The role of mammalian poly(A)-binding proteins in co-ordinating mRNA turnover
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
856 Biochemical Society Transactions (2012) Volume 40, part 4
The role of mammalian poly(A)-binding proteins
in co-ordinating mRNA turnover
Matthew Brook and Nicola K. Gray1
MRC Centre for Reproductive Health, Queen’s Medical Research Institute, University of Edinburgh, 47 Little France Crescent, Edinburgh EH16 4TJ, Scotland, U.K.
Abstract
The function of cytoplasmic PABPs [poly(A)-binding proteins] in promoting mRNA translation has been
intensively studied. However, PABPs also have less clearly defined functions in mRNA turnover including
roles in default deadenylation, a major rate-limiting step in mRNA decay, as well as roles in the regulation
of mRNA turnover by cis-acting control elements and in the detection of aberrant mRNA transcripts. In
the present paper, we review our current understanding of the complex roles of PABP1 in mRNA turnover,
focusing on recent progress in mammals and highlighting some of the major questions that remain to be
addressed.
Introduction absence of cis-acting regulatory elements, so-called default
The extent to which an mRNA is utilized is determined by its deadenylation occurs. Initial poly(A) removal occurs at a
lifespan and rate of translation. These are intimately linked constant low rate due to the distributive nature of PAN2–
and can be regulated by cis-acting elements in response to PAN3 which has a high on/off rate of RNA binding, therefore
the cellular context. In eukaryotes, the primary determinants only removing exposed poly(A). In contrast, CCR4–CAF1
of both mRNA translation and turnover are the 5 m7 GTP acts processively, presumably evicting PABP1, and rapidly
(7-methylguanosine triphosphate) ‘cap’ and 3 poly(A) tail removes the remaining ∼110 nucleotides to leave an oligo(A)
which act by binding eIF (eukaryotic initiation factor) 4F tail. This oligo(A) tail is bound by the LSm (like-Sm) 1-7–Pat1
(comprising eIF4E, eIF4G and eIF4A) and PABPs [poly(A)- complex which promotes removal of the 5 cap (decapping)
binding proteins] respectively. The association of these by the DCP (decapping protein) 1–DCP2 complex, allowing
5 - and 3 -bound factors, primarily through eIF4G–PABP access of the 5 →3 exonuclease XRN1 (exoribonuclease 1)
interactions, forms a ‘closed-loop’ mRNA conformation which degrades the body of the mRNA in concert with 3 →5
(Figure 1). This enhances ribosomal recruitment [1] and exonucleolytic activities (i.e. exosome and/or the LSm1-7–
protects both ends of the mRNA, most likely by increasing Pat1 complex) [3–5].
binding of these factors to block access of the mRNA Cytoplasmic PABPs are conserved across eukaryotes and
degradation machinery. are best characterized as central regulators of mRNA trans-
mRNA turnover is multi-step process in which deadenyla- lation (reviewed in [6–8]). Although mammals encode five
tion [shortening of the poly(A) tail] is usually the initial PABPs, almost all studies have focused on the prototypical
and primary rate-limiting step. In mammals, deadenylation family member, PABP1 (also known as PABPC1). PABP1
is a biphasic process where poly(A) tails are shortened to contains four non-identical RRMs (RNA-recognition motifs)
∼110 nucleotides (from approximately 250 nucleotides) by and a C-terminal region. RRMs 1 and 2 bind poly(A) with
the PAN [poly(A) nuclease] 2–PAN3 deadenylase complex high specificity, whereas RRMs 3 and 4 bind poly(A)
and subsequently to oligo(A) (approximately A10–18 ) by with reduced affinity and can bind adenine/uridine-rich
the CCR4 (carbon catabolite repressor protein 4)–CAF1 RNA, although their RNA-binding specificity remains less
(CCR4-associated factor 1) complex (reviewed in [2]). In the clear [6]. Interestingly, the RRMs also mediate important
protein interactions, including those with eIF4G [8].
The C-terminal region is composed of the PABC (PABP
Key words: cytoplasmic poly(A)-binding protein (PABP), deadenylation, microRNA (miRNA),
mRNA decay, nonsense-mediated decay, translation factor.
C-terminal domain)/MLLE domain which interacts with
Abbreviations used: ARE, adenine/uridine-rich element; AUBP, ARE-binding protein; AUF1, numerous PAM2 (PABP-interacting motif 2)-containing
adenine/uridine-rich RNA-binding factor 1; CCR4, carbon catabolite repressor protein 4; CAF1, proteins {e.g. eRF (eukaryotic release factor) 3 [8],
CCR4-associated factor 1; CRE, cytosine-rich element; DCP, decapping protein; eIF, eukaryotic
initiation factor; EJC, exon-junction complex; ErEN, erythroid-cell-specific endonuclease; eRF,
deadenylases [2,9,10] and TNRC6 (trinucleotide repeat-
eukaryotic release factor; LSm, like-Sm; mCRD, major coding-region determinant; miRNA, containing 6) [11]}, and a proline-rich linker that is involved
microRNA; NF-L, neurofilament light; NMD, nonsense-mediated decay; NOT, negative on TATA; in PABP1–PABP1 interactions which enhance co-operative
NSAP1, NS1-associated protein 1; PABP, poly(A)-binding protein; ePAPB, embryonic PABP; PABC,
PABP C-terminal domain; PAIP1, PABP-interacting protein 1; PAM2, PABP-interacting motif 2;
poly(A) binding [6,12,13]. The known protein partners of
PAN, poly(A) nuclease; PARN, poly(A)-specific ribonuclease; PTC, premature termination codon; PABP1 provide some insight into its complex functions in
PTM, post-translational modification; RISC, RNA-induced silencing complex; RRM, RNA-recognition both mRNA translation and stability, and in the present paper
motif; TNRC6, trinucleotide repeat-containing 6; TOB, transducer of ErbB2; TTP, tristetraprolin; UNR,
upstream of N-ras; UPF, up-frameshift; UTR, untranslated region; XRN1, exoribonuclease 1.
we review the current state of knowledge regarding its role in
1
To whom correspondence should be addressed (email Nicola.Gray@ed.ac.uk). regulating mRNA turnover in mammals.
C The Authors Journal compilation
C 2012 Biochemical Society Biochem. Soc. Trans. (2012) 40, 856–864; doi:10.1042/BST20120100
RNA UK 2012 857
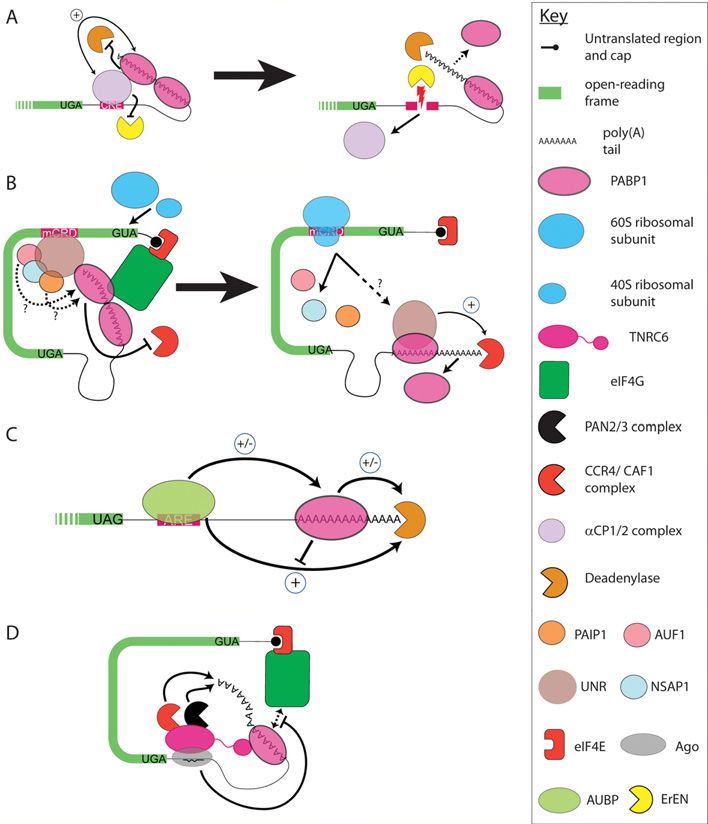
Figure 1 Role of PABP1 in the closed-loop mRNA conformation the CCR4–NOT–CAF1 complex via TOB (transducer of
mRNAs are bound at the cap by eIF4E, as part of the eIF4F complex (eIF4E, ErbB2) 1 (or TOB2) [2]. Somewhat counterintuitively,
eIF4G, eIF4A) and at the poly(A) tail by PABP1, protecting the mRNA given its poly(A)-protective effect, PABP1 also appears
ends from decapping and deadenylation. Both eIF4E and PABP1 bind the to enhance PAN2–PAN3 complex activity and, via TOB-
scaffolding protein eIF4G, which effectively bridges the 5 and 3 ends of mediated recruitment, CCR4–CAF1 deadenylase activity
the mRNA to form a ‘closed-loop’. This conformation increases the affinity [2,17] (Figure 2, panel 1). Such observations, although
of eIF4E for the cap and is proposed to increase the affinity of PABP1 confusing, support complex roles for PABP1 as both as
for the poly(A) tail, promoting small ribosomal subunit recruitment [1] a facilitator of deadenylation and as a negative regulator
and further protecting mRNAs from decay (not depicted). For simplicity, of deadenylation, with its interactions with deadenylases
other initiation factors that interact with PABP1, including those that raising the possibility that PABP1 may inhibit their
contribute to the closed-loop, are not shown. function directly as well as sterically. Since it was proposed
recently that only non-co-operatively bound PABP1 may
be available for PABC-mediated interactions [12], it is
tempting to speculate that co-operatively bound PABP1
acts as a steric block with non-co-operatively bound
PABP1 recruiting/stimulating deadenylases.
Might PABP1 be removed from poly(A) tails
during translation termination?
Work in yeast first highlighted potential links between
deadenylation and translational termination, by showing that
the translation termination factor eRF3 interacts with PAB-1.
This led to a model in which eRF3–PAB-1 interaction during
termination facilitates the removal of a PAB-1 molecule from
the poly(A) tail, rendering the 3 end open to limited 3 →5
exonucleolytic attack [18]. Subsequently, mammalian PABP1
was shown to interact in a mutually exclusive manner with
eRF3, PAN3 and TOB, and, intriguingly, switching of PABP1
from an eRF3-bound to PAN3-bound state requires active
translation since it can be blocked by cycloheximide [19].
eRF3, PAN3 and TOB all contain a PAM2 motif, and ectopic
expression of PAM2-mutant versions of these proteins was
PABP1 function in default mRNA found to exert opposing effects on mRNA deadenylation
deadenylation and decay rates [19]. Thus, similar to yeast, a model was proposed
in which formation of a PABP1–eRF3–eRF1 termination
The dual role of PABP1 in both poly(A) tail complex causes PABP1 to dissociate from the poly(A) tail
protection and deadenylation (Figure 2, panels 3 and 4). Displaced PABP1 leaves the
The stabilizing effect of a poly(A) tail on a reporter mRNA PABP1–eRF3–eRF1 complex upon eRF1 stimulation of
in vitro is negated by sequestration or depletion of PABP1, eRF3-mediated GTP hydrolysis [19,20] and it, or the 3 -most
and can be rescued by addition of exogenous PABP1, PABP1, may then recruit/stimulate PAN3 or TOB1–CCR4–
with non-adenylated mRNAs being rapidly turned over CAF1.
independently of PABP1 status [14]. Since intermediates A link between translation termination and incremental
corresponding to the loss of footprints of consecutive PABP shortening of the poly(A) tail could provide a molecular
molecules can be observed [15,16], this led to the idea timer for mRNA lifespan, limiting the number of times
that PABP1 may act as a steric block to deadenylases that an mRNA molecule can be translated [18,19]. However,
(Figure 2, panel 1). Such a function will be determined most mRNAs will be translated by many more transiting
by the dissociation rate (K d ) of PABP1 (Figure 2, panel ribosomes than would be required to evict the number of
2) which is influenced by whether it is bound non-co- PABP1 molecules bound to the poly(A) tail, suggesting that
operatively or co-operatively to poly(A), with the latter eviction does not occur at every termination event. Moreover,
exhibiting higher affinity [12]. Mammalian cells contain it is unlikely that all default deadenylation is linked to
three major poly(A)-specific 3 exoribonuclease activities: termination, since reducing the translational efficiency of
the CCR4–NOT (negative on TATA)–CAF1 complex, the a reporter mRNA by introducing a hairpin-loop into the
PAN2–PAN3 complex and PARN [poly(A)-specific 5 -UTR (untranslated region) does not block deadenylation
ribonuclease]. Intriguingly, given its poly(A)-protective [21]. As discussed above (Figure 2, panel 2), this termination-
effect, PABP1 interacts directly with PARN and the PAN3 independent deadenylation may simply reflect the off rate
subunit of the PAN2–PAN3 complex and indirectly with of PABP1 and/or, since end-to-end complexes may still
C The Authors Journal compilation
C 2012 Biochemical Society
858 Biochemical Society Transactions (2012) Volume 40, part 4
Figure 2 Role of PABP1 in default deadenylation
(1) PABP1 bound to the poly(A) tail prevents access of deadenylases (blunted arrow), but can also recruit and stimulate
deadenylases (thin arrow), an activity that is blocked by its interaction with eRF3 which competes for the same binding site
within PABC. (2–4) Removal of PABP1 from the poly(A) tail (arrow) facilitates access of deadenylases either as a result of
(2) the inherent ‘off’ rate of PABP1 or (3) due to translation termination, since indirect interaction of PABP1 with terminating
ribosomes, via an eRF3–eRF1 bridge, causes (4) PABP1 to be evicted from the poly(A) tail. (5) Deadenylated mRNAs
are decapped by DCP1–DCP2 (not depicted) and degraded by XRN1 (not depicted), the exosome and/or LSm 1-7–Pat1.
TOB1–TOB2 is not depicted.
form on these mRNAs, PABP1-mediated co-ordination of an apparent K d of 150 nM [24], suggesting that, under certain
deadenylase activity. circumstances, poly(A) tail-associated PABP1 could directly
protect mRNAs from decapping. However, this binding only
A direct role for PABP1 in regulating decapping? occurs in the absence of the cap-binding factor eIF4E, thus it
Deadenylation triggers decapping in mammalian cells ([22] is unclear whether this interaction can, or does, occur in cells.
and reviewed in [23]). Both PABP1 and the poly(A)
tail are required to protect mRNAs from DCP2-mediated
decapping in vitro [24,25], suggesting that protection of Regulation of aberrant transcript turnover
the poly(A) tail by PABP1 is likely to be sufficient to PTCs (premature termination codons) have been described in
account for this effect. Intriguingly, however, poly(A)- approximately 30 % of known disease-associated mutations
bound PABP1, but not free PABP1, can bind the RNA- [26] and promote aberrant as well as premature translation
incorporated cap structure, but not free cap, in vitro with termination. As a result of their aberrant translation
C The Authors Journal compilation
C 2012 Biochemical Society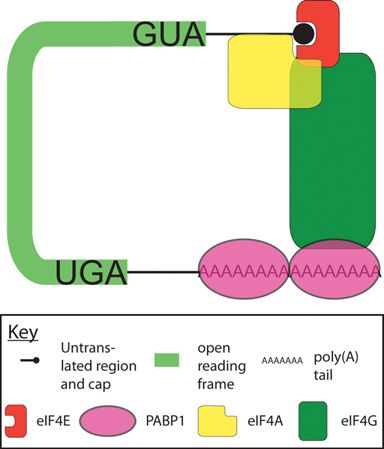
RNA UK 2012 859
Figure 3 PABP1 suppresses NMD
(1) mRNAs are initially bound by the nuclear CBC80–CBC20 cap-binding complex with the EJC–UPF2-containing complex
marking splicing boundaries. As an mRNA emerges from the nucleus, CBC20–CBC80 is exchanged for the cytoplasmic
cap-binding complex [only eIF4E and eIF4G (4G) are shown], allowing bridging interactions with PABP1. (2) During translation,
the transiting ribosome displaces the EJC and PABP1–eRF3 complexes promote correct termination ( + ) by interacting with
eRF1 (not shown). Formation of this complex blocks recruitment of the NMD factor UPF1 (blunted arrow). (3) In contrast,
when PTCs occur 5 to an EJC, the EJC is not displaced and PABP1 is not proximal to the stop codon (4). Thus normal
PABP1–eRF3-mediated termination does not occur (blunted arrow) and UPF1 binds the ribosome-associated eRF3, recruiting
SMG1. (5) The stalled ribosome exits the PTC and the UPF1-containing complex contacts the EJC–UPF2-containing complex
and inhibits further translation initiation (blunted arrow). (6) The endonuclease SMG6 then cleaves the mRNA 3 to the PTC
with (7) the mRNA fragments then being degraded in a deadenylation-independent manner. For simplicity, not all NMD
factors are depicted.
termination, PTC-containing mRNAs can be recognized by NMD factors and deadenylation-independent endonuc-
the mRNA-surveillance machinery and subjected to NMD leolytic cleavage of the mRNA [27,28]. PABP1 therefore
(nonsense-mediated decay). In mammals, NMD appears to plays a role in deciding whether to commit mRNAs for
be triggered by UPF (up-frameshift) 2-containing EJCs NMD, since its interaction with eRF3 on the terminating
(exon-junction complexes) located 3 of PTCs. Prematurely ribosomes at bona fide stop codons promotes ‘normal’
terminating ribosomes undergo abnormal termination, termination and prevents eRF3–UPF1 interaction [29]
with eRF3 being bound by UPF1, which also interacts with (Figure 3). The relative proximity of PABP1 to the
EJC-associated UPF2, leading to the recruitment of further termination codon appears to be important in this decision,
C The Authors Journal compilation
C 2012 Biochemical Society860 Biochemical Society Transactions (2012) Volume 40, part 4
as NMD can be suppressed by tethering PABP1 in the RNA-binding factor 1), NSAP1 (NS1-associated protein 1)
vicinity of PTCs but 5 to EJCs [30–32]. However, although and PABP1, bridging a functional interaction between the
introduction of a PTC which creates a long ‘faux’ (or false) mCRD and the poly(A) tail and preventing deadenylation
3 -UTR leads to NMD, many mammalian mRNAs contain [40] (Figure 4B). The key interaction in forming this bridging
long 3 -UTRs and are not subject to NMD [27], underscoring complex occurs between PABP1 and UNR, although PAIP1
the importance of a 3 EJC as well as PABP1 proximity. and AUF1 are also known PABP1-interacting proteins, but
Nonetheless, increasing the distance between poly(A)-bound it is unclear whether they interact directly with PABP1
PABP and a normal stop codon by extending the 3 -UTR in this context [41]. During c-fos mRNA translation, the
can induce EJC-independent NMD, which is antagonized by transiting ribosome displaces the mCRD-bound complex
reinstating PABP1 proximity [33]. It is not clear how this [40,41], which may remain associated with poly(A)-bound
form of NMD is promoted, but one idea is that unknown PABP1, disrupting the association between the mCRD and
3 -UTR-associated factors prevent PABP1 from interacting the poly(A) tail and apparently permitting recruitment of the
with eRF3 and promote loading of UPF1 to initiate NMD CCR4 deadenylase by UNR [41]. Thus the translation of c-
[33]. fos mRNA acts as a trigger for its destruction, thereby limiting
the expression of this oncogene by restricting the lifetime of
its mRNA.
mRNA-specific regulation of mRNA
stability by PABP1 PABP1, AREs and AUBPs
Although AUBPs (ARE-binding proteins) can enhance or
The rate of mRNA decay can be modulated by sequences
reduce mRNA stability and/or translation, the presence of
and/or structural features within mRNAs that, most often,
an ARE generally confers reduced basal stability to mRNAs
recruit specific trans-acting factors (reviewed in [2,23,34]).
by enhancing deadenylation [2,42]. Thus it seems likely that
These cis-acting elements are normally located within
there is functional cross-talk between AREs and PABP, and,
the 3 -UTR as exemplified by AREs (adenine/uridine-rich
in support of this, PABP1 associates with AUBPs and can
elements) and the α-globin mRNA CRE (cytosine-
bind to A/U-rich RNA, including functional AREs, in vitro
rich element) or less frequently, as is the case for c-fos mRNA,
[43]. However, despite the large number of identified ARE-
in the coding region.
containing mRNAs and the contribution of misregulated
ARE-mediated control to disease (e.g. chronic inflammatory
PABP1-mediated regulation of endonuclease disorders and cancer [34,44,45]), our understanding of the
recruitment role of PABPs in ARE-mediated mRNA decay is limited and
Erythrocytes are enucleated and post-transcriptionally its role may vary between mRNAs (Figure 4C).
regulate the synthesis of α- and β-globin subunits for For instance, an interaction between ARE-bound HuR
haemoglobin production, with an imbalance in their synthesis and poly(A)-bound PABP1 was posited to enhance the
leading to thalassaemias, defective erythropoiesis and/or ability of PABP1 to protect the poly(A) tail of β-casein
peripheral haemolysis [35,36]. The 3 -UTR of α-globin mRNA, contributing to its stability during lactation [46].
mRNA contains a CRE which is bound by the poly(C)- The underlying mechanism remains to be determined, but
binding protein complex αCP1–αCP2 [15]. Interaction may reflect altered interaction of PABP1 with poly(A) or
between PABP1 and αCP1–αCP2 causes an increase in deadenylases. In contrast, the AUBP TTP (tristetraprolin)
both αCP1–αCP2 affinity for the CRE and PABP1-poly(A) destabilizes mRNA by directly recruiting CAF1 deadenylase
tail binding, simultaneously blocking ErEN (erythroid-cell- [47,48], with TTP–PABP1 interactions being reported to
specific endonuclease) from cleaving at the CRE [15,37] and inhibit TTP-mediated deadenylation [49]. However, TTP
reducing deadenylation to stabilize the mRNA (Figure 4A). dephosphorylation not only enhances its ability to recruit
A similar mechanism is employed for the stabilization of NF- CAF1, but also enhances its interaction with PABP1, which
L (neurofilament light) mRNA, whose misregulation results appears at odds with its proposed inhibition of TTP-mediated
in motor neuron degeneration. PABP1 blocks aldolase C- deadenylation [47]. Thus the functional links between PABP1
mediated endonucleolytic cleavage within the 3 -UTR of NF- and TTP await clarification.
L mRNA, with in vitro studies showing that it binds a 68 nt PABP1 also interacts with another mRNA-destabilizing
polypyrimidine-rich region [38,39], implying that PABP1 AUBP, AUF1, whose deficiency in mice leads to the
may bind to sequences other than poly(A) or A/U. stabilization of ARE-containing mRNAs associated with
chronic inflammatory diseases [50]. Whereas PABP1 is
PABP1-mediated regulation of mRNA stability proposed to alter TTP function, AUF1 is posited to alter
via open reading frame cis-acting elements PABP1 function. Although the AUF1–PABP1 interaction
The c-fos mRNA mCRD (major coding-region determinant) does not directly affect PABP1 poly(A) binding, it was
is an 87 nt G/A-rich element that directs translationally proposed that it causes PABP1 eviction from the poly(A)
coupled deadenylation. The c-fos mCRD binds a mul- tail, possibly by modulating the PABP1–eIF4G interaction,
tiprotein complex containing UNR (upstream of N-ras), leading to deadenylation. Consistent with this model, Hsp70
PAIP1 (PABP-interacting protein 1), AUF1 (A/U-rich (heat-shock protein 70), which stabilizes ARE-containing
C The Authors Journal compilation
C 2012 Biochemical SocietyRNA UK 2012 861
Figure 4 Roles of PABP1 in regulating mRNA-specific mRNA turnover
(A) Left: α-globin mRNA is protected from the action of ErEN and deadenylation (blunted arrows) by 3 -UTR CRE-associated
αCP1–αCP2 complex which interacts with PABP1, promoting both CRE and poly(A) binding (double-headed arrow). Right:
in the absence of the αCP1–αCP2 complex binding, ErEN can cleave within the CRE (zigzag arrow), PABP1 poly(A) binding
will be reduced (broken arrow) and the cleaved mRNA fragments will be subject to decay. (B) Left: on non-translating c-fos
mRNA, the mCRD is bound by UNR in complex with AUF1, PAIP1 and NSAP1. UNR (and possibly other complex components,
depicted as broken arrows) interacts with PABP1 and inhibits c-fos mRNA deadenylation (blunted arrow) by an unknown
mechanism. Right: upon translation of c-fos mRNA, the transiting ribosome displaces the UNR-containing complex from
the mCRD, although it may remain associated with the poly(A) tail via PABP1 (broken arrow). UNR is then able to recruit
CCR4–CAF1, and possibly also PAN2–PAN3 (not shown), to promote ( + ) deadenylation. (C) ARE-containing mRNAs are bound
by one or more AUBPs. AUBPs can promote deadenylation directly through recruitment and stimulation of deadenylases and
this can be antagonized by PABP1. AUBPs may also promote deadenylation indirectly by altering PABP1 poly(A) binding or
modulating PABP1-mediated stimulation of deadenylases. Conversely, AUBPs may also inhibit deadenylation by enhancing
PABP1 poly(A) binding and blocking its ability to stimulate deadenylase activity. Positive and inhibitory actions are depicted by
+ or − respectively. (D) PABP1 function is modulated during miRNA-mediated translational repression and deadenylation.
RISC (only TNRC6 and Ago shown) is brought to the mRNA via miRNA complementarity. TNRC6 interacts with PABP1 directly
(shown) and indirectly (not shown) and disrupts its interaction with eIF4G (broken arrow), an interaction which promotes the
closed-loop conformation and may also enhance PABP1 poly(A) binding (not depicted). This results in repressed translation
(blunted arrow), facilitating deadenylation. TNRC6 also directly recruits deadenylases and may bring them into close proximity
to the poly(A) tail via its interaction with PABP1, to promote mRNA deadenylation (arrows). Deadenylase (A and C, see key)
denotes events in which a specific deadenylase(s) has not been implicated.
C The Authors Journal compilation
C 2012 Biochemical Society862 Biochemical Society Transactions (2012) Volume 40, part 4
mRNAs [51], disrupts the PABP–AUF1 interaction [51]. PABP in oocytes and early embryos [66,67] and has mainly
However, ARE binding by AUF1 inhibits its interaction with has been studied in X. laevis, where it is required for
PABP1 in vitro [52], suggesting that this interaction does not both oocyte maturation [68] and early development [64].
occur on ARE-containing mRNAs that are subject to AUF1- X. laevis ePABP binds poly(A) [64,66] and has been shown
mediated deadenylation. Since this observation is inconsistent to promote mRNA-specific cytoplasmic polyadenylation by
with an ARE-specific role for this interaction, it raises protecting newly synthesized poly(A) tails in vitro and in vivo
questions regarding the role of PABP1 in AUF1-mediated [68,69] and to prevent deadenylation of ARE-containing
destabilization. Moreover, AUF1 can also bind poly(A) tails mRNAs in vitro [66]. It is also likely to inhibit default
and other components of the closed-loop complex directly deadenylation via its ability to promote translation initiation
[52,53], emphasizing further that the role of PABP1 in AUF1- through eIF4F interaction [67] and translation termin-
mediated mRNA turnover remains to be fully characterized. ation through interaction with eRF3 [70]. Recent studies in
mice have shown that mammalian ePABP shares the ability
to promote cytoplasmic polyadenylation and is required for
PABP1 and miRNAs (microRNAs)
oocyte maturation [71].
miRNAs play a crucial role in post-transcriptional regulation,
and their dysregulation is implicated in the aetiology of a
wide spectrum of human diseases (reviewed in [54]). miRNA-
mediated silencing utilizes a host of protein factors, including
the RISC (RNA-induced silencing complex) (reviewed
Perspectives
Although it is apparent that PABP1 has pleiotropic roles in
extensively in [55,56]) and PABP1 and is generally considered
mRNA turnover, it is also clear that there are currently more
to involve translational repression and deadenylation. To
questions pertaining to those roles than answers. One reason
date, the order of these events and the extent to which they
for the current lack of clarity likely stems from the use of
promote decay remain controversial.
multiple experimental systems, each optimized to analyse
PABP1 interacts with TNRC6 [56] (Figure 4D), the
specific steps in the mRNA translation/turnover pathway.
RISC component paralogue of GW182, of which there
Thus the development of an in vitro system that retains
are isoforms A–C in mammals. TNRC6 interacts with
regulated coupled mRNA translation/turnover would be a
PABC via a non-canonical PAM2 motif and indirectly
boon for studies of post-transcriptional control, and PABPs
with the RRM region that binds eIF4G, via an unknown
in particular. Another reason may be an overreliance on a
partner [56–58]. The TNRC6–PABP1 interaction is re-
small number of reporter mRNAs (e.g. β-globin reporter),
quired for effective miRNA-mediated silencing [58,59],
which are often used in cell types for which they may be
but, intriguingly, TNRC6 recruits both CCR4–CAF1 and
inappropriate.
PAN2–PAN3 deadenylase complexes directly, and PABP1-
A major question pertaining to PABP1 function in
independently, to effect deadenylation [60–62]. Thus TNRC6
default deadenylation relates to understanding the apparent
may disrupt the PABP1–eIF4G interaction, which is
fundamental contradiction between the abilities of poly(A)-
important for the closed-loop conformation and may also
bound PABP1 to both protect an mRNA from deadenylation
strengthen PABP1 poly(A) binding, to repress translation and
and to recruit/enhance deadenylase activities. It is tempting
enhance deadenylation. Alternatively, TNRC6 may utilize
to speculate that this may be controlled, at least in part,
its interaction with PABP1 to bring the poly(A) tail into
by competition between PAN2–PAN3, the TOB1–TOB2–
proximity of its associated deadenylases [63]. These models
CCR4–CAF1 complex and other PABC partners, which
are not mutually exclusive.
have differing affinities (e.g. eRF3RNA UK 2012 863
Although even less well understood, mRNA-specific 12 Lin, J., Fabian, M., Sonenberg, N. and Meller, A. (2012) Nanopore
roles for PABP1 in mRNA turnover are fast emerging and detachment kinetics of poly(A) binding proteins from RNA molecules
reveals the critical role of C-terminus interactions. Biophys. J. 102,
supported by multiple findings, although such roles remain 1427–1434
to be demonstrated in vivo. However, a better understanding 13 Melo, E.O., Dhalia, R., Martins de Sa, C., Standart, N. and de Melo Neto,
of the role of PABP1 in global mRNA turnover may first be O.P. (2003) Identification of a C-terminal poly(A)-binding protein
(PABP)–PABP interaction domain: role in co-operative binding to poly(A)
necessary to inform studies of its mRNA-specific functions. and efficient cap distal translational repression. J. Biol. Chem. 278,
Thus whether PABP1 contributes to the regulation of 46357–46368
specific mRNAs in multiple ways or whether a unifying 14 Bernstein, P., Peltz, S.W. and Ross, J. (1989) The poly(A)–poly(A)-binding
protein complex is a major determinant of mRNA stability in vitro. Mol.
mechanism exists remains an important question. A final Cell. Biol. 9, 659–670
important, and extremely challenging, question pertains 15 Wang, Z., Day, N., Trifillis, P. and Kiledjian, M. (1999) An mRNA stability
to the contribution of PABP1-mediated regulation of complex functions with poly(A)-binding protein to stabilize mRNA
in vitro. Mol. Cell. Biol. 19, 4552–4560
both default deadenylation/mRNA stabilization and 16 Korner, C.G., Wormington, M., Muckenthaler, M., Schneider, S., Dehlin, E.
mRNA-specific decay to mammalian physiology. and Wahle, E. (1998) The deadenylating nuclease (DAN) is involved in
poly(A) tail removal during the meiotic maturation of Xenopus oocytes.
EMBO J. 17, 5427–5437
17 Ezzeddine, N., Chen, C.Y. and Shyu, A.B. (2012) Evidence providing new
Acknowledgements insights into TOB-promoted deadenylation and supporting a link
between TOB’s deadenylation-enhancing and anti-proliferative activities.
Mol. Cell. Biol. 32, 1089–1098
We thank the organizers of RNA UK 2012 for organizing an excellent
18 Hoshino, S., Imai, M., Kobayashi, T., Uchida, N. and Katada, T. (1999) The
meeting and past and present members of the Gray laboratory for eukaryotic polypeptide chain releasing factor (eRF3/GSPT) carrying the
helpful discussions of this paper. translation termination signal to the 3 -poly(A) tail of mRNA: direct
association of erf3/GSPT with polyadenylate-binding protein. J. Biol.
Chem. 274, 16677–16680
19 Funakoshi, Y., Doi, Y., Hosoda, N., Uchida, N., Osawa, M., Shimada, I.,
Tsujimoto, M., Suzuki, T., Katada, T. and Hoshino, S. (2007) Mechanism of
Funding mRNA deadenylation: evidence for a molecular interplay between
translation termination factor eRF3 and mRNA deadenylases. Genes Dev.
Work in the Gray laboratory is funded by the Medical Research 21, 3135–3148
20 Hauryliuk, V., Zavialov, A., Kisselev, L. and Ehrenberg, M. (2006) Class-1
Council (transition funding and a Program Grant) and project grants release factor eRF1 promotes GTP binding by class-2 release factor eRF3.
from the Biotechnology and Biological Sciences Research Council Biochimie 88, 747–757
and Wellcome Trust. 21 Chen, C.Y., Xu, N. and Shyu, A.B. (1995) mRNA decay mediated by two
distinct AU-rich elements from c-fos and granulocyte-macrophage
colony-stimulating factor transcripts: different deadenylation kinetics and
uncoupling from translation. Mol. Cell. Biol. 15, 5777–5788
22 Yamashita, A., Chang, T.C., Yamashita, Y., Zhu, W., Zhong, Z., Chen, C.Y.
References and Shyu, A.B. (2005) Concerted action of poly(A) nucleases and
1 Kahvejian, A., Svitkin, Y.V., Sukarieh, R., M’Boutchou, M.N. and decapping enzyme in mammalian mRNA turnover. Nat. Struct. Mol. Biol.
Sonenberg, N. (2005) Mammalian poly(A)-binding protein is a 12, 1054–1063
eukaryotic translation initiation factor, which acts via multiple 23 Franks, T.M. and Lykke-Andersen, J. (2008) The control of mRNA
mechanisms. Genes Dev. 19, 104–113 decapping and P-body formation. Mol. Cell 32, 605–615
2 Chen, C.Y. and Shyu, A.B. (2010) Mechanisms of deadenylation- 24 Khanna, R. and Kiledjian, M. (2004) Poly(A)-binding-protein-mediated
dependent decay. Wiley Interdiscip. Rev.: RNA 2, 167–183 regulation of hDcp2 decapping in vitro. EMBO J. 23, 1968–1976
3 Tharun, S. (2009) Roles of eukaryotic Lsm proteins in the regulation of 25 Wang, Z., Jiao, X., Carr-Schmid, A. and Kiledjian, M. (2002) The hDcp2
mRNA function. Int. Rev. Cell Mol. Biol. 272, 149–189 protein is a mammalian mRNA decapping enzyme. Proc. Natl. Acad. Sci.
U.S.A. 99, 12663–12668
4 Ling, S.H., Qamra, R. and Song, H. (2011) Structural and functional
26 Bhuvanagiri, M., Schlitter, A.M., Hentze, M.W. and Kulozik, A.E. (2010)
insights into eukaryotic mRNA decapping. Wiley Interdiscip. Rev.: RNA 2,
NMD: RNA biology meets human genetic medicine. Biochem. J. 430,
193–208
365–377
5 Lykke-Andersen, S., Tomecki, R., Jensen, T.H. and Dziembowski, A.
27 Muhlemann, O. and Lykke-Andersen, J. (2011) How and where are
(2011) The eukaryotic RNA exosome: same scaffold but variable
nonsense mRNAs degraded in mammalian cells? RNA Biol. 7, 28–32
catalytic subunits. RNA Biol. 8, 61–66
28 Schoenberg, D.R. and Maquat, L.E. (2012) Regulation of cytoplasmic
6 Brook, M., Smith, J.W.S. and Gray, N.K. (2009) The DAZL and PABP
mRNA decay. Nat. Rev. Genet. 13, 246–259
families: RNA-binding proteins with interrelated roles in translational 29 Maquat, L.E., Hwang, J., Sato, H. and Tang, Y. (2010) CBP80-promoted
control in oocytes. Reproduction 137, 595–617 mRNP rearrangements during the pioneer round of translation,
7 Jackson, R.J., Hellen, C.U. and Pestova, T.V. (2010) The mechanism of nonsense-mediated mRNA decay, and thereafter. Cold Spring Harbor
eukaryotic translation initiation and principles of its regulation. Nat. Rev. Symp. Quant. Biol. 75, 127–134
Mol. Cell Biol. 11, 113–127 30 Silva, A.L., Ribeiro, P., Inacio, A., Liebhaber, S.A. and Romao, L. (2008)
8 Derry, M.C., Yanagiya, A., Martineau, Y. and Sonenberg, N. (2006) Proximity of the poly(A)-binding protein to a premature termination
Regulation of poly(A)-binding protein through PABP-interacting proteins. codon inhibits mammalian nonsense-mediated mRNA decay. RNA 14,
Cold Spring Harbor Symp. Quant. Biol. 71, 537–543 563–576
9 Kozlov, G., Menade, M., Rosenauer, A., Nguyen, L. and Gehring, K. 31 Singh, G., Rebbapragada, I. and Lykke-Andersen, J. (2008) A competition
(2010) Molecular determinants of PAM2 recognition by the MLLE between stimulators and antagonists of Upf complex recruitment
domain of poly(A)-binding protein. J. Mol. Biol. 397, 397–407 governs human nonsense-mediated mRNA decay. PLoS Biol. 6, e111
10 Siddiqui, N., Mangus, D.A., Chang, T.C., Palermino, J.M., Shyu, A.B. and 32 Ivanov, P.V., Gehring, N.H., Kunz, J.B., Hentze, M.W. and Kulozik, A.E.
Gehring, K. (2007) Poly(A) nuclease interacts with the C-terminal (2008) Interactions between UPF1, eRFs, PABP and the exon junction
domain of polyadenylate-binding protein domain from poly(A)-binding complex suggest an integrated model for mammalian NMD pathways.
protein. J. Biol. Chem. 282, 25067–25075 EMBO J. 27, 736–747
11 Tritschler, F., Huntzinger, E. and Izaurralde, E. (2010) Role of GW182 33 Eberle, A.B., Stalder, L., Mathys, H., Orozco, R.Z. and Muhlemann, O.
proteins and PABPC1 in the miRNA pathway: a sense of déjà vu. Nat. (2008) Posttranscriptional gene regulation by spatial rearrangement of
Rev. Mol. Cell Biol. 11, 379–384 the 3 untranslated region. PLoS Biol. 6, e92
C The Authors Journal compilation
C 2012 Biochemical Society864 Biochemical Society Transactions (2012) Volume 40, part 4
34 Chatterjee, S. and Pal, J.K. (2009) Role of 5 - and 3 -untranslated regions 54 Esteller, M. (2011) Non-coding RNAs in human disease. Nat. Rev. Genet.
of mRNAs in human diseases. Biol. Cell 101, 251–262 12, 861–874
35 Peixeiro, I., Silva, A.L. and Romao, L. (2011) Control of human β-globin 55 van Kouwenhove, M., Kedde, M. and Agami, R. (2011) MicroRNA
mRNA stability and its impact on β-thalassemia phenotype. regulation by RNA-binding proteins and its implications for cancer. Nat.
Haematologica 96, 905–913 Rev. 11, 644–656
36 Voon, H.P. and Vadolas, J. (2008) Controlling α-globin: a review of 56 Huntzinger, E. and Izaurralde, E. (2011) Gene silencing by microRNAs:
α-globin expression and its impact on β-thalassemia. Haematologica contributions of translational repression and mRNA decay. Nat. Rev.
93, 1868–1876 Genet. 12, 99–110
37 Wang, Z. and Kiledjian, M. (2000) The poly(A)-binding protein and an 57 Huntzinger, E., Braun, J.E., Heimstadt, S., Zekri, L. and Izaurralde, E.
mRNA stability protein jointly regulate an endoribonuclease activity. Mol. (2010) Two PABPC1-binding sites in GW182 proteins promote
Cell. Biol. 20, 6334–6341 miRNA-mediated gene silencing. EMBO J. 29, 4146–4160
38 Stefanizzi, I. and Canete-Soler, R. (2007) Coregulation of light 58 Fabian, M.R., Mathonnet, G., Sundermeier, T., Mathys, H., Zipprich, J.T.,
neurofilament mRNA by poly(A)-binding protein and aldolase C: Svitkin, Y.V., Rivas, F., Jinek, M., Wohlschlegel, J., Doudna, J.A. et al.
implications for neurodegeneration. Brain Res. 1139, 15–28 (2009) Mammalian miRNA RISC recruits CAF1 and PABP to affect
39 Canete-Soler, R., Reddy, K.S., Tolan, D.R. and Zhai, J. (2005) Aldolases A PABP-dependent deadenylation. Mol. Cell 35, 868–880
and C are ribonucleolytic components of a neuronal complex that 59 Jinek, M., Fabian, M.R., Coyle, S.M., Sonenberg, N. and Doudna, J.A.
regulates the stability of the light-neurofilament mRNA. J. Neurosci. 25, (2010) Structural insights into the human GW182–PABC interaction in
4353–4364 microRNA-mediated deadenylation. Nat. Struct. Mol. Biol. 17, 238–240
40 Grosset, C., Chen, C.Y., Xu, N., Sonenberg, N., Jacquemin-Sablon, H. and 60 Fabian, M.R., Cieplak, M.K., Frank, F., Morita, M., Green, J., Srikumar, T.,
Shyu, A.B. (2000) A mechanism for translationally coupled mRNA Nagar, B., Yamamoto, T., Raught, B., Duchaine, T.F. et al. (2011)
turnover: interaction between the poly(A) tail and a c-fos RNA coding miRNA-mediated deadenylation is orchestrated by GW182 through two
determinant via a protein complex. Cell 103, 29–40 conserved motifs that interact with CCR4-NOT. Nat. Struct. Mol. Biol. 18,
41 Chang, T.C., Yamashita, A., Chen, C.Y., Yamashita, Y., Zhu, W., Durdan, S., 1211–1217
Kahvejian, A., Sonenberg, N. and Shyu, A.B. (2004) UNR, a new partner 61 Braun, J.E., Huntzinger, E., Fauser, M. and Izaurralde, E. (2011) GW182
of poly(A)-binding protein, plays a key role in translationally coupled proteins directly recruit cytoplasmic deadenylase complexes to miRNA
mRNA turnover mediated by the c-fos major coding-region determinant. targets. Mol. Cell 44, 120–133
Genes Dev. 18, 2010–2023 62 Chekulaeva, M., Mathys, H., Zipprich, J.T., Attig, J., Colic, M., Parker, R. and
42 Clark, A., Dean, J., Tudor, C. and Saklatvala, J. (2009) Post-transcriptional Filipowicz, W. (2011) miRNA repression involves GW182-mediated
gene regulation by MAP kinases via AU-rich elements. Front. Biosci. 14, recruitment of CCR4-NOT through conserved W-containing motifs. Nat.
847–871 Struct. Mol. Biol. 18, 1218–1226
43 Sladic, R.T., Lagnado, C.A., Bagley, C.J. and Goodall, G.J. (2004) Human 63 Walters, R.W., Bradrick, S.S. and Gromeier, M. (2010) Poly(A)-binding
PABP binds AU-rich RNA via RNA-binding domains 3 and 4. Eur. J. protein modulates mRNA susceptibility to cap-dependent
Biochem. 271, 450–457 miRNA-mediated repression. RNA 16, 239–250
44 Hitti, E. and Khabar, K.S. (2012) Sequence variations affecting AU-rich 64 Gorgoni, B., Richardson, W.A., Burgess, H.M., Anderson, R.C., Wilkie, G.S.,
element function and disease. Front. Biosci. 17, 1846–1860 Gautier, P., Martins, J.P., Brook, M., Sheets, M.D. and Gray, N.K. (2011)
45 Khabar, K.S. (2010) Post-transcriptional control during chronic Poly(A)-binding proteins are functionally distinct and have essential roles
inflammation and cancer: a focus on AU-rich elements. Cell. Mol. Life Sci. during vertebrate development. Proc. Natl. Acad. Sci. U.S.A. 108,
67, 2937–2955 7844–7849
46 Nagaoka, K., Suzuki, T., Kawano, T., Imakawa, K. and Sakai, S. (2006) 65 Burgess, H.M. and Gray, N.K. (2012) An integrated model for the
Stability of casein mRNA is ensured by structural interactions between nucleo-cytoplasmic transport of cytoplasmic poly(A)-binding proteins.
the 3 -untranslated region and poly(A) tail via the HuR and Commun. Integr. Biol. 5, 243–247
poly(A)-binding protein complex. Biochim. Biophys. Acta 1759, 66 Voeltz, G.K., Ongkasuwan, J., Standart, N. and Steitz, J.A. (2001) A novel
132–140 embryonic poly(A) binding protein, ePAB, regulates mRNA
47 Marchese, F.P., Aubareda, A., Tudor, C., Saklatvala, J., Clark, A.R. and deadenylation in Xenopus egg extracts. Genes Dev. 15, 774–788
Dean, J.L. (2010) MAPKAP kinase 2 blocks tristetraprolin-directed mRNA 67 Wilkie, G.S., Gautier, P., Lawson, D. and Gray, N.K. (2005) Embryonic
decay by inhibiting CAF1 deadenylase recruitment. J. Biol. Chem. 285, poly(A)-binding protein stimulates translation in germ cells. Mol. Cell.
27590–27600 Biol. 25, 2060–2071
48 Clement, S.L., Scheckel, C., Stoecklin, G. and Lykke-Andersen, J. (2011) 68 Friend, K., Brook, M., Betül Bezirci, F., Sheets, M., Gray, N.K. and Seli, E.
Phosphorylation of tristetraprolin by MK2 impairs AU-rich element mRNA (2012) Embryonic poly(A)-binding protein (ePAB) phosphorylation is
decay by preventing deadenylase recruitment. Mol. Cell. Biol. 31, required for Xenopus oocyte maturation. Biochem. J. 445, 93–100
256–266 69 Kim, J.H. and Richter, J.D. (2007) RINGO/cdk1 and CPEB mediate poly(A)
49 Rowlett, R.M., Chrestensen, C.A., Schroeder, M.J., Harp, M.G., Pelo, J.W., tail stabilization and translational regulation by ePAB. Genes Dev. 21,
Shabanowitz, J., Derose, R., Hunt, D.F., Sturgill, T.W. and Worthington, 2571–2579
M.T. (2008) Inhibition of tristetraprolin (TTP) deadenylation by poly(A) 70 Cosson, B., Berkova, N., Couturier, A., Chabelskaya, S., Philippe, M. and
binding protein. Am. J. Physiol. Gastrointest. Liver Physiol. 295, Zhouravleva, G. (2002) Poly(A)-binding protein and eRF3 are associated
G421–G430 in vivo in human and Xenopus cells. Biol. Cell 94, 205–216
50 Lu, J.Y., Sadri, N. and Schneider, R.J. (2006) Endotoxic shock in AUF1 71 Guzelogylu-Kayisli, O., Lalioti, M.D., Aydiner, F., Sasson, I., Ilbay, O.,
knockout mice mediated by failure to degrade proinflammatory cytokine Sakkas, D., Lowther, K.M., Mehlmann, L.M. and Seli, M. (2012)
mRNAs. Genes Dev. 20, 3174–3184 Embryonic poly(A)-binding protein (EPAB) is required for oocyte
51 Laroia, G., Cuesta, R., Brewer, G. and Schneider, R.J. (1999) Control of maturation and female fertility in mice. Biochem. J. 446, 47–58
mRNA decay by heat shock–ubiquitin–proteasome pathway. Science 72 Brook, M., McCracken, L., Reddington, J.P., Lu, Z.L., Morrice, N.A. and
284, 499–502 Gray, N.K. (2012) The multifunctional poly(A)-binding protein (PABP) 1 is
52 Lu, J.Y., Bergman, N., Sadri, N. and Schneider, R.J. (2006) Assembly of subject to extensive dynamic post-translational modification, which
AUF1 with eIF4G-poly(A) binding protein complex suggests a translation molecular modelling suggests plays an important role in co-ordinating its
function in AU-rich mRNA decay. RNA 12, 883–893 activities. Biochem. J. 441, 803–812
53 Sagliocco, F., Laloo, B., Cosson, B., Laborde, L., Castroviejo, M.,
Rosenbaum, J., Ripoche, J. and Grosset, C. (2006) The ARE-associated
factor AUF1 binds poly(A) in vitro in competition with PABP. Biochem. J. Received 16 April 2012
400, 337–347 doi:10.1042/BST20120100
C The Authors Journal compilation
C 2012 Biochemical SocietyYou can also read

















































