migraine attack
clinical evidence for the sequential recruitment of spinal and supraspinal nociceptive neurons in migraine.
Brain J. Neurol. 2000, 123 Pt 8, 1703–1709. [CrossRef]
79. Moran, M.M. TRP Channels as Potential Drug Targets. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 309–330.
[CrossRef]
80. Wei, X.; Edelmayer, R.M.; Yan, J.; Dussor, G. Activation of TRPV4 on dural afferents produces
headache-related behavior in a preclinical rat model. Cephalalgia 2011, 31, 1595–1600. [CrossRef]
81. Cenac, N.; Altier, C.; Chapman, K.; Liedtke, W.; Zamponi, G.; Vergnolle, N. Transient receptor potential
vanilloid-4 has a major role in visceral hypersensitivity symptoms. Gastroenterology 2008, 135, 937–946.
[CrossRef]
82. Grant, A.D.; Cottrell, G.S.; Amadesi, S.; Trevisani, M.; Nicoletti, P.; Materazzi, S.; Altier, C.; Cenac, N.;
Zamponi, G.W.; Bautista-Cruz, F.; et al. Protease-activated receptor 2 sensitizes the transient receptor
potential vanilloid 4 ion channel to cause mechanical hyperalgesia in mice. J. Physiol. 2007, 578, 715–733.
[CrossRef]
83. Sipe, W.E.; Brierley, S.M.; Martin, C.M.; Phillis, B.D.; Cruz, F.B.; Grady, E.F.; Liedtke, W.; Cohen, D.M.;
Vanner, S.; Blackshaw, L.A.; et al. Transient receptor potential vanilloid 4 mediates protease activated receptor
2-induced sensitization of colonic afferent nerves and visceral hyperalgesia. Am. J. Physiol. Gastrointest. Liver
Physiol. 2008, 294, G1288–G1298. [CrossRef]
84. Zhang, X.C.; Levy, D. Modulation of meningeal nociceptors mechanosensitivity by peripheral
proteinase-activated receptor-2: The role of mast cells. Cephalalgia 2008, 28, 276–284. [CrossRef]
85. Hassler, S.N.; Ahmad, F.B.; Burgos-Vega, C.C.; Boitano, S.; Vagner, J.; Price, T.J.; Dussor, G. Protease activated
receptor 2 (PAR2) activation causes migraine-like pain behaviors in mice. Cephalalgia 2018, 39, 111–122.
[CrossRef]
86. Moore, C.; Gupta, R.; Jordt, S.E.; Chen, Y.; Liedtke, W.B. Regulation of Pain and Itch by TRP Channels.
Neurosci. Bull. 2018, 34, 120–142. [CrossRef]
87. Weyer, A.D.; Lehto, S.G. Development of TRPM8 Antagonists to Treat Chronic Pain and Migraine.
Pharmaceuticals 2017, 10, 37. [CrossRef]
88. Chen, S.P.; Fuh, J.L.; Chung, M.Y.; Lin, Y.C.; Liao, Y.C.; Wang, Y.F.; Hsu, C.L.; Yang, U.C.; Lin, M.W.; Chiou, J.J.;
et al. Genome-wide association study identifies novel susceptibility loci for migraine in Han Chinese resided
in Taiwan. Cephalalgia 2018, 38, 466–475. [CrossRef]
89. Ren, L.; Dhaka, A.; Cao, Y.Q. Function and postnatal changes of dural afferent fibers expressing TRPM8
channels. Mol. Pain 2015, 11, 37. [CrossRef]
90. Ren, L.; Chang, M.J.; Zhang, Z.; Dhaka, A.; Guo, Z.; Cao, Y.Q. Quantitative Analysis of Mouse Dural Afferent
Neurons Expressing TRPM8, VGLUT3, and NF200. Headache 2018, 58, 88–101. [CrossRef]
91. Burgos-Vega, C.C.; Ahn, D.D.; Bischoff, C.; Wang, W.; Horne, D.; Wang, J.; Gavva, N.; Dussor, G. Meningeal
transient receptor potential channel M8 activation causes cutaneous facial and hindpaw allodynia in a
preclinical rodent model of headache. Cephalalgia 2016, 36, 185–193. [CrossRef]
92. Kayama, Y.; Shibata, M.; Takizawa, T.; Ibata, K.; Shimizu, T.; Ebine, T.; Toriumi, H.; Yuzaki, M.; Suzuki, N.
Functional interactions between transient receptor potential M8 and transient receptor potential V1 in the
trigeminal system: Relevance to migraine pathophysiology. Cephalalgia 2018, 38, 833–845. [CrossRef]
93. Kobayashi, K.; Fukuoka, T.; Obata, K.; Yamanaka, H.; Dai, Y.; Tokunaga, A.; Noguchi, K. Distinct expression
of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with aδ/c-fibers and colocalization
with trk receptors. J. Comp. Neurol. 2005, 493, 596–606. [CrossRef]You can also read