Transcriptomic response and immunological responses to chimpanzee adenovirus- and MVA viral-vectored vaccines for RSV in healthy adults - Oxford ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Clinical and Experimental Immunology, 2023, 211, 269–279
https://doi.org/10.1093/cei/uxad003
Advance access publication 9 January 2023
Research Article
Research Article
Transcriptomic response and immunological responses to
chimpanzee adenovirus- and MVA viral-vectored vaccines
for RSV in healthy adults
Downloaded from https://academic.oup.com/cei/article/211/3/269/6979728 by guest on 07 September 2023
C. Green1,2,‡, J. McGinley1,*,‡, , C. Sande1, S. Capone3, S. Makvandi-Nejad5, A. Vitelli3,
L. Silva-Reyes1, S. Bibi1, C. Otasowie1, D. Sheerin1, A. Thompson1, C. Dold1, P. Klenerman4,
E. Barnes4, L. Dorrell5, C. Rollier1, A. Pollard1,§, D. O’Connor1,§
1
Oxford Vaccine Group, Department of Paediatrics and the NIHR Oxford Biomedical Research Centre, University of Oxford, Oxford, UK
2
Institute of Microbiology & Infection, University of Birmingham, Birmingham, UK
3
Experimental Vaccinology Department, ReiThera Srl, Roma, Italy
4
Experimental Medicine Division, Nuffield Department of Medicine, University of Oxford, Oxford, UK
5
Nuffield Department of Medicine, University of Oxford NDM Research Building, Oxford, UK
‡First authors with equal contribution.
§ Joint senior authors.
*
Correspondence: Joseph McGinley, Oxford Vaccine Group, Centre for Clinical Vaccinology & Tropical Medicine, Churchill Hospital, Oxford, UK.
Email: joseph.mcginley@paediatrics.ox.uk
European Medicines Agency’s European Clinical Trials Database (EudraCT; reference 2011-003589-34)
Abstract
Cohorts of healthy younger adults (18–50yrs) and healthy older adults (60–75yrs) were immunized intramuscularly or intranasally with an
adenovirus-vectored RSV vaccine (PanAd3-RSV) as a prime dose and boosted with PanAd3-RSV or a poxvirus-vectored vaccine (MVA-RSV)
encoding the same insert. Whole blood gene expression was measured at baseline, 3- and 7-days post vaccination. Intramuscular prime vaccin-
ation with PanAd3-RSV induced differential expression of 643 genes (DEGs, FDR < 0.05). Intranasal prime vaccination with PanAd3-RSV did not
induce any differentially expressed genes (DEGs) in blood samples at 3 days post vaccination. Intranasally primed participants showed greater
numbers of DEGS on boosting than intramuscularly primed participants. The most highly enriched biological processes related to DEGs after
both prime and boost vaccination were type-1 interferon related pathways, lymphocytic and humoral immune responses.
Received 3 May 2022; Revised 30 October 2022; Accepted for publication 6 January 2023
© The Author(s) 2023. Published by Oxford University Press on behalf of the British Society for Immunology.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/),
which permits unrestricted reuse, distribution, and reproduction in any medium, provided the original work is properly cited.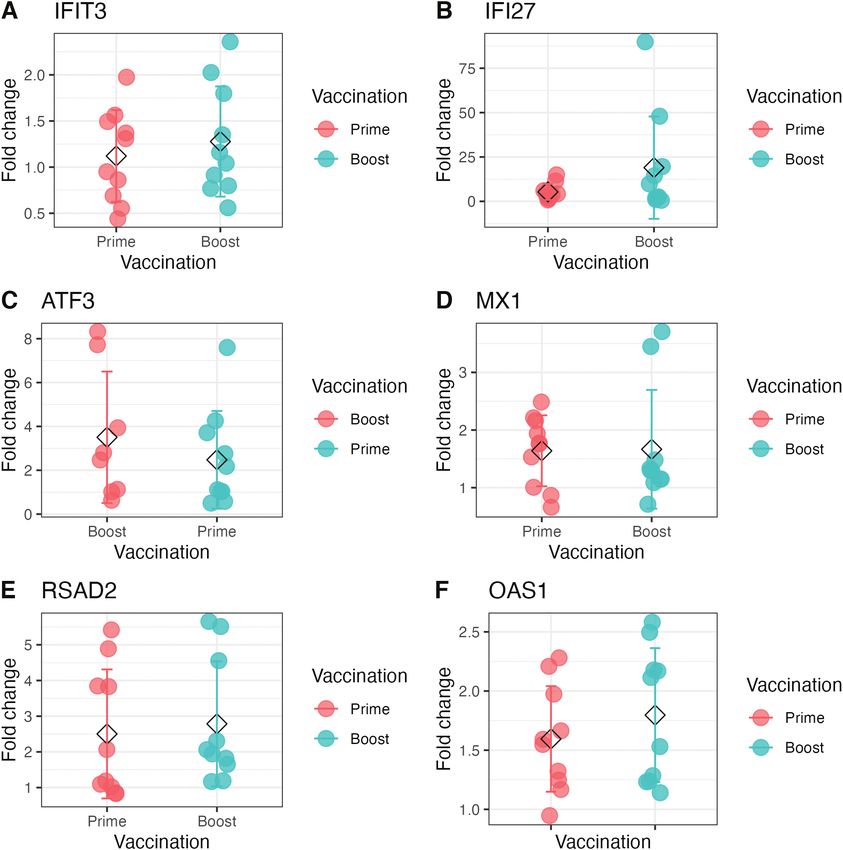
270 Green et al.
Graphical Abstract
Shared differentially expressed genes between PanAd3 Prime and MVA boost treatment
550 genes up in both and 308 genes down in both (N = 955 genes total)
OTOF ISG15 IFIT3
2 IFI44
OAS3 IFI6 EPSTI1 C1QB
USP18 MX1 HERC5 OAS1
CMPK2 HES4
LY6E
MS4A4A
1 XAF1
CDKN1C
Downloaded from https://academic.oup.com/cei/article/211/3/269/6979728 by guest on 07 September 2023
log2 PanAd3 prime
0
−1 ALPL
LOC642342
CHI3L1
−2 ORM1
−2 −1 0 1 2
log2 MVA boost
Keywords: RSV; vaccine; transcriptomics; human trials; antibodies; viral vectors
Abbreviations: DEGs: differentially expressed genes; GO: gene ontology; GSEA: gene set enrichment analysis; IFN: interferon; IM: intramuscular; IN: intranasal;
MVA: modified vaccinia ankara; PanAd: Chimpanzee adenovirus; RSV: respiratory syncytial virus.
Introduction in recent years through the success of INN- Ad26.COV2-S
Viral vectors that deliver gene products from unrelated or- and Vaxzevria vaccines in recent years. Understanding the
ganisms are one of the leading platforms for the development underlying biological pathways that lead to the generation
of novel vaccines [1] and are capable of stimulating both of desirable immunity could profoundly inform the design
antigen-specific antibody and cellular immune responses [2]. of optimal vectors, inserts, or the development of entirely
This provides the potential for control of vaccine-preventable novel technologies [15], particularly in light of the ongoing
diseases of global importance and outbreak pathogens that COVID-19 pandemic, where at the time of writing 13 of the
threaten health security. Significant progress has been made in 44 candidate vaccines currently in trials use viral vectors of
development and testing of viral vectored vaccines for diseases some sort. Interrogation of early pathways induced by vaccin-
such as malaria, tuberculosis, HIV, influenza, rabies, hepatitis ation through analysis of gene expression, and linking early
C, and group B Meningococcus [3–10], and the flexibility of response patterns to immunological endpoints has led to the
their design means that this approach can be used to rap- successful identification of key determinants of vaccine im-
idly develop new vaccines to control mycobacterial and viral munogenicity for several viral and subunit vaccines [16].
epidemics, as recently demonstrated for Ebola in West Africa Analysis is reported here of the early host transcriptomic
[11–13], and now in response to the COVID-19 pandemic responses to two novel respiratory syncytial virus (RSV)
[2, 14]. Several viral vectors have been tested in humans, viral-vectored vaccines in humans (PanAd3-RSV and
including Rhabdovirus (vesicular stomatitis virus, modified MVA-RSV) encoding the same three antigens (F0ΔTM, N,
vaccinia virus Ankara), alphavirus, human adenovirus (Ad5), and M2-1 proteins). In this study, participants were given
and chimpanzee adenovirus (ChAd3, ChAd63, ChAdOx1, PanAd3-RSV either by intramuscular or intra-nasal prime
ChAd155, PanAd3, ad6, ad25, ad35, GAd20, and GrAd32), followed by homologous or heterologous boost, allowing in-
and these biological platforms have been shown to have sight into the host response to the same vaccine given by a
good safety profiles in trials thus far and are immunogenic different route, and to the same antigen delivered by a dif-
[2]. Efficacy of these platforms has been well characterized ferent viral vector. The aims of this study were to identifyClinical and Experimental Immunology, 2023, Vol. 211, No. 3 271 a transcriptomic signature in response to vaccination, and the paired sampling and arrays were weighted by quality demonstrate which elements of this signature are linked to score [20, 21]. A log2 fold change of 0.32 (absolute fold generation of desirable neutralizing antibody responses. The change ≥1.25) from pre-vaccination baseline combined with safety and immunogenicity of this vaccine have been previ- a 5% false discovery rate (FDR) adjusted P-value of
272 Green et al.
Downloaded from https://academic.oup.com/cei/article/211/3/269/6979728 by guest on 07 September 2023
Figure 1. Volcano plot illustrations of early transcriptional responses to RSV vaccination. A: Transcriptional response of N = 21 volunteers who received
PanAd3-RSV prime vaccination by IM injection measured 3-days after vaccination. B: Transcriptional response of N = 21 volunteers who received
PanAd3-RSV prime vaccination by IM injection measured 7 days after vaccination. C: Transcriptional response of N = 11 volunteers who received MVA-
RSV boost vaccination by IM injection after being primed with IM PanAd3-RSV measured 3 days after boost vaccination. D: Transcriptional response of
N = 10 volunteers who received MVA-RSV boost vaccination by IM injection after IN PanAd3 prime vaccination measured 3-days after boost vaccination.
E: Transcriptional response N = 10 volunteers who received MVA-RSV boost vaccination by IM injection after IN priming with PanAd3-RSV measured 7
days after boost vaccination. Red dots denote significantly differentially expressed probes (FDR < 0.05, log2 fold change > 0.26), blue dots significantly
differentially expressed probes with fold change < 1.2, green dots indicate genes with a log2 fold change > 0.26 but which are not significantly
differentially expressed. Horizontal dotted lines represent FDR cutoffs, vertical dotted lines represent log2 fold change cutoffs. (F) Agreement plot of
differentially expressed probes 3 days after IM PanAd3-RSV prime or IM MVA-RSV boost vaccination (IN primed volunteers). Plot of the log2 adjusted
fold changes of all shared differentially expressed genes 3 days after intramuscular priming with PanAd3-RSV and intramuscular boosting with MVA-RSV
in participants primed intramuscularly. Points coloured in purple represent genes that are downregulated post both vaccinations. Blue points are genes
that are upregulated after both vaccinations. No differentially expressed genes were downregulated in response to one vaccination and upregulated in
response to another.
7 days following IN PanAd3-RSV prime vaccination (groups molecular function terms (Supplementary Fig. S2A–D), and
three and four combined). immunological terms (Supplementary Fig. S3A-D). Terms that
In order to further interpret the perturbation of the whole were significantly enriched (FDR < 0.05) were represented
blood transcriptome following prime and boost vaccination, and ranked according to the enrichment score of the term.
a gene set enrichment analysis was performed on all genes The most enriched gene sets 3 days post prime vaccination
on the array ranked by their t-statistic using databases for (Fig. 2A) were type I interferon response genes, with add-
gene ontology (GO) biological process terms (Fig 2A–D), GO itional general anti-viral responses, alongside B cell responses.Clinical and Experimental Immunology, 2023, Vol. 211, No. 3 273
Downloaded from https://academic.oup.com/cei/article/211/3/269/6979728 by guest on 07 September 2023
Figure 2. Gene set enrichment analysis of biological processes of the response to prime vaccination three and seven days after vaccination. Gene
enrichment analysis for terms relating to biological processes was performed on the ranked list of all probes on the array at 3 (A) and 7 days (B) post
prime vaccination and 3 (C) and 7 days (D) post boost vaccination by MVA-RSV in participants primed intranasally. Most significantly enriched terms are
ranked by enrichment score. Dots are coloured based on the FDR of the associated term. Size of dots is based on the number of genes associated with
that term. FDR274 Green et al.
Table 2. Differentially expressed probes 3 and 7 days after IM PanAd3-RSV prime vaccination with significant statistical correlation to clinical/
immunological responses measured at later time points
Vaccination Number of genes Number of genes Number of genes Number of genes Number of genes
correlated with correlated with correlated with VnAb correlated with ASC-G correlated with ASC-A
PRNT 28 days post IFNγ 14 days post 7 days post vaccination 7 days post vaccination 7 days post vaccination
vaccination vaccination
3 Days post PanAd3 43 15 4 23 20
priming
3 days post MVA 6 0 NA 0 0
boost (IM PanAd3
primed)
3 days post MVA 32 20 NA 9 6
Downloaded from https://academic.oup.com/cei/article/211/3/269/6979728 by guest on 07 September 2023
boost (IN PanAd3
primed)
7 Days post PanAd3 42 11 85 73 44
prime
7 days post MVA 15 57 18 94 23
boost (IM PanAd3
primed)
7 days post MVA 12 53 51 19 31
boost (IN PanAd3
primed)
Data from volunteers in study groups one, two, three, and four combined (n = 42). The significant correlations (Pearsons P-value 0.05) between changes in
gene expression from the 634 of differentially expressed probes 3 days after IM PanAd3-RSV prime and the 782 and 94 probes, respectively differentially
expressed after boost vaccination with MVA-RSV after either intramuscular or intranasal priming and log2-transformed fold-change in serum anti-PanAd3
vector neutralising antibody 4-weeks after vaccination (VnAb), log2-transformed fold-change in serum RSV-neutralising antibody titre 28 days after
vaccination (PRNT), log2-transformed anti-F IgG and IgA antibody secreting cell count 7 days after vaccination (ASC-G, ASC-A), and the log2-transformed
fold-change in IFNγ count 14 days after prime.
data have been previously published [18]. Forty-three genes RT-qPCR was performed in this group to analyse the ex-
differentially expressed 3 days post IM prime correlated (either pression of genes found to be of interest based on their re-
positively or negatively) with the fold changes in neutralizing lationship with immunological outcomes in younger adults.
antibody 28 days post prime vaccination (Table 2), 15 genes Six genes were selected for differential expression analysis in
with the fold change expansion of F-specific IFNγ producing T older adults. These genes were chosen as they had the most
cells 14 days post prime vaccination (Table 2), four genes cor- significant correlations with neutralizing antibody responses
related with PanAd3 (vector)-specific serum neutralizing anti- in older adults from transcriptomic data. Of the genes selected
body titres 28 days post vaccination (Table 2), and 23 and 20 for differential expression analysis by RT-qPCR, IFI27, MX1,
genes correlated with the number of antibody secreting cells OAS1, and RSAD2 were found to be differentially expressed,
producing IgG and IgA in response to RSV F protein 7 days while IFIT3 and ATF3 were not found to be expressed in
post vaccination, respectively (Table 2). Two hundred fifty-five older adults (fig. 3A–F). After boost vaccination with MVA-
were also shown to have correlations with immunological out- RSV, ATF3, IFI27, MX1, OAS1, and RSAD2 were found to
come measures seven days post vaccination, despite the lack of be differentially expressed, while IFIT3 was not found to be
differential expression at this time point (Table 2). significantly differentially expressed, but none of these genes
Participants who had been primed intramuscularly with correlated with changes in neutralising antibody in older
PanAd3 and then boosted with MVA elicited similar re- adults (Fig. 4A–F). Gene expression analysis by RT-qPCR was
sponses, with 33 genes differentially expressed 3 days post not possible in younger adult samples as these had been fully
boost that correlated with neutralising antibody 28 days depleted before commencement of RT-qPCR experiments.
post boost vaccination(Table 2), 20 genes were correlated
with the magnitude of F-specific IFNγ producing T-cells 7
days post boost (Table 2), and nine and six genes correlated Discussion
with the number of antibody-secreting cells producing IgG In this whole-blood gene expression analysis, we found a rich
and IgA at seven days post vaccination in response to RSV repertoire of 634 differentially expressed genes 3 days after
F protein, respectively (Table 2). Boosting with IM PanAd3 intramuscular prime vaccination with PanAd3-RSV, and 94
after intranasal vaccination with IN PanAd3 RSV induced no differentially expressed genes 3 days after subsequent boost
significant differential expression (not shown). vaccination with MVA-RSV. Intranasal prime vaccination
with PanAd3-RSV did not induce differential gene expression
Early gene regulation replicated in older adults but in blood, but did induce differential expression after boosting.
did not correlate with vaccine responses Seven hundred and eighty-two genes differentially expressed
A further cohort of older adults was vaccinated with PanAd3 genes were detected 3 days after MVA-RSV boost, suggesting
RSV and MVA RSV (Table 1). Gene expression analysis with that either there was an effect from the intra-nasal routeClinical and Experimental Immunology, 2023, Vol. 211, No. 3 275
Downloaded from https://academic.oup.com/cei/article/211/3/269/6979728 by guest on 07 September 2023
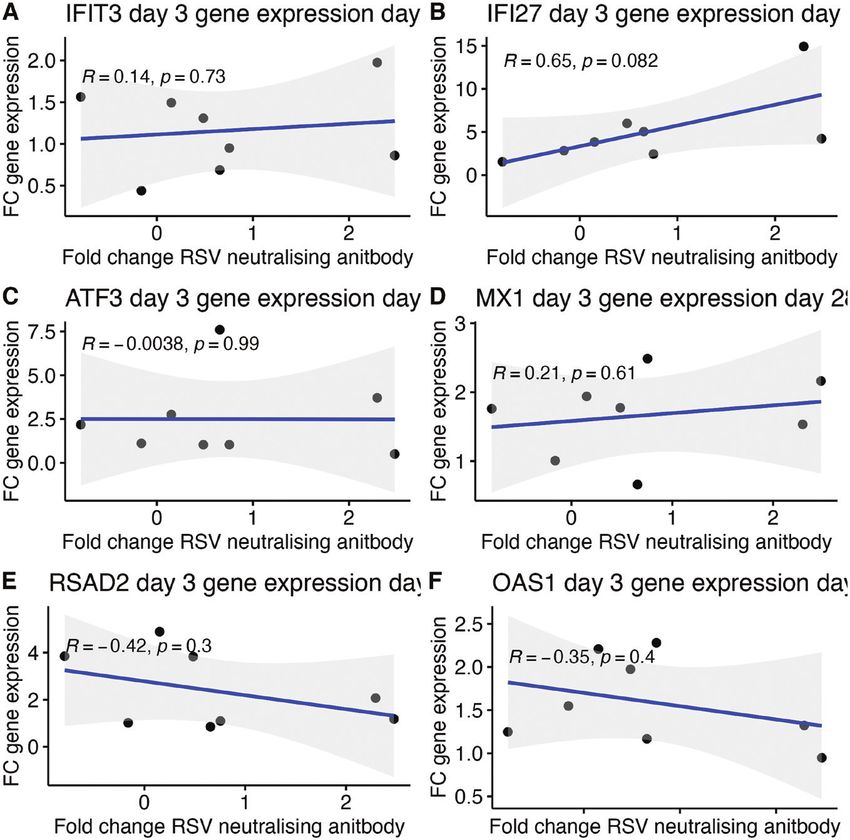
Figure 3. (A–F) IFI27, MX1, OAS1, and RSAD2 are significantly differentially expressed in older adult grouping after prime vaccination with PanAd Titres
of genes determined to be relevant to the generation of immunological outcomes such as neutralising antibody response were measured in older
adults by qPCR. Results are presented as ddCT between day of vaccination and 28 days post vaccination. Asterisk indicates differential expression of
that gene compared with baseline levels (P < 0.05), genes after prime vaccination are coloured in red while genes after boost are coloured in blue.
of priming that could not be appreciated in blood or MVA hospital with severe RSV-bronchiolitis [24]. These data indi-
boosting results in greater gene expression where participants cate that several genes are differentially expressed following
have not been intramuscularly primed. In younger adults, the vaccination and are representative of the host response to
magnitude of expression of several genes correlated with the adenoviral vector vaccines, and are likely to have had bio-
immunological responses to vaccination measured at later logically relevant roles in the host response to these viral vec-
time points, suggesting some linkage between expression of tors and/or the RSV gene inserts in the vector. The fact that
these genes and the development of neutralizing antibody re- strong and consistent expression of interferon markers is seen
sponses. after vaccination is a promising indicator of efficacy, as these
One notable gene was IFI27, which had the greatest fold- responses have been linked to longer and more effective pro-
changes in induction after IM PanAd3-RSV and IM MVA- tection in viral vectored vaccination previously, and serve as a
RSV 3 days post vaccination. IFI27 expression is consistently promising indicator of immunogenicity.
observed in RSV infection with a high degree of consist- The PanAd3 vector applied by intra-nasal spray (used as
ency between RSV-specific signatures derived from PBMCs, an influenza vaccine in mice and RSV vaccine in rodents and
whole blood, and airway epithelial cells [23]. IFI27, together new-born calves) induced immune responses in the lungs and
with other genes that were differentially expressed, such spleen without any notable changes in immunity measured
as IFI44, OAS3, EPSTII, and IFI44L, are generally signifi- in blood that were protective to experimental challenges
cantly up-regulated in whole blood from infants admitted to [25–27]. Notably, the live-attenuated IN flu vaccine (LAIV,276 Green et al.
Downloaded from https://academic.oup.com/cei/article/211/3/269/6979728 by guest on 07 September 2023
Figure 4 (A–F) Correlation between gene expression and neutralising antibody in older adults Pearson correlation between changes in gene expression
3 days post prime vaccination in older adults as measured by rt-qPCR and log 2-fold change in RSV neutralizing antibody measured at 28 days post
vaccination. Grey area indicates confidence interval for the data. Line represents degree of correlation between fold change in gene expression and fold
change in neutralizing antibody.
FluMist) protects from infection despite a relative paucity expression 7 days after vaccination and only in children less
of responses measurable in blood [28, 29]. RSV-specific im- than 5-years old [30].
mune responses to IN PanAd3-RSV prime in humans were While our analyses showed a modest number of differen-
equally absent or minimal in blood, but these responses tially expressed genes seven days post prime vaccination, with
were inducible with the first dose of IM vaccine at boost PanAd3-RSV and boost vaccination with MVA-RSV, ranked
(IM PanAd3-RSV or IM MVA-RSV). Of particular interest gene set enrichment analysis (ranked using t-statistics for dif-
was the absence of expected transcriptional responses to the ferential gene expression) of the entire probeset showed en-
boosting with IM PanAd3-RSV (after IN prime) when com- richment for a number of terms related to the development
pared with the greater number of differentially expressed of memory CD4+ and CD8+ T-cell responses and humoral
probes induced following intramuscular boosting after immunity at these time points. The development of these
intranasal priming. These observations, combined with the memory and humoral immune responses seven days post vac-
gene set enrichment analysis, indicated a subtle response cination with a viral vector vaccine has been previously ob-
to mucosal immunisation with potential to alter the blood served, and is consistent with what is accepted and desirable
transcriptomic response to an IM booster vaccine. Similar for these vaccinations [31]. Several genes correlated with clin-
to what was observed here, the injectable inactivated in- ical and antigen-specific immune responses, giving an indica-
fluenza vaccine (TIV) induced the expression of interferon tion of potential pathways that lead towards safe and durable
genes shortly after vaccination. In contrast, intra-nasal live- protective immunity, however these genes may not neces-
attenuated influenza vaccine (LAIV) induced interferon gene sarily reflect the process by which these immune responsesClinical and Experimental Immunology, 2023, Vol. 211, No. 3 277
are generated in at risk groups such as older adults. It is dif- tracked over an RSV season and rate at which RSV infections
ficult to control for the effect of previous RSV exposure in are developed, as well as the severity of those infections is
conducting analysis on responses to RSV vaccination, as all tracked and compared with gene expression markers.
participants will have differing degrees of protection against As PanAd3 and MVA constructs without the RSV insert
RSV, which will make it more difficult to understand the rela- were not included in this study, it was impossible to make the
tionship between immune transcriptomic signalling and anti- comparison between the response to the vector and the more
body mediated immunological protection. specific responses to the insert.
This was a post hoc analysis and subject to limitations
common to first-in-man clinical trials in that there were
only a small number of volunteers in each prime/boost study Conclusions
group and the potential for confounding chance observa- A small, shared core of transcriptional responses 3 days after
tions. This included insufficient statistical power for direct vaccination, mostly related to type I interferon responses typ-
between-group comparisons. The within group analyses ically followed each dose of IM vaccine, with MVA respon-
used here were applied with all effort to correct for baseline sible for a greater magnitude of transcriptional changes than
Downloaded from https://academic.oup.com/cei/article/211/3/269/6979728 by guest on 07 September 2023
heterogeneity using the lmfit model, but we cannot assume the adenovirus vector. Analysis of the gene expression profiles
that differences in the magnitude of gene expression was has revealed the potential for expression of discrete and inter-
driven by vaccine alone. For example, more differentially dependent molecular pathways in response to different route
expressed probes were detected 3 days after IM PanAd3- of immunisation and different viral vectors.
RSV in study group 2 (compared with study group one),
when 6/10 group 2 individuals were actually sampled 2 days
after vaccination and all other samples were obtained 3 days Supplementary data
after vaccination (this was permissible in the trial protocol). Supplementary data is available at Clinical and Experimental
Adding the day of sampling as well as the baseline serum Immunology online.
PanAd3 (vector) neutralizing antibody titre, which was also
significantly different between study groups 1 and 2 vo-
lunteers, as covariates into the lmfit model did not signifi- Acknowledgements
cantly alter the number of differentially expressed probes We thank the Oxford Genomics Centre at the Wellcome
following IM PanAd3-RSV prime (data not shown). Using Centre for Human Genetics (funded by Wellcome Trust
DNA microarray data from whole blood is representative of grant reference 203141/Z/16/Z) for the generation and initial
the gene expression from a diversity of cell populations in processing of the microarray data. We also wish to acknow-
circulation. What remains unclear is the extent to which the ledge the team of research staff at the Oxford Vaccine Group,
fold-change in gene expression from whole blood is influ- University of Oxford, the Jenner Institute Transcriptomics
enced by relative changes in different cell populations after Core Facility, University of Oxford, and especially thank
vaccination. For example, neutrophils transiently declined 3 the many volunteers who were willing to contribute to this
days after vaccination (data not shown) which was not in- research. E.B. acknowledges the support of the Oxford
cluded in the lmfit model. NIHR Biomedical Research Centre and is an NIHR Senior
The findings of this study should be confirmed in fur- Investigator. The views expressed in this article are those of
ther studies. The capacity to predict neutralizing antibody the author and not necessarily those of the NHS, the NIHR,
responses should ideally be modelled in a controlled study or the Department of Health.
designed specifically to measure the relationship between
expression of these genes and neutralization. This could be
done in further mouse and human experiments where partici- Ethics and dissemination
pants are vaccinated with RSV001, and expression of specific RSV001 has clinical trial authorization from the Medicines
genes are assessed in a controlled study designed to deter- and Healthcare Products Regulatory Agency (MHRA) and
mine the relationship between expression of these genes and ethics approval from NRES Berkshire (reference 13/SC/0023).
a neutralization response. It would also be interesting to as- All study procedures adhere to International Conference on
sess if the protein expression of the genes identified in this Harmonisation (ICH) Good Clinical Practice guidelines.
study are also associated with neutralization via an assay that The results of the trial are to be published in peer-reviewed
could quantify protein expression, such as western blot. This journals, conferences, and academic forums. All animal re-
would allow us to assess if the effects on neutralization of searches conducted in this study were conducted according to
these genes is due to some downstream genes regulated by the ARRIVE guidelines.
genes identified in this study, or by expression of the identified
genes themselves.
It is difficult with this study design to determine the efficacy Informed consent
of the vaccine in the populations studied, as rates of infection Volunteers were healthy adults aged 18–50 years (YA) or
after vaccination were not tested. While it is possible in this 65–75 years (OA) who gave written informed consent to
study to compare gene expression to markers of vaccine effi- enter the trial, which included the analysis of host genetic fac-
cacy, such as neutralization and expression of markers such tors. The eligibility criteria, screening and selection of volun-
as interferons, there is not yet a single widely accepted cor- teers is described in detail in published material(Green et al,
relate for protection against RSV. Testing for relationship of 2015a). Note the terms ‘prime’ and boost’ used by convention
this biomarker with protective efficacy of the vaccine would from studies in antigen-naïve populations, but for RSV all vo-
require a study where infection rates of each participant are lunteers have natural immunity from previous exposure and278 Green et al.
the terms are used in relation to the first and second dose of • Clinical Trials Authorisation from the UK Medicines and
vaccine. Healthcare Products Regulatory Agency (MHRA refer-
ence 35082/0003/001-0001)
• University of Oxford Genetic Modification Safety
Funding Committee (GSMC reference GM462.11.64)
The RSV001 clinical trial was funded and sponsored by • Research Ethics Committee (NRES Berkshire reference
ReiThera S.r.l. (formerly Okairos S.r.l), which was ac- 13/SC/0023)
quired by GlaxoSmithKline Biologicals SA during the trial. • Local site and NHS R&D approvals
Further funding and support was provided by the NIHR
Oxford Biomedical Research and for Charles Sande and Paul
Klenerman from the Wellcome Trust (WT 091663MA). References
1. Ewer KJ, Lambe T, Rollier CS, Spencer AJ, Hill AV, Dorrell L. Viral
vectors as vaccine platforms: from immunogenicity to impact. Curr
Author contributions Opin Immunol 2016, 41, 47–54. doi:10.1016/j.coi.2016.05.014.
Downloaded from https://academic.oup.com/cei/article/211/3/269/6979728 by guest on 07 September 2023
2. Folegatti PM, Ewer KJ, Aley PK, Angus B, Becker S, Belij-
C.G. was the lead trial physician and in equal contribution
Rammerstorfer S, et al. Safety and immunogenicity of the ChAdOx1
with J.M. wrote the manuscript. J.M. performed the GSEA
nCoV-19 vaccine against SARS-CoV-2: a preliminary report of a
analyses and together with C.S., D.O.’C., and J.M. who wrote phase 1/2, single-blind, randomised controlled trial. Lancet 2020,
the R script used for data analysis. C.O. and J.M. performed 396, 467–78.
the wet lab work and QC analyses. The analysis plan for 3. Ewer KJ, O’Hara GA, Duncan CJ, Collins KA, Sheehy SH, Reyes-
differentially expressed genes was performed by J.M., C.G., Sandoval A, et al. Protective CD8 + T-cell immunity to human ma-
C.S., and, D.O.’C. GSEA by C.G. and J.M. Correlates with laria induced by chimpanzee adenovirus-MVA immunisation. Nat
clinical/immunological outcomes were performed by C.G., Commun 2013, 4, 2836.
J.M., and C.S. P.K. and A.P. were named Investigators for 4. Sheehy SH, Duncan CJ, Elias SC, Collins KA, Ewer KJ, Spencer AJ,
RSV001 and together with C.G. instigated this work on vac- et al. Phase Ia clinical evaluation of the plasmodium falciparum
cine transcriptomics. All authors had input into the manu- blood-stage antigen MSP1 in ChAd63 and MVA vaccine vectors.
Mol Ther 2011, 19, 2269–76. doi:10.1038/mt.2011.176.
script and have approved the manuscript for publication.
5. Hayton EJ, Rose A, Ibrahimsa U, Del Sorbo M, Capone S, Crook A,
C.O. conducted qPCRs used in this manuscript. S.B. provided et al. Safety and tolerability of conserved region vaccines vectored
training and consultation on all laboratory assays. L.D. and by plasmid DNA, simian adenovirus and modified vaccinia virus
E.B. provided validation data alongside guidance on its in- Ankara administered to human immunodeficiency virus type 1-un-
terpretation. S.C. and S.-M.N. did data analyses and vector infected adults in a randomized, single-blind phase i trial. PLoS
antibody work on the validation study. C.D., D.S., and L.S.-R. One 2014, 9, e101591. doi:10.1371/journal.pone.0101591.
designed and conducted mouse vaccination plan and RSV 6. Smaill F, Jeyanathan M, Smieja M, Medina MF, Thanthrige-Don N,
neutralization assays. Ad vaccines used in mice were devel- Zganiacz A, et al. A human type 5 adenovirus-based tuberculosis
oped by C.R. and C.D. and work involving them under was vaccine induces robust T cell responses in humans despite preex-
conducted under the supervision of C.R. and C.D. S.C. and isting anti-adenovirus immunity. Sci Transl Med 2013, 5, 205ra134.
7. McShane H, Pathan AA, Sander CR, Keating SM, Gilbert SC,
A.V. developed the vaccine at Reithera s.r.l and facilitated this
Huygen K, et al. Recombinant modified vaccinia virus Ankara
study. expressing antigen 85A boosts BCG-primed and naturally acquired
antimycobacterial immunity in humans. Nat Med 2004, 10, 1240–
4. doi:10.1038/nm1128.
Conflict of interests 8. Antrobus RD, Coughlan L, Berthoud TK, Dicks MD, Hill AV, Lambe
A.P. has previously conducted clinical trials of vaccines on T, et al. Clinical assessment of a novel recombinant simian adeno-
behalf of Oxford University funded by GlaxoSmithKline virus ChAdOx1 as a vectored vaccine expressing conserved influenza
Biologicals SA and ReiThera S.r.l, but does not receive a antigens. Mol Ther 2014, 22, 668–74. doi:10.1038/mt.2013.284.
any personal payments from them. A.P. is chair of the UK 9. Swadling L, Capone S, Antrobus RD, Brown A, Richardson R,
Department of Health’s (DH) Joint Committee on Vaccination Newell EW, et al. A human vaccine strategy based on chimpanzee
adenoviral and MVA vectors that primes, boosts, and sustains
and Immunisation (JCVI), but the views expressed in this
functional HCV-specific T cell memory. Sci Transl Med 2014, 6,
manuscript do not necessarily represent the views of JCVI or 261ra153.
DH. A.V. is a named inventor on patent applications covering 10. Capone S, D’Alise AM, Ammendola V, Colloca S, Cortese R, Nic-
RSV antigen expression system (WO 2012/089833). The re- osia A, et al. Development of chimpanzee adenoviruses as vaccine
maining authors declare they have no competing interests. vectors: challenges and successes emerging from clinical trials. Ex-
pert Rev Vaccines 2013, 12, 379–93. doi:10.1586/erv.13.15.
11. Milligan ID, Gibani MM, Sewell R, Clutterbuck EA, Campbell D,
Data and materials availability Plested E, et al. Safety and immunogenicity of novel adenovirus
The RSV001 clinical trial was conducted in accordance type 26-and modified vaccinia Ankara-vectored Ebola vaccines: a
with the UK Medicines for Human Use (Clinical Trials) randomized clinical trial. JAMA 2016, 315, 1610–23.
12. Henao-Restrepo AM, Camacho A, Longini I, Watson C, Edmunds
Regulations 2004, and subsequent amendment. RSV001 was
W, Egger M, et al. Efficacy and effectiveness of an rVSV-vectored
registered with clinicaltrials.gov (reference NCT01805921), vaccine expressing Ebola surface glycoprotein: interim results from
the European Medicines Agency’s European Clinical Trials the Guinea ring vaccination cluster-randomised trial. Lancet 2015,
Database (EudraCT; reference 2011-003589-34) and was 386, 857–66.
portfolio adopted by the National Institute for Health Clinical 13. Ewer K, Rampling T, Venkatraman N, Bowyer G, Wright D, Lambe
Research Network. The following regulatory approvals were T, et al. A monovalent chimpanzee adenovirus Ebola vaccine
granted before study procedures commenced. boosted with MVA. N Engl J Med 2016, 374, 1635–46.Clinical and Experimental Immunology, 2023, Vol. 211, No. 3 279
14. Zhu F-C, Li Y-H, Guan X-H, Hou L-H, Wang W-J, Li J-X, et al. 23. Ioannidis I, McNally B, Willette M, Peeples ME, Chaussabel D,
Safety, tolerability, and immunogenicity of a recombinant adeno- Durbin JE, et al. Plasticity and virus specificity of the airway ep-
virus type-5 vectored COVID-19 vaccine: a dose-escalation, open- ithelial cell immune response during respiratory virus infection. J
label, non-randomised, first-in-human trial. Lancet 2020, 395, Virol 2012, 86, 5422–36. doi:10.1128/jvi.06757-11.
1845–54. doi:10.1016/s0140-6736(20)31208-3. 24. Fjaerli HO, Bukholm G, Krog A, Skjaeret C, Holden M, Nakstad
15. Blohmke CJ, O’Connor D, Pollard AJ. The use of systems biology B, et al. Whole blood gene expression in infants with respiratory
and immunological big data to guide vaccine development. Ge- syncytial virus bronchiolitis. BMC Infect Dis 2006, 6.
nome Med 2015, 7, 114. 25. Pierantoni A, Esposito ML, Ammendola V, Napolitano F, Grazioli
16. Querec T, Bennouna S, Alkan S, Laouar Y, Gorden K, Flavell R, et F, Abbate A, et al. Mucosal delivery of a vectored RSV vaccine is
al. Yellow fever vaccine YF-17D activates multiple dendritic cell safe and elicits protective immunity in rodents and nonhuman
subsets via TLR2, 7, 8, and 9 to stimulate polyvalent immunity. J primates. Mol Ther - Methods Clin Dev 2015, 2, 15018.
Exp Med 2006, 203, 413–24. doi:10.1084/jem.20051720. 26. Taylor G, Thom M, Capone S, Pierantoni A, Guzman E, Herbert
17. Green CA, Scarselli E, Voysey M, Capone S, Vitelli A, Nicosia A, R, et al. Efficacy of a virus-vectored vaccine against human and bo-
et al. Safety and immunogenicity of novel respiratory syncytial vine respiratory syncytial virus infections. Sci Transl Med 2015, 7,
virus (RSV) vaccines based on the RSV viral proteins F, N and 300ra127.
Downloaded from https://academic.oup.com/cei/article/211/3/269/6979728 by guest on 07 September 2023
M2-1 encoded by simian adenovirus (PanAd3-RSV) and MVA 27. Vitelli A, Quirion MR, Lo C-Y, Misplon JA, Grabowska AK,
(MVA-RSV); protocol for an open-label, dose-escalation, single- Pierantoni A, et al. Vaccination to conserved influenza antigens in
centre, phase 1 clinical trial in healthy adults. BMJ Open 2015, 5, mice using a novel simian adenovirus vector, PanAd3, derived from
e008748. doi:10.1136/bmjopen-2015-008748. the Bonobo Pan paniscus. PLoS One 2013, 8, e55435. doi:10.1371/
18. Green CA, Sande CJ, Scarselli E, Capone S, Vitelli A, Nicosia A, et journal.pone.0055435.
al. Novel genetically-modified chimpanzee adenovirus and MVA- 28. Beyer WEP, Palache AM, De Jong JC, Osterhaus ADME. Cold-
vectored respiratory syncytial virus vaccine safely boosts humoral adapted live influenza vaccine versus inactivated vaccine: systemic
and cellular immunity in healthy older adults. J Infect 2019, 78, vaccine reactions, local and systemic antibody response, and vac-
382–92. doi:10.1016/j.jinf.2019.02.003. cine efficacy: a meta-analysis. Vaccine 2002, 20, 1340–53.
19. Smyth GK. Linear models and empirical Bayes methods for 29. Sasaki S, Jaimes MC, Holmes TH, Dekker CL, Mahmood K, Kem-
assessing differential expression in microarray experiments. Stat ble GW, et al. Comparison of the influenza virus-specific effector
Appl Genet Mol Biol 2004, 3, 1Article3–25. doi:10.2202/1544- and memory B-cell responses to immunization of children and
6115.1027. adults with live attenuated or inactivated influenza virus vaccines. J
20. Smyth GK, Michaud J, Scott HS. Use of within-array replicate spots Virol 2007, 81, 215–28. doi:10.1128/jvi.01957-06.
for assessing differential expression in microarray experiments. Bio- 30. Cao RG, Suarez NM, Obermoser G, Lopez SMC, Flano E, Mertz
informatics 2005, 21, 2067–75. doi:10.1093/bioinformatics/bti270. SE, et al. Differences in antibody responses between trivalent in-
21. Reiner A, Yekutieli D, Benjamini Y. Identifying differentially activated influenza vaccine and live attenuated influenza vaccine
expressed genes using false discovery rate controlling procedures. correlate with the kinetics and magnitude of interferon signaling
Bioinformatics 2003, 19, 368–75. doi:10.1093/bioinformatics/ in children. J Infect Dis 2014, 210, 224–33. doi:10.1093/infdis/
btf877. jiu079.
22. Subramanian A, Tamayo P, Mootha V, Mukherjee S, Ebert B, 31. Wang C, Hart M, Chui C, Ajuogu A, Brian IJ, de Cassan SC, et al.
Gillette M, et al. Gene set enrichment analysis: a knowledge-based Germinal center B cell and T follicular helper cell responses to viral
approach for interpreting genome-wide expression profiles. Proc vector and protein-in-adjuvant vaccines. J Immunol 2016, 197,
Natl Acad Sci USA 2005, 102, 15545–50. 1242–51. doi:10.4049/jimmunol.1502472.You can also read

















































